
Monitoring and Inference of Behavioral Resistance in Beneficial Insects to Insecticides in Two Pest Control Systems: IPM and Organic


Abstract
:1. Introduction
2. Materials and Methods
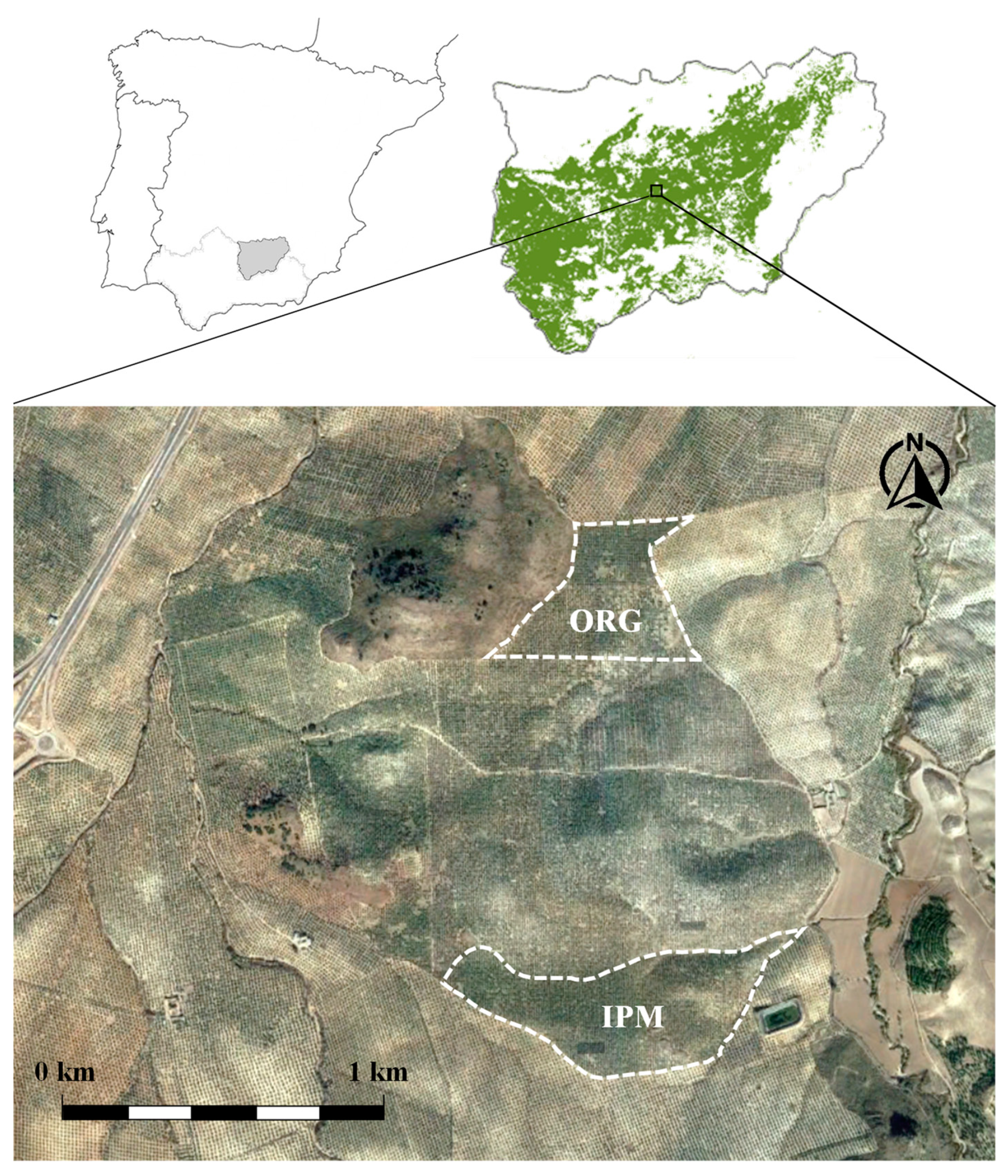
2.1. Description of the Study Area
2.2. Experimental Design
2.3. Statistical Analysis
3. Results
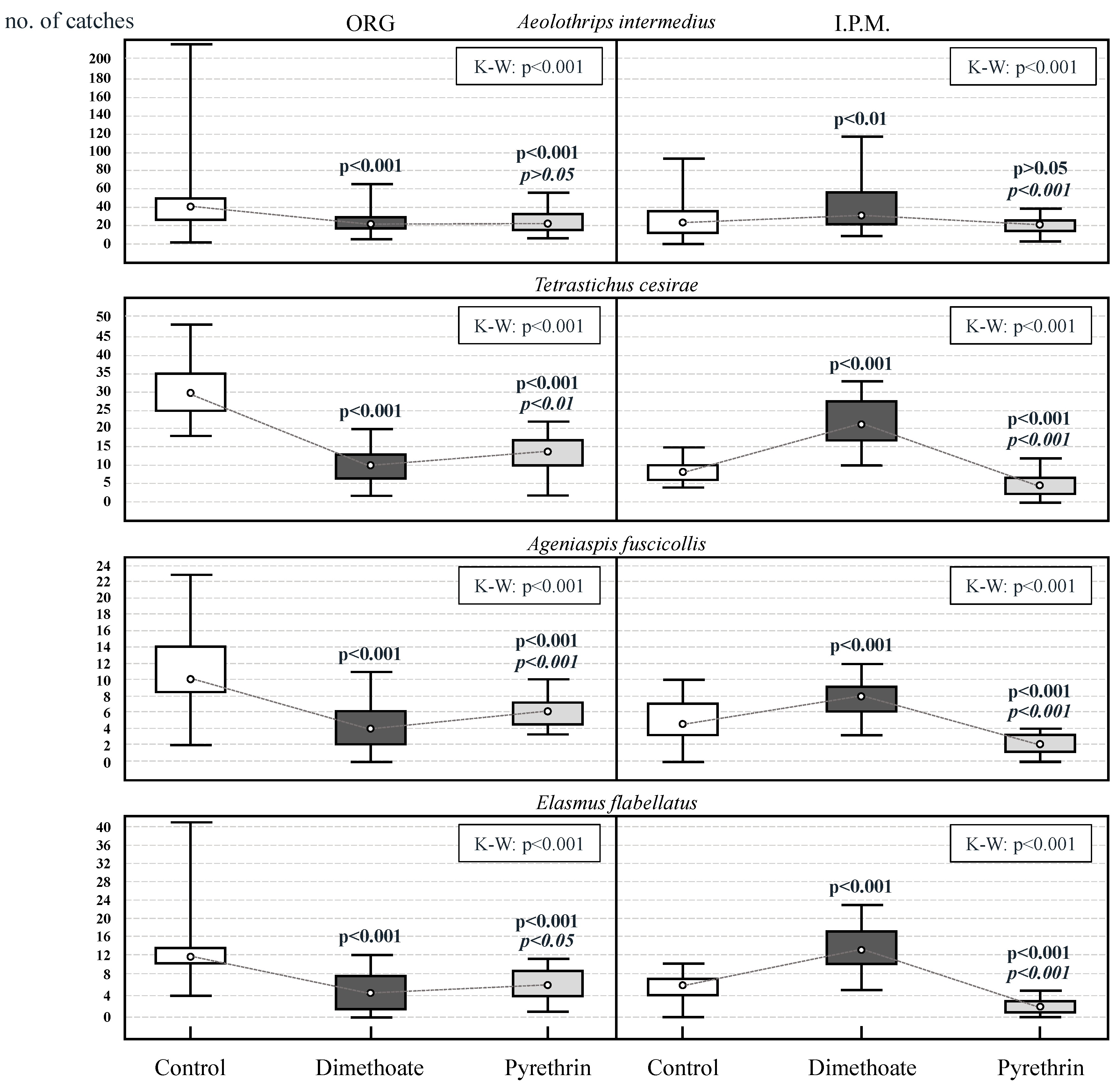
3.1. Analysis of the Relative Abundance of Beneficial Insects (Control Plots)
3.2. Behavioral Resistance Assessment (Plots Treated with Dimethoate/Pyrethrins)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stark, J.D.; Jepson, P.C.; Mayer, D.F. Limitations to use of topical toxicity data for predictions of pesticide side effects in the field. J. Econ. Entomol. 1995, 88, 1081–1088. [Google Scholar] [CrossRef]
- Lund, A.E.; Hollingworth, R.M.; Shankland, D.L. Chlordimeform: Plant protection by a sublethal, noncholinergic action on the central nervous system. Pestic. Biochem. Physiol. 1979, 11, 117–128. [Google Scholar] [CrossRef]
- Haynes, K.F. Sublethal effects of neurotoxic insecticides on insect behavior. Annu. Rev. Entomol. 1988, 33, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- De França, S.M.; Breda, M.O.; Barbosa, D.R.; Araujo, A.M.; Guedes, C.A. The sublethal effects of insecticides in insects. In Biological Control of Pest and Vector Insects; IntechOpen: London, UK, 2017; pp. 23–39. [Google Scholar]
- Lee, C.Y. Sublethal effects of insecticides on longevity, fecundity and behaviour of insect pests: A review. J. Biosci. 2000, 11, 107–112. [Google Scholar]
- Kongmee, M.; Prabaripai, A.; Akratanakul, P.; Bangs, M.J.; Chareonviriyaphap, T. Behavioral responses of Aedes aegypti (Diptera: Culicidae) exposed to deltamethrin and possible implications for disease control. J. Med. Entomol. 2004, 41, 1055–1063. [Google Scholar] [CrossRef]
- Correa, Y.D.C.G.; Faroni, L.R.; Haddi, K.; Oliveira, E.E.; Pereira, E.J.G. Locomotory and physiological responses induced by clove and cinnamon essential oils in the maize weevil Sitophilus zeamais. Pestic. Biochem. Physiol. 2015, 125, 31–37. [Google Scholar] [CrossRef]
- Haddi, K.; Oliveira, E.E.; Faroni, L.R.; Guedes, D.C.; Miranda, N.N. Sublethal exposure to clove and cinnamon essential oils induces hormetic-like responses and disturbs behavioral and respiratory responses in Sitophilus zeamais (Coleoptera: Curculionidae). J. Econ. Entomol. 2015, 108, 2815–2822. [Google Scholar] [CrossRef]
- Kennedy, J.S. The excitant and repellent effects on mosquitos of sub-lethal contacts with DDT. Bull. Entomol. Res. 1947, 37, 593–607. [Google Scholar] [CrossRef]
- Rolff, J.; Reynolds, S. (Eds.) Insect Infection and Immunity: Evolution, Ecology, and Mechanisms (No. 25); Oxford University Press on Demand: Oxford, UK, 2009. [Google Scholar]
