Abstract
Watermelon is one of the most desirable vegetable crops in the world. Recently, grafting is common in watermelons worldwide, but not all grafting methods are compatible with polyploids. In this study, diploid, triploid, and tetraploid from one watermelon variety, “Mi Mei”, were grafted on the “Xijiaqiangsheng” squash rootstock to study the effect of genome duplication on graft compatibility. Three grafting methods (splice, hole, and tongue) were used to determine graft compatibility. Significant differences in survival rates, hormones, antioxidants (AOX), sugars, and starch contents were observed between compatible/incompatible combinations. Compatible combinations with high survival rates showed high levels of hormones, AOX, carbohydrates, and low hydrogen peroxide H2O2 compared to incompatible plants. The hole grafting method was more efficient with diploid, while splice was efficient with a tetraploid, and both methods can be used for triploid. Compatibility is a combined effect of hormone, carbohydrate, and antioxidant activities. We predict that compatibility is a complex process and that further molecular studies must be performed to dig deep into this phenomenon.
1. Introduction
Seedless watermelon cultivars (Citrullus lanatus L.) are the most desirable by consumers and have a high price and more excellent quality than seeded watermelon [1]. Seedless watermelons are triploids (3n = 33) produced by crossing a tetraploid (4n = 44) seed parent with a diploid (2n = 22) pollen parent [2,3]. Tetraploid induction can be done by different methods, like applying aqueous chemicals (Colchicine and Oryzalin) solution to the apical meristem of diploid seedlings [4,5,6].
Polyploidy is the heritable condition of possessing more than two complete sets of chromosomes, and very common among plants, and are usually fit and well adapted [7]. One of the main advantages of the polyploidy in the watermelon is triploid (seedless) fruits; another benefit, conferred by gene redundancy, is diversifying gene function by altering redundant copies of important or essential genes [7,8,9]. But generally, after genome doubling, the artificially induced autopolyploids always exhibit new characters, such as DNA contents, high secondary metabolite organisms, large tissues and organs, improved yield, higher contents of chlorophyll, lycopene, fructose, and glucose, and higher tolerance to both abiotic and biotic stress, than diploid, all of which are distinct advantages of polyploidy [2,3,5,10,11,12,13].
Recently, grafting has been used to enhance the resistance to biotic and abiotic stresses and improve yields [14,15]. Graft compatibility is a complex process of anatomical and biochemical interactions [16,17] that starts with wound response, then callus formation, and finished with cambium and functional vascular system between rootstock and scion creation [18]. All of these steps can determine the future of a grafted plant [19]. According to (Melnyk et al., 2017), who explained the graft junction formation steps at the first 1–2 days after grafting: (1) adhesion of scion and stock (2) cell divisions initiate and callus formation (3) callus makes contact between scion and stock. A week or more after grafting: (4) callus differentiate to xylem and phloem elements, (5) vascular connections are re-established to [20].
Several watermelon grafting methods have been reported [21,22]. The tongue (T), hole (H), and splice (S) are the most used methods in vegetable grafting, T is one of the initial techniques that are performed, and unpracticed producers prefer it, due to its simplicity, high success rate, and that it does not require much care or healing chambers [21,23,24]. Experienced watermelon growers prefer the H method, due to the limited space needed and the workers’ speedy process. [23]; on the other hand, it requires expert workers and more time than other methods and also needs careful environmental conditions control (light, humidity, temperature) after grafting [21,25]. Even though the S method is relatively simple, it is unpopular, due to the lower survival rate if the healing environment is not perfect [25,26]. However, grafting with the splice method gives a faster, stronger, and larger union region area than other methods [17,27,28].
IAA, and cytokinin, play an essential role in regulating scion–stock interactions [29] and are required for wound healing and vascular regeneration in the graft region area [30,31]. Graft incompatibility in the fruit trees occurs, due to hormone imbalance [32,33,34,35]. Auxin is a plant hormone that causes the elongation of cells in shoots and regulates plant growth. Low concentrations of auxin promote intact roots’ growth, whereas higher concentrations inhibit growth [36,37]. The growth rate of maize roots is more sensitive to changes in IAA concentration [38]. Cytokinin in tobacco increases root growth, primarily by increasing the size of the root apical meristem. Similarly, mutations that partially disrupt cytokinin perception also cause enhanced root growth and accelerate the process of vascular differentiation in the root tip [37].
Reactive Oxygen Species (ROS) accumulation leads to oxidative damage and cell death [11,39,40]. Increasing activity of defense-related enzymes, such as Catalase (CAT) and Peroxidase (POD), can scavenge ROS in plants and improve the resistance of plants to stress [41,42]. POX and CAT convert H2O2 to H2O [43]. The formation of graft union has coincided with antioxidants (AOX) activities in dealing with oxidative stress [44,45]. In comparison, incompatible grafts showed higher H2O2 levels and lower AOX activities in the grafting region [44,46,47].
Furthermore, carbohydrates play an essential role in plants’ cellular activities by providing energy [48,49]. Graft union development is due to the content of carbohydrates present in scion and rootstock at grafting time, higher carbohydrates levels, and auxins and cytokinin, essential for effective callus formation. Interactions among rootstock and scion cultivars for utilizing carbohydrates can influence starch depletion, which occurs throughout callus development; the survival rate increases by increasing starch content [49,50].
Rootstock-scion compatibility depends on anatomical, physiological, and genetic variables. Oda et al. (1993) Suggested that the reduction in the difference in diameter of the hypocotyls between the scion and rootstock was effective in increasing the survival rate and promoting cucumber plants’ growth, while the number of vascular bundles did not affect the survival rate [51]. Traka-Mavrona et al. (2000) [52] also showed that the differences in stem diameter between Cucurbita and Cucumis reduced grafts’ survival rate.
This research studied the effect of genome doubling on graft compatibility at physiological and anatomical levels, which maybe led to a good explanation of the compatibility mechanisms. It will also open the door for breeders to produce tetraploid rootstocks, which have high survival rates and high resistance to biotic and abiotic stress.
2. Materials and Methods
2.1. Plant Materials
Seedless watermelons are triploids (3n = 33) produced by crossing a tetraploid seed parent with a diploid (2n = 22) pollen parent. Tetraploid induction was done by applying colchicine to the growing apex of diploid seedlings. Polyploid seeds for one variety (mimei), which is homozygous and genetically stable, and passed the achievement appraisal of the Chinese Department of Agriculture in 1990 and won the second prize of science technology progress of the Department of Agriculture in 1991 [53], were used as scions and squash as rootstock, which is an interspecific hybrid (between Cucurbita moschata × Cucurbita maxima) (xijiaqiangsheng) and widely used in China as a rootstock, obtained from the polyploid watermelon group—Zhengzhou Fruit research institute (CAAS) China.
Plants were cultivated in a glasshouse with the growth conditions were set to 25–30 °C with 60–85% humidity with ordinary light. The sensor in each experimental plot’s center was used to record temperature and humidity (THtool-V151_En; Campbell Scientific Ltd., Beijing, China). We closed the healing room for seven days. After that, we start to open gradually until 12 days after grafting, and after 15 days, we collected survival rates data and samples.
2.2. Methods
2.2.1. Grafting Methods
This study used the three most commercial methods used in watermelon grafting, namely, tongue, splice, and hole methods (Figure 1).

