
Providing Alternative Hosts and Nectar to Aphid Parasitoids in a Plum Orchard to Determine Resource Complementarity and Distance Range Effect on Biological Control

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Trophic Resources Provided by the Central Inter-Row
2.4. Estimation of Aphid Incidence
2.5. Parasitism Rate
2.6. Statistical Analyses
2.6.1. Resources Provided in the Inter-Row
2.6.2. Aphid and Parasitism Incidences and Parasitism Rates
3. Results
3.1. Availability of Resources in the Inter-Row
3.2. Aphid Incidence
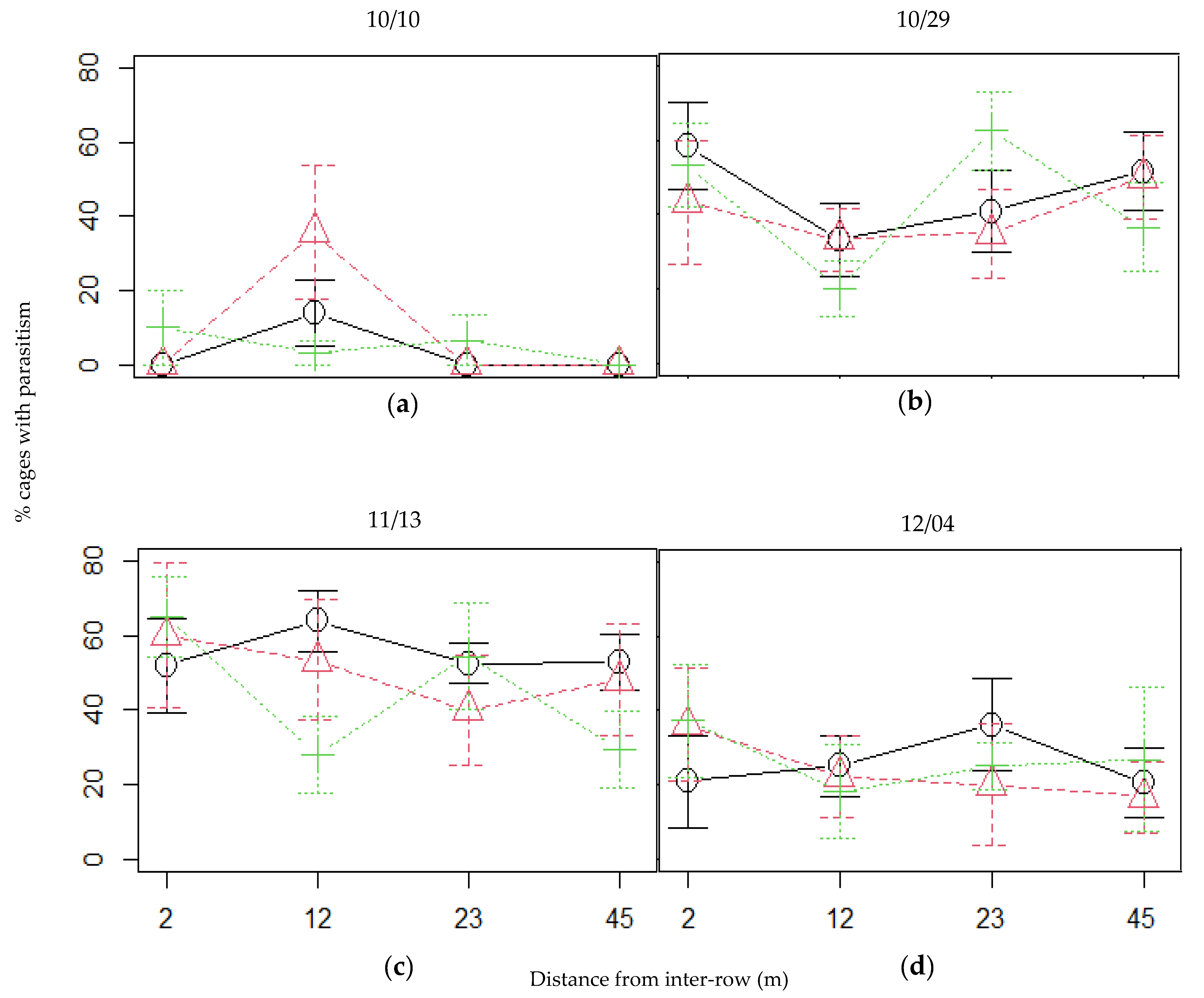
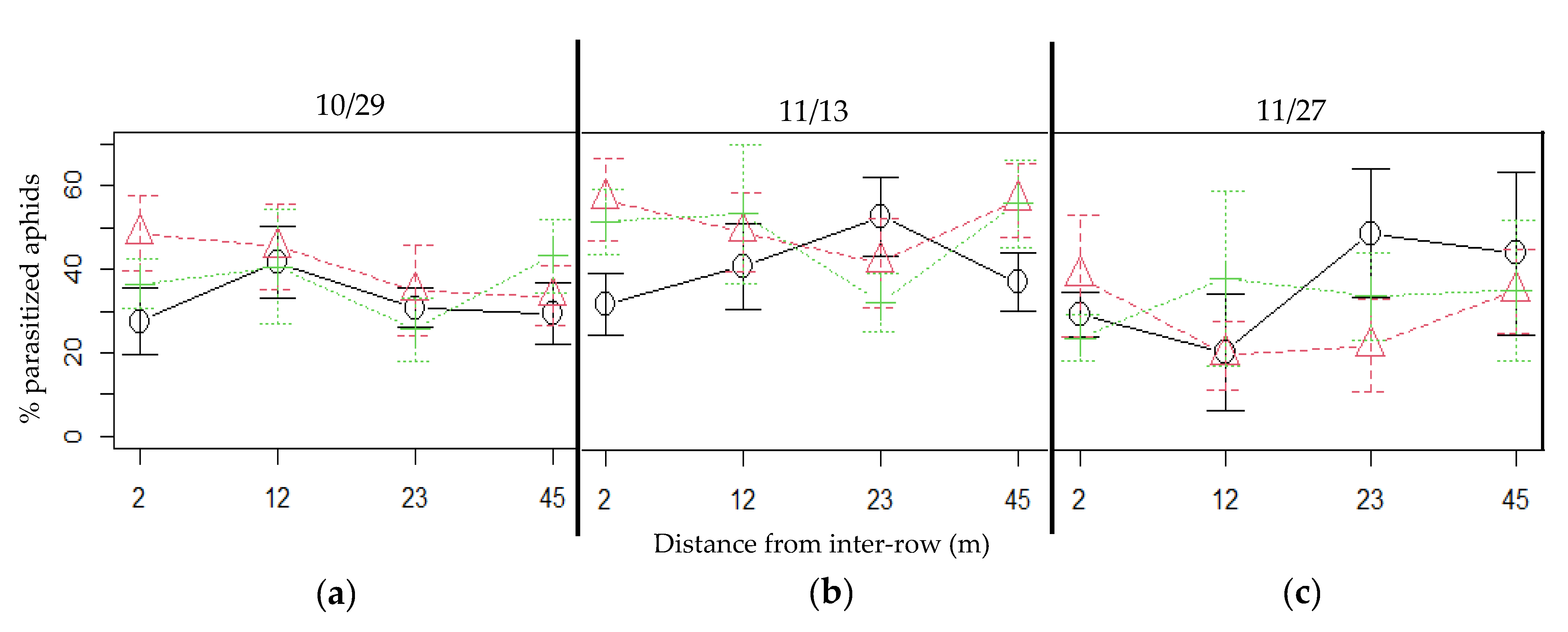
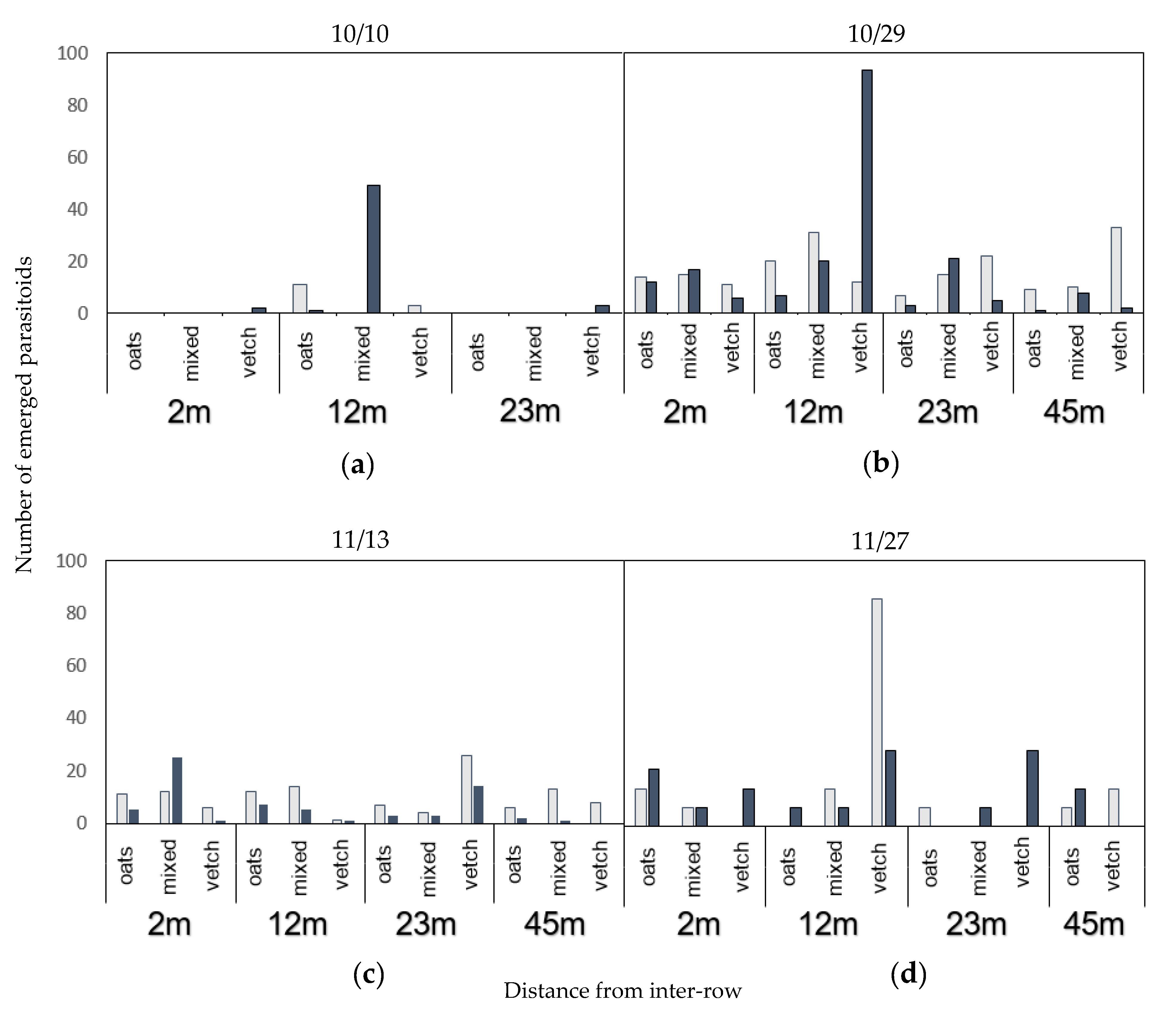
3.3. Incidence of Parasitism and Parasitism Rate
4. Discussion
4.1. Abundance, Quality and Temporality of Carbohydrates Provided in the Inter-Row
4.2. Abundance, Quality and Temporality of Alternative Hosts and Parasitism in the Inter-Row
4.3. Dispersal of Parasitoids between the Intercrops and the Plum Trees
4.4. Potential Negative Effect of the Inter-Row on Parasitism in Plum Trees
4.5. Plum Aphids Incidence and Their Regulation
4.6. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Langevelde, F.; Prins, H.H.T. Introduction to Resource Ecology. In Resource Ecology; Prins, H.H.T., Van Langevelde, F., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–6. ISBN 978-1-4020-6848-5. [Google Scholar]
- Prins, H.H.T.; Van Langevelde, F. Assembling a Diet from Different Places. In Resource Ecology; Prins, H.H.T., Van Langevelde, F., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 129–155. ISBN 978-1-4020-6848-5. [Google Scholar]
- Gillespie, M.A.K.; Gurr, G.M.; Wratten, S.D. Beyond Nectar Provision: The Other Resource Requirements of Parasitoid Biological Control Agents. Entomol. Exp. Appl. 2016, 159, 207–221. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Monographs in Behavior and Ecology; Princeton University Press: Princeton, NJ, USA, 1994; Volume 67, ISBN 0-691-00047-6. [Google Scholar]
- Leather, S.R.; Walters, K.F.A.; Bale, J.S. The Ecology of Insect Overwintering; Cambridge University Press: Cambridge, UK, 1995; ISBN 978-0-521-55670-5. [Google Scholar]
- Ballesteros, G.I.; Gadau, J.; Legeai, F.; Gonzalez-Gonzalez, A.; Lavandero, B.; Simon, J.-C.; Figueroa, C.C. Expression Differences in Aphidius Ervi (Hymenoptera: Braconidae) Females Reared on Different Aphid Host Species. PeerJ 2017, 5, e3640. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, L.S.; Nguyen, L.T.H.; Amiens-Desneux, E.; Luo, C.; Lavoir, A.; Gatti, J.; Desneux, N. The Preference–Performance Relationship as a Means of Classifying Parasitoids According to Their Specialization Degree. Evol. Appl. 2019, 12, 1626–1640. [Google Scholar] [CrossRef]