- Van Dame, R.; Meled, M.; Colin, M.E.; Belzunces, L.P. Alteration of the homing-flight in the honey bee Apis mellifera L. Exposed to sublethal dose of deltamethrin. Environ. Toxicol. Chem. Int. J. 1995, 14, 855–860. [Google Scholar] [CrossRef]
- Pike, K.S.; Mayer, D.F.; Glazer, M.; Kious, C. Effects of permethrin on mortality and foraging behavior of honey bees in sweet corn. Environ. Entomol. 1982, 11, 951–953. [Google Scholar] [CrossRef]
- Quisenberry, S.S.; Lockwood, J.A.; Byford, R.L.; Wilson, H.K.; Sparks, T.C. Pyrethroid Resistance in the Horn Fly., Haematobia irritans (L.) (Diptera: Muscidae). J. Econ. Entomol. 1984, 77, 1095–1098. [Google Scholar] [CrossRef]
- Haynes, K.F.; Li, W.G.; Baker, T.C. Control of pink bollworm moth (Lepidoptera: Gelechiidae) with insecticides and pheromones (attracticide): Lethal and sublethal effects. J. Econ. Entomol. 1986, 79, 1466–1471. [Google Scholar] [CrossRef]
- Moore, R.F. Behavioral and biological effects of NRDC-161 as factors in control of the boll weevil. J. Econ. Entomol. 1980, 73, 265–267. [Google Scholar] [CrossRef]
- Chareonviriyaphap, T.; Bangs, M.J.; Suwonkerd, W.; Kongmee, M.; Corbel, V.; Ngoen-Klan, R. Review of insecticide resistance and behavioral avoidance of vectors of human diseases in Thailand. Parasites Vectors 2013, 6, 280. [Google Scholar] [CrossRef] [Green Version]
- González-Ruiz, R.; Varela, J.L.M. Chemical control of Prays oleae (Lep., Yponomeutidae) and its influence on natural enemies of olive grove pests. Phytoma España 2000, 115, 24–30. (In Spanish) [Google Scholar]
- Gómez-Guzmán, J.A.; García-Marín, F.J.; Sáinz-Pérez, M.; González-Ruiz, R. Behavioural Resistance in Insects: Its Potential Use as Bio Indicator of Organic Agriculture. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2017; Volume 95, p. 042038. [Google Scholar]
- Alvarado, M.; Civantos, M.; Duran, J.M. Plagas. In El Cultivo del Olivo; Junta de Andalousia and Editiones Mundi-Prensa: Madrid, Spain, 1997; pp. 401–459. [Google Scholar]
- Civantos, M. Defensa Fitosanitaria en Sistemas de Producción Integrada. In Cultivo del Olivar en Zonas de Especial Protección Ambiental. Informaciones Técnicas 65/99; Consejería de Agricultura y Pesca; Junta de Andalucía: Seville, Spain, 1999. [Google Scholar]
- Gerold, J.L.; Laarman, J.J. Behavioural responses to contact with DDT in Anopheles atroparvus. Nature 1967, 215, 518–520. [Google Scholar] [CrossRef]
- Gould, F. Role of behavior in the evolution of insect adaptation to insecticides and resistant host plants. Bull. ESA 1984, 30, 34–41. [Google Scholar] [CrossRef]
