
Biocontrol of Macrophomina phaseolina Using Bacillus amyloliquefaciens Strains in Cowpea (Vigna unguiculata L.)



 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Characterization of Microorganisms
2.2. DNA Extraction and Phylogenetic Analysis
2.3. Characterization of Growth Conditions for Bacterial Isolates
2.4. Evaluation of Plant Growth-Promoting Activities
2.5. Production of Cell Wall-Degrading Enzymes
2.6. In Vitro Fungal Antagonism
2.7. Fungal Growth Inhibition by Bacterial Volatile Organic and Diffusible Compounds
2.8. Effect of Bacteria Isolates on the Germination of M. phaseolina Microsclerotia
2.9. Biocontrol Assay in Planta
2.10. Statistical Analysis
3. Results and Discussion
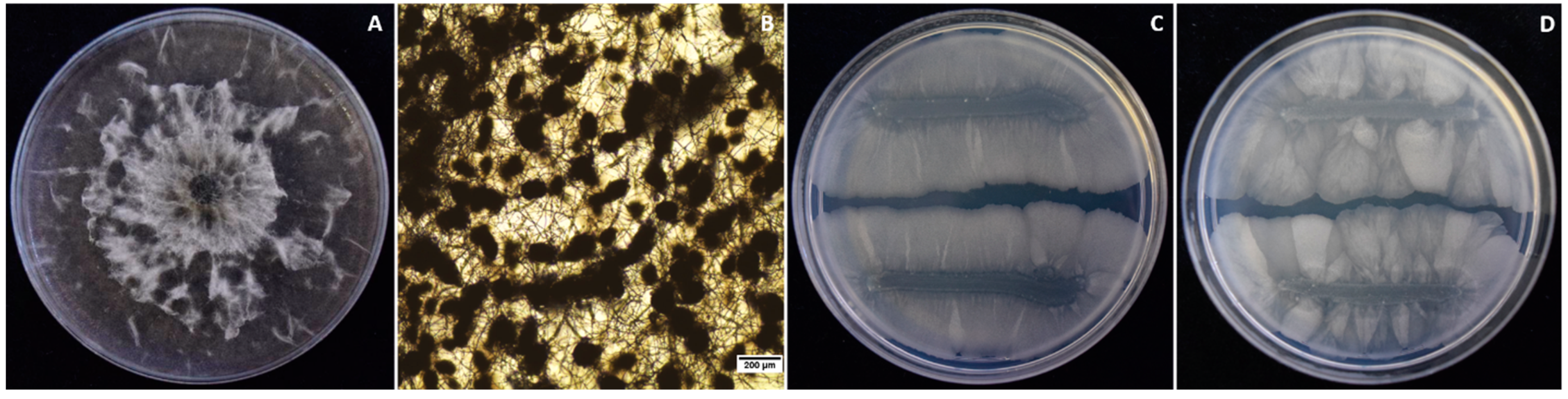
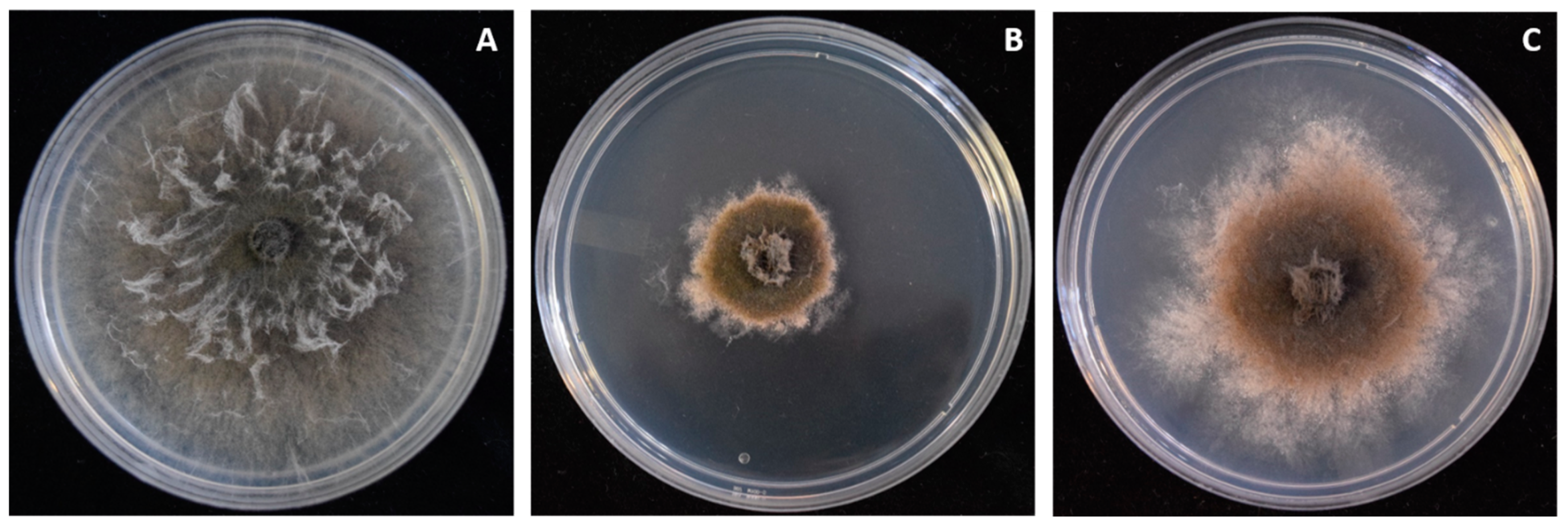
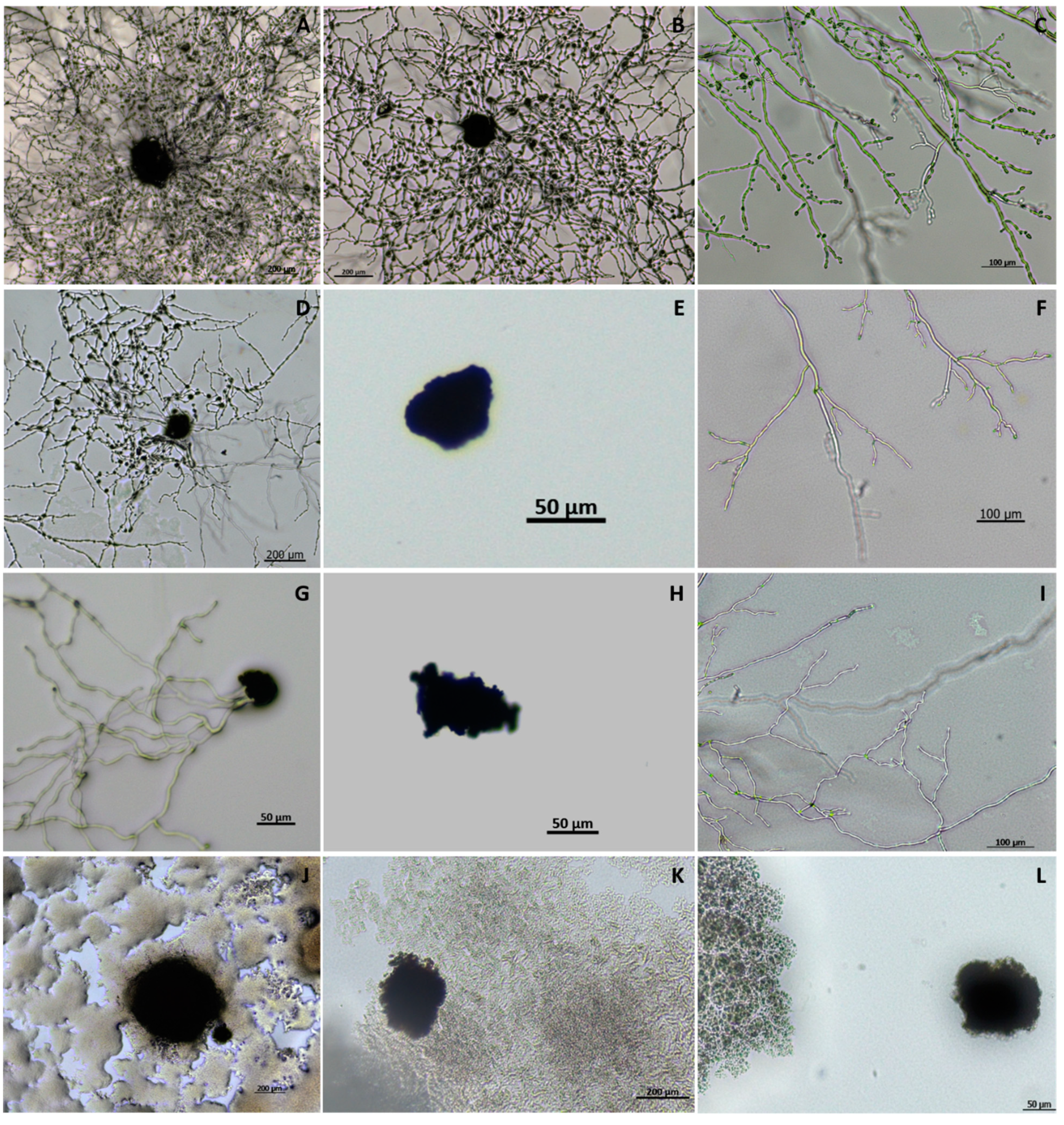
3.1. Isolation and Identification of Macrophomina phaseolina from Strawberry Crown Rot
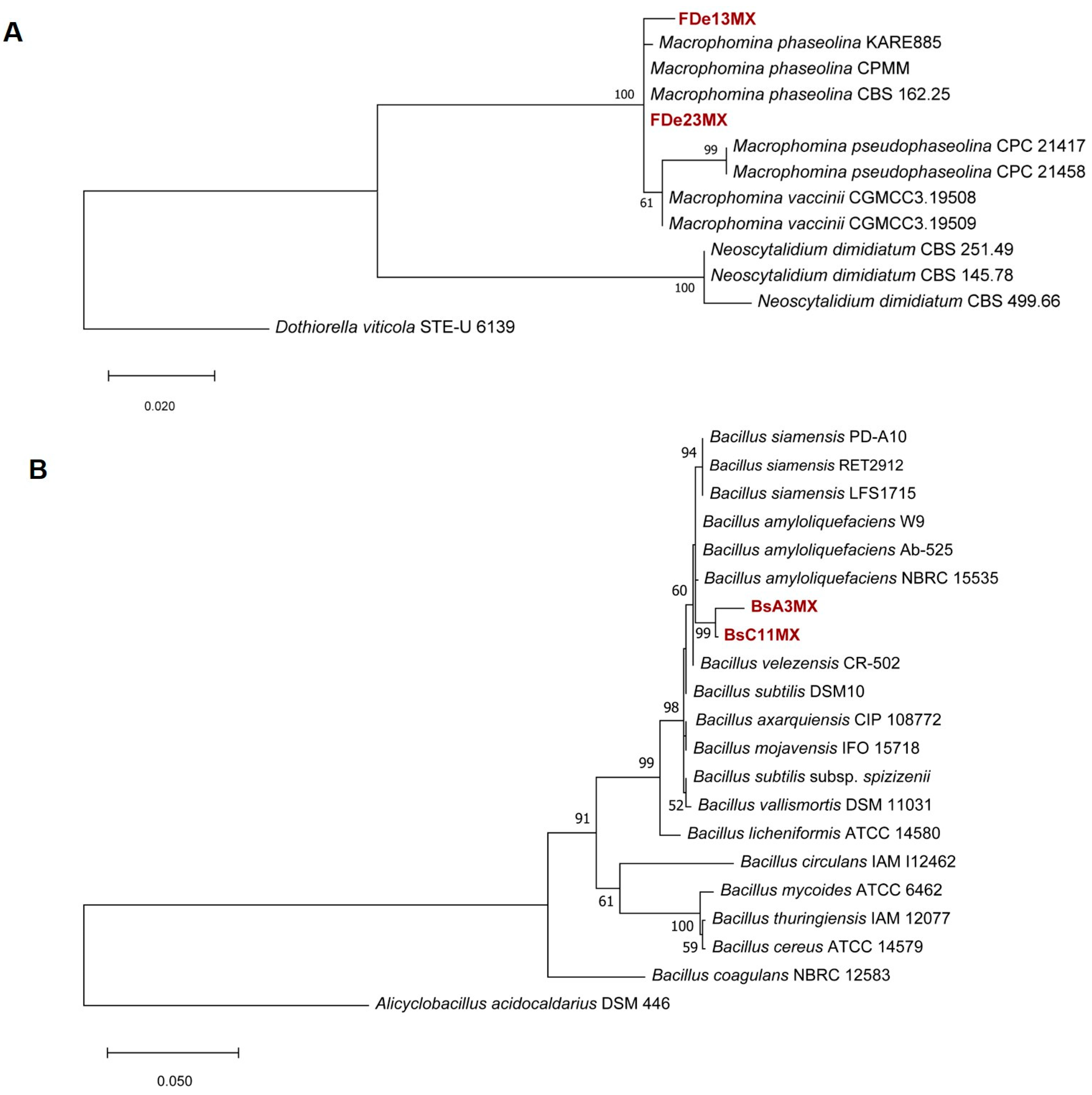
3.2. Bacterial Isolate Identification
3.3. Characterization of Bacillus amyloliquefaciens Isolates
3.4. Characterization of B. amyloliquefaciens as Plant Growth Promoters
3.5. Production of Hydrolytic Enzymes
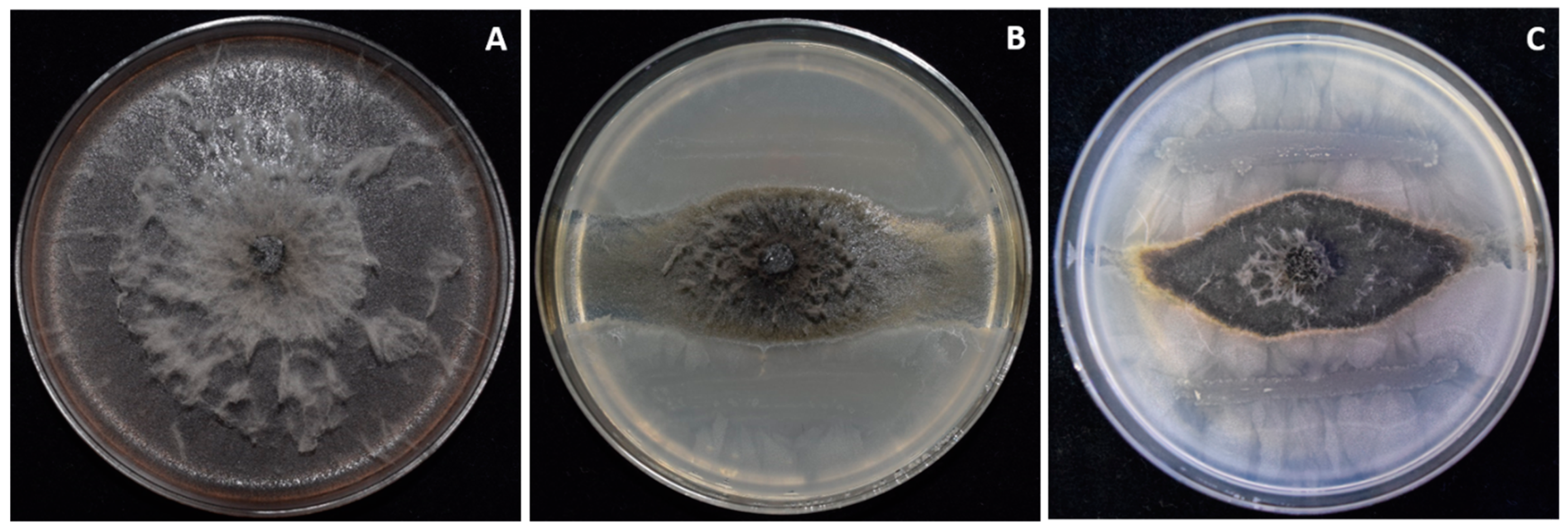
3.6. Two Bacillus amyloliquefaciens Isolates Inhibit Fungal Growth of Macrophomina In Vitro
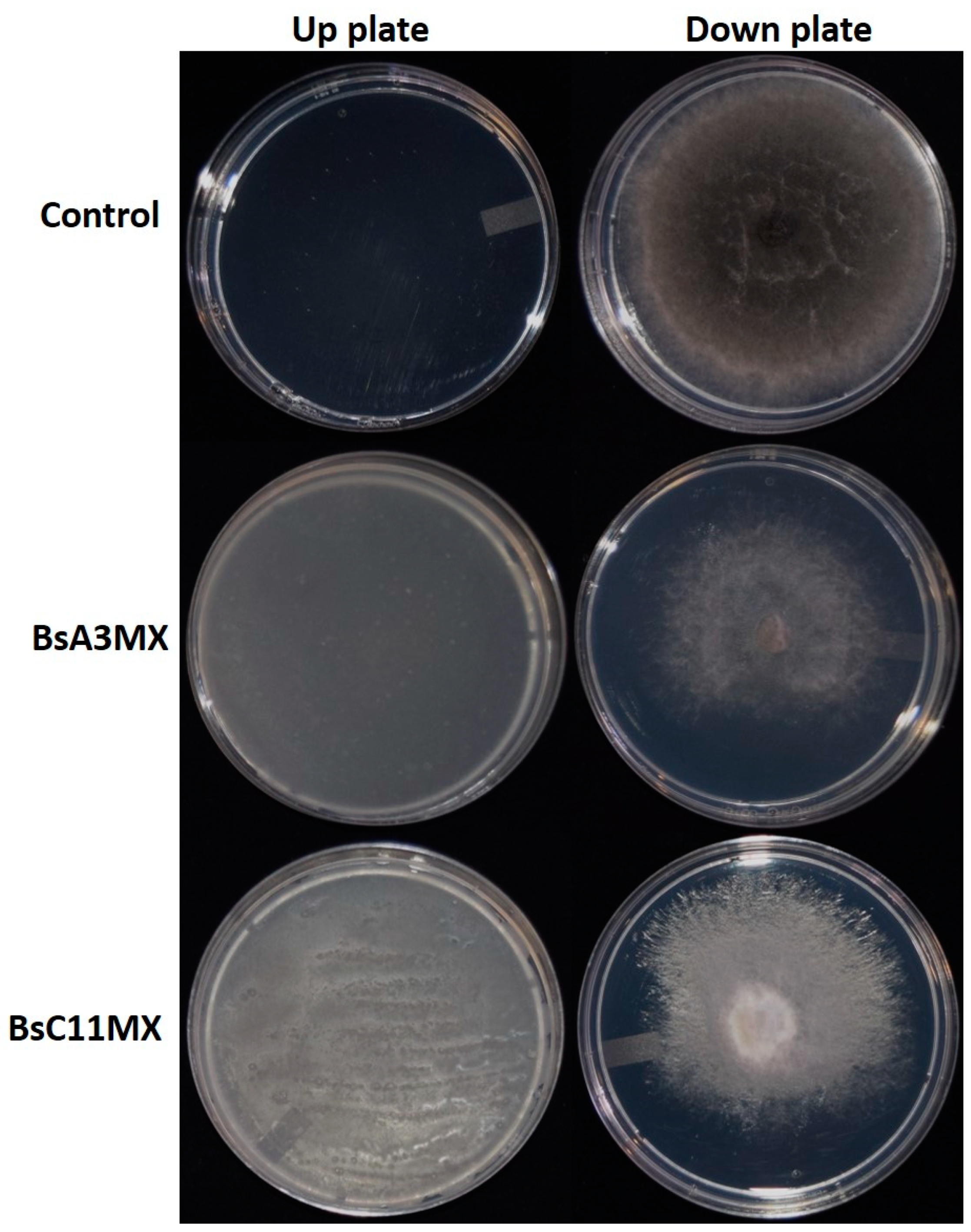
3.7. B. amyloliquefaciens BsA3MX and BsC11MX Inhibit M. phaseolina by the Combined Action of Volatile Organic Compounds and Diffusible Compounds
3.8. Effect of B. amyloliquefaciens on the Germination of M. phaseolina Microsclerotia
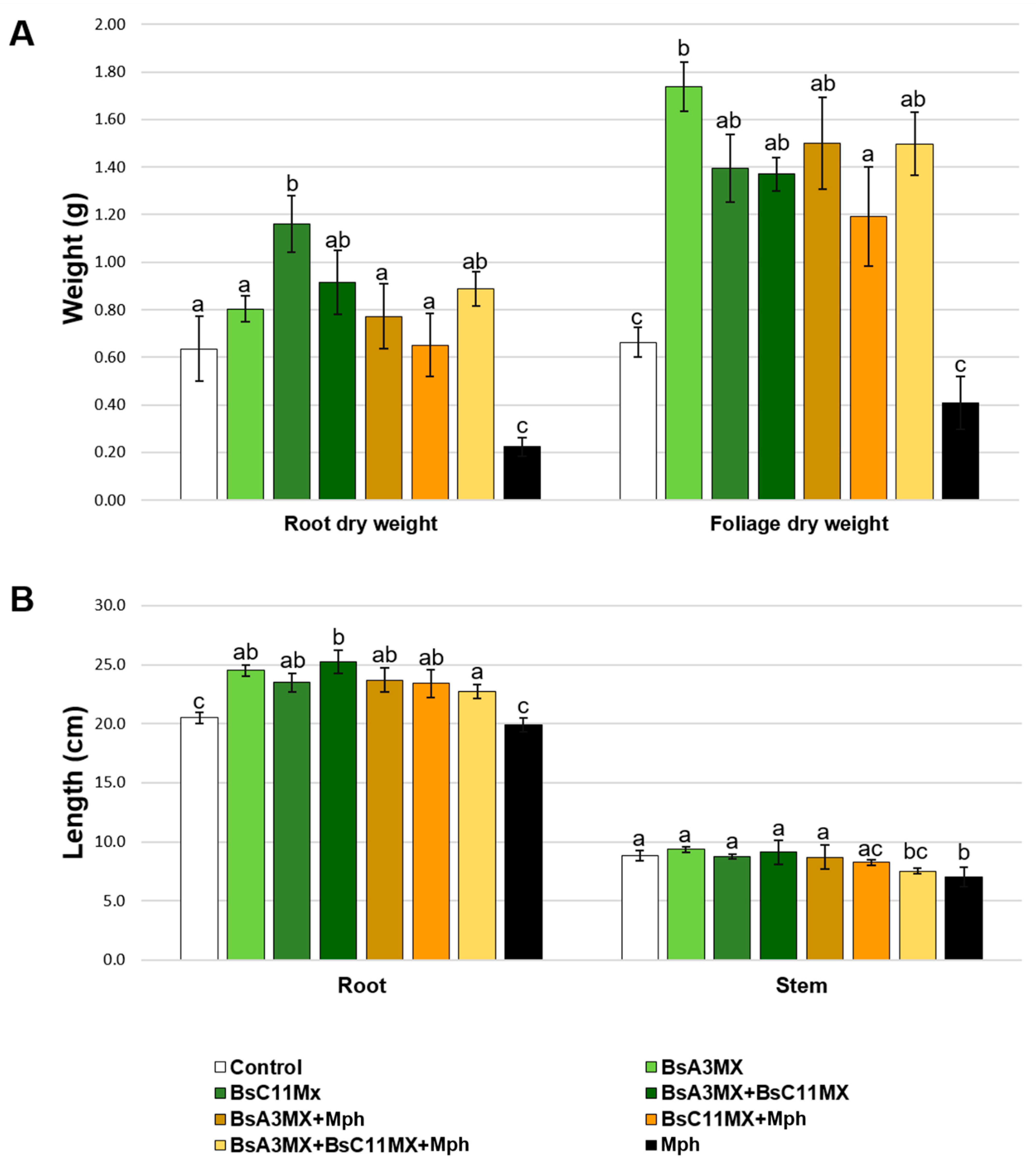
3.9. Bacillus amyloliquefaciens Isolates Promote Plant Growth and Suppress M. phaseolina in Planta
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhingra, O.D.; Sinclair, J.B. Biology and Pathology of Macrophomina phaseolina; Univeridade Feederal de Vicosa: Viçosa, Brazil, 1978; p. 166. [Google Scholar]
- Marquez, N.; Giachero, M.L.; Declerck, S.; Ducasse, D.A. Macrophomina phaseolina: General Characteristics of Pathogenicity and Methods of Control. Front. Plant Sci. 2021, 12, 634397. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Suh, S.-O.; Schneider, R.W.; Russin, J.S. Host Specialization in the Charcoal Rot Fungus, Macrophomina phaseolina. Phytopathology 2001, 91, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.K.; Sharma, S.K.; Ramteke, R. Biology, Epidemiology and Management of the Pathogenic Fungus Macrophomina phaseolina (Tassi) Goid with Special Reference to Charcoal Rot of Soybean (Glycine max (L.) Merrill). J. Phytopathol. 2012, 160, 167–180. [Google Scholar] [CrossRef]
- Kunwar, I.K.; Singh, T.; Machado, C.C.; Sinclair, J.B. Histopathology of soybean seed and seedling infection by Macrophomina phaseolina. Phytopathology 1986, 76, 532–535. [Google Scholar] [CrossRef]
- Zveibil, A.; Freeman, S. First Report of Crown and Root Rot in Strawberry Caused by Macrophomina phaseolina in Israel. Plant Dis. 2005, 89, 1014. [Google Scholar] [CrossRef]
