Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes

 ,
, 
 ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Field Experiments
2.2. Evaluation of Agronomic Traits
2.3. Statistical Analysis
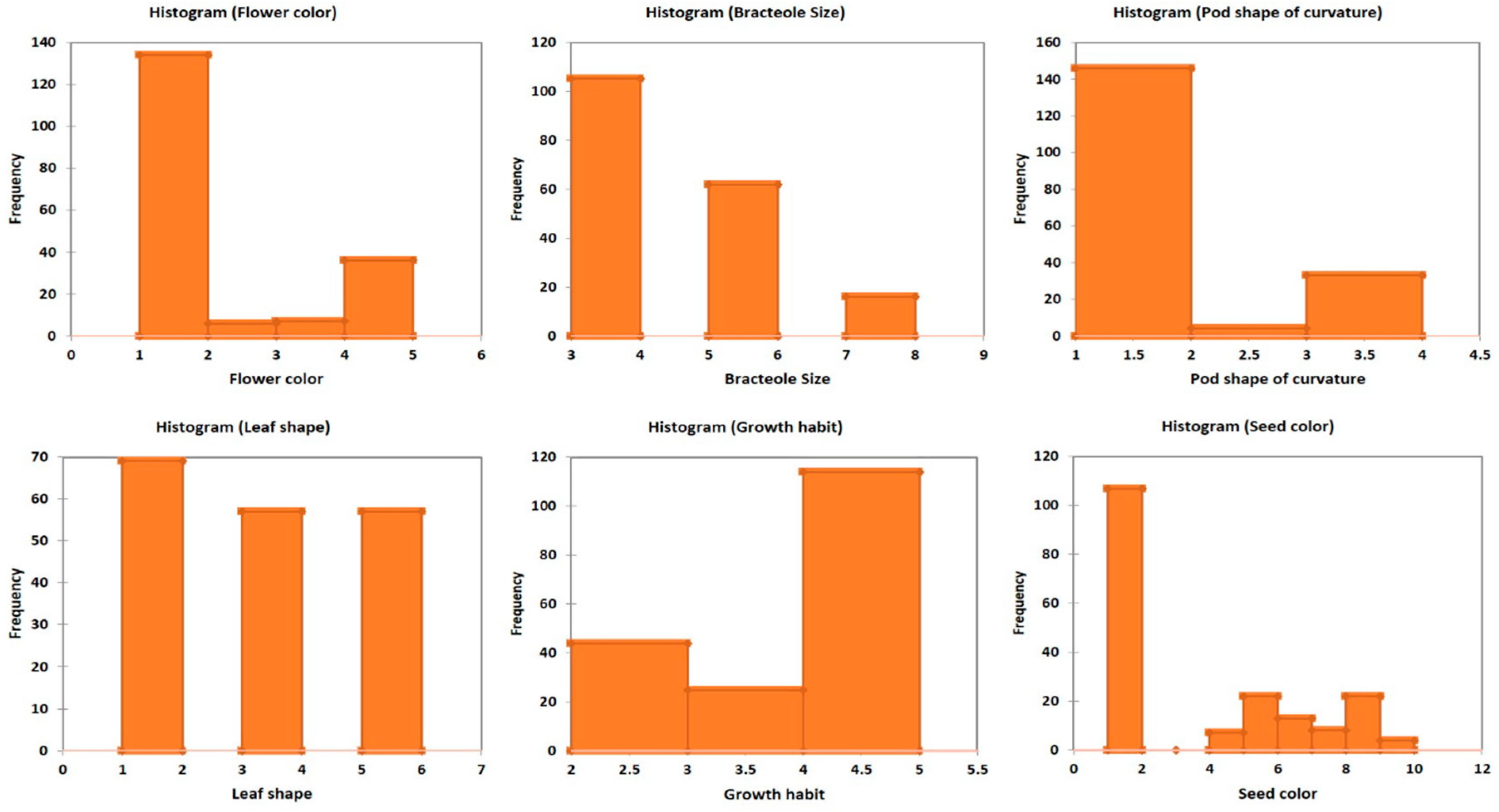
3. Results
Correlation Coefficients Among the Different Quantitative Traits and Multivariate Analysis
4. Discussion
Selection of Superior Accessions for the Characters of Interest
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Khush, G.S.; Lee, S.; Cho, J.I.; Jeon, J.S. Biofortification of crops for reducing malnutrition. Plant Biotechnol. Rep. 2012, 6, 195–202. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.A.; Gündoğdu, M.; Ercişli, S.; Karaköy, T.; Saracoğlu, O.; Habyarimana, E.; Lin, X.; Hatipoğlu, R.; Nawaz, M.A.; Sameeullah, M.; et al. Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean. Genes 2020, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Baloch, F.S.; Alsaleh, A.; Shahid, M.Q.; Çiftçi, V.; De Miera, L.E.; Aasim, M.; Nadeem, M.A.; Aktaş, H.; Özkan, H.; Hatipoğlu, R. A whole genome DArTseq and SNP analysis for genetic diversity assessment in durum wheat from central fertile crescent. PLoS ONE 2017, 12, e0167821. [Google Scholar] [CrossRef]
- Islam, F.M.A.; Basford, K.E.; Redden, R.J.; Beebe, S. Preliminary evaluation of the common bean core collection at CIAT. Plant Genet. Resour. Newsl. 2006, 145, 29–37. [Google Scholar]
- Singh, S.P.; Terán, H.; Muñoz, C.G.; Takegami, J.C. Two cycles of recurrent selection for seed yield in common bean. Crop Sci. 1999, 39, 391–397. [Google Scholar] [CrossRef]