- Vinson, S.B. Biochemical Coevolution between Parasitoids and their Hosts. In Evolutionary Strategies of Parasitic Insects and Mites; Price, P.W., Ed.; Springer: Boston, MA, USA, 1975; pp. 14–48. ISBN 978-1-4615-8734-7. [Google Scholar]
- Neuville, S.; Le Ralec, A.; Outreman, Y.; Jaloux, B. The Delay in Arrival of the Parasitoid Diaeretiella Rapae Influences the Efficiency of Cabbage Aphid Biological Control. BioControl 2016, 61, 115–126. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Gagic, V.; Bommarco, R. Time Will Tell: Resource Continuity Bolsters Ecosystem Services. Trends Ecol. Evol. 2015, 30, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, A.; Klein, A.-M.; Weisser, W.W.; Tscharntke, T. Multitrophic Effects of Experimental Changes in Plant Diversity on Cavity-Nesting Bees, Wasps, and Their Parasitoids. Oecologia 2012, 169, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Tylianakis, J.M.; Tscharntke, T.; Klein, A.-M. Diversity, Ecosystem Function, and Stability of Parasitoid-Host Interactions Across a Tropical Habitat Gradient. Ecology 2006, 87, 3047–3057. [Google Scholar] [CrossRef]
- Aquilino, K.M.; Cardinale, B.J.; Ives, A.R. Reciprocal Effects of Host Plant and Natural Enemy Diversity on Herbivore Suppression: An Empirical Study of a Model Tritrophic System. Oikos 2005, 108, 275–282. [Google Scholar] [CrossRef]
- Wäckers, F.L. Suitability of (extra-)floral nectar, pollen, and honeydew as insect food sources. In Plant-Provided Food for Carnivorous Insects; Wackers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 17–74. ISBN 978-0-511-54222-0. [Google Scholar]
- Tena, A.; Senft, M.; Desneux, N.; Dregni, J.; Heimpel, G.E. The Influence of Aphid-Produced Honeydew on Parasitoid Fitness and Nutritional State: A Comparative Study. Basic Appl. Ecol. 2018, 29, 55–68. [Google Scholar] [CrossRef]
- Wäckers, F.L.; van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a Food Source for Natural Enemies: Making the Best of a Bad Meal? Biol. Control 2008, 45, 176–184. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Wratten, S.D. Improved Fitness of Aphid Parasitoids Receiving Resource Subsidies. Ecology 2004, 85, 658–666. [Google Scholar] [CrossRef]