Figure 1.
Three grafting methods were used for watermelon grafting, (A) tongue grafting method (T), (B) splice grafting method (S), and (C) hole grafting method (H).
2.2.2. Survival Rates
Survival rates were investigated after 15 days using the formula [54].
Survival rates were measured after 15 days from the grafting by counting the successful seedlings and dividing them by the total number of the grafting seedlings. The individual plants were counted from three biological replicates; each replicate contains 30 plants from the splice and hole grafting methods (which had significant differences between polyploids).
Samples were collected at three different stages (0, 3, and 15 days) after grafting for the determinations and histological studies. Samples were taken at 0 DAG as a control to study the grafting process development. At 3 DAG is the critical time for grafting, as it has the highest rate of cellular activity, which is determined based on compatibility/incompatibility in the grafting. At 15 DAG, samples were taken after survival and success compatibility. The most critical time in the healing process is 2–3 DAG during the healing stage [55], there is the highest rate of stress (complete dark, high humidity, and temperature), and the highest increase of hormones and antioxidants leads to graft compatibility [56].
2.2.3. Estimate Hormone Contents by ELISA Method
Three biological replicates were collected from the graft union at 0, 3, and 15 days after grafting (DAG). Indole-3-acetic acid (IAA) and Zeatin Riboside (ZR) contents were measured with the Enzyme-Linked Immunosorbent Assay (ELISA) method (China Agricultural University). The determination of hormonal contents was performed as outlined by [29].
2.2.4. Assay for Antioxidants Enzymes Activity and H2O2 Contents
Superoxide dismutase (SOD) activity in the supernatant was assayed for its ability to inhibit the photochemical reduction of nitro blue tetrazolium (NBT) by reading the absorbance at 560 nm wavelength using SOD assay kit/ YX-C-A500 for the measurement of SOD activity. Catalase activity was determined by measuring the rate of change of H2O2 absorbance in 60 s at 240 nm, using a spectrophotometer by CAT assay kit/BC0200.
Peroxidase (POD) can catalyze the oxidation of phenols and amines by H2O2, which has the dual function of eliminating the toxicity of H2O2, phenols, and amines. POD catalyzes the oxidation of specific substrates by H2O2 and has characteristic light absorption at 470 nm using POD assay kit/ YX-C-A502. H2O2 forms a yellow titanium peroxide complex with titanium sulfate, with characteristic absorption at 415 nm wavelength using H2O2 assay kit/ YX-C-A400 (Sino best biological technology co., Beijing, China), according to the manufacturer’s instructions. Three biological replications from grafting union at 0, 3, and 15 DAG from three different plants for each replicate were collected for analysis.
2.2.5. Quantification of Carbohydrates, Sugars, and Starch
All experiments were carried out using starch assay kit/ YX-C-C400 for starch content estimation at 620 nm wavelength, sugars assay kit/YX-C-B629 for sugars content using 540 nm wavelength (Sino best biological technology co., Beijing, China), according to the manufacturer’s instructions. Three biological replications from grafting union at 0, 3, and 15 DAG from three different plants for each replicate were collected for analysis.
2.2.6. Rootstocks Roots Architecture and Development Measurements (Scanning of Rootstock Roots Morphology)
At different five stages (0, 3, 5, 7, and 15 DAG), roots from five grafting combination plants were taken, washed twice with water. The root morphology was carried by a scanner (i800 plus EZ V2.61p, Selangor, Malaysia) the image analysis system (Wseen LA-S) was used to analyze the root morphology parameters.
2.2.7. Anatomical Observation
Five samples (graft union region) were collected from each replicate in Diploid, Triploid, and Tetraploid watermelons after 15 DAG, which grafted with splice and hole grafting method and fixed in FAA solution for 48 h then dehydrated in an ethanol series (30, 50, 70, and 95%), each dehydrated step for 90 min, then softened in glycerol: alcohol mixture (1:1) about one week. Samples were embedded in paraffin wax (52–54 °C), then a ten μm thickness section was sectioned on a sliding microtome [17]. Finally, paraffin sections were examined with an epifluorescence microscope (Leica DM-LS, Leica Microsystems, Wetzlar, Germany). Images were captured with HPCCD-5 Color Microscope Camera with ScopePhoto, V3.1 software.
2.3. Statistical Analysis
The present experiment was designed based on a completely randomized design. The general linear model by two-way ANOVA, 2 × 3 were applied to analyze the data using SPSS 26 (IBM).
The two-way ANOVA model included the main effects of species (diploid or triploid or tetraploid), the grafting method (Splice or Hole), and interactions between species and grafting methods. Data are shown as the means of three replicates. Correlation between the characters was determined at 95%. Hormones, AOX, sugars, and starch contents in the graft union were subjected to Principal Component Analysis (PCA) to verify the interaction between different grafting methods among polyploid watermelons synthesize the considered variables using Excel (Microsoft Corporation, Redmond, WA, USA).
3. Results
3.1. Survival Rates of Di, Tri, and Tetraploid Watermelon Grafted by Different Grafting Methods
Plant Survive rate was measured at 15 DAG in diploid (Di), triploid (Tri), and tetraploid (Tetra) watermelons, which were grafted by using Tongue (T), Splice (S), and Hole (H) grafting methods during two consecutive seasons, March and August 2019. The statistical analysis showed significant differences between Di, Tri, and Tetra when S and H methods were used (Figure 2).
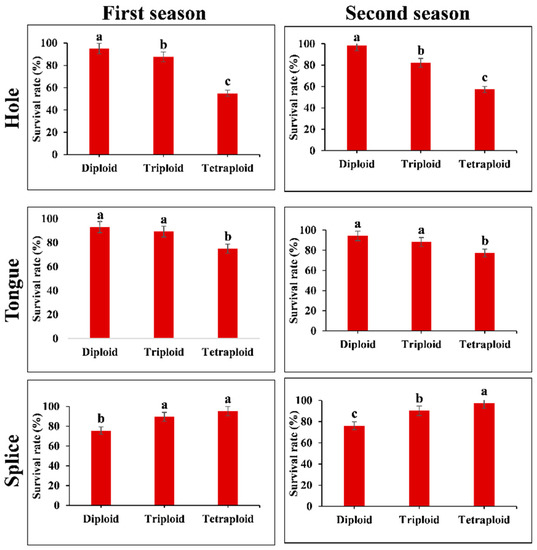
Figure 2.
The survival rate of different grafting methods among different ploidy watermelon at 15 days after grafting during March and August 2019 cropping seasons. The bars with different letters differ significantly (p < 0.05).
The results showed that the splice method was more compatible with tetraploid and triploid and gave a higher success rate than the diploid. On the contrary, the hole method was compatible with diploid more and gave a survival rate higher than the tetraploid. While the tongue method was lower in survival rate results and no high differences between polyploids, the following measurements will be completed on the splice and hole methods to study compatibility mechanisms and study the effect of genome duplication in graft compatibility.
3.2. Measurement of Indole-3-Acetic Acid (IAA) and Zeatin Riboside (ZR) in the Grafting Union among Di, Tri, and Tetraploid Watermelon at Different Days after Grafting
Diploid, triploid, and tetraploid watermelon grafted with splice and hole methods showed a significant effect on the graft union’s hormones contents during the grafting process (Figure 3). In this study, hormone contents increased during the grafting process, especially in 2–3 DAG; our results also showed that the hormone contents in Tetra were more than Tri and Di. The contents of hormones and their increment rates differ according to the grafting method, as there was a difference between splice and hole methods.