- Islam, S.; Haque, S.; Islam, M.M.; Emdad, E.M.; Halim, A.; Hossen, Q.M.M.; Hossain, Z.; Ahmed, B.; Rahim, S.; Rahman, S.; et al. Tools to kill: Genome of one of the most destructive plant pathogenic fungi Macrophomina phaseolina. BMC Genom. 2012, 13, 493. [Google Scholar] [CrossRef]
- Ijaz, S.; Sadaqat, H.A.; Khan, M.N. A review of the impact of charcoal rot (Macrophomina phaseolina) on sunflower. J. Agric. Sci. 2013, 151, 222–227. [Google Scholar] [CrossRef]
- Salahlou, R.; Safaie, N.; Shams-Bakhsh, M. Genetic diversity of Macrophomina phaseolina populations, the causal agent of sesame charcoal rot using inter-simple sequence repeat markers. J. Agric. Sci. 2016, 18, 277–287. [Google Scholar]
- Vibha. Macrophomina phaseolina: The Most Destructive Soybean Fungal Pathogen of Global Concern. In Current Trends in Plant Disease Diagnostics and Management Practices; Kumar, P., Gupta, V., Tiwari, A., Kamle, M., Eds.; Fungal Biology Series; Springer: Cham, Switzerland, 2016; pp. 193–205. [Google Scholar]
- Meyer, W.A.; Sinclair, J.B.; Khare, M.N. Factors affecting charcoal rot of soybean seedlings. Phytopathology 1974, 64, 845–849. [Google Scholar] [CrossRef]
- Pandey, A.K.; Basandrai, A.K. Will Macrophomina phaseolina spread in legumes due to climate change? A critical review of current knowledge. J. Plant Dis. Prot. 2021, 128, 9–18. [Google Scholar] [CrossRef]
- Kanaan, H.; Medina, S.; Krassnovsky, A.; Raviv, M. Survival of Macrophomina phaseolina s.l. and Verticillium dahliae during solarization as affected by composts of various maturities. Crop Prot. 2015, 76, 108–113. [Google Scholar] [CrossRef]
- Lokesh, R.; Rakholiya, K.B.; Thesiya, M.R. Evaluation of Different Fungicides against Macrophomina phaseolina (Tassi) Goid. Causing Dry Root Rot of Chickpea (Cicer arietinum L.) invitro. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 901–911. [Google Scholar] [CrossRef]
- Almeida, Á.M.; Sosa-Gomez, D.R.; Binneck, E.; Marin, S.R.; Zucchi, M.I.; Abdelnoor, R.V.; Souto, E.R. Effect of crop rotation on specialization and genetic diversity of Macrophomina phaseolina. Trop. Plant Pathol. 2008, 33, 257–264. [Google Scholar] [CrossRef]
- Adrees, H.; Haider, M.S.; Anjum, T.; Akram, W. Inducing systemic resistance in cotton plants against charcoal root rot pathogen using indigenous rhizospheric bacterial strains and chemical elicitors. Crop Prot. 2019, 115, 75–83. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Decrease in the incidence of charcoal root rot in common bean (Phaseolus vulgaris L.) by Bacillus amyloliquefaciens B14, a strain with PGPR properties. Biol. Control 2017, 113, 1–8. [Google Scholar] [CrossRef]
- Villarreal-Delgado, M.F.; Villa-Rodríguez, E.D.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Parra-Cota, F.I.; de los Santos-Villalobos, S. The genus Bacillus as a biological control agent and its implications in the agricultural biosecurity. Mex. J. Phytopathol. 2018, 36, 95–130. [Google Scholar] [CrossRef]