- Jado, R.H.; Araj, S.-E.; Abu-Irmaileh, B.; Shields, M.W.; Wratten, S.D. Floral Resources to Enhance the Potential of the Parasitoid Aphidius Colemani for Biological Control of the Aphid Myzus Persicae. J. Appl. Entomol. 2019, 143, 34–42. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E.; Leibee, G.L. Comparing Floral Nectar and Aphid Honeydew Diets on the Longevity and Nutrient Levels of a Parasitoid Wasp. Entomol. Exp. Appl. 2004, 111, 189–199. [Google Scholar] [CrossRef]
- Idris, A.B.; Grafius, E. Wildflowers as Nectar Sources for Diadegma Insulare (Hymenoptera: Ichneumonidae), a Parasitoid of Diamondback Moth (Lepidoptera: Yponomeutidae). Environ. Entomol. 1995, 24, 1726–1735. [Google Scholar] [CrossRef]
- Heil, M. Nectar: Generation, Regulation and Ecological Functions. Trends Plant Sci. 2011, 16, 191–200. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E.; van Driesche, R.G. Nectar Availability and Parasitoid Sugar Feeding. In Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, HI, USA, 14–18 January 2002; Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2002; pp. 14–18. [Google Scholar]
- Koptur, S. Extrafloral Nectary-Mediated Interactions between Insects and Plants. In Insect-Plant Interactions; CRC Press: Boca Raton, FL, USA, 1992; pp. 89–138. [Google Scholar]
- Tena, A.; Pekas, A.; Cano, D.; Wäckers, F.L.; Urbaneja, A. Sugar Provisioning Maximizes the Biocontrol Service of Parasitoids. J. Appl. Ecol. 2015, 52, 795–804. [Google Scholar] [CrossRef]
- Irvin, N.A.; Pinckard, T.R.; Perring, T.M.; Hoddle, M.S. Evaluating the Potential of Buckwheat and Cahaba Vetch as Nectar Producing Cover Crops for Enhancing Biological Control of Homalodisca Vitripennis in California Vineyards. Biol. Control 2014, 76, 10–18. [Google Scholar] [CrossRef]
- Jamont, M.; Dubois-Pot, C.; Jaloux, B. Nectar Provisioning Close to Host Patches Increases Parasitoid Recruitment, Retention and Host Parasitism. Basic Appl. Ecol. 2014, 15, 151–160. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Wäckers, F.L. Effects of Flower Attractiveness and Nectar Availability in Field Margins on Biological Control by Parasitoids. Biol. Control 2008, 46, 400–408. [Google Scholar] [CrossRef]
- Azzouz, H.; Giordanengo, P.; Wäckers, F.L.; Kaiser, L. Effects of Feeding Frequency and Sugar Concentration on Behavior and Longevity of the Adult Aphid Parasitoid: Aphidius Ervi (Haliday) (Hymenoptera: Braconidae). Biol. Control 2004, 31, 445–452. [Google Scholar] [CrossRef]