- Lockwood, J.A.; Sparks, T.C.; Story, R.N. Evolution of insect resistance to insecticides: A reevaluation of the roles of physiology and behavior. Bull. ESA 1984, 30, 41–51. [Google Scholar] [CrossRef]
- Pluthero, F.G.; Singh, R.S. Insect behavioural responses to toxins: Practical and evolutionary considerations. Can. Entomol. 1984, 116, 57–68. [Google Scholar] [CrossRef]
- Singh, J.P.; Marwaha, K.K. Effect of sublethal concentrations of some insecticides on growth and development of maize stalk borer, Chilo partellus (Swinhoe) larvae. Shashpa 2000, 7, 181–186. [Google Scholar]
- Kevan, P.G. Pollinators as bioindicators of the state of the environment: Species, activity and diversity. Agric. Ecosyst. Environ. 1999, 74, 373–393. [Google Scholar] [CrossRef]
- Thompson, H.M. Behavioural effects of pesticides in bees—Their potential for use in risk assessment. Ecotoxicology 2003, 12, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Guedes, N.M.P.; Guedes, R.N.C.; Ferreira, G.H.; Silva, L.B. Flight take-off and walking behavior of insecticide-susceptible and–resistant strains of Sitophilus zeamais exposed to deltamethrin. Bull. Entomol. Res. 2009, 99, 393–400. [Google Scholar] [CrossRef]
- Prodescon, S.A. Characterization of the Spanish Organic Production Sector in Terms of Value and Market, Referring to the Year 2015; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente: Madrid, Spain, 2016. [Google Scholar]
- Moral, A.M.; Uclés, D.F.; Jurado, E.B.; Viruel, M.J.M. The commitment to organic farming in the olive grove. A market for the future. Boletín Inst. Estud. Giennenses 2017, 216, 353–376. [Google Scholar]
- ESYRCE. Encuesta Sobre Superficie y Rendimientos de los Cultivos. 2020. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/esyrce/ (accessed on 11 July 2021).
- Gómez-Guzmán, J.A.; Sainz-Pérez, M.; González-Ruiz, R. Induction of sublethal effects for the characterization of Olive groves under different pest management systems. Rev. Bras. Frutic. 2021, 43. [Google Scholar] [CrossRef]
- Poolred. Available online: http://www.poolred.com/ (accessed on 7 September 2021).
- Henry, C.S.; Brooks, S.J.; Thierry, D.; Duelli, P.; Johnson, J.B.; McEwen, P.K.; New, T.R.; Whittington, A.E. The common green lacewing (Chrysoperla carnea s. lat.) and the sibling species problem. In Lacewings in the Crop Environment; Cambridge University Press: Cambridge, UK, 2001; pp. 29–42. [Google Scholar]
- Regnault-Roger, C.; Philogène, B.J.R.; Vincent, C. (Eds.) Produits phytosanitaires insecticides d’origine végétale: Promesses d’hier et d’aujourd’hui. In Biopesticides D’origine Végétale; Tec&Doc Editions: London, UK, 2002; pp. 1–18. [Google Scholar]