- Maughan, H.; Van der Auwera, G. Bacillus taxonomy in the genomic era finds phenotypes to be essential though often misleading. Infect. Genet. Evol. 2011, 11, 789–797. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Khan, M.; Salman, M.; Jan, S.A.; Shinwari, Z.K. Biological control of fungal phytopathogens: A comprehensive review based on Bacillus species. MOJ Biol. Med. 2021, 6, 90–92. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Martínez-Salgado, S.J.; Andrade-Hoyos, P.; Parraguirre Lezama, C.; Rivera-Tapia, A.; Luna-Cruz, A.; Romero-Arenas, O. Biological control of charcoal rot in peanut crop through strains of Trichoderma spp., in Puebla, Mexico. Plants 2021, 10, 2630. [Google Scholar] [CrossRef] [PubMed]
- Khambhati, V.H.; Abbas, H.K.; Sulyok, M.; Tomaso-Peterson, M.; Shier, W.T. First report of the production of mycotoxins and other secondary metabolites by Macrophomina phaseolina (Tassi) Goid. Isolates from soybeans (Glycine max L.) symptomatic with charcoal rot disease. J. Fungi 2020, 6, 332. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.P.R.; Mueller, D.; Mengistu, A.; Singh, A.K.; Hartman, G.L.; Wise, K.A. Advancing Our Understanding of Charcoal Rot in Soybeans. J. Integr. Pest Manag. 2017, 8, 8. [Google Scholar] [CrossRef]
- Reyes-Franco, M.C.; Hernández-Delgado, S.; Beas-Fernández, R.; Medina-Fernández, M.; Simpson, J.; Mayek-Pérez, N. Pathogenic and genetic variability within Macrophomina phaseolina from Mexico and other countries. J. Phytopathol. 2006, 154, 447–453. [Google Scholar] [CrossRef]
- Leyva-Mir, S.G.; Velázquez-Martínez, G.C.; Tlapal-Bolaños, B.; Tovar-Pedraza, J.M.; Rosas-Saito, G.H.; Alvarado-Gómez, O.G. Caracterización morfológica y molecular de aislados de Macrophomina phaseolina asociados a caña de azúcar en México. Rev. Argent. Microbiol. 2015, 47, 143–147. [Google Scholar] [CrossRef][Green Version]
- Ayala-Armenta, Q.A.; Tovar-Pedraza, J.M.; Apodaca-Sánchez, M.A.; Correia, K.C.; Sauceda-Acosta, C.P.; Camacho-Tapia, M.; Beltrán-Peña, H. Phylogeny and pathogenicity of soilborne fungi associated with wilt disease complex of tomatillo (Physalis ixocarpa) in northern Sinaloa, Mexico. Eur. J. Plant Pathol. 2020, 157, 733–749. [Google Scholar] [CrossRef]
- Martínez-Salgado, S.J.; Romero-Arenas, O.; Morales-Mora, L.A.; Luna-Cruz, A.; Rivera-Tapia, J.A.; Silva-Rojas, H.V.V.; Andrade-Hoyos, P. First Report of Macrophomina phaseolina Causing Charcoal Rot of Peanut (Arachis hypogaea) in Mexico. Plant Dis. 2021, 105, 2722. [Google Scholar] [CrossRef]
- SIAP. Servicio de Información y Estadística Agroalimentaria y Pesquera; Ministerio de Agricultura de Mexico, Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA): Mexico City, Mexico, 2020. [Google Scholar]
- Williams-Alanís, H.; Pecina-Quintero, V.; Montes-García, N.; Zavala-García, F.; Arcos-Cavazos, G.; Gámez-Vázquez, A.J. Evaluation of sorghum [Sorghum bicolor (L.) Moench.] hybrids for resistance to charcoal rot [Macrophomina phaseolina (Tassi) Goid.] in Tamaulipas, Mexico. Revista Mex. Fitopatol. 2009, 27, 69–72. [Google Scholar]
- Mayek-Pérez, N.; López-Salinas, E.; Cumpián-Gutiérrez, J.; Acosta-Gallegos, J.A. Inheritance of field resistance to Macrophomina phaseolina (Tassi) Goid. in recombinant endogamic lines of common bean (Phaseolus vulgaris L.). Revista Mex. Fitopatol. 2009, 27, 1–10. [Google Scholar]