- Rivero, A.; West, S.A. The Physiological Costs of Being Small in a Parasitic Wasp. Evol. Ecol. Res. 2002, 14, 407–420. [Google Scholar]
- Weisser, W.W.; Völkl, W. Dispersal in the Aphid Parasitoid, Lysiphlebus Cardui (Marshall) (Hym., Aphidiidae). J. Appl. Entomol. 1997, 121, 23–28. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Bianchi, F.J.J.A.; Hsu, C.L. Movement of Entomophagous Arthropods in Agricultural Landscapes: Links to Pest Suppression. Annu. Rev. Entomol. 2014, 59, 559–581. [Google Scholar] [CrossRef]
- Lavandero, B.; Wratten, S.; Shishehbor, P.; Worner, S. Enhancing the Effectiveness of the Parasitoid Diadegma Semiclausum (Helen): Movement after Use of Nectar in the Field. Biol. Control 2005, 34, 152–158. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E. Impact of Flowering Buckwheat on Lepidopteran Cabbage Pests and Their Parasitoids at Two Spatial Scales. Biol. Control 2005, 34, 290–301. [Google Scholar] [CrossRef]
- Sirot, E.; Bernstein, C. Time Sharing between Host Searching and Food Searching in Parasitoids: State-Dependent Optimal Strategies. Behav. Ecol. 1996, 7, 189–194. [Google Scholar] [CrossRef][Green Version]
- Lu, Z.-X.; Zhu, P.-Y.; Gurr, G.M.; Zheng, X.-S.; Read, D.M.Y.; Heong, K.-L.; Yang, Y.-J.; Xu, H.-X. Mechanisms for Flowering Plants to Benefit Arthropod Natural Enemies of Insect Pests: Prospects for Enhanced Use in Agriculture: Flowering Plants Benefit Natural Enemies. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef]
- Frank, S.D. Biological Control of Arthropod Pests Using Banker Plant Systems: Past Progress and Future Directions. Biol. Control 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Orre, G.U.S.; Tompkins, J.-M.; Jonsson, M.; Jacometti, M.A.; Wratten, S.D. Provision of Floral Resources for Biological Control Restoring an Important Ecosystem Service. Funct. Ecosyst. Commun. 2007, 1, 86–94. [Google Scholar]
- Koptur, S.; Jones, I.M.; Peña, J.E. The Influence of Host Plant Extrafloral Nectaries on Multitrophic Interactions: An Experimental Investigation. PLoS ONE 2015, 10, e0138157. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.W.; Mathews, C.R.; Krawczyk, G. Extrafloral Nectar in an Apple Ecosystem to Enhance Biological Control. J. Econ. Entomol. 2010, 103, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.W.; Mathews, C.R. Conservation Biological Control of Rosy Apple Aphid, Dysaphis Plantaginea (Passerini), in Eastern North America. Environ. Entomol. 2007, 36, 1131–1139. [Google Scholar] [CrossRef]
