
CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Gene Expression Analysis
2.3. Cloning of CgbZIP1 Gene and Construction of Expression Vector
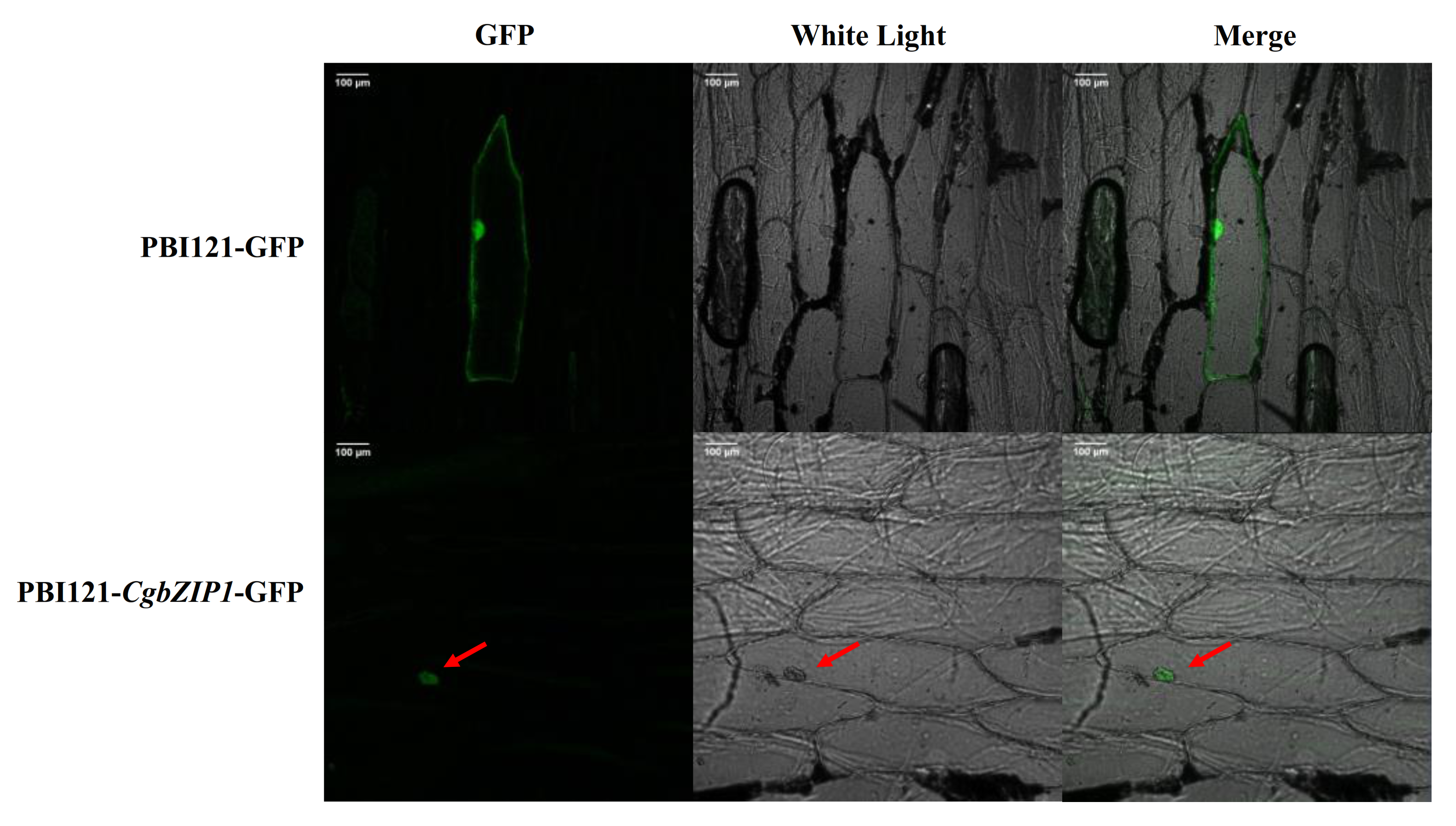
2.4. Subcellular Localization of CgbZIP1 Protein
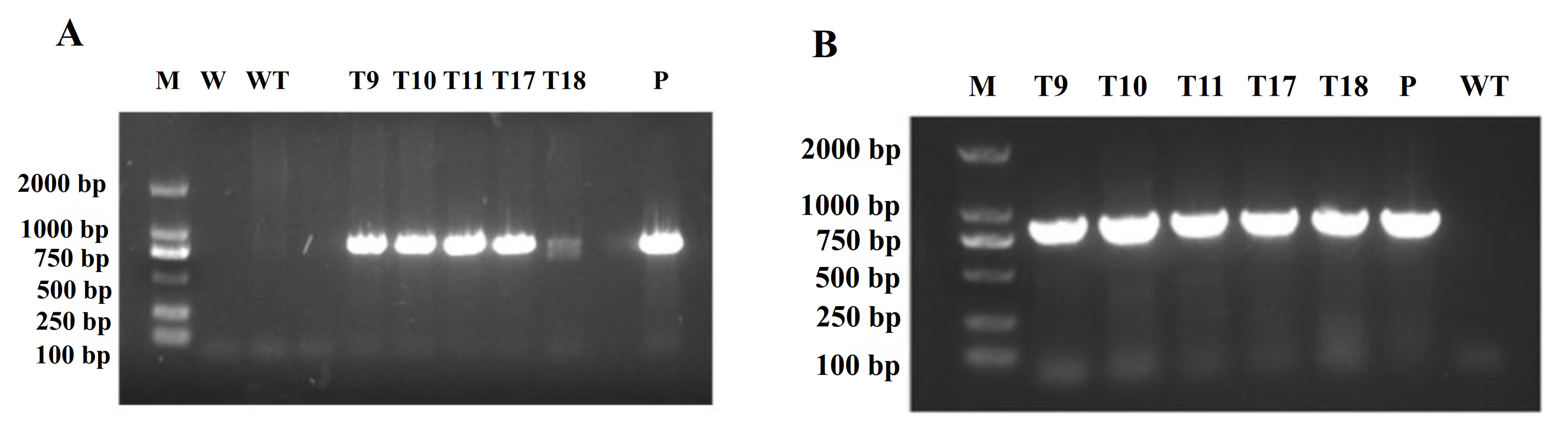
2.5. Production and Identification of Transgenic Tobacco
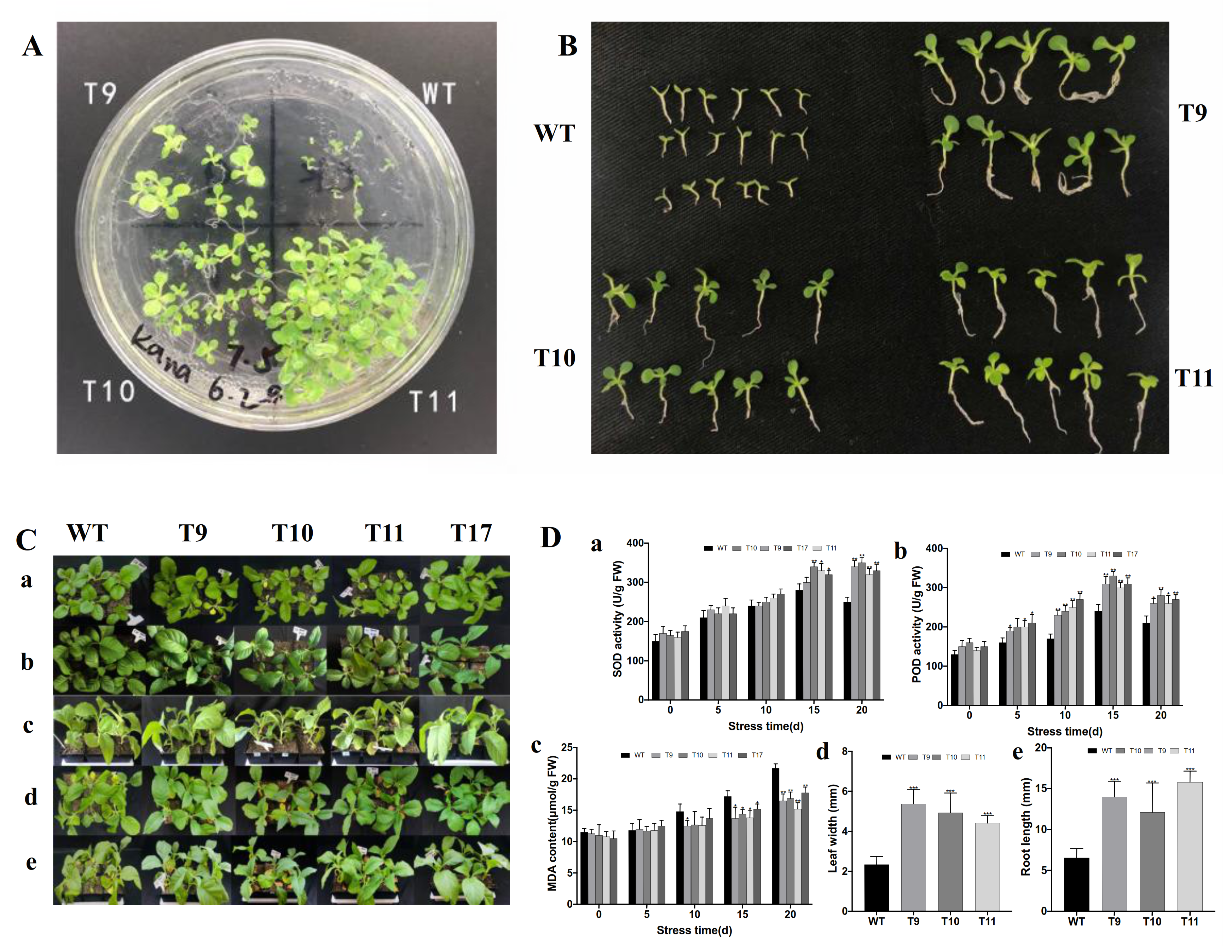
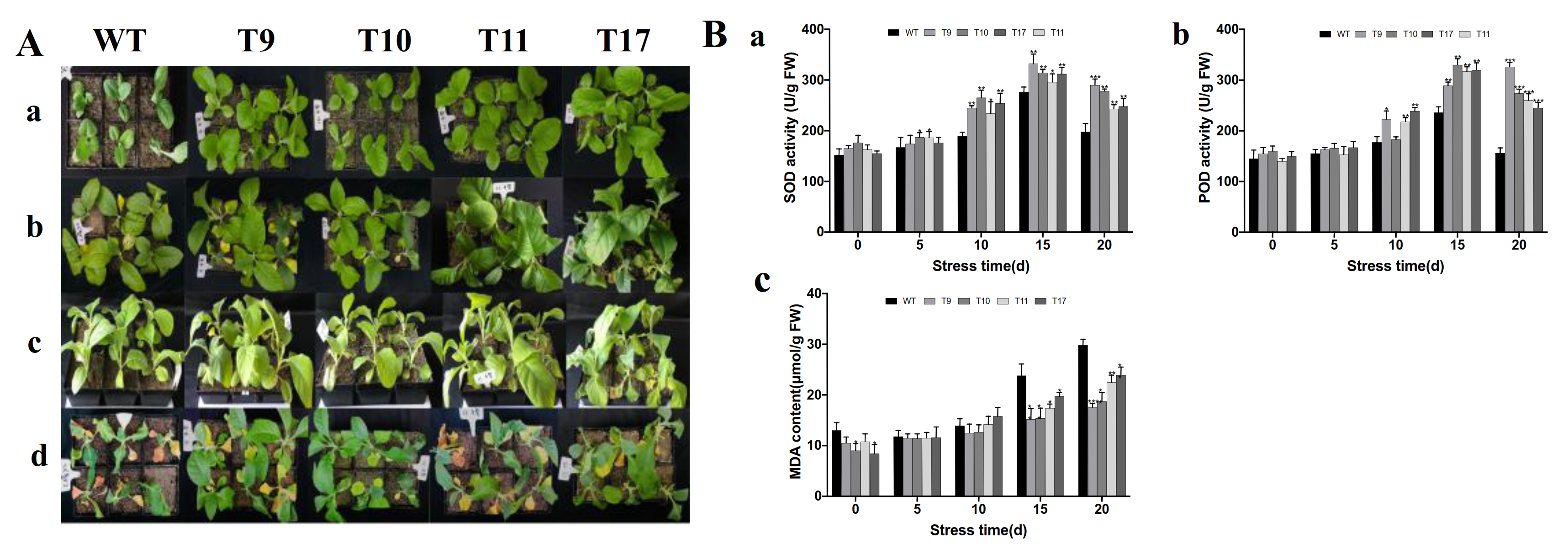
2.6. Assessment of Stress Resistance in Transgenic Tobacco
3. Results
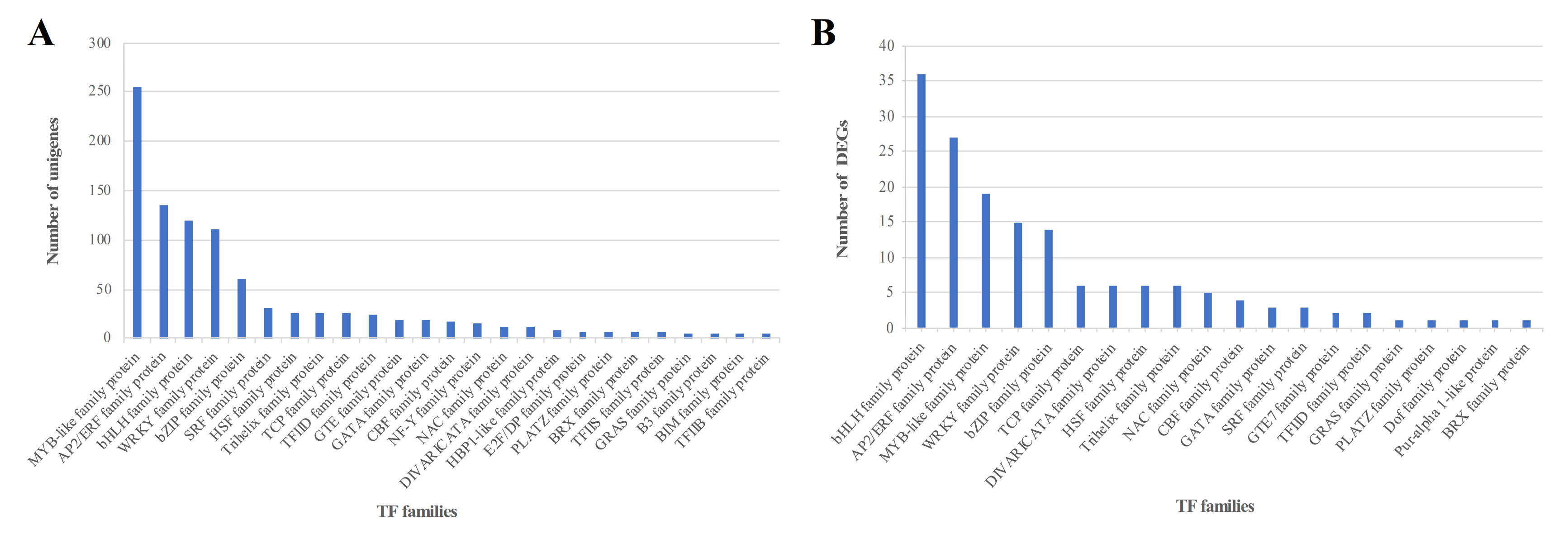
3.1. Identification of Differentially Expressed TF Genes in C. grandiflora in Response to Salt Stress
3.2. Molecular Characterization of CgbZIP1
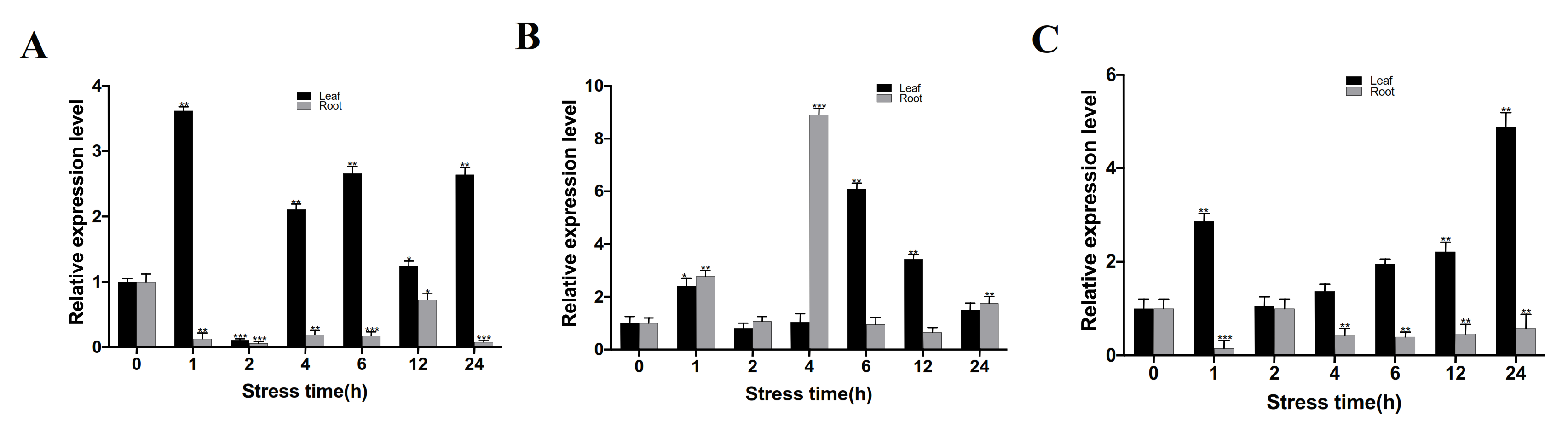
3.3. Spatiotemporal Expression Pattern of CgbZIP1 Gene under Abiotic Stresses and ABA Treatment
3.4. Subcellular Localization of CgbZIP1
3.5. Production of Transgenic Tobacco Overexpressing CgbZIP1
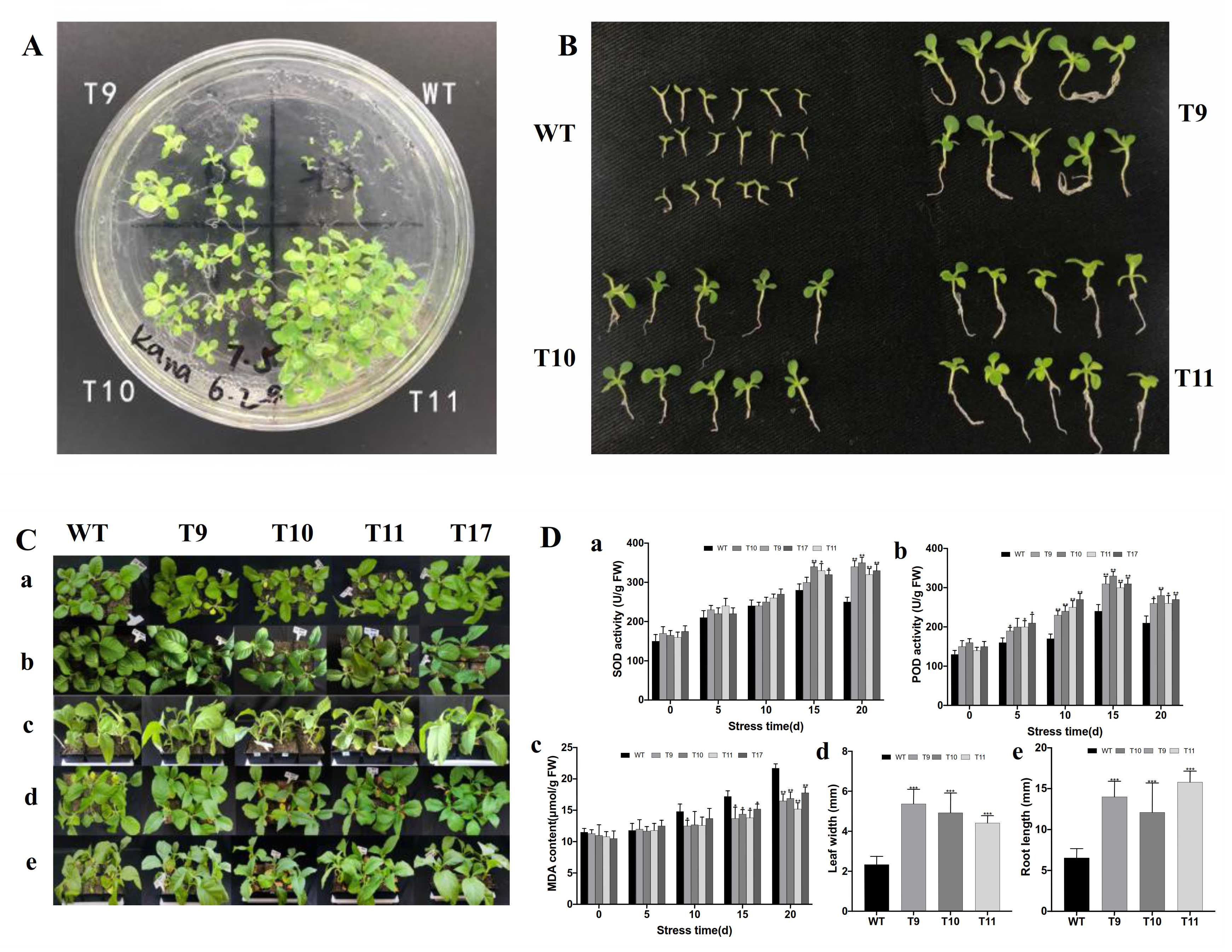
3.6. Transgenic Plants Overexpressing CgbZIP1 Exhibit Enhanced Tolerance to Salt Stress
3.7. Transgenic Plants Overexpressing CgbZIP1 Exhibit Enhanced Tolerance to Drought Stress
4. Discussion
4.1. TF Analysis Based on Transcriptome in Plants under Salt Stress
4.2. Molecular Mechanism of the bZIP TF-Mediated Plant Response to Abiotic Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, J.; Yang, R.; Li, W.; Pan, Y.; Zheng, M.; Zhang, Z. Effect of biochar amendment on water infiltration in a coastal saline soil. J. Soils Sediments 2018, 18, 3271–3279. [Google Scholar] [CrossRef]
- Zelm, E.V.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) Role in Plant Development and Coping with Different Stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef] [PubMed]
- Fricke, W. Energy costs of salinity tolerance in crop plants: Night-time transpiration and growth. New Phytol. 2020, 225, 1152–1165. [Google Scholar] [CrossRef]