- Pickel, B.; Dai, N.; Maymon, M.; Elazar, M.; Tanami, Z.; Frenkel, O.; Toamy, M.A.; Mor, N.; Freeman, S. Development of a reliable screening technique for determining tolerance to Macrophomina phaseolina in strawberry. Eur. J. Plant Pathol. 2020, 157, 707–718. [Google Scholar] [CrossRef]
- Lee, S.B.; Milgroom, M.G.; Taylor, J.W. A rapid, high yield mini-prep method for isolation of total genomic DNA from fungi. Fungal Genet. Rep. 1988, 35, 11. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Cowan, S.T. Cowan and Steel’s Manual for the Identification of Medical Bacteria; Cambridge University Press: Cambridge, UK, 2004; p. 352. [Google Scholar]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. [6] Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Badgujar, M.D.; Sonawane, H.M.; Mhaske, M.M.; Chincholkar, S.B. Production of microbial iron chelators (siderophores) by fluorescent Pseudomonas. Indian J. Biotechnol. 2005, 4, 484–490. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizosferic bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric Estimation of Indoleacetic Acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J.W. Experiments with Some Microorganisms which Utilize Ethane and Hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef]
- Govindasamy, V.; Senthilkumar, M.; Mageshwaran, V.; Annapurna, K. Detection and Characterization of ACC Deaminase in Plant Growth Promoting Rhizobacteria. J. Plant Biochem. Biotechnol. 2009, 18, 71–76. [Google Scholar] [CrossRef]
- Baldani, J.I.; Reis, V.M.; Videira, S.S.; Boddey, L.H.; Baldani, V.L.D. The art of isolating nitrogen-fixing bacteria from non-leguminous plants using N-free semi-solid media: A practical guide for microbiologists. Plant Soil 2014, 384, 413–431. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Corrales Ramírez, L.C.; Sánchez Leal, L.C.; Arevalo Galvez, Z.Y.; Moreno Burbano, V.E. Bacillus: Género bacteriano que demuestra ser un importante solubilizador de fosfato. Nova 2014, 12, 165–178. [Google Scholar] [CrossRef]
- Franco-Correa, M. Evaluación de Caracteres PGPR en Actinomicetos e Interacciones de Estas Rizobacterias con Hongos Formadores de Micorriza. Ph.D. Thesis, Facultad de Ciencias, Universidad de Granada,, Granada, Spain, 2008. [Google Scholar]
- Komal, R.; Kalavati, P. Isolation and screening of zinc mobilizing microorganisms from rhizospheric soil. Paripex Indian J. Res. 2018, 7, 58–60. [Google Scholar] [CrossRef]
- Tagele, S.B.; Kim, S.W.; Lee, H.G.; Lee, Y.S. Potential of novel sequence type of Burkoldera cenocepacia for biological control of root rot maize (Zea mays L.) cause by Fusarium temperatum. Int. J. Mol. Sci. 2019, 20, 1005. [Google Scholar] [CrossRef]
- Agrawal, T.; Kotashane, S.A. Chitinolytic assay of indigenous Thrichoderma isolates collected from diferente geographical locations of Chhattisgarh in Central India. SpringerPlus 2012, 1, 1–10. [Google Scholar] [CrossRef]
