
Heat Waves and Broomrape Are the Major Constraints for Lentil Cultivation in Southern Spain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Statistical Analysis
2.2.1. Variances Analyses
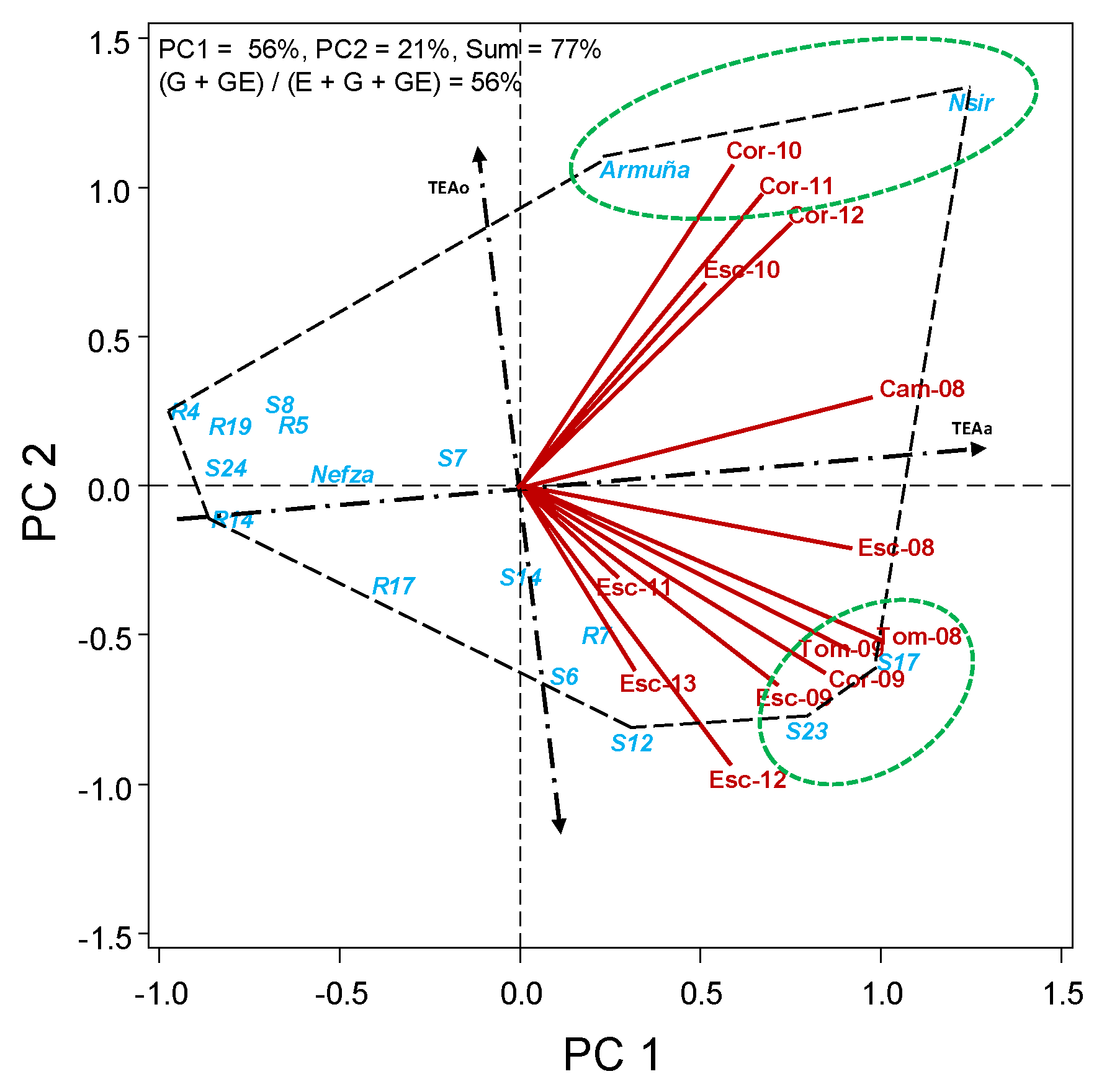
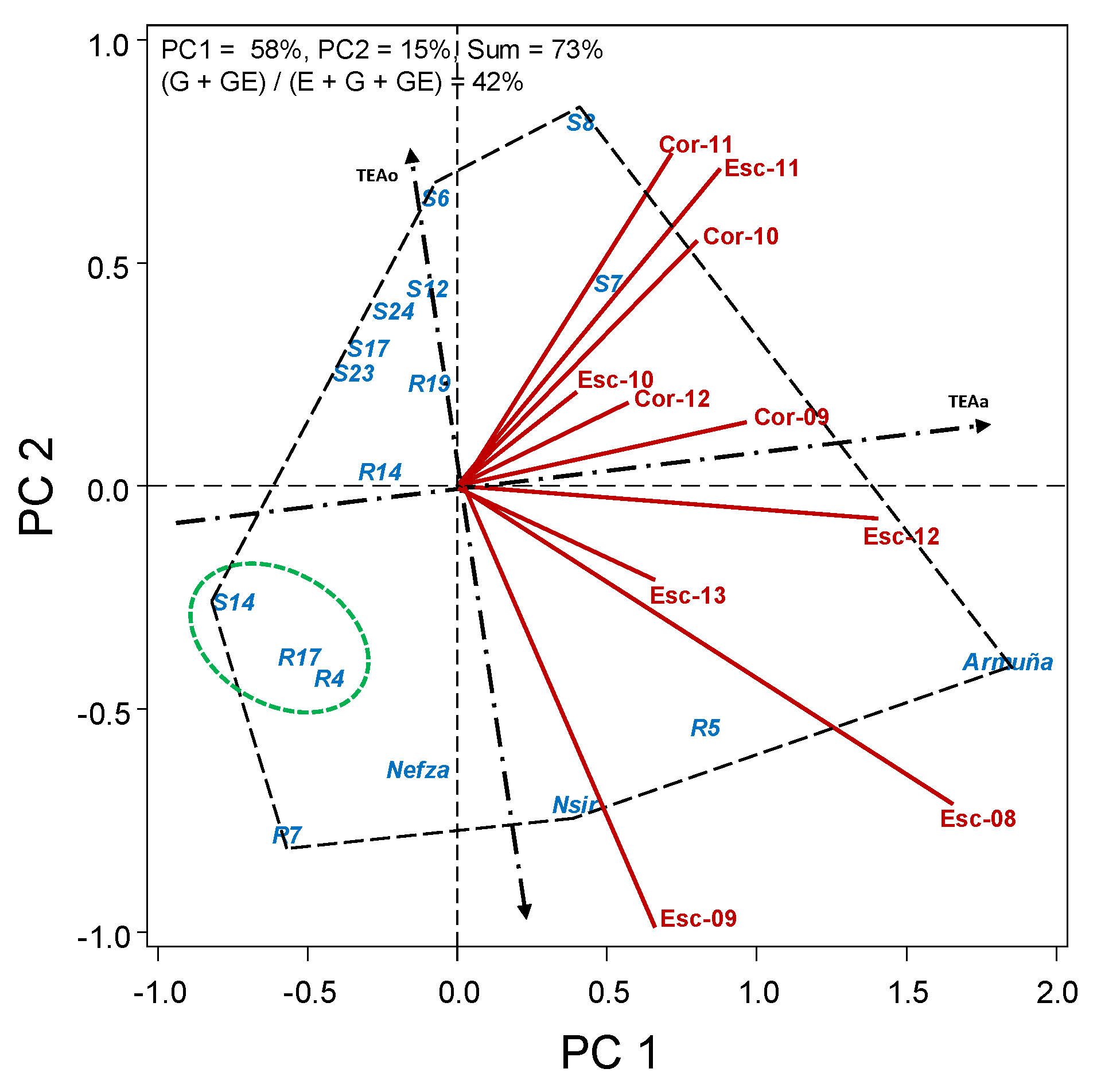
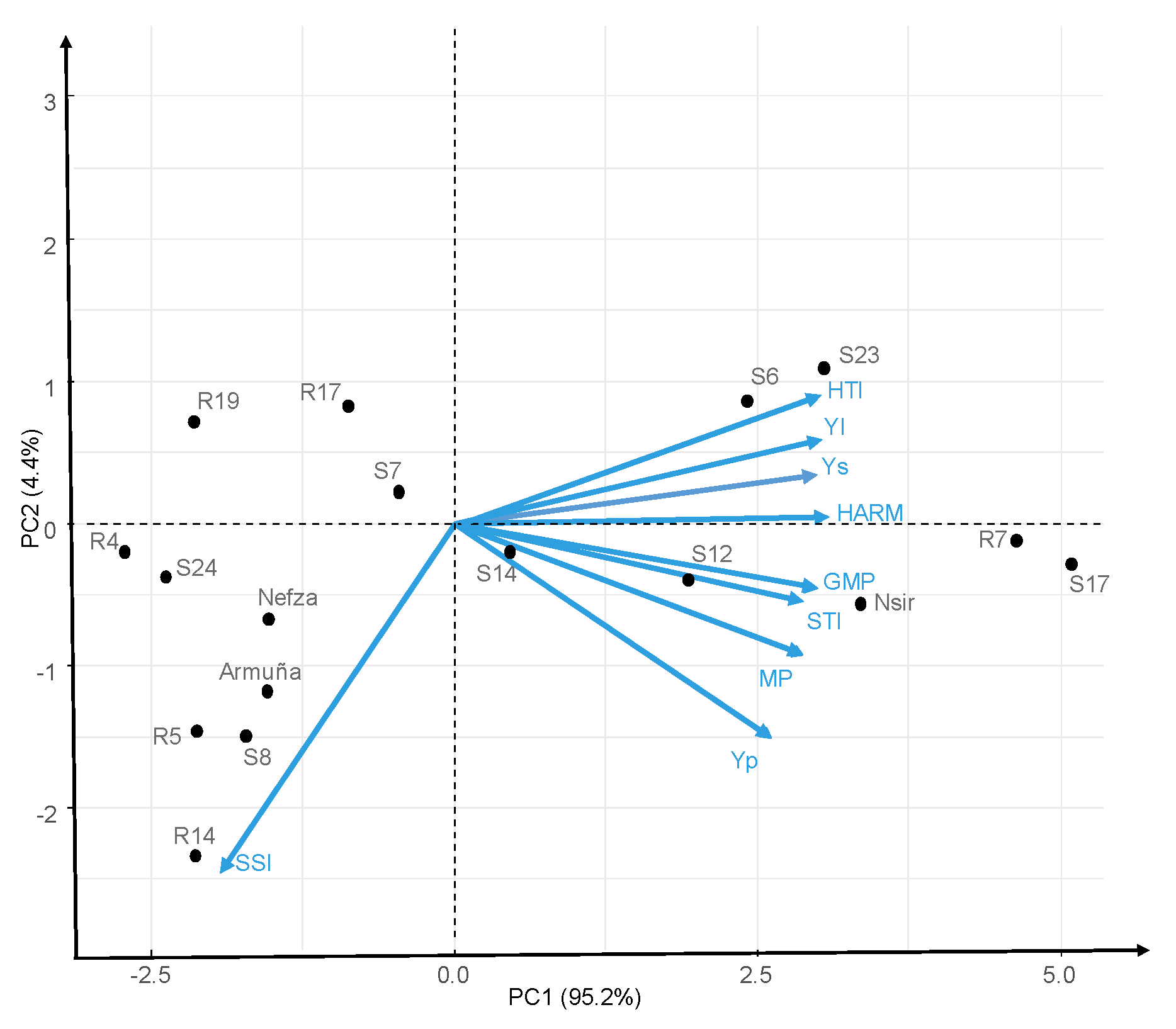
2.2.2. HA-GGE Biplots
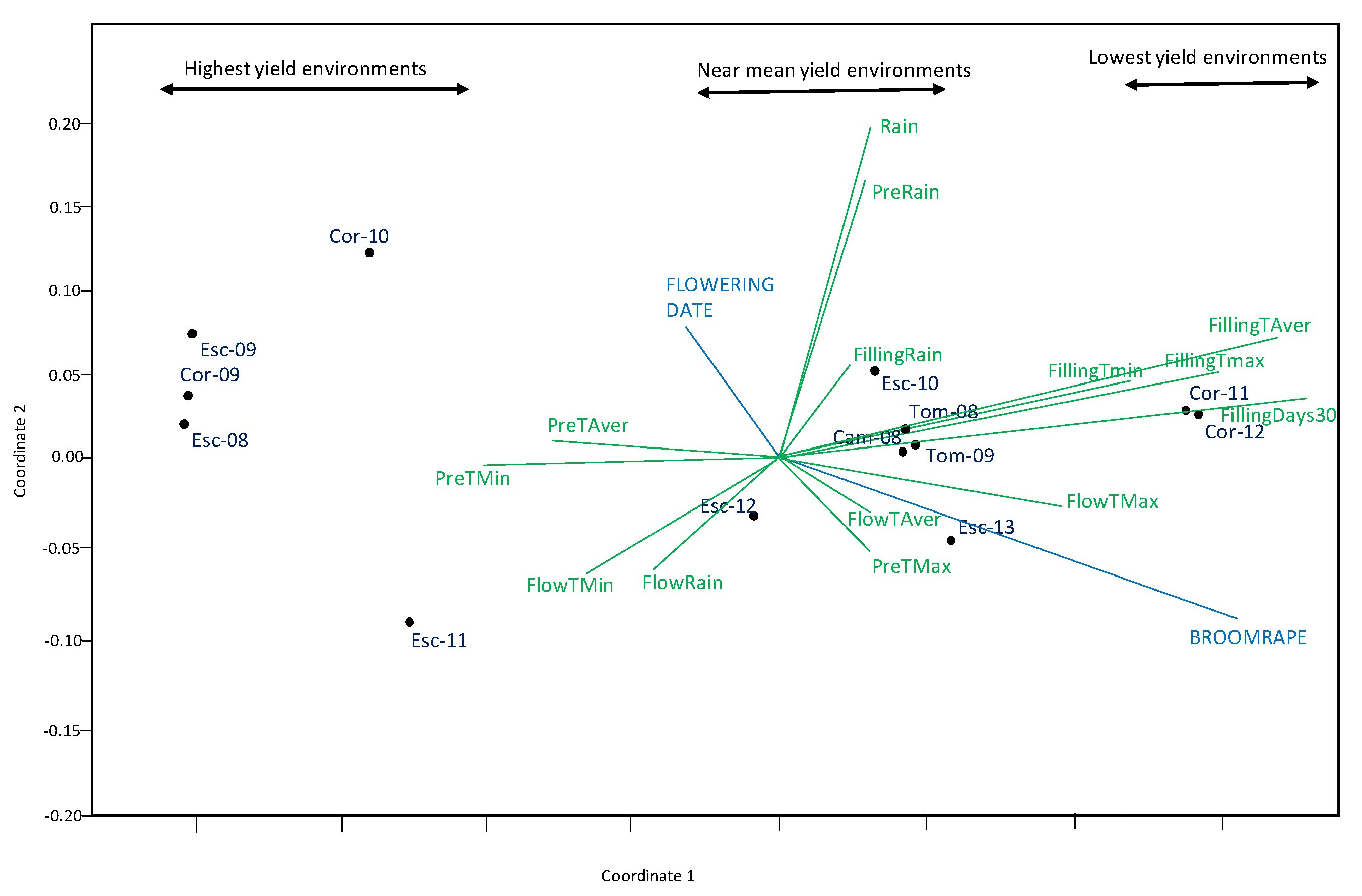
2.2.3. Non-Metric Multi-Dimensional Scaling Ordination (NMDS)
2.2.4. Heat Tolerance Indices
- Geometric mean productivity (GMP) = √(Ypi × Ysi) [18]
- Yield index (YI) = Ysi/Ys [19]
- Mean productivity (MP) = (Ypi + Ysi)/2 [20]
- Stress susceptibility index (SSI) = (1 − (Ysi/Ypi))/SI [21]
- Stress tolerance index (STI) = (Ypi × Ysi)/Y2 p [18]
- Harmonic mean (HARM) = 2 (Ypi × Ysi)/(Ypi + Ysi) [22]
- Heat tolerance index (HTI) = (Y2 si/Ypi) × (Yp/Y2 s) [23]
3. Results
4. Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT 2021. Available online: http://www.fao.org/faostat (accessed on 4 July 2021).
- Wright, D.M.; Neupane, S.; Heidecker, T.; Haile, T.A.; Chan, C.; Coyne, C.J.; McGee, R.J.; Udupa, S.; Henkrar, F.; Barilli, E.; et al. Understanding photothermal interactions will help expand production range and increase genetic diversity of lentil (Lens culinaris Medik.). Plants People Planet 2021, 3, 171–181. [Google Scholar] [CrossRef]
- Erskine, W.; Tufail, M.; Russell, A.; Tyagi, M.C.; Rahman, M.M.; Saxena, M.C. Current and future strategies in breeding lentil for resistance to biotic and abiotic stresses. Euphytica 1994, 73, 127–135. [Google Scholar] [CrossRef]
- Ali, A.; Johnson, D.L.; Stushnoff, C. Screening lentil (Lens culinaris) for cold hardiness under controlled conditions. J. Agric. Sci. 1999, 133, 313–319. [Google Scholar] [CrossRef]
- Muehlbauer, F.J.; Cho, S.; Sarker, A.; McPhee, K.E.; Coyne, C.J.; Rajesh, P.N.; Rebecca, R. Application of biotechnology in breeding lentil for resistance to biotic and abiotic stress. Euphytica 2006, 147, 149–165. [Google Scholar] [CrossRef]
- Sellami, M.H.; Pulvento, C.; Lavini, A. Selection of Suitable Genotypes of Lentil (Lens culinaris Medik.) under Rainfed Conditions in South Italy Using Multi-Trait Stability Index (MTSI). Agronomy 2021, 11, 1807. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Haddar, A. Parasitic plants. In The Lentil: Botany, Production and Uses; Erskine, W., Muehlbauer, F.J., Sarker, A., Sharma, B., Eds.; CABI: Wallingfore, UK, 2009; pp. 343–349. ISBN 978-1-84593-487-3. [Google Scholar]
- Beniwal, S.P.S.; Bayaa, B.; Weigand, S.; Makkouk, K.H.; Saxena, M.C. Field Guide for Lentil Diseases and Insect Pests; ICARDA: Aleppo, Syria, 1993. [Google Scholar]