- Zapata, N.; Medina, P.; Viñuela, E.; Budia, F. Toxicidad de malation, pimetrocina, piretrinas naturales+ PBO y triflumuron en adultos del parasitoide Psyttalia concolor (Szepligeti)/(Hym.: Braconidae) según el modo de aplicación. Bol. San. Veg. Plagas 2005, 31, 111–118. [Google Scholar]
- Lafargue, G.L.; Medina, J.M.A.; Acosta, A.L.; Llanes, Y.M. Piretrinas y Piretroides. Anu. Cienc. UNAH 2018, 16, 4–13. [Google Scholar]
- Henry, C.W.; Shamsi, S.A.; Warner, I.M. Separation of natural pyrethrum extracts using micellar electrokinetic chromatography. J. Chromatogr. A 1999, 863, 89–103. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Bloomquist, J.R. Neurotoxic actions of pyrethroid insecticides. Annu. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef]
- Crosby, D.G. Environmental Fate of Pyrethrins; Oxford University Press: New York, NY, USA, 1995; pp. 194–213. [Google Scholar]
- Lybrand, D.B.; Xu, H.; Last, R.L.; Pichersky, E. How plants synthesize Pyrethrins: Safe and biodegradable insecticides. Trends Plant Sci. 2020, 25, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.V.; Holle, S.G.; Hutchison, W.D.; Koch, R.L. Lethal and sublethal effects of conventional and organic insecticides on the parasitoid Trissolcus japonicus, a biological control agent for Halyomorpha halys. Front. Insect Sci. 2021, 1, 5. [Google Scholar] [CrossRef]
- Todd, G.D.; Wohlers, D.; Citra, M.J. Toxicological Profile for Pyrethrins and Pyrethroids; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2003. [Google Scholar]
- Liu, F.; Wang, Q.; Xu, P.; Andreazza, F.; Valbon, W.R.; Bandason, E.; Chen, M.; Yan, R.; Feng, B.; Smith, L.B.; et al. A dual-target molecular mechanism of pyrethrum repellency against mosquitoes. Nat. Commun. 2021, 12, 2553. [Google Scholar] [CrossRef]
- Yan, R.; Zhou, Q.; Xu, Z.; Wu, Y.; Zhu, G.; Wang, M.; Guo, Y.; Dong, K.; Chen, M. Pyrethrins elicit olfactory response and spatial repellency in Aedes albopictus. Pest Manag. Sci. 2021, 77, 3706–3712. [Google Scholar] [CrossRef]
- Fine, B.C. Pattern of pyrethrin-resistance in houseflles. Nature 1961, 191, 884–885. [Google Scholar] [CrossRef] [PubMed]
- Cochran, D.G. Effects of synergists on bendiocarb and pyrethrins resistance in the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 1987, 80, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Weinzierl, R.; Henn, T. Botanical insecticides and insecticidal soaps. In Handbook of Integrated Pest Management for Turf and Ornamentals; CRC Press: Boca Raton, FL, USA, 1994; pp. 541–555. ISBN 9780138752798. Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/9780138752798-58/botanical-insecticides-insecticidal-soaps-richard-weinzierl-tess-henn (accessed on 26 January 2022).