- Ozturk, I.; Kara, M.; Yildiz, C.; Ercisli, S. Physico-mechanical seed properties of the common Turkish bean (Phaseolus vulgaris) cultivars ‘Hinis’ and ‘Ispir’. N. Z. J. Crop Hortic. Sci. 2009, 37, 41–50. [Google Scholar] [CrossRef]
- Bozoğlu, H.; Sozen, O.A. sample for biodiversity in Turkey: Common bean (Phaseolus vulgaris L.) landraces from Artvin. Afr. J. Biotechnol. 2011, 10, 13789–13796. [Google Scholar] [CrossRef]
- Angioi, S.A.; Rau, D.; Attene, G.; Nanni, L.; Bellucci, E.; Logozzo, G.; Negri, V.; ZeuliR, P.L.S.; Papa, R. Beans in Europe: Origin and structure of the European landraces of Phaseolus vulgaris L. Theor. Appl. Genet. 2010, 121, 829–843. [Google Scholar] [CrossRef]
- Şehirali, S. Yemeklik Dane Baklagiller; Ankara Üniversitesi Ziraat Fakültesi: Ankara, Turkey, 1988; Pulses. [Google Scholar]
- Aydın, M.F.; Baloch, F.S. Exploring the genetic diversity and population structure of Turkish common bean germplasm by the iPBS-retrotransposons markers. Legume Res. 2018, LR–423, 1–7. [Google Scholar]
- Nadeem, M.A.; Habyarimana, E.; Çiftçi, V.; Nawaz, M.A.; Karaköy, T.; Comertpay, G.; Shahid, M.Q.; Hatipoğlu, R.; Yeken, M.Z.; Ali, F.; et al. Characterization of genetic diversity in Turkish common bean gene pool using phenotypic and whole-genome DArTseq-generated silicoDArT marker information. PLoS ONE 2018, 13, e0205363. [Google Scholar] [CrossRef] [PubMed]
- FAO (Food and Agriculture Organization of the United Nations). Available online: http://www.fao.org/news/archive/news-by-date/2010/en/?page=5&ipp=10 (accessed on 5 September 2018).