- Bhandari, K.; Siddique, K.H.; Turner, N.C.; Kaur, J.; Singh, S.; Agrawal, S.K.; Nayyar, H. Heat stress at reproductive stage disrupts leaf carbohydrate metabolism, impairs reproductive function, and severely reduces seed yield in lentil. J. Crop Improv. 2016, 30, 118–151. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Effects of drought, heat and their interaction on the growth, yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity. Front. Plant Sci. 2017, 8, 1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.K.; Holland, J.B. A heritability-adjusted GGE biplot for test environment evaluation. Euphytica 2010, 171, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Fernández, T.; Carrillo-Perdomo, E.; Flores, F.; Rubiales, D. Identification and multi-environment validation of resistance to pea weevil (Bruchus pisorum) in Pisum germplasm. J. Pest Sci. 2018, 91, 205–214. [Google Scholar] [CrossRef]
- Rubiales, D.; Emeran, A.A.; Flores, F. Adaptation of grass pea (Lathyrus sativus) to Mediterranean environments. Agronomy 2020, 10, 1295. [Google Scholar] [CrossRef]
- Burgueño, J.; Crossa, J.; Vargas, M. SAS Programs for Graphing GE and GGE Biplots; CIMMYT: El Batan, Mexico, 2003. [Google Scholar]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Red de Información Agroclimática de Andalucía (RIA). Available online: https://www.juntadeandalucia.es/agriculturaypesca/ifapa/ria/servlet/FrontController (accessed on 7 August 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Fernandez, G.C. Effective Selection Criteria for Assessing Plant Stress Tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar]
- Gavuzzi, P.; Rizz, F.; Palumbo, M.; Campaline, R.G.; Ricciardi, G.L.; Borghi, B. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci. 1997, 77, 523–531. [Google Scholar] [CrossRef]
- Rosielle, A.A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environment. Crop Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars, I. Grain yield response. Austr. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Schneider, K.A.; Rosales-Serna, R.; Ibarra-Perez, F.; Cazares-Enriquez, B.; Acosta-Gallegos, J.A.; Ramirez-Vallejo, P.; Wassimi, N.; Kelly, J.D. Improving common bean performance under drought stress. Crop Sci. 1997, 37, 43–50. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Z.R.; Li, D.; Wei, J.W.; Qiao, W.C.; Meng, X.H.; Sun, S.L.; Li, H.M.; Zhao, M.H.; Chen, X.M.; et al. Evaluation of a new method for quantification of heat tolerance in different wheat cultivars. J. Integr. Agric. 2018, 17, 786–795. [Google Scholar] [CrossRef] [Green Version]
- Poarch, T.G. Application of stress indices for heat tolerance screening of common bean. J. Agron. Crop Sci. 2006, 192, 390–394. [Google Scholar] [CrossRef]
- Ouji, A.; Sayar, R.; Mouelhi, M.; Amri, M.; Bouhadida, M.; Omri, N.; Abbes, Z.; Ben Salah, H.; Halila, M.H.; Kumar, S.; et al. Advances in lentil (Lens culinaris L.) breeding in Tunisia. In Proceedings of the International Conference on Pulses, Marrakesh, Morocco, 18–20 April 2016; p. 76. [Google Scholar]
- MAPA. 2021. Available online: https://www.mapa.gob.es/en/ (accessed on 13 June 2021).
- Yan, W.; Rajcan, I. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef]