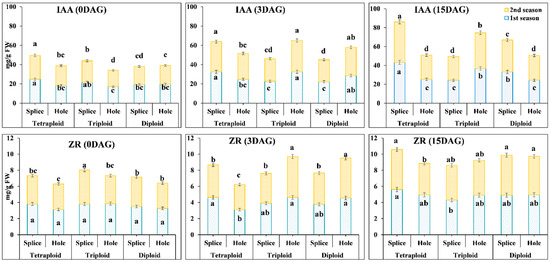
Figure 3.
IAA and ZR hormones contents in different ploidy watermelon grafted with splice and hole methods at 0, 3, and 15 DAG, during March and August 2019 cropping seasons grafted with splice and hole methods. The bars with different letters differ significantly (p < 0.05). IAA: Indole acetic acid, ZR: Zeatin Riboside, DAG: Days after grafting.
In the splice method at 0 DAG, IAA contents in Tetra were 1.28 and 1.32-fold higher than Di, while in Tri, IAA was 1.13 and 1.18-fold higher than Di in the first and second season, respectively. Also, the increment rate of IAA in the graft union at 3 DAG were 14.48, 3.93, and 30.24% in the first season and 21.97, 5.14, and 27.32% in the second season in Di, Tri, and Tetra watermelon, respectively. The increment rate of IAA in the graft union at 15 DAG were 48.63, 7.07, and 33.32% in the first season and 48.55, 7.1, and 36.62% in the second season in Di, Tri, and Tetra watermelon, respectively. While in hole method, the results showed different direction as were the results of IAA showed that no significant differences between different ploidy watermelon at 0 DAG, but the increment rates at 3 DAG were 45.12, 89.67, and 34.71% in the first season and 51.75, 91.34, and 31.74% in the second season in Di, Tri, and Tetra watermelon, respectively. The increment rates at 15 DAG were −14.57, 13.04, and 3.35% in the first season and −11.2, 16.31, and −4.2% in the second season in Di, Tri Tetra watermelon, respectively.
Whereas ZR results showed no significant differences between Di and Tetra at 0 DAG in both grafting methods, while at 3 and 15 DAG, there were highly significant differences during both seasons (Figure 3). ZR content increases significantly in the different ploidy watermelon during the grafting process, varying with the grafting method. In splice methods, ZR contents at 3 DAG were increased by 8.17, 2.85, and 21.52% in the first season and 5.32, −12.5, and 14.77% in the second season in Di, Tri, and Tetra watermelon, respectively. The increment rate at 15 DAG was 29.26, 9.9, 21.12% in the first season and 28.99, 16.73, and 23.35% in the second season in Di, Tri, and Tetra watermelon, respectively. While with hole methods, the content of ZR at 3 DAG was increased by 39.1, 20.13, and −0.16% in the first season and 58.72, 46.91, and −1.88% in the second season in Di, Tri, and Tetra watermelon, respectively. The increment rate at 15 DAG was 8.53, 5.92, and 58.29% in the first season and −3.57, −14.74, and 25.3% in the second season in Di, Tri, and Tetra watermelon, respectively.
3.3. Measurement of POD, SOD, CAT, and H2O2 Contents in the Grafting Union among Di, Tri, and Tetraploid Watermelon at Different Days after Grafting
Our results showed highly significant differences between different ploidy watermelons during both seasons in POD (Figure 4); when the splice method was used, the highest POD activities were recorded in Tri at 0 DAG, 4.1 and 2.82-fold, compared to Di, and Tetra, in the first season and 3.7 and 1.43-fold in the second season. At 3 DAG, POD activities’ increment rates were 114.4, 24, and 223.53% in the first season, 116.71, 32.82, and 219.13% during the second season Di, Tri, and Tetra watermelon, respectively. While when the hole method was used, the highest POD activities were recorded in Tri at 0 DAG, 2.28, and 1.74-fold, compared to Di and Tetra, in the first season and 2.28 and at least 1.74-fold and the second season. At 3 DAG, POD activities’ increment rates were 421.39, 22.58, and 219.95% in both seasons and 421.39, 22.58, and 219.95% second season in Di, Tri, and Tetra watermelon, respectively.
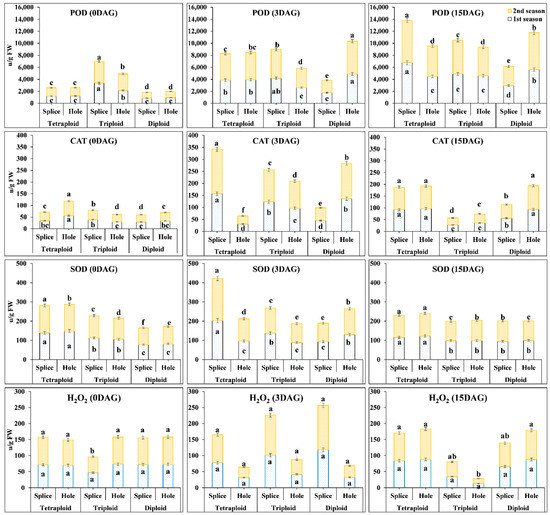
Figure 4.
Peroxidase (POD), Superoxide dismutase (SOD), Catalase (CAT), and Hydrogen peroxide (H2O2) contents in different ploidy watermelons grafted with splice and hole methods at 0, 3, and 15 DAG during March and August 2019 cropping seasons. The bars with different letters differ significantly (p < 0.05). DAG: Days after grafting.
Significantly higher CAT content was observed in Tri than Tetra and Di (Figure 4), at 0 DAG with splice method, CAT activities were 1.35 and 1.3-fold higher in Tri than Di in the first and second season, respectively, and 1.12 and 1.1-fold higher than Tetra in the first and second season, respectively. CAT activities in the graft union start to increase at 3 DAG in all polyploids with increment rates of 57, 217.17, and 354.8% in the first season, 70.1, 226.82, and 400% in the second season in Di, Tri, and Tetra watermelon, respectively. While when the hole method was used, the highest CAT activities were recorded in Tetra at 0 DAG, 1.67, and 1.66-fold, compared with Di and Tri in the first season and 1.74, and 1.99-fold in the second season, respectively. At 3 DAG, the increment rates of CAT activities were 303.25, 252.15, and −45.97% in the first season and 311.39, 261.79, and −45.45% in the second season in Di, Tri, and Tetra watermelon, respectively; at 3 DAG stage, it was observed that catalase activities start to decrease in tetraploid when the hole method was used. In contrast, itis starts to increase with the splice method.