- Shrestha, S.; Asch, F.; Dusserre, J.; Ramanantsoanirina, A.; Brueck, H. Climate effects on yield components as affected by genotypic responses to variable environmental conditions in upland rice systems at different altitudes. Field Crop Res. 2012, 134, 216–228. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F.; Hardy-Weinberg, E. Introduction to Quantitative Genetics, 4th ed.; Addison Wesley Longman: Harlow, UK, 1996. [Google Scholar]
- Gauch, H.G.; Zobel, R.W. AMMI analysis of yield trials. In Genotype by Environment Interacrtion; Kang, M.S., Gauch, H.G., Eds.; CRC Press: New York, NY, USA, 1996; pp. 85–122. [Google Scholar]
- Martins, S.R.; Vences, F.J.; Miera, L.E.S.; Barroso, M.R.; Carnide, V. RAPD analysis of genetic diversity among and within Portuguese landraces of common white bean (Phaseolus vulgaris L.). Sci. Hortic. 2006, 108, 133–142. [Google Scholar] [CrossRef]
- Boros, L.; Wawer, A.; Borucka, K. Morphological, phenological and agronomical characterisation of variability among common bean (Phaseolus vulgaris L.) local populations from The National Centre for Plant Genetic Resources: Polish Genebank. J. Hortic. Res. 2014, 22, 123–130. [Google Scholar] [CrossRef]
- Stoilova, T.; Pereira, G.; De Sousa, M. Morphological characterization of a small common bean (Phaseolus vulgaris L.) collection under different environments. J. Cent. Eur. Agric. 2013, 14, 19. [Google Scholar] [CrossRef]
- Scarano, D.; Rubio, F.; Ruiz, J.J.; Rao, R.; Corrado, G. Morphological and genetic diversity among and within common bean (Phaseolus vulgaris L.) landraces from the Campania region (Southern Italy). Sci. Hortic. 2014, 180, 72–80. [Google Scholar] [CrossRef]
- Da Costa Vaz, D.; de Morais Júnior, O.P.; Peixoto, N. Agro-morphological characterization and genetic divergence assessment in bush snap bean genotypes. Pesqui. Agropecu. Trop. 2017, 47, 134–144. [Google Scholar]
- Rana, J.C.; Sharma, T.R.; Tyagi, R.K.; Chahota, R.K.; Gautam, N.K.; Singh, M.; Sharma, P.N.; Ojha, S.N. Characterisation of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica 2015, 205, 441–457. [Google Scholar] [CrossRef]
- Madakbaş, S.Y.; Ergin, M. Morphological and phenological characterization of Turkish bean (Phaseolus vulgaris L.) genotypes and their present variation states. Afr. J. Agric. Res. 2011, 6, 6155–6166. [Google Scholar]
- Khaidizar, M.I.; Haliloglu, K.; Elkoca, E.; Aydin, M.; Kantar, F. Genetic diversity of common bean (Phaseolus vulgaris L.) landraces grown in northeast Anatolia of Turkey assessed with simple sequence repeat markers. Turk. J. Field Crop. 2012, 17, 145–150. [Google Scholar]
- Ceylan, A.; Öcal, N.; Akbulut, M. Genetic diversity among the Turkish common bean cultivars (Phaseolus vulgaris L.) as assessed by SRAP, POGP and cpSSR markers. Biochem. Syst. Ecol. 2014, 1, 219–229. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 1984. [Google Scholar]
- International Board for Plant Genetic Resources (IPGRI). Phaseolus Vulgaris Descriptors; International Board for Plant Genetic Resources: Rome, Italy, 1982. [Google Scholar]
- Aghamir, F.; Bahrami, H.; Malakouti, M.J.; Eshghi, S.; Sharifi, F. Seed germination and seedling growth of bean (Phaseolus vulgaris) as influenced by magnetized saline water. Eurasian J. Soil Sci. 2016, 5, 39–46. [Google Scholar] [CrossRef]
- Federer, W.T. Augmented (or hoonuiaku) designs. Hawaii Plant Rec. 1956, 55, 191–208. [Google Scholar]
- Rathore, A.; Parsad, R.; Gupta, V.K. Computer aided construction and analysis of augmented designs. J. Indian Soc. Agric. Stat. 2004, 57, 320–344. [Google Scholar]
- Pereira, S.H.; Alvares, R.C.; De Cássia Silva, F.; De Faria, L.C.; Cunha Melo, L. Genetic, environmental and genotype x environment interaction effects on the common bean grain yield and commercial quality. Semin. Cienc. Agrar. 2017, 38, 1241–1250. [Google Scholar] [CrossRef][Green Version]
- Habyarimana, E.; Bonardi, P.; Laureti, D.; Di Bari, V.; Casentino, S.; Lorenzoni, C. Multilocational evaluation of biomass sorghum hybrids under two stand densities and variable water supply in Italy. Ind. Crop. Prod. 2004, 20, 3–9. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 20 August 2019).