- Barghi, S.S.; Mostafaii, H.; Peighami, F.; Zakaria, R.A. Path analysis of yield and its components in lentil under end season heat condition. Int. J. Agric. Res. 2012, 2, 969–974. [Google Scholar]
- El Haddad, N.; Rajendran, K.; Smouni, A.; Es-Safi, N.E.; Benbrahim, N.; Mentag, R.; Nayyar, H.; Maalouf, F.; Kumar, S. Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress. Agronomy 2020, 10, 1036. [Google Scholar] [CrossRef]
- Choukri, H.; Hejjaoui, K.; El-Baouchi, A.; El Haddad, N.; Smouni, A.; Maalouf, F.; Thavarajah, D.; Kumar, S. Heat and drought stress impact on phenology, grain yield, and nutritional quality of lentil (Lens culinaris Medikus). Front. Nutr. 2020, 7, 596307. [Google Scholar] [CrossRef]
- Delahunty, A.; Nuttall, J.; Nicolas, M.; Brand, J. Response of lentil to high temperature under variable water supply and carbon dioxide enrichment. Crop Pasture Sci. 2018, 69, 1103–1112. [Google Scholar] [CrossRef]
- Kumar, J.; Sen Gupta, D.; Djalovic, I. Breeding, genetics, and genomics for tolerance against terminal heat in lentil: Current status and future directions. Legume Sci. 2020, 2, e38. [Google Scholar] [CrossRef] [Green Version]
- Lake, L.; Kutchartt, D.G.; Calderini, D.F.; Sadras, V.O. Critical developmental period for grain yield and grain protein concentration in lentil. Field Crops Res. 2021, 270, 108203. [Google Scholar] [CrossRef]
- Choudhury, D.R.; Tarafdar, S.; Das, M.; Kundagrami, S. Screening lentil (Lens culinaris Medik.) germplasms for heat tolerance. Trends Biosci. 2012, 5, 143–146. [Google Scholar]
- Kumar, J.; Kant, R.; Kumar, S.; Basu, P.S.; Sarker, A.; Singh, N.P. Heat tolerance in lentil under field conditions. Legume Genom. Genet. 2016, 7, 1–11. [Google Scholar]
- Chakherchaman, A.; Mostafaei, S.H.; Imanparast, L.; Eivazian, M.R. Evaluation of drought tolerance in lentil advanced genotypes in Ardabil region, Iran. J. Food Agric. Environ. 2009, 7, 283–288. [Google Scholar]
- Siahsar, B.A.; Ganjali, S.; Allahdoo, M. Evaluation of drought tolerance indices and their relationship with grain yield of lentil lines in drought-stressed and irrigated environments. Austr. J. Basic Appl. Sci. 2010, 4, 4336–4346. [Google Scholar]
- Singh, D.; Singh, C.K.; Singh, T.; Pal, M. Genetics and molecular mapping of heat tolerance for seedling survival and pod set in lentil. Crop Sci. 2017, 57, 3059–3067. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Identification of high-temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Front. Plant Sci. 2017, 8, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, J.; Basu, P.S.; Gupta, S.; Dubey, S.; Gupta, D.S.; Singh, N.P. Physiological and molecular characterisation for high temperature stress in Lens culinaris. Funct. Plant Biol. 2018, 45, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Silim, S.N.; Saxena, M.C.; Erskine, W. Effect of sowing date on the growth and yield of lentil in a rainfed Mediterranean environment. Expl. Agric. 