- Che-Othman, M.H.; Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat mitochondrial respiration shifts from the tricarboxylic acid cycle to the GABA shunt under salt stress. New Phytol. 2020, 225, 1166–1180. [Google Scholar] [CrossRef]
- de la Torre-González, A.; Navarro-León, E.; Blasco, B.; Ruiz, J.M. Nitrogen and photorespiration pathways, salt stress genotypic tolerance effects in tomato plants (Solanum lycopersicum L.). Acta Physiol Plant. 2020, 42, 1–8. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Rodriguez-Uribe, L.; O’Connell, M.A. A root-specific bZIP transcription factor is responsive to water deficit stress in tepary bean (Phaseolus acutifolius) and common bean (P. vulgaris). J. Exp. Bot. 2006, 57, 1391–1398. [Google Scholar] [CrossRef]
- Schütze, K.; Harter, K.; Chaban, C. Post-translational regulation of plant bZIP factors. Trends Plant Sci. 2008, 13, 247–255. [Google Scholar] [CrossRef]
- Amir Hossain, M.; Lee, Y.; Cho, J.I.; Ahn, C.H.; Lee, S.K.; Jeon, J.S.; Kang, H.; Lee, C.H.; An, G.; Park, P.B. The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol. Biol. 2010, 72, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Zhang, D.F.; Fu, J.; Shi, Y.S.; Song, Y.C.; Wang, T.Y.; Li, Y. Cloning and characterization of a maize bZIP transcription factor, ZmbZIP72, confers drought and salt tolerance in transgenic Arabidopsis. Planta 2012, 235, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.H.; Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya Tsai, Y.C.; Chan, M.T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta 2010, 231, 1459–1473. [Google Scholar] [CrossRef]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wu, Y.H.; Tian, X.Q.; Bai, Z.Y.; Liang, Q.Y.; Liu, Q.L.; Pan, Y.Z.; Zhang, L.; Jiang, B.B. Overexpression of DgWRKY4 Enhances Salt Tolerance in Chrysanthemum Seedlings. Front Plant Sci. 2017, 8, 1592. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhong, M.; Wu, Y.H.; Bai, Z.Y.; Liang, Q.Y.; Liu, Q.L.; Pan, Y.Z.; Zhang, L.; Jiang, B.B.; Jia, Y.; et al. Overexpression of a chrysanthemum transcription factor gene DgNAC1 improves the salinity tolerance in chrysanthemum. Plant Cell Rep. 2017, 36, 571–581. [Google Scholar] [CrossRef]
- Liu, Q.L.; Xu, K.D.; Zhao, L.J.; Pan, Y.Z.; Jiang, B.B.; Zhang, H.Q.; Liu, G.L. Overexpression of a novel chrysanthemum NAC transcription factor gene enhances salt tolerance in tobacco. Biotechnol. Lett. 2011, 33, 2073–2082. [Google Scholar] [CrossRef]
- Zhou, L.; Shi, Q.; Qi, Y.; Miao, K.; Wang, Y. Transcriptome Analysis of Paeonia lutea and Its Differentially Expressed Genes Screening at Different Flower Opening Stages. Molecular Plant Breeding. 2018. Available online: http://kns.cnki.net/kcms/detail/46.1068.s.20181008.0928.016.html (accessed on 1 January 2022).
- Guo, S.C. Tanscriptome Sequencing Analysis of Sunflower Responsive to Verticillium Dahliae Infection and Mining of Resistance-related Genes. Ph.D. Thesis, Inner Mongolia University, Inner Mongolia, Hohhot, China, March 2017. [Google Scholar]
- Liu, C. Transcriptome Dynamics and Functions of RhIAA16 Genes during Rose Petal Abscission. Ph.D. Thesis, China Agricultural University, Beijing, China, September 2016. [Google Scholar]
- Wang, H.; Wang, W.; Wang, D.; Zhang, S.; Hu, X.; Lu, S.; Gong, X. De novo assembly and functional annotation of transcriptome data of Rhododendron pulchurum cv. Baifeng 4 leaf. Acta Agric. Zhejiangensis 2018, 30, 1149–1159. [Google Scholar]
- Zhang, J.; Shi, X.; Liu, H.; Ma, G.; Zou, Q.; Zhu, K.; Zhou, J.; Mao, J. Study on the Differential Accumulation of Anthocyanin in Different-colored Phalaenopsis Based on Transcriptomics. Mol. Plant Breed. 2018, 16, 4530–4542. [Google Scholar]