- Mazzucotelli, C.A.; Ponce, A.G.; Kotlar, C.E.; Moreira, M.D.R. Isolation and characterization of bacterial strains with a hydrolytic profile with potential use in bioconversion of agroindustial by-products and waste. Food Sci. Technol. 2013, 33, 295–303. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, R. Production of alkaline pectinase by bacteria (Cocci sps.) isolated from decomposing fruit materials. J. Phytol. 2012, 4, 1–5. [Google Scholar]
- Hankin, L.; Anagnostakis, S.L. The use of solid media for detection of enzyme production by fungi. Mycologia 1975, 67, 597–607. [Google Scholar] [CrossRef]
- Fokkema, N.J. Antagonism between fungal saprophytes and pathogens on aerial plant surfaces. In Microbiology of Aerial Plant Surfaces; Dickinson, C.H., Preece, T.F., Eds.; Academic Press: London, UK, 1976; pp. 487–505. [Google Scholar]
- Ammon, V.; Wyllie, T.D.; Brown, M.F. Investigation of the infection process of Macrophomina phaseolina on the surface of soybean roots using scanning electron microscopy. Mycopathologia 1975, 55, 77–81. [Google Scholar] [CrossRef]
- Aviles, M.; Castillo, S.; Bascon, J.; Zea-Bonilla, T.; Martin-Sanchez, P.M.; Pérez-Jiménez, R.M. First report of Macrophomina phaseolina causing crown and root rot of strawberry in Spain. Plant Pathol. 2008, 57, 382. [Google Scholar] [CrossRef]
- Baino, O.M.; Salazar, S.M.; Ramallo, A.C.; Kirschbaum, D.S. First Report of Macrophomina phaseolina Causing Strawberry Crown and Root Rot in Northwestern Argentina. Plant Dis. 2011, 95, 1477. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, S.; Gambardella, M.; Henríquez, J.L.; Diaz, I. First Report of Crown Rot of Strawberry Caused by Macrophomina phaseolina in Chile. Plant Dis. 2013, 97, 996. [Google Scholar] [CrossRef]
- Hutton, D.G.; Gomez, A.O.; Mattner, S.W. Macrophomina phaseolina and its association with strawberry crown rot in Australia. Int. J. Fruit Sci. 2013, 13, 149–155. [Google Scholar] [CrossRef]
- Koike, S.T. Crown Rot of Strawberry Caused by Macrophomina phaseolina in California. Plant Dis. 2008, 92, 1253. [Google Scholar] [CrossRef]
- Koike, S.T.; Arias, R.S.; Hogan, C.S.; Martin, F.N.; Gordon, T.R. Status of Macrophomina phaseolina on strawberry in California and preliminary characterization of the pathogen. Int. J. Fruit Sci. 2016, 16 (Suppl. 1), 148–159. [Google Scholar] [CrossRef]
- Priest, F.G.; Goodfellow, M.; Shute, L.A.; Berkeley, R.C.W. Bacillus amyloliquefaciens sp. nov., nom. rev. Int. J. Syst. Evol. Microbiol. 1987, 37, 69–71. [Google Scholar] [CrossRef]
- Fan, B.; Blom, J.; Klenk, H.P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis form an “operational group B. amyloliquefaciens” within the B. subtilis species complex. Front. Microbiol. 2017, 8, 22. [Google Scholar] [CrossRef]
- Chavera, G.F.S.; Macuer, M.A.; Torres, P.M. Endospore-Forming Bacteria Present in a Commercial Stabilized Poultry Manure Determines the Fusarium Biocontrol and the Tomato Growth Promotion. Agronomy 2020, 10, 1636. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-Producing Rhizobacteria as a Promising Tool for Empowering Plants to Cope with Iron Limitation in Saline Soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, N.; Maheshwari, R.; Negi, M.; Suneja, P. Optimization of IAA production by endophytic Bacillus spp. from Vigna radiate for their potential use as plant growth promoters. Isr. J. Plant Sci. 2018, 65, 83–96. [Google Scholar] [CrossRef]