SOD activity was observed higher in Tetra and Tri watermelon than Di watermelon (Figure 4). With the splice method at 0 DAG, SOD activities in Tetra were 1.23 and 1.79-fold higher than Di and Tri in the first season and 1.24 and 1.63-fold in the second season. At 3 DAG, SOD activities start to increase in Tetra than compared to Tri and Di. The increment rates of SOD activities were 20.57, 22.25, and 46% in the first season and 10.2, 13.1, and 53.66% during the second season in Di, Tri, and Tetra watermelon, respectively. With the hole method, the highest SOD activities were recorded in Tetra at 0 DAG, 1.83, and 1.41-fold, compared with Di and Tri in the first season, and 1.53 and 1.26-fold in the second season. At 3 DAG, the increment rates of SOD activities were different with different ploidy where it was increased in Di but decreased in Tetra and Tri; SOD activities were 59.6, −15.9, and −34.81% in the first season and 49.1, −10.66, and −16.39% in the second season in Di, Tri, and Tetra watermelon, respectively, at 3 DAG stage. It was observed that SOD activity with the hole method was very high in Di, but it decreased in the Tri and decreased further in the Tetra.
Reactive Oxygen Species (ROS), like hydrogen peroxide (H2O2) accumulation leads to cell death, depends on the balance between the production of AOX and ROS. H2O2 contents were lower in Tri than Di and Tetra at 0 DAG stage when the splice method was used (Figure 4). It was 0.65 and 0.65-fold lower in Tri than Di and Tetra in the first season and 0.59 and 0.58-fold in the second season. But at 3 DAG, the increment rates of H2O2 activities in the graft union started to increase: 64.1, 116.49, and 8.59% in the first season and 65.57, 153.79, and 2.33% in Di, Tri, and Tetra, respectively. There were no significant differences between Di and Tetra in H2O2 contents at 0 DAG, but the increase at 3 DAG was significant in the Di and not significant in the Tri and Tetra. With the hole method, no significant differences between Di, Tri, and Tetra in the content of H2O2 in both seasons. At 3 DAG, the increment rates of H2O2 contents were 124.95, 77.37, 118.5% in the first season, 134.4, 85.44, and 149.89% in the second season in Di, Tri, and Tetra watermelon, respectively, at the 3 DAG stage.
3.4. Carbohydrates (Starch and Sugars) Contents in the Grafting Union among Di, Tri, and Tetraploid Watermelon with Different Grafting Methods at Different Days after Grafting
Generally, carbohydrates provide energy in the plant. The cotyledons are the source of carbohydrates in the seedlings; the survival rate was positively correlated with increased starch content during the grafting process. The sugars results (Figure 5) showed significant differences between different ploidy watermelons with hole and splice methods in both seasons at 0 DAG, where in Tetra were higher than Tri and Di. With the splice method, total sugars’ contents in Tetra were 1.54 and 1.54-fold in the first season and 2.12 and 2.12-fold in the second season, higher than Di and Tri, respectively. At 3 DAG, the contents of sugars start to decrease, where it was 288.5, 228.94, and 47.7% in the first season and 288.5, 229, 47.7% in the second season in Di, Tri, and Tetra, respectively; it was observed that the decrement rate is significantly lower in Tetra than Di and Tri at 3 DAG with splice method. At 15 DAG, sugars’ contents start to increase 322.29, 42.66, and 12.51% in the first season and 322.29, 42.66, and 12.5% in the second season in Di, Tri, and Tetra, respectively. While with the hole method, sugars’ contents at 0 DAG were higher in Tetra with 2.12 and 2.29-fold than Di and Tri, respectively, in both seasons. At 3 DAG, sugars’ contents start to increase in Di and Tri by 8.9 and 54, 55% in both seasons, while in Tetra, the sugars’ contents begin to decrease by 46% in both seasons. Also, at 15 DAG, sugar contents increased just in Di by 151% in both seasons, while Tri and Tetra decreased by 33.42 and 96.1%, respectively.
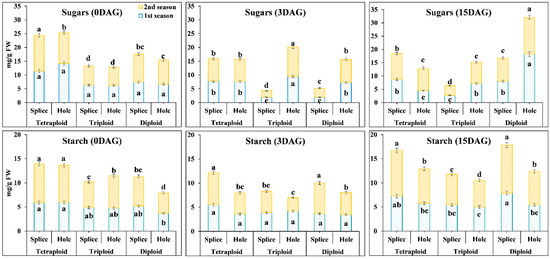
Figure 5.
Sugars and Starch contents at different ploidy watermelon grafted with splice and hole methods at 0, 3, and 15 DAG. The bars with different letters differ significantly (p < 0.05). DAG: Days after grafting.
Starch results in (Figure 5) at 0 DAG using the splice method showed no significant differences between different ploidy watermelons in the first season. In the second season, Tetra starch contents are highly significant (1.31 and 1.51-fold) than compared to Di and Tri. Starch contents start to decrease at 3 DAG in all ploidy. Still, the decrement rates at Tetra were less than Di and Tri, 41.66, 25.31, and 7.38% at the first season, while in the second season no significant differences between 0 and 3 DAG with Di, but the decrement rates were high in Tri and Tetra by 21.11, and 20.9%, respectively. The results showed differences between different ploidy during the grafting process (0, 3, and 15 DAG) but no significance with the hole method. There are significant differences at 0 DAG in the second season, as the starch contents in Tetra were higher than Di and Tri by 1.63 and 1.24-fold in the first season and 1.79, 1.15 in the second season, respectively. At 3 DAG, no significant differences in Di during this stage but with Tri and Tetra’s content start to decrease by 13.38 and 65.68% in the first season and 143.14 and 74.31% in the second season, respectively. At 15 DAG, starch contents increase significantly by 53.85, 18.63, and 60.68% in the first season and 50.14, 101.78, and 62.18% in the second season in Di, Tri, and Tetra, respectively.
3.5. Rootstock Roots Growth and Development by Morphological Parameters
The four grafting combinations’ root morphology parameters (control without grafting, Di, Tri, and Tetra) grafted by splice and hole methods were shown in Figure 6, Figure 7 and Figure 8. There were significant differences in the coefficients of root morphology between levels of ploidy. The rootstock grafted with Tetra watermelon plant roots showed higher length, surface area, projected area, and fresh weight than the other combinations with both methods. The results showed that the growth of rootstock roots length increment rates was higher with Tetra, followed by Tri and Di in both methods. On the other side, Di and Tri’s roots length were higher than the splice method with the hole method. Surface area is an indicator for new roots production; there are significant differences between different ploidy in surface area during the grafting process with both methods. The results showed that the growth of new roots (surface area) was higher with Tetra in both methods than Di and Tri. Tri was a higher increment rate than Di in the surface area with the splice method, while the hole method had no significant differences. The increased production of new roots and the increase in length led to the increased projected area and fresh weight. It was observed in our results that the rootstock grafted with Tetra watermelon plants roots exhibited higher projected area and fresh weight than Di and Tri with both methods, also with the hole method, no significant differences between Di and Tri in the projected area and fresh weight, but it is highly significant with control. During the grafting and healing process, there are significant differences in volume, average diameter, and dry weight in the rootstock roots grafted with Di, Tri, and Tetra watermelon, using splice and hole grafting methods. Rootstock grafted with Tri gave a high increase in roots volume with the splice method followed by Di and Tetra, while with the hole method, Tetra gave a high increase in roots volume followed by Tri and Di. Because the control (without grafting) growing in roots length was less, the average diameter was higher. The results showed that the control rootstock roots gave a high increase in roots diameter than grafted plants in both methods. Finally, rootstock roots dry weight, which grafted with Di and Tri watermelon, gave highly significant weight than Tri and control in both methods.