- Wang, L.; Yang, Y.; Liu, Y.; Yang, L.; Liu, Y.; Zhou, Y. Transcriptome analysis of Chrysanthemum × grandiflora in salt stress based on high-through-put sequencing. Mol. Plant Breed. 2020, 18, 1419–1427. [Google Scholar]
- Gao, W.J.; He, M.; Liu, J.; Ma, X.; Zhang, Y.; Dai, S.; Zhou, Y. Overexpression of Chrysanthemum lavandulifolium ClCBF1 in Chrysanthemum morifolium White Snow improves the level of salinity and drought tolerance. Plant Physiol. Biochem. 2018, 124, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Song, A.; Jiang, J.; Zhang, T.; Sun, H.; Wang, Y.; Chen, S.; Chen, F. CmWRKY1 enhances the dehydration tolerance of chrysanthemum through the regulation of ABA-associated genes. PLoS ONE 2016, 11, e0150572. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Chen, S.; Jiang, J.; Chen, F.; Chen, Y.; Gu, C.; Li, P.; Song, A.; Zhu, X.; Gao, H.; et al. Heterologous expression of the chrysanthemum r2r3-myb transcription factor cmmyb2 enhances drought and salinity tolerance, increases hypersensitivity to aba and delays flowering in arabidopsis thaliana. Mol. Biotechnol. 2012, 51, 160–173. [Google Scholar]
- Gallois, P.; Marinho, P. Leaf disk transformation using Agrobacterium tumefaciens-expression of heterologous genes in tobacco. Methods Mol. Biol. 1995, 49, 39–48. [Google Scholar] [CrossRef]
- Liu, Y.; He, M.; Dong, F.; Cai, Y.; Gao, W.; Zhou, Y.; Dai, S. The Chrysanthemum lavandulifolium ClNAC9 gene positively regulates saline, alkaline and drought stress in transgenic Chrysanthemum grandiflorum. J. Am. Soc. Hortic. Sci. 2019, 144, 280–288. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Li, X.; Zhang, D.; Gao, B.; Yang, H.; Wang, Y.; Guan, K.; Wood, A.J. ScDREB8, a novel A-5 type of DREB gene in the desert moss Syntrichia caninervis, confers salt tolerance to Arabidopsis. Plant Physiol. Biochem. 2017, 120, 242–251. [Google Scholar] [CrossRef]