- Yasmin, H.; Naz, R.; Nosheen, A.; Hassan, M.; Ilyas, N.; Sajjad, M.; Anjum, S.; Gao, X.; Geng, Z. Identification of New Biocontrol Agent against Charcoal Rot Disease Caused by Macrophomina phaseolina in Soybean (Glycine Max L.). Sustainability 2020, 12, 6856. [Google Scholar] [CrossRef]
- Singh, N.; Pandey, P.; Dubey, R.; Maheshwari, D.K. Biological control of root rot fungus Macrophomina phaseolina and growth enhancement of Pinus roxburghii (Sarg.) by rhizosphere competent Bacillus subtilis BN1. World J. Microbiol. Biotechnol. 2008, 24, 1669–1679. [Google Scholar] [CrossRef]
- Afouda, L.; Schulz, D.; Wolf, G.; Wydra, K. Biological control of Macrophomina phaseolina on cowpea (Vigna unguiculata) under dry conditions by bacterial antagonists. Int. J. Biol. Chem. Sci. 2012, 6, 5068–5077. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Radhakrishnan, R.; Kumar, A. Plant defense approach of Bacillus subtilis (BERA 71) against Macrophomina phaseolina (Tassi) Goid in mung bean. J. Plant Interact. 2017, 12, 390–401. [Google Scholar] [CrossRef]
- Bojórquez-Armenta, Y.d.J.; Mora-Romero, G.A.; López-Meyer, M.; Maldonado-Mendoza, I.E.; Castro-Martínez, C.; Romero-Urías, C.D.L.A.; Cordero-Ramírez, J.D.; Martínez-Álvarez, J.C. Evaluation of Bacillus spp. isolates as potential biocontrol agents against charcoal rot caused by Macrophomina phaseolina on common bean. J. Gen. Plant Pathol. 2021, 87, 377–386. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Humayun, P.; Kiran, B.K.; Kannan, I.G.K.; Vidya, M.S.; Deepthi, K.; Rupela, O. Evaluation of bacteria isolated from rice rhizosphere for biological control of charcoal rot of sorghum caused by Macrophomina phaseolina (Tassi) Goid. World J. Microbiol. Biotechnol. 2011, 27, 1313–1321. [Google Scholar] [CrossRef]
- Torres, M.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M. Antagonistic effects of Bacillus subtilis subsp. subtilis and B. amyloliquefaciens against Macrophomina phaseolina: SEM study of fungal changes and UV-MALDI-TOF MS analysis of their bioactive compounds. Microbiol. Res. 2016, 182, 31–39. [Google Scholar] [CrossRef]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin-and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soil-borne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Ali, N.Y.; Suleman, P. Biological control of Sclerotinia sclerotiorum (Lib.) de Bary with Trichoderma harzianum and Bacillus amyloliquefaciens. Crop Prot. 2008, 27, 1354–1359. [Google Scholar] [CrossRef]
- Ji, S.H.; Paul, N.C.; Deng, J.X.; Kim, Y.S.; Yun, B.-S.; Yu, S.H. Biocontrol Activity of Bacillus amyloliquefaciens CNU114001 against Fungal Plant Diseases. Mycobiology 2013, 41, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciensPPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 108, 386–395. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Maharachchikumbura, S.S.; Al-Sadi, A.M.; Kang, S.M.; Yun, B.W.; Lee, I.J. Biocontrol potential of Bacillus amyloliquefaciens against Botrytis pelargonii and Alternaria alternata on Capsicum annuum. J. Fungi 2021, 7, 472. [Google Scholar] [CrossRef]