- Cochran, D.G. Resistance to pyrethrins in the German cockroach: Inheritance and gene-frequency estimates in field-collected populations (Dictyoptera: Blattellidae). J. Econ. Entomol. 1994, 87, 280–284. [Google Scholar] [CrossRef]
- Kaufman, P.E.; Scott, J.G.; Rutz, D.A. Monitoring insecticide resistance in house flies (Diptera: Muscidae) from New York dairies. Pest Manag. Sci. 2001, 57, 514–521. [Google Scholar] [CrossRef]
- Speare, R.; Koehler, J.M. A case of pubic lice resistant to pyrethrins. Aust. Fam. Physician 2001, 30, 572–574. [Google Scholar]
- Fine, B.C. The present status of resistance to pyrethroid insecticides. Pyrethrum Post 1963, 7, 18–21. [Google Scholar]
- Lloyd, C.J.; Parkin, E.A. Further studies on a pyrethrum-resistant strain of the granary weevil, Sitophilus granarius (L.). J. Sci. Food Agric. 1963, 14, 655–663. [Google Scholar] [CrossRef]
- Lloyd, C.J. The toxicity of pyrethrins and five synthetic pyrethroids, to Tribolium castaneum (Herbst), and susceptible and pyrethrin-resistant Sitophilus granarius (L.). J. Stored Prod. Res. 1973, 9, 77–92. [Google Scholar] [CrossRef]
- Zettle, J.L.; McDonald, L.L.; Redlinger, L.M.; Jones, R.D. Plodia interpunctella and Cadra cautella resistance in strains to malathion and synergized pyrethrins. J. Econ. Entomol. 1973, 66, 1049–1050. [Google Scholar] [CrossRef]
- Zettler, L.J. Pesticide resistance in Tribolium castaneum and T. confusum (Coleoptera: Tenebrionidae) from flour mills in the United States. J. Econ. Entomol. 1991, 84, 763–767. [Google Scholar] [CrossRef]
- Lloyd, C.J.; Ruczkowski, G.E. The cross-resistance to pyrethrins and eight synthetic pyrethroids, of an organophosphorus-resistant strain of the rust-red flour beetle Tribolium castaneum (herbst). Pestic. Sci. 1980, 11, 331–340. [Google Scholar] [CrossRef]
- Stasinakis, P.; Katsares, V.; Mavragani-Tsipidou, P. Organophosphate Resistance and Allelic Frequencies of Esterases in the Olive Fruit Fly Bactrocera oleae (Diptera: Tephritidae). J. Agric. Urban Entomol. 2001, 18, 157–168. [Google Scholar]
- Skouras, P.G.; Margaritopoulos, J.T.; Seraphides, N.A.; Ioannides, I.M.; Kakani, E.G.; Mathiopoulos, K.D.; Tsitsipis, J.A. Organophosphate resistance in olive fruit fly, Bactrocera oleae, populations in Greece and Cyprus. Pest Manag. Sci. 2007, 63, 42–48. [Google Scholar] [CrossRef]
- Ramos, P.; Ramos, J.M. Veinte años de observaciones sobre la depredación oófaga en Prays oleae Bern. Granada (España), 1970–1989. Boletín Sanid. Veg. Plagas 1990, 16, 119–127. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insect: An Outline of Entomology; John Wiley & Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- Kranthi, K.R.; Ali, S.S.; Banerjee, S.K. Baseline toxicity of spinosad on the cotton bollworm, Helicoverpa armigera (Hub.), in India. Resist. Pest Manag. 2000, 11, 9–12. [Google Scholar]
- Moulton, J.K.; Pepper, D.A.; Dennehy, T.J. Beet armyworm (Spodoptera exigua) resistance to spinosad. Pest Manag. Sci. Former. Pestic. Sci. 2000, 56, 842–848. [Google Scholar] [CrossRef]
- Mau, R.F.; Gusukuma-Minuto, L. Diamondback moth resistance to spinosad (Success and Tracer, Dow Agro-Sciences) in Hawaii: Confirmation, review of causal factors and establishment of a mitigation plan. In Proceedings of the 5th International Seminar on Technology of Cole Crops Production, Celaya, Guanajuato, Mexico, 17–18 May 2001; pp. 75–80. [Google Scholar]
- Gunning, R.V.; Balfe, M.E. Spinosad resistance in Australian Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). In Proceedings of the 10th IUPAC International Congress on the Chemistry of Crop Protection, Basel, Switzerland, 4–9 August 2002; p. 290. [Google Scholar]