- Barili, L.D.; Vale, N.M.; Prado, A.L.; Carneiro, J.E.; Silva, F.F.; Nascimento, M. Genotype-environment interaction in common bean cultivars with carioca grain, recommended for cultivation in Brazil in the last 40 years. Crop Breed. Appl. Biotechnol. 2015, 15, 244–250. [Google Scholar] [CrossRef]
- Oliveira, G.V.; Carneiro, P.C.; De Souza Carneiro, J.E.; Cruz, C.D. Adaptabilidade e estabilidade de linhagens de feijão comum em Minas Gerais. Pesqui. Agropecu. Bras. 2006, 41, 257–265. [Google Scholar] [CrossRef][Green Version]
- Rocha, G.S.; Carneiro, J.E.; Rezende Júnior, L.D.; Menezes Júnior, J.Â.; Carneiro, P.C.; Cecon, P.R. Effect of environments on the estimated genetic potential of segregating common bean populations. Crop Breed. Appl. Biotechnol. 2013, 13, 241–248. [Google Scholar] [CrossRef][Green Version]
- Okii, D.; Mukankusi, C.; Sebuliba, S.; Tukamuhabwa, P.; Tusiime, G.; Talwana, H.; Odong, T.; Namayanja, A.; Paparu, P.; Nkalubo, S.; et al. Genetic variation, Heritability estimates and GXE effects on yield traits of Mesoamerican common bean (Phaseolus vulgaris L) germplasm in Uganda. Plant Genet. Resour. 2018, 16, 237–248. [Google Scholar] [CrossRef]
- Robinson, H.F. Quantitative genetics in relation to breeding of the centennial of mendalism. Indian J. Genet. 1966, 26, 171–187. [Google Scholar]
- Phuke, R.M.; Anuradha, K.; Radhika, K.; Jabeen, F.; Anuradha, G.; Ramesh, T.; Hariprasanna, K.; Mehtre, S.P.; Deshpande, S.P.; Anil, G.; et al. Genetic variability, genotype 9 environment interaction, correlation, and GGE biplot analysis for grain iron and zinc concentration and other agronomic traits in RIL population of sorghum (Sorghum bicolor L. Moench). Front. Plant Sci. 2017, 8, 712. [Google Scholar] [CrossRef] [PubMed]
- Wondimu, W.; Bogale, A. Genetic Variability, Heritability and Genetic Advance of Some Haricot Bean (Phaseolus vulgaris L.) Varieties at Bench-Maji Zone, Southwest Ethiopia. Asian J. Crop Sci. 2017, 9, 133–140. [Google Scholar] [CrossRef]
- Scully, B.T.; Wallace, D.H.; Viands, D.R. Heritability and correlation of biomass, growth rates, harvest index, and phenology to the yield of common beans. J. Am. Soc. Hortic. Sci. 1991, 116, 127–130. [Google Scholar] [CrossRef]
- De La Fuente, M.; González, A.M.; De Ron, A.M.; Santalla, M. Patterns of genetic diversity in the Andean gene pool of common bean reveal a candidate domestication gene. Mol. Breed. 2013, 31, 501–516. [Google Scholar] [CrossRef]
- Yeken, M.Z.; Kantar, F.; Çancı, H.; Özer, G.; Çiftçi, V. Breeding of Dry Bean Cultivars Using Phaseolus vulgaris Landraces in Turkey. Int. J. Agric. Wild. Sci. 2018, 4, 45–54. [Google Scholar] [CrossRef][Green Version]
- Oswaldo, V. Variedades de Fríjol en América Latina y su Origen; Centro Internacional de Agricultura Tropical (CIAT): Cali, Colombia, 1983. [Google Scholar]