- Liu, X.F.; Xiang, L.L.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. The identification of a MYB transcription factor controlling anthocyanin biosynthesis regulation in Chrysanthemum flowers. Sci. Hortic. 2015, 194, 278–285. [Google Scholar] [CrossRef]
- Zhou, X.T.; Jia, L.J.; Wang, H.Y.; Zhao, P.; Wang, W.Y.; Liu, N.; Song, S.W.; Wu, Y.; Su, L.; Zhang, J.; et al. The potato transcription factor StbZIP61 regulates dynamic biosynthesis of salicylic acid in defense against Phytophthora infestans infection. Plant J. 2018, 95, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. A novel sweetpotato bZIP transcription factor gene, IbbZIP1, is involved in salt and drought tolerance in transgenic Arabidopsis. Plant Cell Rep. 2019, 38, 1373–1382. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Shi, J.; Zhao, L. The CmbZIP1 transcription factor of chrysanthemum negatively regulates shoot branching. Plant Physiol. Biochem. 2020, 151, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Mohamed, S.; Regnault, T.; Mieulet, D.; Guiderdoni, E.; Sentenac, H.; Véry, A.-A. Constitutive Contribution by the Rice OsHKT1;4 Na+ Transporter to Xylem Sap Desalinization and Low Na+ Accumulation in Young Leaves Under Low as High External Na+ Conditions. Front. Plant Sci. 2020, 11, 1130. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Bao, X.; Zhi, Y.; Wu, Q.; Guo, Y.; Yin, X.; Zeng, L.; Li, J.; Zhang, J.; He, W.; et al. Overexpression of a MYB Family Gene, OsMYB6, Increases Drought and Salinity Stress Tolerance in Transgenic Rice. Front. Plant Sci. 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.B.; Wei, T.; Wang, L.H.; Hu, Y.W.; Liu, X.W.; Liu, Y.S. Kiwifruit (Actinidia chinensis) R1R2R3-MYB transcription factor AcMYB3R enhances drought and salinity tolerance in Arabidopsis thaliana. J. Integr. Agric. 2019, 18, 417–427. [Google Scholar] [CrossRef]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional regulation of bHLH during plant response to stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef]