- García, V.B.Q. La Resistencia a Spinosad en Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Ph.D. Thesis, Universidad Politécnica de Cartagena, Cartagena, Spain, 2007. [Google Scholar]
- Bollero, A.L.; Moya, J.H.; Macías, V.V.; Mohedano, D.P.; Moya, J.C.H. Introducción al Olivar Ecológico en Andalucía; Instituto de Investigación y Formación Agraria y Pesquera, Junta de Andalucía: Seville, Spain, 2017. [Google Scholar]
- Delvare, G.; Aberlenc, H.P.; Adis, J.; Springate, N.D.; Stork, N.E.; Didham, R.K.; Basset, Y. A review of methods for sampling arthropods in tree canopies. Canopy Arthropods 1997, 27, 52. [Google Scholar]
- Young, M. Insects in flight. In Insect Sampling in Forest Ecosystems; Blackwell Publishing: Hoboken, NJ, USA, 2005; pp. 116–145. [Google Scholar]
- Dimitrova, A.; Livieratos, I.; Gkisakis, V. Trapping methodologies for functional canopy arthropod diversity in olive agroecosystem. In Proceedings of the 2nd Mediterranean Forum Research and Innovation as Tools for Sustainable Agriculture, Food & Nutrition Security, Bari, Italy, 18–20 September 2018. [Google Scholar]
- Arambourg, Y. Traité D’entomologie Oléicole; Conseil Oléicole International: Madrid, Spain, 1986. [Google Scholar]
- Hodkinson, I.D.; Hughes, M.K. La Fitofagia en los Insectos; Oikos-Tau: Barcelona, Spain, 1993. [Google Scholar]
- Andrés-Cantero, F. Enfermedades y Plagas del Olivo, 4th ed.; Riquelme y Vargas Ediciones, SL Jaén: Jaen, Spain, 2001; p. 646. [Google Scholar]
- Guerrero García, A. Nueva Olivicultura; Mundi-Prensa: Madrid, Spain, 2003; Volume 5, p. 225. [Google Scholar]
- Burrack, H.J.; Fornell, A.M.; Connell, J.H.; O’Connell, N.V.; Phillips, P.A.; Vossen, P.M.; Zalom, F.G. Intraspecific larval competition in the olive fruit fly (Diptera: Tephritidae). Environ. Entomol. 2009, 38, 1400–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M. Contribución al estudio de la entomocenosis de Prays oleae Bern. (Lep. Hyponomeutidae) en Granada (España). Acta Oecol. 1981, 2, 27–35. [Google Scholar]
- Varela, J.L.M.; González-Ruiz, R. Bases Metodológicas para la Evaluación del Impacto Ocasionado por las Aplicaciones Insecticidas Sobre los Enemigos Naturales de las Plagas del Olivo (II); Phytoma España: València, Spain, 1999; pp. 32–42. [Google Scholar]
- De Liñán, C. Entomología Agroforestal, Insectos y Ácaros que Dañan Montes, Cultivos y Jardines; Ediciones Aerotécnicas: Madrid, Spain, 1998. [Google Scholar]
- Nikolova, I.; Georgieva, N.; Tahsin, N. Toxicity of neem and pyrethrum products applied alone and in combination with different organic products to some predators and their population density. Rom. Agric. Res. 2015, 32, 291–301. [Google Scholar]
- Canard, M. Chrysopides (Neuroptera) Récoltés Dans les Oliveraies en Grèce; Bio. Gall. Hell: Toulouse, France, 1979. [Google Scholar]
- Alrouechdi, K. Les Chrysopides (Neuroptera) Récoltés dans une Oliveraie du sud-est de la France; Acta Oecologica/Oecologia Aplicata: Paris, France, 1980. [Google Scholar]
- Campos, M.; Ramos, P. Some relationships between the number of Prays oleae eggs laid on olive fruits and their predation by Chrysoperla carnea. In Integrated Pesticide Control Olive-Groves, Proceedings of the CEC/FAO/IOBC International Joint Meeting, Pisa, Italy, 3–6 April 1984; Balkema for CEC: Rotterdam, The Netherlands, 1985; pp. 237–241. [Google Scholar]
- Bozsik, A.; González-Ruiz Ruíz, R.; Lara, B.H. Distribution of the Chysoperla carnea Complex in Southern Spain (Neuroptera: Chrysopidae). In Protecția Mediului: Analele Universității din Oradea, Fascicula; Oradea, Romania, 2009; Volume 14, pp. 60–65. Available online: https://protmed.uoradea.ro/facultate/anale/protectia_mediului/2009/agr/11.Bozsikv%20Andras%202.pdf (accessed on 1 January 2022).