- Yuan, J.; Raza, W.; Shen, Q.; Huang, Q. Antifungal Activity of Bacillus amyloliquefaciens NJN-6 Volatile Compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. MicrobiologyOpen 2019, 8, e00813. [Google Scholar] [CrossRef]
- Raza, W.; Ling, N.; Yang, L.; Huang, Q.; Shen, Q. Response of tomato wilt pathogen Ralstonia solanacearum to the volatile organic compounds produced by a biocontrol strain Bacillus amyloliquefaciens SQR-9. Sci. Rep. 2016, 6, 24856. [Google Scholar] [CrossRef]
- Asari, S.; Matzén, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple effects of Bacillus amyloliquefaciens volatile compounds: Plant growth promotion and growth inhibition of phytopathogens. FEMS Microbiol. Ecol. 2016, 92, fiw070. [Google Scholar] [CrossRef]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Tahir, H.A.S.; Gu, Q.; Wu, H.; Gao, X. Volatile Compounds of Endophytic Bacillus spp. have Biocontrol Activity Against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef]
- Gotor-Vila, A.; Teixidó, N.; Di Francesco, A.; Usall, J.; Ugolini, L.; Torres, R.; Mari, M. Antifungal effect of volatile organic compounds produced by Bacillus amyloliquefaciens CPA-8 against fruit pathogen decays of cherry. Food Microbiol. 2017, 64, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb. Cell Fact. 2009, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, P.; Zafari, D.; Mahmoodi, S.B.; Hashemi, M.; Gholamhoseini, M.; Dolatabadian, A.; Ataei, R. Histopathology of charcoal rot disease (Macrophomina phaseolina) in resistant and susceptible cultivars of soybean. Rhizosphere 2018, 7, 27–34. [Google Scholar] [CrossRef]
- Lodha, S.; Mawar, R. Population dynamics of Macrophomina phaseolina in relation to disease management: A review. J. Phytopathol. 2020, 168, 1–17. [Google Scholar] [CrossRef]
- Francis, I.; Holsters, M.; Vereecke, D. The Gram-positive side of plant–microbe interactions. Environ. Microbiol. 2010, 12, 1–12. [Google Scholar] [CrossRef]
- Wang, D.-C.; Jiang, C.-H.; Zhang, L.-N.; Chen, L.; Zhang, X.-Y.; Guo, J.-H. Biofilms Positively Contribute to Bacillus amyloliquefaciens 54-induced Drought Tolerance in Tomato Plants. Int. J. Mol. Sci. 2019, 20, 6271. [Google Scholar] [CrossRef]
- Martínez-Austria, P.F.; Jano-Pérez, J.A. Climate Change and Extreme Temperature Trends in the Baja California Peninsula, Mexico. Air Soil Water Res. 2021, 14, 11786221211010702. [Google Scholar] [CrossRef]
- Sezen, A.; Ozdal, M.; Koc, K.; Algur, O.F. Isolation and characterization of plant growth promoting rhizobacteria (PGPR) and their effects on improving growth of wheat. J. Appl. Biol. Sci. 2019, 10, 41–46. [Google Scholar] [CrossRef]
- Delgado-Ramírez, C.S.; Hernández-Martínez, R.; Sepúlveda, E. Rhizobacteria associated with a native solanaceae promote plant growth and decrease the effects of Fusarium oxysporum in tomato. Agronomy 2021, 11, 579. [Google Scholar] [CrossRef]