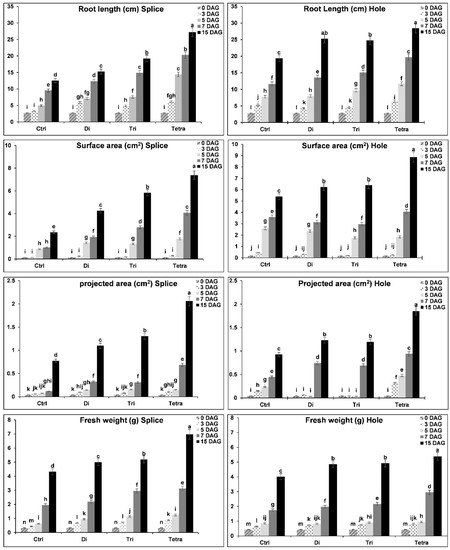
Figure 6.
Roots’ growth and development of Xijiaqiangsheng rootstock grafted with splice and hole grafting methods among Cntl, Di, Tri, and Tetra watermelon at 0, 3, 5, 7, and 15 DAG. The bars with different letters differ significantly (p < 0.05). Cntl: control without grafting, Di: Diploid, Tri: triploid, Tetra: Tetraploid, DAG: Days after grafting.
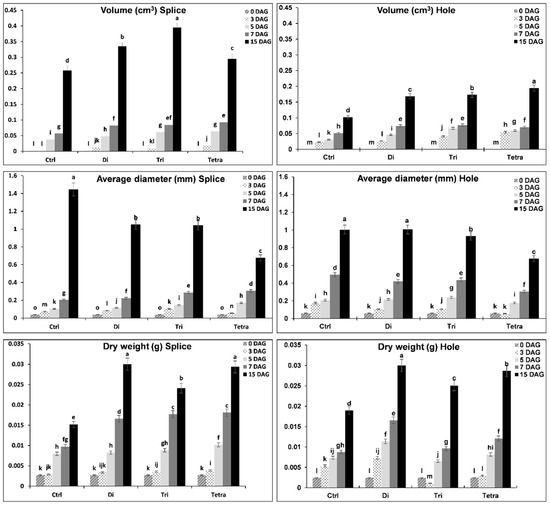
Figure 7.
Roots’ growth and development of Xijiaqiangsheng rootstock grafted with splice and hole grafting methods among Cntl, Di, Tri, and Tetra watermelon at 0, 3, 5, 7, and 15 DAG. The bars with different letters differ significantly (p < 0.05). Cntl: control without grafting, Di: Diploid, Tri: triploid, Tetra: Tetraploid, DAG: Days after grafting.
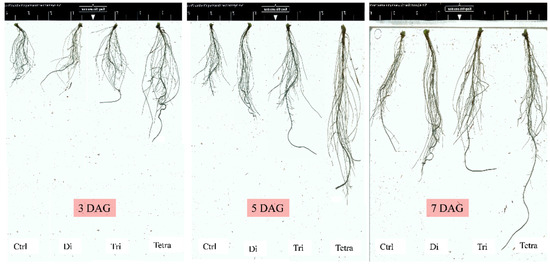
Figure 8.
Roots’ growth and development of Xijiaqiangsheng rootstock grafted with splice methods among Cntl, Di, Tri, and Tetra watermelon at 3, 5, and 7 DAG. Cntl: control without grafting, Di: Diploid, Tri: triploid, Tetra: Tetraploid, DAG: Days after grafting.
3.6. Anatomical Observation
Comparative anatomical studies at the grafting union region (Figure 9) between different ploidy watermelon grafted with splice and hole methods reveal apparent differences between compatible and incompatible combinations (Figure 9). Long adhesion region and new tissues between the scion and rootstock were observed in the compatible plants of Tetra with splice method and Di and Tri with hole method. Conversely, the small adhesion regions were found in the graft union in the incompatible plants of Tetra with hole and Di with the splice method.
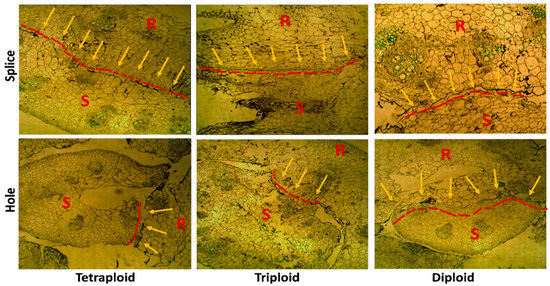
Figure 9.
Anatomical observation of Di, Tri, and Tetra watermelon grafted on squash rootstock using splice and hole grafting methods at 15 Days after grafting (zoom 40×), arrows indicate connection region between scions and rootstock. R: rootstock, S: Scion.
3.7. Principal Component Analysis
PCA can indicate the essential traits contributing to graft compatibility in the materials and methods under study (Figure 10). The resulting PCA scatterplot with the splice method showed a clear cluster for AOX, hormones, sugars, starch (on the right side of PC1), and H2O2 (on the left side of PC1). Other clustered groups showed additional separation in more than one group, in the second, third, and fourth quarters of the centroid, respectively, for Di, Tri, Tetra at different stages (0, 3, 15 DAG) using Splice (S) and Hole (H) grafting methods.
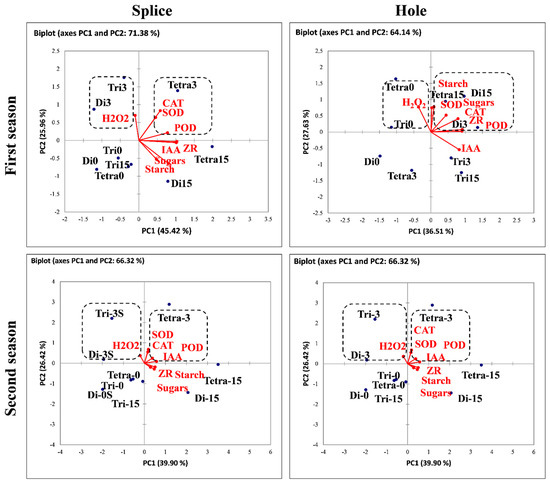
Figure 10.
Principal Component Analysis (PCA) scatterplot based on total concentrations of the AOX, Hormones, starch, and sugars as a function of the grafting combination in Di, Tri, and Tetra watermelon at three stages (0, 3, 15 DAG). Di: Diploid, Tri: triploid, Tetra: Tetraploid.
4. Discussion
In this study, we compared hole, splice, and tongue grafting methods among diploid, triploid, and tetraploid watermelon. The data showed that the hole method gave a high survival rate in Di and triploid watermelon, while the splice method gave a high survival rate with tetraploid watermelon. While with the tongue method, the differences were not very high.
Plant hormones play a vital role in callus initiation and vascular bundle formation at grafting union [29,30,31,34,35,54,56,57,58,59,60,61,62]. During the grafting process, the maximum contents of IAA were observed until 3 DAG in the scion [56,63]. We compared the IAA and ZR contents in Di, Tri, and Tetra watermelon graft unions (Figure 3) using splice and hole grafting methods on different days after grafting. The IAA and ZR contents in Tetra combinations were significantly high compared to Di and Tri combinations with the splice method. Di and Tri had a high content of IAA and ZR when the hole method was used. Our results showed high compatibility in Tetra grafted with splice method and high content and higher increment rates of hormones than Di, especially at 3 DAG (critical period) and 15 DAG. While Di and Tri gave high survival rate with hole method and high increment rates of IAA and ZR than Tetra, these results can explain the high survival rates in Tetra with the splice and the high survival rate in Di and Tri with hole method; these results were in agreement with [56,64,65].