- González-Ruiz, R.G.; Al-Asaad, S.; Bozsik, A. Influencia de las masas forestales en la diversidad y abundancia de los crisópidos (Neur.: “Chrysopidae”) del olivar. Cuad. Soc. Española Cienc. For. 2008, 26, 33–38. [Google Scholar]
- González-Ruiz, R.; Gómez-Guzmán, J.A. Agricultural management greatly affects the beneficial entomofauna of the olive groves. Am. J. Biomed. Sci. Res. 2019, 1, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.A.; Pereira, J.A.; Torres, L.M.; Nogueira, A.J. Evaluation of the effects, on canopy arthropods, of two agricultural management systems to control pests in olive groves from north-east of Portugal. Chemosphere 2007, 67, 131–139. [Google Scholar] [CrossRef]
- Petacchi, R.; Minnocci, A. Impact of different Bactrocera oleae (Gmel) control strategies on olive-grove entomofauna. In Proceedings of the II International Symposium on Olive Growing, Jerusalem, Israel, 1 January 1994; ISHS Acta Horticulturae 356. Volume 356, pp. 399–402. [Google Scholar] [CrossRef]
- Rodríguez, E.; Peña, A.; Raya, A.J.S.; Campos, M. Evaluation of the effect on arthropod populations by using deltamethrin to control Phloeotribus scarabaeoides Bern. (Coleoptera: Scolytidae) in olive orchards. Chemosphere 2003, 52, 127–134. [Google Scholar] [CrossRef]
- Araya, M.; Araya, J.; Guerrero, M. Efectos de algunos insecticidas en dosis subletales sobre adultos de Aphidius ervi Haliday (Hymenoptera: Aphidiidae). Boletín Sanid. Vegetal. Plagas 2004, 30, 247–254. [Google Scholar]
- Umoru, P.A.; Powell, W. Sub-lethal effects of the insecticides pirimicarb and dimethoate on the aphid parasitoid Diaeretiella rapae (Hymenoptera: Braconidae) when attacking and developing in insecticide-resistant hosts. Biocontrol Sci. Technol. 2002, 12, 605–614. [Google Scholar] [CrossRef]
- Pascual, S.; Cobos, G.; Seris, E.; Sánchez-Ramos, I.; González-Núñez, M. Spinosad bait sprays against the olive fruit fly (Bactrocera oleae (Rossi)): Effect on the canopy non-target arthropod fauna. Int. J. Pest Manag. 2014, 60, 258–268. [Google Scholar] [CrossRef]
- Cochran, D.G. Insecticide Resistance. Understanding and Controlling the German Cockroach; Oxford University Press: New York, NY, USA, 1995; pp. 171–192. [Google Scholar]
- Busvine, J.R. Resistance to pyrethrins. Bull. World Health Organ. 1960, 22, 592. [Google Scholar] [PubMed]
- Morales, J.; Budia, F.; Viñuela, E. Efectos secundarios de cinco insecticidas sobre los diferentes estadios de desarrollo del parasitoide Hyposoter didymator (Thunberg) (Hymenoptera: Ichneumonidae). Bol. San. Veg. Plagas 2004, 30, 773–782. [Google Scholar]
- Schalk, J.M.; Shepard, B.M.; Stoner, K.A. Response of caterpillar pests and the parasite Diadegma insulare to collard cultivars and a pyrethrin insecticide. HortScience 1993, 28, 308–310. [Google Scholar] [CrossRef] [Green Version]
- Viñuela, E.; Handel, U.; Vogt, H. Evaluación en campo de los efectos secundarios de dos plaguicidas de origen botánico, una piretrina natural y un extracto de neem, sobre Chrysoperla carnea Steph. (Neuroptera: Chrysopidae). Bol. San. Veg. Plagas 1996, 22, 97–106. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Euzophera pinguis | Prays oleae | Phloeotribus scarabaeoides | Bactrocera oleae | ||||
|---|---|---|---|---|---|---|---|
| IPM | Pesticide: active principles | Pyrinex®: Chlorpyrifos 48% | Danadim®: Dimethoate 40% | Danadim®: Dimethoate 40% | Danadim®: Dimethoate 40% | ||
| Active ingredient per ha | 0.3 L/ha | 0.3 L/ha | 0.45 L/ha | 0.9 L/ha | |||