- Bouaziz, D.; Pirrello, J.; Ben Amor, H.; Hammami, A.; Charfeddine, M.; Dhieb, A.; Bouzayen, M.; Gargouri-Bouzid, R. Ectopic expression of dehydration responsive element binding proteins (StDREB2) confers higher tolerance to salt stress in potato. Plant Physiol. Biochem. 2012, 60, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, D.; Li, X.; Guan, K.; Yang, H. Novel DREB A-5 subgroup transcription factors from desert moss (Syntrichia caninervis) confers multiple abiotic stress tolerance to yeast. J. Plant Physiol. 2016, 194, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D. An Abscisic Acid Signaling Pathway Starting from Plastid/Chloroplast. Chin. Bull. Bot. 2011, 46, 361–369. [Google Scholar]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Jiang, M.; Li, P.; Chu, Z. Genome-wide identification and evolutionary analyses of the PP2C gene family with their expression profiling in response to multiple stresses in Brachypodium distachyon. BMC Genom. 2016, 17, 175. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Meng, Z.; Meng, Z.; Malik, W.; Yan, R.; Lwin, K.M.; Lin, F.; Wang, Y.; Sun, G.; Zhou, T.; et al. GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum L.). Sci. Rep. 2016, 6, 35040. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.Y.; Hu, Y.F.; Li, J.J.; Yao, X.; Liu, K.D. BnaABF2, a bZIP transcription factor from rapeseed (Brassica napus L.), enhances drought and salt tolerance in transgenic Arabidopsis. Bot. Stud. 2016, 57, 12. [Google Scholar] [CrossRef] [Green Version]
- Llorca, C.M.; Potschin, M.; Zentgraf, U. bZIPs and WRKYs: Two large transcription factor families executing two different functional strategies. Front. Plant Sci. 2014, 5, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chu, Z. Genome-wide evolutionary characterization and analysis of bZIP transcription factors and their expression profiles in response to multiple abiotic stresses in Brachypodium distachyon. BMC Genom. 2015, 16, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Dal Santo, S.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281. [Google Scholar] [CrossRef] [Green Version]