1991, 27, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Alcántara, C.; Pérez-de-Luque, A.; Gil, J.; Sillero, J.C. Infection of chickpea (Cicer arietinum) by crenate broomrape (Orobanche crenata) as influenced by sowing date and weather conditions. Agronomie 2003, 23, 359–362. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Sillero, J.C.; Moral, A.; Cubero, J.I.; Rubiales, D. Effect of sowing date and host resistance on the establishment of Orobanche crenata in faba bean and common vetch. Weed Res. 2004, 44, 282–288. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Flores, F.; Rubiales, D. Differences in crenate broomrape parasitism dynamics on three legume crops using a Thermal Time Model. Front. Plant Sci. 2016, 7, 1910. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in parasitic weeds management in legume crops. A review. Agron. Sustain. Devel. 2012, 32, 433–449. [Google Scholar] [CrossRef]
- Grenz, J.H.; Manschadi, A.M.; Uygur, F.N.; Sauerborn, J. Effects of environment and sowing date on the competition between faba bean (Vicia faba) and the parasitic weed Orobanche crenata. Field Crop Res. 2005, 93, 300–313. [Google Scholar] [CrossRef]
- Rubiales, D.; Barilli, E.; Flores, F. Broomrape (Orobanche crenata) as a major constraint for grass pea (Lathyrus sativus) production in Mediterranean rain-fed environments. Agronomy 2020, 10, 1931. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Sillero, J.C.; Pérez-de-Luque, A.; Rubiales, D. Identification of sources of resistance to crenate broomrape (Orobanche crenata) in Spanish lentil (Lens culinaris) germplasm. Weed Res. 2008, 48, 85–94. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Origin/Derived from Accession no. |
|---|---|
| R4 | ILL5755 |
| R5 | ILL6002 |
| R7 | ILL6258 |
| R14 | ILL7517 |
| R17 | ILL8707 |
| R19 | ILL9903 |
| S6 | ILL10074 |
| S7 | ILL10079 |
| S8 | ILL10170 |
| S12 | ILL10174 |
| S14 | ILL10273 |
| S17 | ILL10278 |
| S23 | ILL10648 |
| S24 | ILL10653 |
| Armuña | Spanish landrace |
| Nefza | Tunisian cv. |
| Nsir | Tunisian cv. |
| Environment | Season | Site, Level of Broomrape (Oc) Infestation | Soil type | Soil pH | Latit. | Longit. | Altit. | Average Tmax (°C) | Average Tmin (°C) | Rain (mm) |
|---|---|---|---|---|---|---|---|---|---|---|
| Cam-08 | 2007–2008 | Campillo: low Oc | Vertisol | 7.5–8 | 37°20′ N | 4°51′ W | 461 | 18.0 | 6.8 | 264 |
| Cor-09 | 2008–2009 | Córdoba: high Oc | Cambisol | 6.5–7 | 37°50′ N | 4°50′ W | 90 | 19.2 | 6.9 | 279 |
| Cor-10 | 2009–2010 | Córdoba: high Oc | Cambisol | 6.5–7 | 37°50′ N | 4°50′ W | 90 | 20.1 | 8.4 | 1053 |
| Cor-11 | 2010–2011 | Córdoba: high Oc | Cambisol | 6.5–7 | 37°50′ N | 4°50′ W | 90 | 22.5 | 10.3 | 513 |
| Cor-12 | 2011–2012 | Córdoba: high Oc | Cambisol | 6.5–7 | 37°50′ N | 4°50′ W | 90 | 20.8 | 5.9 | 163 |
| Esc-08 | 2007–2008 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 20.1 | 9.3 | 391 |