The grafting process faced more stresses like injury or wound, complete dark, and high humidity during healing time [54,55]. The most critical time in the healing process is the 2nd and 3rd days after grafting. [56]. Polyploids are more tolerant of abiotic stresses than Di, and genome doubling gave high resistance to stresses because of the high contents of hormones and antioxidants’ high activities [66,67,68,69].
The results in both seasons showed that POD, SOD, and CAT activities were significantly different between different ploidy watermelons among splice and hole methods (Figure 4). In this study, POD, SOD, and CAT activities showed a high increase during the Tetra healing process with the splice method, while the increment rates were high in Di and Tri with the hole method. On the contrary, the content of hydrogen peroxide (H2O2) did not increase during the healing process in Tetra with the splice method. It didn’t increase in Di and Tri with the hole method. These results were because of the high activities of antioxidants, which leads to scavenging oxygen radicals and H2O2. According to [43,46].
Carbohydrates provide energy to plants, and the cotyledons are the source of carbohydrates in the seedlings [27,50]. The survival rate was positively correlated with an increase in starch content [51]. An increase in the percentage of carbohydrates before grafting by adding sucrose to the rootstocks led to a higher survival rate [50]. In our results, with the splice method, the accumulation of sugars and starch contents in the graft union at 0 DAG was significantly higher in Tetra than in Tri and Di. The decrement rates of sugars and starch were meager compared to Di and Tri. These results may explain the increment in the survival rate in Tetra with the splice method. Same with the hole method, Di and Tri gave a higher survival rate because of the higher content of starch and sucrose at 0 DAG and the lower decrement rates during 3 DAG than tetraploid watermelon. These results are consistent with [50,55], who suggested that higher carbohydrate contents in grafted tissues could lead to successful grafting.
Just as hormones are essential in the process of compatibility and callus formation, so are carbohydrates [50,51]. The experiment has proved that increasing the percentage of carbohydrates increased the grafting survival rate [50].
The roots are more sensitive to auxin than the stem. The roots can be elongated by adding auxin [36,37,38,67]. In the Arabidopsis, after germination, the root meristem growth is controlled by the interaction between cytokinin (promoting differentiation) and auxin (promoting division) [70]. Our experiment measured the rootstock roots’ growth rate and compared the effect of grafting by Di, Tri, and Tetra on roots’ development. It was found that in the case of tetraploid, the rate of rootstock roots growth was much higher during the grafting process than diploid and triploid. This could be due to the high hormone contents and the high interaction between cytokinin and auxin in tetraploid watermelon than diploid and triploid [36,37,38,67,70].
Rootstock–scion compatibility depends on anatomical, physiological, and genetic variables, and the survival rate increases with fewer differences between origin and taste [71]. The survival rate was inversely related to the difference between the diameters of the scion and the rootstock. Our anatomical study showed that the adhesion region in tetraploid with the splice method was long compared to the hole method. Since the tetraploid seedling is flat (not round), the hole method is not suitable for grafting. In the diploid, the slides showed that the adhesion area in the hole is bigger and longer than the splice; this may explain the high survival rate in the diploid with the hole method. Likewise, for triploid, the splice’s adhesion area was bigger than the hole, explaining the increased survival rate in the splice more than the hole.
The PCA scatter plot highlighted that the hormones, AOX, sugars, and starch had a pivotal role in the graft compatibility of Di, Tri, and Tetra watermelon during 0, 3, and 15 DAG using splice and hole grafting methods. Indeed, all AOX, hormones, and carbohydrates in the grafting union clustered together on the positive side of PC1. The opposite was noticed with H2O2 on the negative side of PC1 in both seasons.
5. Conclusions
This study investigated physiological and anatomical changes in graft-compatible and graft-incompatible polyploid watermelon with different grafting methods. Our results suggest that the higher content of hormones, antioxidants activities, and carbohydrates, would contribute to superior physiology, callus initiation, differentiation, and growth performance in the graft-compatible combination. These results provide us some advice, such as exogenous applications with sugars or hormones before grafting and reduce the stresses on the plant during the healing period to increase the survival rate; additionally, to study the effect of genome doubling on graft compatibility. Our findings should further understand the compatibility mechanisms to improve plant growth performance and cause incompatible rootstock grafting to influence plant growth. Also, it can open the door for breeders to produce tetraploid rootstock, which has high compatibility and high resistance for abiotic and biotic stresses.
Author Contributions
M.O.K. and W.L. visualize and designed the experiments. M.O.K. performed the experiments. M.O.K. and M.J.U. analyzed the data and wrote the manuscript. E.M., W.D., P.Y., H.Z., S.Z., X.L., N.H., M.A. and E.E.-r. participated in the experimentations and analysis. M.J.U., M.A., H.Z. and W.L. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2021-ZFRI), China Agriculture Research System (CARS-25-03), National Key R&D Program of China (2018YFD0100704), the National Natural Science Foundation of China (Nos. 31471893; 31672178)
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors declare that supporting data for the study findings are presented in the manuscript, and available from the corresponding author upon request.