- Baloglu, M.C.; Eldem, V.; Hajyzadeh, M.; Unver, T. Genome-wide analysis of the bZIP transcription factors in cucumber. PLoS ONE 2014, 9, e96014. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Liu, G.; Liu, Y.; Zheng, L.; Nie, X.; Wang, Y. The bZIP protein from Tamarix hispida, ThbZIP1, is ACGT elements binding factor that enhances abiotic stress signaling in transgenic Arabidopsis. BMC Plant Biol. 2013, 13, 151. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Liu, J.; Jia, J.; Kong, X. A novel wheat bZIP transcription factor, TabZIP60, confers multiple abiotic stress tolerances in transgenic Arabidopsis. Physiol. Plant. 2015, 153, 538–554. [Google Scholar] [CrossRef]
- Agarwal, P.; Baranwal, V.K.; Khurana, P. Genome-wide Analysis of bZIP Transcription Factors in wheat and Functional Characterization of a TabZIP under Abiotic Stress. Sci. Rep. 2019, 9, 4608. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene #ID | log2FC | Pfam_annotation | nr_annotation |
|---|---|---|---|
| c26412.graph_c0 | 3.98 | bZIP TF; Basic region leucine zipper | predicted protein [Hordeum vulgare subsp. vulgare] |
| c38401.graph_c0 | 1.18 | -- | PREDICTED: TF VIP1 isoform X2 [Nelumbo nucifera] |
| c39230.graph_c0 | −3.49 | bZIP TF; Basic region leucine zipper | hypothetical protein JCGZ_11804 [Jatropha curcas] |
| c43517.graph_c0 | 2.29 | bZIP TF | PREDICTED: TF RF2a-like [Solanum tuberosum] |
| c44927.graph_c0 | −2.56 | Homeobox domain; bZIP TF | hypothetical protein CISIN_1g0032502mg, partial [Citrus sinensis] |
| c45723.graph_c0 | −1.39 | Basic region leucine zipper; bZIP TF | PREDICTED: uncharacterized protein LOC103705064 [Phoenix dactylifera] |