| Esc-09 | 2008–2009 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 20.3 | 8.8 | 252 |
| Esc-10 | 2009–2010 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 19.2 | 9.4 | 1206 |
| Esc-11 | 2010–2011 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 21.6 | 11.4 | 534 |
| Esc-12 | 2011–2012 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 20.5 | 9.1 | 134 |
| Esc-13 | 2012–2013 | Escacena: high Oc | Fluvisol | 7–7.5 | 37°25′ N | 6°15′ W | 88 | 19.4 | 9.2 | 411 |
| Tom-08 | 2007–2008 | Tomejil: low Oc | Vertisol | 7–7.5 | 37°30′ N | 5°57′ W | 12 | 21.7 | 8.7 | 399 |
| Tom-09 | 2008–2009 | Tomejil: low Oc | Vertisol | 7–7.5 | 37°30′ N | 5°57′ W | 12 | 21.3 | 7.1 | 219 |
| Grain yield (kg/ha) | Random Effects | Estimate | Standard Error | Z value | Pr > Z |
| E | 45084 | 20177 | 2.23 | 0.0127 | |
| Block(E) | 4073 | 1915 | 2.13 | 0.0167 | |
| G × E | 34344 | 5238 | 6.56 | <0.0001 | |
| Residual | 47537 | 3296 | 14.42 | <0.0001 | |
| Fixed Effects | Numerator df | Denominator df | F value | Pr > F | |
| G | 16 | 192 | 5.3 | <0.0001 | |
| Broomrape infection (Oc/plant) | Random Effects | Estimate | Standard Error | Z value | Pr > Z |
| E | 0.0483 | 0.0247 | 1.95 | 0.0255 | |
| Block(E) | 0.0080 | 0.0035 | 2.28 | 0.0113 | |
| G×E | 0.0078 | 0.0032 | 2.39 | 0.0084 | |
| Residual | 0.0524 | 0.0041 | 12.65 | <0.0001 | |
| Fixed Effects | Numerator df | Denominator df | F value | Pr > F | |
| G | 16 | 144 | 5.18 | <0.0001 | |
| Dates to flowering (dtf) | Random Effects | Estimate | Standard Error | Z value | Pr > Z |
| E | 28.19 | 12.22 | 2.31 | 0.0105 | |
| Block(E) | 0 | . | . | . | |
| G × E | 23.57 | 3.05 | 7.73 | <0.0001 | |
| Residual | 18.17 | 1.22 | 14.87 | <0.0001 | |
| Fixed Effects | Numerator df | Denominator df | F value | Pr > F | |
| G | 16 | 192 | 30.63 | <0.0001 |
| Accession | Cam-08 | Cor-09 | Cor-10 | Cor-11 | Cor-12 | Esc-08 | Esc-09 | Esc-10 | Esc-11 | Esc-12 | Esc-13 | Tom-08 | Tom-09 | Mean | SE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nsir | 635 | 901 | 1854 | 370 | 437 | 1125 | 933 | 921 | 871 | 567 | 387 | 510 | 493 | 770 | 71 |
| S23 | 476 | 1450 | 358 | 139 | 150 | 966 | 1413 | 516 | 775 | 938 | 247 | 470 | 461 | 643 | 85 |
| S17 | 424 | 1215 | 684 | 163 | 170 | 1254 | 965 | 332 | 342 | 982 | 251 | 630 | 635 | 619 | 73 |
| S12 | 416 | 794 | 145 | 135 | 144 | 974 | 1174 | 139 | 829 | 1504 | 487 | 404 | 385 | 579 | 76 |
| R7 | 128 | 970 | 669 | 108 | 105 | 1204 | 577 | 174 | 847 | 602 | 264 | 625 | 606 | 529 | 66 |
| S6 | 342 | 748 | 227 | 89 | 93 | 892 | 1127 | 162 | 1189 | 429 | 429 | 406 | 388 | 502 | 68 |
| S14 | 345 | 800 | 325 | 105 | 84 | 888 | 917 | 269 | 480 | 306 | 596 | 318 | 318 | 442 | 55 |
| Armuña | 268 | 608 | 1311 | 325 | 225 | 807 | 640 | 261 | 171 | 276 | 119 | 283 | 200 | 423 | 54 |
| R17 | 293 | 395 | 420 | 74 | 85 | 594 | 516 | 166 | 396 | 978 | 836 | 303 | 261 | 409 | 53 |
| Nefza | 216 | 609 | 829 | 53 | 41 | 741 | 643 | 244 | 311 | 276 | 409 | 244 | 204 | 371 | 43 |