- Kishinevsky, M.; Cohen, N.; Chiel, E.; Wajnberg, E.; Keasar, T. Sugar Feeding of Parasitoids in an Agroecosystem: Effects of Community Composition, Habitat and Vegetation. Insect Conserv. Divers. 2018, 11, 50–57. [Google Scholar] [CrossRef]
- Jevremović, D.; Paunović, S.A.; Petrović-Obradović, O. Flight Dynamics and Species Composition of Aphids Landing on Plum and Apricot Leaves in the Orchards in Western Serbia. Phytoparasitica 2016, 44, 501–511. [Google Scholar] [CrossRef]
- Dedryver, C.-A.; Le Ralec, A.; Fabre, F. The Conflicting Relationships between Aphids and Men: A Review of Aphid Damage and Control Strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Bell, A.C. The Life-History of the Leaf-Curling Plum Aphid Brachycaudus Helichrysi in Northern Ireland and Its Ability to Transmit Potato Virus YC(AB). Ann. Appl. Biol. 1983, 102, 1–6. [Google Scholar] [CrossRef]
- Madsen, H.F.; Blair Bailey, J. Biology and Control of the Leaf Curl Plum Aphid in Northern California. J. Econ. Entomol. 1958, 51, 226–229. [Google Scholar] [CrossRef]
- Song, B.; Tang, G.; Sang, X.; Zhang, J.; Yao, Y.; Wiggins, N. Intercropping with Aromatic Plants Hindered the Occurrence of Aphis Citricola in an Apple Orchard System by Shifting Predator–Prey Abundances. Biocontrol Sci. Technol. 2013, 23, 381–395. [Google Scholar] [CrossRef]
- Song, B.Z.; Wu, H.Y.; Kong, Y.; Zhang, J.; Du, Y.L.; Hu, J.H.; Yao, Y.C. Effects of Intercropping with Aromatic Plants on the Diversity and Structure of an Arthropod Community in a Pear Orchard. BioControl 2010, 55, 741–751. [Google Scholar] [CrossRef]
- Payton Miller, T.L.; Rebek, E.J. Banker Plants for Aphid Biological Control in Greenhouses. J. Integr. Pest. Manag. 2018, 9, 9. [Google Scholar] [CrossRef]
- Huang, N.; Enkegaard, A.; Osborne, L.S.; Ramakers, P.M.J.; Messelink, G.J.; Pijnakker, J.; Murphy, G. The Banker Plant Method in Biological Control. Crit. Rev. Plant Sci. 2011, 30, 259–278. [Google Scholar] [CrossRef]
- Goh, H.G.; Kim, J.H.; Han, M.W. Application of Aphidius Colemani Viereck for Control of the Aphid in Greenhouse. J. Asia-Pac. Entomol. 2001, 4, 171–174. [Google Scholar] [CrossRef]
- Encyclop’Aphid. Available online: https://www6.inrae.fr/encyclopedie-pucerons (accessed on 22 September 2021).