| c48234.graph_c0 | 1.11 | Basic region leucine zipper; bZIP TF | unnamed protein product [Vitis vinifera] |
| c48785.graph_c1 | 7.80 | bZIP TF; Basic region leucine zipper | PREDICTED: ocs element-binding factor 1-like [Sesamum indicum] |
| c51234.graph_c0 | 1.33 | -- | PREDICTED: ABSCISIC ACID-INSENSITIVE 5-like protein 5 [Sesamum indicum] |
| c54119.graph_c1 | −1.33 | Basic region leucine zipper; bZIP TF | bZIP TF FD homolog [Chrysanthemum seticuspe f. boreale] |
| c54748.graph_c0 | 3.13 | G-box binding protein MFMR; bZIP TF; Basic region leucine zipper | hypothetical protein PRUPE_ppa006173mg [Prunus persica] |
| c55167.graph_c0 | 1.23 | bZIP TF; Basic region leucine zipper | BnaC04g52230D [Brassica napus] |
| c56024.graph_c0 | 1.21 | bZIP TF; Basic region leucine zipper | PREDICTED: G-box-binding factor 4 [Vitis vinifera] |
| c59327.graph_c0 | 1.05 | bZIP TF; Basic region leucine zipper | PREDICTED: probable TF PosF21 [Nicotiana tomentosiformis] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, L.; Liu, C.; Yin, H.; Liu, H.; Luo, H.; He, M.; Zhou, Y. CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress. Agronomy 2022, 12, 556. https://doi.org/10.3390/agronomy12030556
Liu Y, Wang L, Liu C, Yin H, Liu H, Luo H, He M, Zhou Y. CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress. Agronomy. 2022; 12(3):556. https://doi.org/10.3390/agronomy12030556
Chicago/Turabian StyleLiu, Yu, Lin Wang, Chenxu Liu, Hang Yin, He Liu, Hong Luo, Miao He, and Yunwei Zhou. 2022. "CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress" Agronomy 12, no. 3: 556. https://doi.org/10.3390/agronomy12030556
APA StyleLiu, Y., Wang, L., Liu, C., Yin, H., Liu, H., Luo, H., He, M., & Zhou, Y. (2022). CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress. Agronomy, 12(3), 556. https://doi.org/10.3390/agronomy12030556