| S8 | 234 | 292 | 469 | 76 | 76 | 771 | 536 | 588 | 558 | 233 | 618 | 203 | 164 | 371 | 44 |
| S7 | 183 | 847 | 339 | 117 | 109 | 780 | 546 | 355 | 164 | 239 | 336 | 330 | 272 | 355 | 42 |
| S24 | 313 | 315 | 312 | 62 | 69 | 579 | 1017 | 202 | 591 | 169 | 429 | 163 | 146 | 336 | 50 |
| R5 | 157 | 549 | 589 | 108 | 72 | 733 | 581 | 257 | 191 | 211 | 220 | 209 | 142 | 309 | 36 |
| R14 | 174 | 557 | 120 | 73 | 66 | 806 | 265 | 166 | 725 | 250 | 342 | 218 | 141 | 300 | 48 |
| R4 | 162 | 454 | 541 | 68 | 98 | 553 | 291 | 108 | 575 | 231 | 689 | 237 | 135 | 319 | 37 |
| R19 | 146 | 559 | 404 | 87 | 95 | 533 | 444 | 240 | 389 | 208 | 356 | 174 | 163 | 292 | 37 |
| Mean | 289 | 710 | 564 | 127 | 125 | 835 | 740 | 300 | 553 | 494 | 412 | 337 | 301 | 445 | |
| SE | 20 | 50 | 67 | 22 | 23 | 31 | 51 | 47 | 67 | 59 | 44 | 21 | 28 | 15 |
| Accession | Cam-08 | Cor-09 | Cor-10 | Cor-11 | Cor-12 | Esc-08 | Esc-09 | Esc-10 | Esc-11 | Esc-12 | Esc-13 | Tom-08 | Tom-09 | Mean | SE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Armuña | 115 | 121 | 122 | 125 | 99 | 111 | 115 | 102 | 107 | 109 | 108 | 115 | 117 | 113 | 1.2 |
| Nsir | 105 | 110 | 106 | 118 | 96 | 102 | 105 | 94 | 103 | 95 | 100 | 107 | 107 | 104 | 1.1 |
| Nefza | 100 | 105 | 102 | 113 | 90 | 97 | 99 | 90 | 99 | 87 | 94 | 101 | 102 | 98 | 1.2 |
| S8 | 97 | 95 | 106 | 120 | 90 | 94 | 90 | 84 | 93 | 91 | 90 | 99 | 96 | 96 | 1.5 |
| S7 | 95 | 95 | 106 | 119 | 93 | 90 | 90 | 78 | 86 | 87 | 87 | 98 | 92 | 94 | 1.7 |
| R4 | 95 | 96 | 106 | 113 | 77 | 93 | 95 | 75 | 92 | 96 | 90 | 98 | 96 | 94 | 1.7 |
| R19 | 96 | 94 | 106 | 116 | 90 | 90 | 88 | 76 | 98 | 86 | 88 | 97 | 95 | 94 | 1.8 |
| R7 | 91 | 91 | 92 | 88 | 79 | 90 | 86 | 80 | 99 | 86 | 88 | 91 | 90 | 89 | 1.0 |
| S12 | 91 | 95 | 99 | 98 | 85 | 87 | 84 | 74 | 95 | 85 | 85 | 92 | 88 | 89 | 1.4 |
| S17 | 91 | 95 | 101 | 90 | 83 | 88 | 84 | 72 | 101 | 82 | 85 | 94 | 86 | 89 | 1.4 |
| S14 | 88 | 91 | 92 | 88 | 79 | 89 | 84 | 87 | 90 | 84 | 89 | 88 | 86 | 87 | 0.7 |
| S23 | 87 | 91 | 92 | 88 | 81 | 87 | 84 | 74 | 96 | 84 | 85 | 90 | 86 | 87 | 1.0 |
| S24 | 88 | 91 | 98 | 88 | 82 | 87 | 90 | 76 | 87 | 85 | 86 | 87 | 86 | 87 | 0.9 |
| R5 | 86 | 91 | 92 | 77 | 67 | 89 | 85 | 88 | 91 | 81 | 87 | 87 | 88 | 85 | 1.3 |
| S6 | 87 | 95 | 101 | 88 | 80 | 84 | 84 | 61 | 101 | 85 | 82 | 88 | 86 | 86 | 1.8 |
| R17 | 82 | 85 | 76 | 77 | 65 | 84 | 84 | 75 | 89 | 78 | 83 | 82 | 86 | 81 | 1.4 |
| R14 | 81 | 89 | 59 | 78 | 74 | 81 | 84 | 75 | 74 | 81 | 80 | 81 | 86 | 79 | 1.4 |
| Mean | 93 | 96 | 97 | 99 | 83 | 91 | 90 | 80 | 94 | 87 | 89 | 94 | 93 | 91 | |
| SE | 1.2 | 1.2 | 2.0 | 2.4 | 1.3 | 1.0 | 1.2 | 1.8 | 1.3 | 1.0 | 1.0 | 1.2 | 1.2 | 0.4 |
| Accession | Cam-08 | Cor-09 | Cor-10 | Cor-11 | Cor-12 | Esc-08 | Esc-09 | Esc-10 | Esc-11 | Esc-12 | Esc-13 | Tom-08 | Tom-09 | Mean | SE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Armuña | 0.00 | 0.35 | 0.33 | 1.03 | 0.72 | 0.41 | 0.96 | 0.41 | 0.67 | 0.67 | 0.62 | 0.00 | 0.00 | 0.49 | 0.10 |