- Starý, P. The Aphidiidae of Chile (Hymenoptera, Ichneumonoidea, Aphidiidae). Dtsch. Entomol. Z. 1995, 42, 113–138. [Google Scholar] [CrossRef]
- Alfaro-Tapia, A.; Alvarez-Baca, J.K.; Tougeron, K.; Lavandero, B.; Le Lann, C.; Van Baaren, J. Overwintering Strategies and Life-History Traits of Different Populations of Aphidius Platensis along a Latitudinal Gradient in Chile. Entomol. Gen. 2021, 98641. [Google Scholar] [CrossRef]
- Alvarez-Baca, J.K.; Alfaro-Tapia, A.; Lavandero, B.; Le Lann, C.; Van Baaren, J. Suitability and Profitability of a Cereal Aphid for the Parasitoid Aphidius Platensis in the Context of Conservation Biological Control of Myzus Persicae in Orchards. Insects 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Géneau, C.E.; Wäckers, F.L.; Luka, H.; Daniel, C.; Balmer, O. Selective Flowers to Enhance Biological Control of Cabbage Pests by Parasitoids. Basic Appl. Ecol. 2012, 13, 85–93. [Google Scholar] [CrossRef]
- Repullo-Ruibérriz de Torres, M.A.; Moreno-García, M.; Ordóñez-Fernández, R.; Rodríguez-Lizana, A.; Cárceles Rodríguez, B.; García-Tejero, I.F.; Durán Zuazo, V.H.; Carbonell-Bojollo, R.M. Cover Crop Contributions to Improve the Soil Nitrogen and Carbon Sequestration in Almond Orchards (SW Spain). Agronomy 2021, 11, 387. [Google Scholar] [CrossRef]
- McKinlay, R.G. Vegetable Crop Pests, 1st ed.; Palgrave Macmillan: London, UK, 1992. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Petrović, A.; Mitrović, M.; Kavallieratos, N.G.; Starý, P.; Rakhshani, E.; Rakhshanipour, M.; Popović, A.; Shukshuk, A.H.; Ivanović, A. Molecular and Morphological Variability within the Aphidius Colemani Group with Redescription of Aphidius Platensis Brethes (Hymenoptera: Braconidae: Aphidiinae). Bull. Entomol. Res. 2014, 104, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Mansion-Vaquié, A.; Wezel, A.; Ferrer, A. Wheat Genotypic Diversity and Intercropping to Control Cereal Aphids. Agric. Ecosyst. Environ. 2019, 285, 106604. [Google Scholar] [CrossRef]
- Xu, Q.; Hatt, S.; Lopes, T.; Zhang, Y.; Bodson, B.; Chen, J.; Francis, F. A Push–Pull Strategy to Control Aphids Combines Intercropping with Semiochemical Releases. J. Pest. Sci. 2018, 91, 93–103. [Google Scholar] [CrossRef]
- Ndzana, R.A.; Magro, A.; Bedoussac, L.; Justes, E.; Journet, E.-P.; Hemptinne, J.-L. Is There an Associational Resistance of Winter Pea-Durum Wheat Intercrops towards Acyrthosiphon Pisum Harris? J. Appl. Entomol. 2014, 138, 577–585. [Google Scholar] [CrossRef]
- Seidenglanz, M.; Huňady, I.; Poslušná, J.; Løes, A.-K. Influence of Intercropping with Spring Cereals on the Occurrence of Pea Aphids (Acyrthosiphon Pisum Harris, 1776) and Their Natural Enemies in Field Pea (Pisum Sativum L.). Plant Protect. Sci. 2011, 47, 25–36. [Google Scholar] [CrossRef]
- Heimpel, G.E. Linking Parasitoid Nectar Feeding and Dispersal in Conservation Biological Control. Biol. Control 2019, 132, 36–41. [Google Scholar] [CrossRef]
- Hales, D.; Tomiuk, J.; Wöhrmann, K.; Sunnucks, P. Evolutionary and Genetic Aspects of Aphid Biology: A Review. Eur. J. Entomol. 1997, 94, 1–55. [Google Scholar]
- Ohta, I.; Honda, K. Use of Sitobion Akebiae (Hemiptera: Aphididae) as an Alternative Host Aphid for a Banker-Plant System Using an Indigenous Parasitoid, Aphidius Gifuensis (Hymenoptera: Braconidae). Appl. Entomol. Zool. 2010, 45, 233–238. [Google Scholar] [CrossRef][Green Version]
- Yano, E. Augmentation of Natural Enemies for Pest Control in Protected Culture. Pestic. Outlook 2003, 14, 247–250. [Google Scholar] [CrossRef]
- Starý, P. Aphidius Platensis Brethes, Its Distribution and Host Range (Hym.: Aphidiidae). Orient. Insects 1972, 6, 359–370. [Google Scholar] [CrossRef]
- Luquet, M.; Moulin, C.; Cortesero, A.-M.; Anton, S.; Jaloux, B. Early Experience Influences Several Steps of the Host Selection Process Differentially in Aphidius Ervi (Hymenoptera, Braconidae). J. Asia-Pac. Entomol. 2020, 23, 1235–1240. [Google Scholar] [CrossRef]
- Derocles, S.A.P.; Navasse, Y.; Buchard, C.; Plantegenest, M.; Le Ralec, A. “Generalist” Aphid Parasitoids Behave as Specialists at the Agroecosystem Scale. Insects 2019, 11, 6. [Google Scholar] [CrossRef]