- Eke, P.; Kumar, A.; Sahu, K.P.; Wakam, L.N.; Sheoran, N.; Ashajyothi, M.; Patel, A.; Fekam, F.B. Endophytic bacteria of desert cactus (Euphorbia trigonas Mill) confer drought tolerance and induce growth promotion in tomato (Solanum lycopersicum L.). Microbiol. Res. 2019, 228, 126302. [Google Scholar] [CrossRef] [PubMed]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef] [PubMed]
- Zafar-Ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress. Agronomy 2019, 9, 343. [Google Scholar] [CrossRef]
- Anderson, R.; Bayer, P.E.; Edwards, D. Climate change and the need for agricultural adaptation. Curr. Opin. Plant Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | BsA3MX | BsC11MX |
|---|---|---|
| Gram stain | + | + |
| Colony morphology | Irregular, flat, dry, and dull colonies creamy-white on TSA | |
| Form | Single rod-shaped | Single rod-shaped |
| Motility | + | + |
| Endospore formation | Two-terminal | Two-terminal |
| Biofilm formation | + | + |
| NaCl | ||
| Maximum | 10% | 10% |
| Temperature | ||
| Maximum | 50 °C | 50 °C |
| pH | ||
| Minimum | 5 | 5 |
| Maximum | 9 | 9 |
| Hemolytic activity | - | - |
| Urease | - | - |
| Oxidase | + | + |
| Catalase | + | + |
| Protease | + | + |
| Citrate utilization | - | - |
| Indole test | + | + |
| Acid from: | ||
| Glucose | +++ | +++ |
| Lactose | + | ++ |
| Arabinose | + | +++ |
| Trehalose | +++ | + |
| Dextrose | +++ | +++ |
| Amylose | + | + |
| Xylose | ++ | ++ |
| Mannitol | ++ | +++ |
| Glycogen | +++ | +++ |
| Methyl-α-D-glucoside | - | - |
| Characteristic | BsA3MX | BsC11MX |
|---|---|---|
| Siderophore production | 69.3% | 54.7% |
| IAA production | 7.2 mg·mL−1 | 6.2 mg·mL−1 |
| ACC-deaminase production | + | + |
| Nitrogen fixation | - | - |
| Cyanhydric acid production | - | - |
| P solubilization | + | + |
| K solubilization | - | - |
| Zn solubilization | + | + |
| Biofilm formation | + | + |
| Chitinase | - | - |
| Cellulase | - | - |
| Pectinase | - | - |
| Amylase | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel-Montoya, E.A.; Delgado-Ramírez, C.S.; Sepulveda, E.; Hernández-Martínez, R. Biocontrol of Macrophomina phaseolina Using Bacillus amyloliquefaciens Strains in Cowpea (Vigna unguiculata L.). Agronomy 2022, 12, 676. https://doi.org/10.3390/agronomy12030676
Rangel-Montoya EA, Delgado-Ramírez CS, Sepulveda E, Hernández-Martínez R. Biocontrol of Macrophomina phaseolina Using Bacillus amyloliquefaciens Strains in Cowpea (Vigna unguiculata L.). Agronomy. 2022; 12(3):676. https://doi.org/10.3390/agronomy12030676
Chicago/Turabian StyleRangel-Montoya, Edelweiss Airam, Carmen Sanjuana Delgado-Ramírez, Edgardo Sepulveda, and Rufina Hernández-Martínez. 2022. "Biocontrol of Macrophomina phaseolina Using Bacillus amyloliquefaciens Strains in Cowpea (Vigna unguiculata L.)" Agronomy 12, no. 3: 676. https://doi.org/10.3390/agronomy12030676
APA StyleRangel-Montoya, E. A., Delgado-Ramírez, C. S., Sepulveda, E., & Hernández-Martínez, R. (2022). Biocontrol of Macrophomina phaseolina Using Bacillus amyloliquefaciens Strains in Cowpea (Vigna unguiculata L.). Agronomy, 12(3), 676. https://doi.org/10.3390/agronomy12030676