| Nsir | 0.00 | 0.20 | 0.07 | 0.99 | 0.65 | 0.19 | 0.81 | 0.53 | 0.13 | 0.41 | 0.43 | 0.00 | 0.00 | 0.34 | 0.10 |
| S7 | 0.00 | 0.11 | 0.13 | 1.07 | 0.73 | 0.20 | 0.08 | 0.62 | 0.63 | 0.47 | 0.42 | 0.00 | 0.00 | 0.34 | 0.08 |
| R5 | 0.00 | 0.26 | 0.10 | 1.01 | 0.52 | 0.20 | 0.87 | 0.06 | 0.47 | 0.47 | 0.51 | 0.00 | 0.00 | 0.34 | 0.08 |
| S8 | 0.00 | 0.21 | 0.23 | 1.02 | 0.68 | 0.07 | 0.11 | 0.26 | 0.80 | 0.43 | 0.27 | 0.00 | 0.00 | 0.31 | 0.08 |
| Nefza | 0.00 | 0.06 | 0.07 | 0.94 | 0.28 | 0.06 | 0.67 | 0.11 | 0.17 | 0.32 | 0.43 | 0.00 | 0.00 | 0.25 | 0.07 |
| S6 | 0.00 | 0.18 | 0.23 | 1.24 | 0.37 | 0.04 | 0.00 | 0.33 | 0.17 | 0.22 | 0.44 | 0.00 | 0.00 | 0.25 | 0.08 |
| S12 | 0.00 | 0.11 | 0.10 | 0.90 | 0.46 | 0.03 | 0.08 | 0.37 | 0.43 | 0.36 | 0.46 | 0.00 | 0.00 | 0.25 | 0.07 |
| S24 | 0.00 | 0.15 | 0.13 | 0.73 | 0.77 | 0.04 | 0.05 | 0.41 | 0.33 | 0.25 | 0.18 | 0.00 | 0.00 | 0.24 | 0.07 |
| S17 | 0.00 | 0.08 | 0.03 | 1.24 | 0.33 | 0.02 | 0.19 | 0.11 | 0.23 | 0.23 | 0.52 | 0.00 | 0.00 | 0.23 | 0.09 |
| R19 | 0.00 | 0.09 | 0.20 | 0.97 | 0.39 | 0.05 | 0.39 | 0.13 | 0.37 | 0.24 | 0.22 | 0.00 | 0.00 | 0.23 | 0.07 |
| R14 | 0.00 | 0.10 | 0.03 | 0.56 | 0.26 | 0.06 | 0.08 | 0.11 | 0.47 | 0.28 | 0.56 | 0.00 | 0.00 | 0.21 | 0.06 |
| S23 | 0.00 | 0.09 | 0.13 | 0.88 | 0.33 | 0.08 | 0.01 | 0.22 | 0.27 | 0.19 | 0.35 | 0.00 | 0.00 | 0.20 | 0.06 |
| R4 | 0.00 | 0.03 | 0.03 | 0.63 | 0.53 | 0.03 | 0.36 | 0.09 | 0.20 | 0.26 | 0.37 | 0.00 | 0.00 | 0.19 | 0.06 |
| R7 | 0.00 | 0.00 | 0.03 | 0.59 | 0.24 | 0.05 | 0.69 | 0.20 | 0.17 | 0.23 | 0.10 | 0.00 | 0.00 | 0.18 | 0.09 |
| R17 | 0.00 | 0.12 | 0.00 | 0.59 | 0.48 | 0.06 | 0.16 | 0.11 | 0.07 | 0.15 | 0.28 | 0.00 | 0.00 | 0.15 | 0.06 |
| S14 | 0.00 | 0.06 | 0.03 | 0.41 | 0.30 | 0.01 | 0.17 | 0.06 | 0.20 | 0.12 | 0.28 | 0.00 | 0.00 | 0.13 | 0.03 |
| Mean | 0.00 | 0.13 | 0.11 | 0.87 | 0.47 | 0.09 | 0.33 | 0.24 | 0.34 | 0.31 | 0.38 | 0.00 | 0.00 | 0.26 | |
| SE | 0.00 | 0.02 | 0.02 | 0.04 | 0.06 | 0.01 | 0.06 | 0.04 | 0.04 | 0.02 | 0.04 | 0.00 | 0.00 | 0.02 |
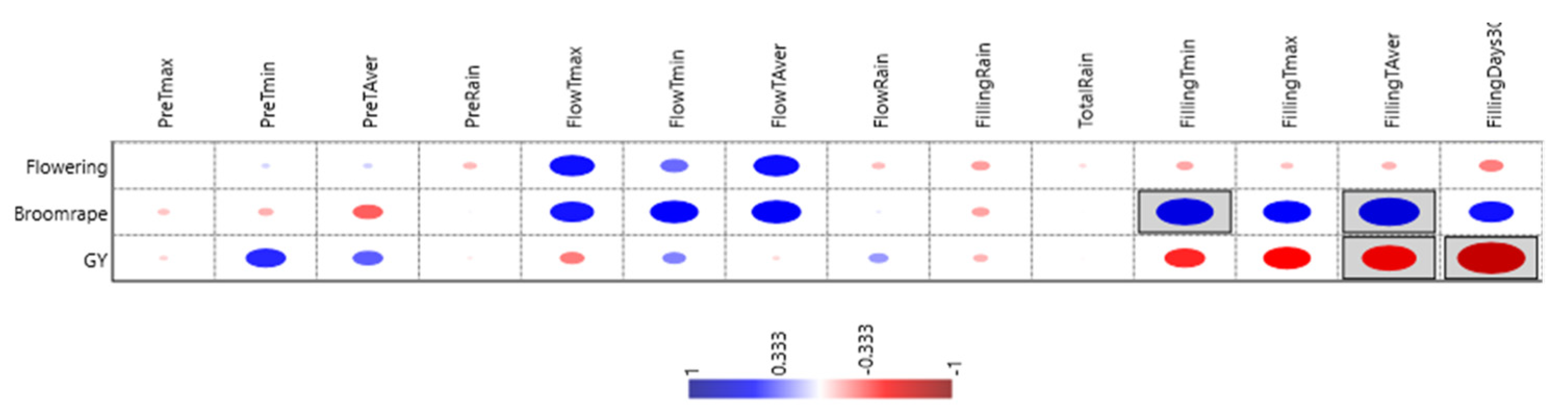
| Grain Yield | Dtf | Broomrape | Rain | Tmin Preflowering | TAver Filling | Numb. Days Tmax > 30 °C | |
|---|---|---|---|---|---|---|---|
| Grain yield | 0.19 | −0.36 | 0.02 | 0.42 | −0.58 * | −0.72 ** | |
| Dtf | −0.07 | −0.06 | 0.08 | −0.15 | −0.25 | ||
| Broomrape | 0.01 | −0.15 | 0.64 * | 0.46 |
| Environments | GY Mean | Broomrape Mean | Tmin at Pre-Flowering | Taver at Flowering | Tmax at Grain Filling | Num. Days Tmax > 30 °C |
|---|---|---|---|---|---|---|
| Cam-08 | 289 | 0.00 | 4.8 | 9.8 | 20.6 | 0 |
| Cor-09 | 710 | 0.13 | 4.1 | 8.8 | 25.5 | 2 |
| Cor-10 | 564 | 0.11 | 5.4 | 10.1 | 24.1 | 2 |
| Cor-11 | 127 | 0.87 | 5.6 | 10.0 | 28.9 | 8 |
| Cor-12 | 125 | 0.47 | 1.2 | 7.7 | 29.1 | 9 |
| Esc-08 | 835 | 0.09 | 8.0 | 12.6 | 22.2 | 0 |
| Esc-09 | 740 | 0.33 | 6.2 | 10.3 | 24.8 | 2 |