- Derocles, S.A.P.; Le Ralec, A.; Besson, M.M.; Maret, M.; Walton, A.; Evans, D.M.; Plantegenest, M. Molecular Analysis Reveals High Compartmentalization in Aphid-Primary Parasitoid Networks and Low Parasitoid Sharing between Crop and Noncrop Habitats. Mol. Ecol. 2014, 23, 3900–3911. [Google Scholar] [CrossRef]
- Langhof, M.; Meyhöfer, R.; Poehling, H.-M.; Gathmann, A. Measuring the Field Dispersal of Aphidius Colemani (Hymenoptera: Braconidae). Agric. Ecosyst. Environ. 2005, 107, 137–143. [Google Scholar] [CrossRef]
- Luquet, M.; Peñalver-Cruz, A.; Satour, P.; Anton, S.; Cortesero, A.-M.; Lavandero, B.; Jaloux, B. Aphid Honeydew May Be the Predominant Sugar Source for Aphidius Parasitoids Even in Nectar-Providing Intercrops. Biol. Control 2021, 158, 104596. [Google Scholar] [CrossRef]
- Snyder, W.E.; Ives, A.R. Interactions Between Specialist and Generalist Natural Enemies: Parasitoids, Predators, and Pea Aphid Biocontrol. Ecology 2003, 84, 91–107. [Google Scholar] [CrossRef]
- Chacón, J.M.; Heimpel, G.E. Density-Dependent Intraguild Predation of an Aphid Parasitoid. Oecologia 2010, 164, 213–220. [Google Scholar] [CrossRef]
- Batuecas, I.; Agustí, N.; Castañé, C.; Alomar, O. Molecular Tracking of Insect Dispersal to Verify Arthropod Predator Movement from an Alfalfa Field to a Peach Orchard. Biol. Control 2021, 158, 104506. [Google Scholar] [CrossRef]
- Raymond, L.; Ortiz-Martínez, S.A.; Lavandero, B. Temporal Variability of Aphid Biological Control in Contrasting Landscape Contexts. Biol. Control 2015, 90, 148–156. [Google Scholar] [CrossRef]
- Pollier, A.; Tricault, Y.; Plantegenest, M.; Bischoff, A. Sowing of Margin Strips Rich in Floral Resources Improves Herbivore Control in Adjacent Crop Fields: Floral Resources Improve Herbivore Control. Agric. For. Entomol. 2018. [Google Scholar] [CrossRef]
- Saeed, R.; Razaq, M.; Hardy, I.C.W. The Importance of Alternative Host Plants as Reservoirs of the Cotton Leaf Hopper, Amrasca Devastans, and Its Natural Enemies. J. Pest. Sci. 2015, 88, 517–531. [Google Scholar] [CrossRef]
- Jacometti, M.; Jørgensen, N.; Wratten, S. Enhancing Biological Control by an Omnivorous Lacewing: Floral Resources Reduce Aphid Numbers at Low Aphid Densities. Biol. Control 2010, 55, 159–165. [Google Scholar] [CrossRef]
- Scarratt, S.L.; Wratten, S.D.; Shishehbor, P. Measuring Parasitoid Movement from Floral Resources in a Vineyard. Biol. Control 2008, 46, 107–113. [Google Scholar] [CrossRef]
- Jones, I.M.; Koptur, S.; von Wettberg, E.J. The Use of Extrafloral Nectar in Pest Management: Overcoming Context Dependence. J. Appl. Ecol. 2017, 54, 489–499. [Google Scholar] [CrossRef]
- Mansion-Vaquié, A.; Ferrer, A.; Ramon-Portugal, F.; Wezel, A.; Magro, A. Intercropping Impacts the Host Location Behaviour and Population Growth of Aphids. Entomol. Exp. Appl. 2019, 168, 41–52. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observation Date | Intercrop Modality | Rp * | Sa | Sm | Md | Sg * | Ap | Ac * | Mp * | Bh | Af * |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 11 July | Oats | 13.6 | 0.8 | 0.07 | 0.27 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 10.8 | 1.13 | 0 | 0.27 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Vetch | 0.13 | 0.33 | 0 | 0 | 0.2 | 0 | 0 | 0 | 0 | 0 | |
| 21 August | Oats | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 0.53 | 0.13 | 0 | 0.07 | 0.07 | 0 | 0 | 0 | 0 | 0 | |
| Vetch | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.93 | 0 | 0 | 0.7 | |
| 26 September | Oats | 7.67 | 2.47 | 2.93 | 4.47 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 1.13 | 0.47 | 0 | 1.1 | 0 | 0.5 | 0.23 | 0.13 | 0.23 | 0.03 | |
| Vetch | 0 | 0 | 0 | 0 | 0 | 2.1 | 1.53 | 0.47 | 0 | 0.73 | |
| 10 October | Oats | 1.47 | 0.07 | 7.87 | 1.33 | 0.4 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 1.83 | 0.57 | 0.97 | 2.1 | 1.3 | 0.6 | 0 | 0 | 0 | 0.07 | |
| Vetch | 0.13 | 0 | 0 | 0 | 0 | 4.33 | 1.33 | 0.27 | 0.07 | 0.13 | |