| No. applications (date) | 1 per year (May) | 1 per year (May) | 1 per year (June) | 2 per year (September + October) | |||
| ORG | Device/Pesticide:active principles | Delta traps + pheromone dispenser | Abanto®: Pyrethrin 4% | B. thuringiensis aizawaii | trap logs | Abanto®: Pyrethrin 4% | Spintor®: Spinosad 0.024% |
| Active ingredient per ha | 1 trap/ha | 0.6 L/ha | 1–2 kg/ha | - | 0.6 L/ha | 1 L/ha | |
| No. applications (date) | 1 per year (May) | 1 out of 4 years (May) | 3 out of 4 years (May) | - | 2 out of 5 years (September + October) | 1/year (October) | |
| Identified Species | Associated Olive Pest Species | Bibliographic References | |
|---|---|---|---|
| Predators | Anthocoris nemoralis (F., 1794) (Hem.: Anthocoridae) | Euphyllura olivina; Liothrips oleae; Prays oleae | Arambourg, 1986; Andrés-Cantero, 1997; Bejarano-Alcázar et al., 2011 |
| Orius laevigatus (Fieber, 1860) (Hem.: Anthocoridae) | Euphyllura olivina; Liothrips oleae; Prays oleae | Arambourg, 1986; Andrés-Cantero, 1997; Bejarano-Alcázar et al., 2011 | |
| Chrysoperla agilis Henry et al., 2003 (Neu.: Chrysopidae) | Euphyllura olivina; Prays oleae | Arambourg, 1986; Bozsik et al., 2009 | |
| Harraphidia laufferi (Navás, 1915) (Rap.: Raphidiidae) | Phloeotribus scarabaeoides | González-Ruiz, 1989; Rozas & González-Ruiz, 2017 | |
| Aeolothrips intermedius Bagnall, 1934 (Thy.: Aeolothripidae) | Liothrips oleae; Aceria oleae; Oxycenus maxwelli | De Liñán, 1998 | |
| Parasitoids | Chelonus elaeaphilus Silvestri, 1908 (Hym.: Braconidae) | Prays oleae | Campos, 1981; Arambourg, 1986; Carrero, 1996 |
| Ageniaspis fuscicollis (Dalman, 1820) (Hym.: Encyrtidae) | Prays oleae | Campos, 1981; Arambourg, 1986; Carrero, 1996 | |
| Pnigalio mediterraneus Ferrierre & Delucchi, 1957 (Hym.: Eulophidae) | Bactrocera oleae; Prays oleae | Neuenschwander et al., 1983; El-Heneidy et al., 2001 | |
| Tetrastichus cesirae Russo, 1938 (Hym.: Eulophidae) | Saissetia oleae; Bactrocera oleae; Liothrips oleae | Arambourg, 1986; De Andrés Cantero, 1997 | |
| Elasmus flabellatus (Fonscolombe, 1832) (Hym.: Eulophidae) | Prays oleae | Campos, 1981; Nave et al., 2017 | |
| Diadegma semiclausum (Hellén, 1949) (Hym.: Ichneumonidae) | Prays oleae | De Andrés Cantero, 1997 | |
| Itoplectis alternans (Gravenhorst, 1829) (Hym.: Ichneumonidae) | Prays oleae | De Andrés Cantero, 1997 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Guzmán, J.A.; Sainz-Pérez, M.; González-Ruiz, R. Monitoring and Inference of Behavioral Resistance in Beneficial Insects to Insecticides in Two Pest Control Systems: IPM and Organic. Agronomy 2022, 12, 538. https://doi.org/10.3390/agronomy12020538
Gómez-Guzmán JA, Sainz-Pérez M, González-Ruiz R. Monitoring and Inference of Behavioral Resistance in Beneficial Insects to Insecticides in Two Pest Control Systems: IPM and Organic. Agronomy. 2022; 12(2):538. https://doi.org/10.3390/agronomy12020538
Chicago/Turabian StyleGómez-Guzmán, José Alfonso, María Sainz-Pérez, and Ramón González-Ruiz. 2022. "Monitoring and Inference of Behavioral Resistance in Beneficial Insects to Insecticides in Two Pest Control Systems: IPM and Organic" Agronomy 12, no. 2: 538. https://doi.org/10.3390/agronomy12020538
APA StyleGómez-Guzmán, J. A., Sainz-Pérez, M., & González-Ruiz, R. (2022). Monitoring and Inference of Behavioral Resistance in Beneficial Insects to Insecticides in Two Pest Control Systems: IPM and Organic. Agronomy, 12(2), 538. https://doi.org/10.3390/agronomy12020538