Acknowledgments
The authors thank all the Polyploid Watermelon Research Group members, Zhengzhou Fruit Research Institute; Chinese Academy of Agricultural Sciences.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marr, C.W.; Gast, K.L.B.; Charles, W.M.; Karen, L.B. Gast 2 1. Reactions by consumers in a farmers’ market to prices for seedless watermelon and ratings of eating quality. HortTechnology 1991, 1, 105–106. [Google Scholar] [CrossRef]
- Jaskani, M.J.; Kwon, S.W.; Kim, D.H. Comparative study on vegetative, reproductive and qualitative traits of seven diploid and tetraploid watermelon lines. Euphytica 2005, 145, 259–268. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- Noh, J.; Sheikh, S.; Chon, H.G.; Seong, M.H.; Lim, J.H.; Lee, S.G.; Jung, G.T.; Kim, J.M.; Ju, H.-J.; Huh, Y.C. Screening different methods of tetraploid induction in watermelon [Citrullus lanatus (thunb.) Manst. and Nakai]. Hortic. Environ. Biotechnol. 2012, 53, 521–529. [Google Scholar] [CrossRef]
- Zhang, N.; Bao, Y.; Xie, Z.; Huang, X.; Sun, Y.; Feng, G.; Zeng, H.; Ren, J.; Li, Y.; Xiong, J. Efficient Characterization of Tetraploid Watermelon. Plants 2019, 8, 419. [Google Scholar] [CrossRef]
- Hassan, J.; Miyajima, I.; Ozaki, Y.; Mizunoe, Y.; Sakai, K.; Zaland, W. Tetraploid Induction by Colchicine Treatment and Crossing with a Diploid Reveals Less-Seeded Fruit Production in Pointed Gourd (Trichosanthes dioica Roxb.). Plants 2020, 9, 370. [Google Scholar] [CrossRef]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Misra, B.B.; Shan, S.; Chen, S.; Soltis, P.S. Polyploidy and the proteome. Biochim. Biophys. Acta BBA Proteins Proteom. 2016, 1864, 896–907. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, S.; Lu, X.; He, N.; Gao, L.; Dou, J.; Bie, Z.; Liu, W. Plant Physiology and Biochemistry Genome duplication improves the resistance of watermelon root to salt stress. Plant Physiol. Biochem. 2018, 133, 11–21. [Google Scholar] [CrossRef]
- Doyle, J.J.; Coate, J.E. Polyploidy, the nucleotype, and novelty: The impact of genome doubling on the biology of the cell. Int. J. Plant Sci. 2019, 180, 1–52. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, S.; Cheng, Z.; Wan, X.; Yan, Z.; King, S. Lycopene and citrulline contents in watermelon (Citrullus lanatus) fruit with different ploidy and changes during fruit development. Acta Hortic. 2010, 871, 543–550. [Google Scholar]
- Ulas, F.; Aydın, A.; Ulas, A.; Yetisir, H. Grafting for Sustainable Growth Performance of Melon (Cucumis melo) Under Salt Stressed Hydroponic Condition. Eur. J. Sustain. Dev. 2019, 8, 201–210. [Google Scholar] [CrossRef]
- Monteiro, G.C.; Goto, R.; Minatel, I.O.; de Sousa da Silva, E.; da Silva, E.G.; Vianello, F.; Lima, G.P.P. Grafting, Agrochemicals, and Oxidative Enzymes as Factor for Plant Biotic Resistance. In Plant Health Under Biotic Stress: Volume 1: Organic Strategies; Ansari, R.A., Mahmood, I., Eds.; Springer: Singapore, 2019; pp. 37–57. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Taji, A.; Backhouse, D.; Oda, M. Anatomy and physiology of graft incompatibility in solanaceous plants. J. Hortic. Sci. Biotechnol. 2008, 83, 581–588. [Google Scholar] [CrossRef]
- El-Gazzar, T.; Dawa, K.K.; Ibrahim, E.; El-Banna, M.; Mohamed, A. Anatomical Study on Watermelon Grafting. J. Plant Prod. 2017, 8, 999–1009. [Google Scholar] [CrossRef][Green Version]
- Assunção, M.; Canas, S.; Cruz, S.; Brazão, J.; Zanol, G.C.; Eiras-Dias, J.E. Graft compatibility of Vitis spp.: The role of phenolic acids and flavanols. Sci. Hortic. 2016, 207, 140–145. [Google Scholar] [CrossRef]
- Pina, A.; Cookson, S.J.; Calatayud, A.; Trinchera, A.; Errea, P. Physiological and molecular mechanisms underlying graft compatibility. Veg. Grafting Princ. Pract. 2017, 10, 132–154. [Google Scholar] [CrossRef]
- Dogra, K.; Kour, K.; Kumar, R.; Bakshi, P.; Kumar, V. Graft Incompatibility in Horticultural Crops. Int. J. Curr. Microbiol. Appl. Sci. 2018. [Google Scholar] [CrossRef]
- Melnyk, C.W. Plant grafting: Insights into tissue regeneration. Regeneration 2017, 4, 3–14. [Google Scholar] [CrossRef]
- Hassell, R.L.; Memmott, F.; Liere, D.G. Grafting Methods for Watermelon Production. HortScience 2008, 43, 1677–1679. [Google Scholar] [CrossRef]
- Kubota, C.; McClure, M.; Burelle, N.; Bausher, M.; Rosskopf, E.; Chellemi, D. How to Graft Cucurbits. Multi-State Project Use of Grafted Seedlings for Methyl Bromide Transition in US Open-Field Fresh Vegetable Production; Arizona University: Tucson, AZ, USA, 2010; Available online: http://cals.arizona.edu/grafting/howto/cucurbits/grafting_methods (accessed on 1 March 2021).
- Oda, J.L.M.; Lee, M. Grafting of herbaceous vegetable and ornamental crops. Hortic. Rev. 2003, 28, 61–124. [Google Scholar]
- Oda, I. New Grafting Methods for Fruit-Bearing Vegetables in Japan. Jpn. Agric. Res. Q. JARQ 1995, 29, 187–194. [Google Scholar]
- Cushman, K. Grafting Techniques for Watermelon. EDIS Florida: HS1075-University of Florida IFAS Extension. USA. 2007. Available online: http://edis.ifas.ufl.edu (accessed on 1 March 2021).
- Memmott, F. Refinement of Innovative Watermelon Grafting Methods with Appropriate Choice of Developmental Stage, Rootstock Type, and Root Treatment to Increase Grafting Success. Master’s Thesis, Clemson University, Clemson, SC, USA, 2010. [Google Scholar]
- Abd El-Wanis, M.; El-Eslamboly, A.; Salama, M. Impact of different grafting methods on yield and quality of watermelon. Res. J. Agric. Biol. Sci. 2013, 9, 330–340. [Google Scholar]
- Khankahdani, H.H.; Zakeri, E.; Saeedi, G.; Shakerdargah, G. Evaluation of different rootstocks and grafting techniques on graft union percent, yield and yield components of watermelon cv. ‘Crimson Sweet’. World Appl. Sci. J. 2012, 18, 645–651. [Google Scholar]
- Mo, Z.; Feng, G.; Su, W.; Liu, Z.; Peng, F. Transcriptomic Analysis Provides Insights into Grafting Union Development in Pecan (Carya illinoinensis). Genes 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R.; Perkins-Veazie, P.; Sakata, Y.; López-Galarza, S.; Maroto, J.V.; Lee, S.-G.; Huh, Y.-C.; Sun, Z.; Miguel, A.; King, S.R.; et al. Cucurbit Grafting. Crit. Rev. Plant Sci. 2008, 27, 50–74. [Google Scholar] [CrossRef]
- Lu, S.; Song, Y. Relation between phytohormone level and vascular bridge differentiation in graft union of explanted internode autografting. Chin. Sci. Bull. 1999, 44, 1874–1878. [Google Scholar] [CrossRef]
- Ren, Y.; Guo, S.r.; Shu, S.; Xu, Y.; Sun, J. Isolation and expression pattern analysis of CmRNF5 and CmNPH3L potentially involved in graft compatibility in cucumber/pumpkin graft combinations. Sci. Hortic. 2018, 227, 92–101. [Google Scholar] [CrossRef]