| 29 October | Oats | 1.13 | 6.27 | 0.73 | 1.07 | 0.6 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 0.77 | 1.33 | 1.9 | 0.33 | 0.73 | 1.57 | 0.43 | 0 | 0 | 0 | |
| Vetch | 0 | 0 | 0 | 0 | 0 | 17.87 | 1.13 | 0 | 0 | 0 | |
| 13 November | Oats | 0.2 | 16.27 | 1.67 | 0.07 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mixed | 0.13 | 4.77 | 0.37 | 0.07 | 0 | 0.8 | 0 | 0 | 0 | 0 | |
| Vetch | 0 | 0 | 0 | 0 | 0 | 13.27 | 0.13 | 0 | 0 | 0 |
| Date | Explained Variable | Fixed Effect (Random Effect) | ANOVA Test Following GLMMs (Df = 2) | Pairwise Comparisons 1 (Emmeans) p-Values (Df = 55) | |||
|---|---|---|---|---|---|---|---|
| Chisq | p-Value | Oats/Mixed | Oats/Vetch | Mixed/Vetch | |||
| 21 August | Alternative hosts | Intercrop (replicate) | 5.1441 | 0.076 | / | / | / |
| Honeydew providers | Intercrop (replicate) | 2.4817 | 0.289 | / | / | / | |
| 26 September | Alternative hosts | Intercrop (replicate) | 12.132 | 0.002 | 0.002 | 0.046 | 0.453 |
| Honeydew providers | Intercrop (replicate) | 31.136 | <0.001 | <0.001 | 0.001 | 0.473 | |
| 10 October | Alternative hosts | Intercrop (replicate) | 1.301 | 0.522 | / | / | / |
| Honeydew providers | Intercrop (replicate) | 2.0423 | 0.360 | / | / | / | |
| 29 October | Alternative hosts | Intercrop (replicate) | 1.0802 | 0.583 | / | / | / |
| Honeydew providers | Intercrop (replicate) | 26.000 | <0.001 | 0.187 | 0.005 | <0.001 | |
| 13 November | Alternative hosts | Intercrop (replicate) | 0.178 | 0.915 | / | / | / |
| Honeydew providers | Intercrop (replicate) | 14.847 | <0.001 | <0.001 | 0.295 | 0.015 | |
| Date | Intercrop Modality | Ae | Ap | Au | Lt | Pg | Pv | P sp. | H | NI | NE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 21 August | Oats | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 1 |
| Mixed | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 4 | |
| Vetch | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 26 September | Oats | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 35 |
| Mixed | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | |
| Vetch | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | |
| 10 October | Oats | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 5 | 2 |
| Mixed | 7 | 1 | 0 | 0 | 4 | 1 | 0 | 0 | 4 | 6 | |
| Vetch | 2 | 0 | 0 | 8 | 0 | 0 | 0 | 5 | 1 | 20 | |
| 29 October | Oats | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 7 |
| Mixed | 1 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 1 | 7 | |
| Vetch | 23 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 1 | 8 | |
| 13 November | Oats | 1 | 0 | 10 | 0 | 0 | 0 | 4 | 0 | 12 | 9 |
| Mixed | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 6 | 9 | |
| Vetch | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lérault, L.; Clavel, E.; Villegas, C.M.; Cabrera, N.; Jaloux, B.; Plantegenest, M.; Lavandero, B. Providing Alternative Hosts and Nectar to Aphid Parasitoids in a Plum Orchard to Determine Resource Complementarity and Distance Range Effect on Biological Control. Agronomy 2022, 12, 77. https://doi.org/10.3390/agronomy12010077
Lérault L, Clavel E, Villegas CM, Cabrera N, Jaloux B, Plantegenest M, Lavandero B. Providing Alternative Hosts and Nectar to Aphid Parasitoids in a Plum Orchard to Determine Resource Complementarity and Distance Range Effect on Biological Control. Agronomy. 2022; 12(1):77. https://doi.org/10.3390/agronomy12010077
Chicago/Turabian StyleLérault, Louise, Elsa Clavel, Cinthya M. Villegas, Nuri Cabrera, Bruno Jaloux, Manuel Plantegenest, and Blas Lavandero. 2022. "Providing Alternative Hosts and Nectar to Aphid Parasitoids in a Plum Orchard to Determine Resource Complementarity and Distance Range Effect on Biological Control" Agronomy 12, no. 1: 77. https://doi.org/10.3390/agronomy12010077
APA StyleLérault, L., Clavel, E., Villegas, C. M., Cabrera, N., Jaloux, B., Plantegenest, M., & Lavandero, B. (2022). Providing Alternative Hosts and Nectar to Aphid Parasitoids in a Plum Orchard to Determine Resource Complementarity and Distance Range Effect on Biological Control. Agronomy, 12(1), 77. https://doi.org/10.3390/agronomy12010077