| Esc-10 | 300 | 0.24 | 7.3 | 10.9 | 27.9 | 7 |
| Esc-11 | 553 | 0.34 | 7.3 | 11.1 | 26.1 | 3 |
| Esc-12 | 494 | 0.31 | 5.2 | 10.5 | 21.7 | 2 |
| Esc-13 | 412 | 0.42 | 6.7 | 10.6 | 24.3 | 2 |
| Tom-08 | 337 | 0.00 | 6.8 | 12.8 | 24.2 | 3 |
| Tom-09 | 301 | 0.00 | 4.9 | 10.2 | 28.1 | 8 |
| Accession | Ys | Yp | GMP | YI | MP | SSI | STI | HARM | HTI |
|---|---|---|---|---|---|---|---|---|---|
| S17 | 635 | 1254 | 892 | 2.11 | 944 | 0.79 | 1.25 | 843 | 2.86 |
| R7 | 606 | 1204 | 854 | 2.01 | 905 | 0.80 | 1.14 | 806 | 2.71 |
| Nsir | 493 | 1125 | 745 | 1.64 | 809 | 0.90 | 0.87 | 686 | 1.92 |
| S23 | 461 | 966 | 668 | 1.53 | 714 | 0.84 | 0.70 | 625 | 1.96 |
| S12 | 385 | 974 | 613 | 1.28 | 680 | 0.97 | 0.59 | 552 | 1.36 |
| S6 | 388 | 892 | 589 | 1.29 | 640 | 0.91 | 0.54 | 541 | 1.51 |
| S14 | 318 | 888 | 531 | 1.06 | 603 | 1.03 | 0.44 | 468 | 1.01 |
| S7 | 272 | 780 | 461 | 0.90 | 526 | 1.05 | 0.33 | 403 | 0.84 |
| Armuña | 200 | 807 | 402 | 0.66 | 504 | 1.20 | 0.25 | 321 | 0.44 |
| R17 | 261 | 594 | 394 | 0.87 | 428 | 0.90 | 0.24 | 363 | 0.89 |
| Nefza | 204 | 741 | 389 | 0.68 | 473 | 1.16 | 0.24 | 320 | 0.50 |
| S8 | 164 | 771 | 355 | 0.54 | 467 | 1.26 | 0.20 | 270 | 0.31 |
| R14 | 141 | 806 | 337 | 0.47 | 474 | 1.32 | 0.18 | 240 | 0.22 |
| R5 | 142 | 733 | 323 | 0.47 | 438 | 1.29 | 0.16 | 238 | 0.25 |
| R19 | 163 | 533 | 295 | 0.54 | 348 | 1.11 | 0.14 | 250 | 0.44 |
| S24 | 146 | 579 | 290 | 0.48 | 362 | 1.19 | 0.13 | 233 | 0.33 |
| R4 | 135 | 553 | 273 | 0.44 | 344 | 1.21 | 0.11 | 217 | 0.29 |
| Ys | Yp | HARM | MP | SSI | STI | YI | HTI | |
|---|---|---|---|---|---|---|---|---|
| Yp | 0.83 *** | |||||||
| HARM | 0.99 *** | 0.88 *** | ||||||
| MP | 0.93 *** | 0.98 *** | 0.96 *** | |||||
| SSI | −0.49 * | 0.05 | −0.42 | −0.15 | ||||
| STI | 0.98 *** | 0.87 *** | 0.99 *** | 0.95 *** | −0.41 | |||
| YI | 0.98 *** | 0.83 *** | 0.99 *** | 0.93 *** | −0.50 * | 0.99 *** | ||
| HTI | 0.97 *** | 0.70 ** | 0.95 *** | 0.84 *** | −0.65 ** | 0.95 *** | 0.98 *** | |
| GMP | 0.98 *** | 0.93 *** | 0.99 *** | 0.99 *** | −0.31 | 0.98 *** | 0.98 *** | 0.91 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubiales, D.; Moral, A.; Flores, F. Heat Waves and Broomrape Are the Major Constraints for Lentil Cultivation in Southern Spain. Agronomy 2021, 11, 1871. https://doi.org/10.3390/agronomy11091871
Rubiales D, Moral A, Flores F. Heat Waves and Broomrape Are the Major Constraints for Lentil Cultivation in Southern Spain. Agronomy. 2021; 11(9):1871. https://doi.org/10.3390/agronomy11091871
Chicago/Turabian StyleRubiales, Diego, Ana Moral, and Fernando Flores. 2021. "Heat Waves and Broomrape Are the Major Constraints for Lentil Cultivation in Southern Spain" Agronomy 11, no. 9: 1871. https://doi.org/10.3390/agronomy11091871
APA StyleRubiales, D., Moral, A., & Flores, F. (2021). Heat Waves and Broomrape Are the Major Constraints for Lentil Cultivation in Southern Spain. Agronomy, 11(9), 1871. https://doi.org/10.3390/agronomy11091871