- Ren, Y.; Xu, Q.; Wang, L.; Guo, S.; Shu, S.; Lu, N.; Sun, J. Plant Physiology and Biochemistry Involvement of metabolic, physiological and hormonal responses in the graft-compatible process of cucumber/pumpkin combinations was revealed through the integrative analysis of mRNA and miRNA expression. Plant Physiol. Biochem. 2018, 129, 368–380. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, J.; Hu, F.; Qin, Y.; Wang, X.; Hu, G. Transcriptome changes between compatible and incompatible graft combination of Litchi chinensis by digital gene expression profile. Sci. Rep. 2017, 7, 3954. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Gabel, A.; Hardcastle, T.J.; Robinson, S.; Miyashima, S.; Grosse, I.; Meyerowitz, E.M. Transcriptome dynamics at Arabidopsis graft junctions reveal an intertissue recognition mechanism that activates vascular regeneration. Proc. Natl. Acad. Sci. USA 2018, 115, E2447–E2456. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, E. Regulation of root growth by plant hormones—roles for auxin and gibberellin. Crit. Rev. Plant Sci. 2005, 24, 249–265. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Pilet, P.-E.; Saugy, M. Effect on root growth of endogenous and applied IAA and ABA: A critical reexamination. Plant Physiol. 1987, 83, 33–38. [Google Scholar] [CrossRef]
- Zafar, S.A.; Patil, S.B.; Uzair, M.; Fang, J.; Zhao, J.; Guo, T.; Yuan, S.; Uzair, M.; Luo, Q.; Shi, J.; et al. DEGENERATED PANICLE AND PARTIAL STERILITY 1 (DPS1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol. 2019, 1. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Noor, R.S.; Wang, Z.; Umair, M.; Yaseen, M.; Ameen, M.; Rehman, S.-U.; Khan, M.U.; Imran, M.; Ahmed, W.; Sun, Y. Interactive Effects of Grafting Techniques and Scion-Rootstocks Combinations on Vegetative Growth, Yield and Quality of Cucumber (Cucumis sativus L.). Agronomy 2019, 9, 288. [Google Scholar] [CrossRef]
- Meng, J.; Wu, S.; Wang, X.; Yu, X.; Jiang, R. Effects of Different Rootstocks on Plant Growth and Fruit Quality of Watermelon. Agric. Biotechnol. 2019, 8, 64–68. [Google Scholar]
- Ahmad, P.; Wani, M.R. Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment: Volume 1; Springer: New York, NY, USA, 2014; pp. 1–376. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, J.; Qin, Y.; Hu, G. Study on the graft compatibility between ‘Jingganghongnuo’ and other litchi cultivars. Sci. Hortic. 2016, 199, 56–62. [Google Scholar] [CrossRef]
- Fernandez-Garcia, N.; Carvajal, M.; Olmos, E. Graft union formation in tomato plants: Peroxidase and catalase involvement. Ann. Bot. 2004, 93, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Aloni, B.; Karni, L.; Deventurero, G.; Levin, Z.; Cohen, R.; Katzir, N.; Lotan-Pompan, M.; Edelstein, M.; Aktas, H.; Turhan, E.; et al. Physiological and biochemical changes at the rootstock-scion interface in graft combinations between Cucurbita rootstocks and a melon scion. J. Hortic. Sci. Biotechnol. 2008, 83, 777–783. [Google Scholar] [CrossRef]
- Nocito, F.F.; Espen, L.; Fedeli, C.; Lancilli, C.; Musacchi, S.; Serra, S.; Sansavini, S.; Cocucci, M.; Sacchi, G.A. Oxidative stress and senescence-like status of pear calli co-cultured on suspensions of incompatible quince microcalli. Tree Physiol. 2010, 30, 450–458. [Google Scholar] [CrossRef][Green Version]
- Rapaka, V.K.; Faust, J.E.; Dole, J.M.; Runkle, E.S. Diurnal carbohydrate dynamics affect postharvest ethylene responsiveness in portulaca (Portulaca grandiflora ‘Yubi Deep Rose’) unrooted cuttings. Postharvest Biol. Technol. 2007, 44, 293–299. [Google Scholar] [CrossRef]
- Dabirian, S.; Miles, C.A. Increasing Survival of Splice-grafted Watermelon Seedlings Using a Sucrose Application. HortScience 2017, 52, 579–583. [Google Scholar] [CrossRef]
- Hunter, J.; Volschenk, C.; Le Roux, D.; Fouché, G.; Adams, L. Plant Material Quality, a compilation of research. In Research Reports; ARC Infruitec-Nietvoorbij: Stellenbosch, South Africa, 2004; p. 49. [Google Scholar]
- Oda, M.; Tsuji, K.; Sasaki, H. Effect of Hypocotyl Morphology on Survival Rate and Growth of Cucumber Seedlings Grafted on Cucurbita spp. Jpn. Agric. Res. Q. 1993, 26, 259. [Google Scholar]
- Tan, S.; Liu, J.; Huang, X.; Li, S. Selection of New Watermelon Variety ‘Mimei Tetraploid’. J. Fruit Tree Sci. 1993, 10, 87–91. [Google Scholar]
- Traka-Mavrona, E.; Koutsika-Sotiriou, M.; Pritsa, T. Response of squash (Cucurbita spp.) as rootstock for melon (Cucumis melo L.). Sci. Hortic. 2000, 83, 353–362. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Schuster, C.; Leyser, O.; Meyerowitz, E.M. A Developmental Framework for Graft Formation and Vascular Reconnection in Arabidopsis thaliana. Curr. Biol. 2015, 25, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Gainza, F.; Opazo, I.; Muñoz, C. Graft incompatibility in plants: Metabolic changes during formation and establishment of the rootstock/scion union with emphasis on Prunus species. Chil. J. Agric. Res. 2015, 75, 28–34. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Goldschmidt, E.E. Plant grafting: New mechanisms, evolutionary implications. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yan, B.; Sun, J.; Jia, P.; Zhang, Z.; Yan, X.; Chai, J.; Ren, Z.; Zheng, G.; Liu, H. Graft-union development: A delicate process that involves cell-cell communication between scion and stock for local auxin accumulation. J. Exp. Bot. 2012, 63, 4219–4232. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.K.; Melnyk, C.W. The role of plant hormones during grafting. J. Plant Res. 2018, 131, 49–58. [Google Scholar] [CrossRef]
- Donner, T.J.; Sherr, I.; Scarpella, E. Regulation of preprocambial cell state acquisition by auxin signaling in Arabidopsis leaves. Development 2014. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, L.; Wu, R. Plant grafting: How genetic exchange promotes vascular reconnection. New Phytol. 2017, 214, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Mazur, E.; Benková, E.; Friml, J. Vascular cambium regeneration and vessel formation in wounded inflorescence stems of Arabidopsis. Nat. Publ. Group 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Zheng, B.S.; Chu, H.L.; Jin, S.H.; Huang, Y.J.; Wang, Z.J.; Chen, M.; Huang, J.Q. cDNA-AFLP analysis of gene expression in hickory (Carya cathayensis) during graft process. Tree Physiol. 2009, 30, 297–303. [Google Scholar] [CrossRef]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef]
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.; Quiñones, A.; Martínez-Alcántara, B.; Aleza, P.; Morillon, R.; Navarro, L.; Primo-Millo, E.; Martínez-Cuenca, M.-R. Effects of salinity on diploid (2x) and doubled diploid (4x) Citrus macrophylla genotypes. Sci. Hortic. 2016, 207, 33–40. [Google Scholar] [CrossRef]
- Jiang, A.; Gan, L.; Tu, Y.; Ma, H.; Zhang, J.; Song, Z.; He, Y.; Cai, D.; Xue, X. The effect of genome duplication on seed germination and seedling growth of rice under salt stress. Aust. J. Crop Sci. 2013, 7, 1814. [Google Scholar]
- Tu, Y.; Jiang, A.; Gan, L.; Hossain, M.; Zhang, J.; Peng, B.; Xiong, Y.; Song, Z. Genome duplication improves rice root resistance to salt stress. Rice 2014, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Perilli, S.; Ioio, R.D.; Di Mambro, R.; Costantino, P.; Sabatini, S. The rate of cell differentiation controls the Arabidopsis root meristem growth phase. Curr. Biol. 2010, 20, 1138–1143. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).