- Singh, S.P.; Gepts, P.; Debouck, D.G. Races of common bean (Phaseolus vulgaris, Fabaceae). Econ. Bot. 1991, 45, 379–396. [Google Scholar] [CrossRef]
- Sinkovič, L.; Pipan, B.; Sinkovič, E.; Meglič, V. Morphological Seed Characterization of Common (Phaseolus vulgaris L.) and Runner (Phaseolus coccineus L.) Bean Germplasm: A Slovenian Gene Bank Example. Biomed. Res. Int. 2019, 2019, 6376948. [Google Scholar] [CrossRef]
- Sharma, B.D.; Rana, J.C. Plant Genetic Resources of Western Himalayas: Status and Propospects; M/s Bishen Singh Mahendra Pal Singh: Dehrdun, India, 2005. [Google Scholar]
- Balkaya, A.; Ergün, A. Diversity and use of pinto bean (Phaseolus vulgaris) populations from Samsun, Turkey. N. Z. J. Crop Hortic. Sci. 2008, 36, 189–197. [Google Scholar] [CrossRef]
- Ozer, S.; Karaköy, T.; Toklu, F.; Baloch, F.S.; Kilian, B.; Özkan, H. Nutritional and physicochemical variation in Turkish kabuli chickpea (Cicer arietinum L.) landraces. Euphytica 2010, 175, 237–249. [Google Scholar] [CrossRef]
- Krasteva, L.; Apostolova, E.; Dimova, D.; Svetleva, D. Correlation dependences between twenty-one traits of some Bulgarian common bean genotypes. In Proceedings of the IV Balkan Symposium on Vegetables and Potatoes, Plovdiv, Bulgaria, 9–12 September 2008; pp. 191–198. [Google Scholar]
- Blair, M.W.; Giraldo, M.C.; Buendia, H.F.; Tovar, E.; Duque, M.C.; Beebe, S.E. Microsatellite marker diversity in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2006, 113, 100–109. [Google Scholar] [CrossRef]
- Sharma, M.K.; Mishra, S.; Rana, N.S. Genetic divergence in French bean (Phaseolus vulgaris L.) pole type cultivars. Legume Res. 2009, 32, 220–223. [Google Scholar]
- Yeken, M.Z.; Nadeem, M.A.; Karaköy, T.; Baloch, F.S.; Çiftçi, V. Determination of Turkish Common Bean Germplasm for Morpho-agronomic and Mineral Variations for Breeding Perspectives in Turkey. KSU J. Agric. Nat. 2019, 22, 38–50. [Google Scholar]
- Ceolin, C.G.; Celeste Gonçalves-Vidigal, M.; Soares Vidigal Filho, P.; Vinícius Kvitschal, M.; Gonela, A.; Alberto Scapim, C. Genetic divergence of the common bean (Phaseolus vulgaris L.) group Carioca using morpho-agronomic traits by multivariate analysis. Hereditas 2007, 144, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Sharma, M.K.; Singh, M.; Yadav, S.K. Genetic diversity of French bean (Bush type) genotypes in North-West Himalayas. Indian J. Plant Genet. Res. 2010, 23, 285–287. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Source of Variation | F Value |
|---|---|---|
| Days to Emergence | Genotypes | 0.394 |
| G × E | 0.261 | |
| Days to 1st Flower | Genotypes | 3.07 ** |
| G × E | 1.34 | |
| Days to 50% Flowering | Genotypes | 1.88 * |
| G × E | 2.13 * | |
| 1st Pod Height | Genotypes | 12.99 **** |
| G × E | 1.89 * | |
| Primary Branches/Plant | Genotypes | 3.82 *** |
| G × E | 1.54 | |
| Secondary Branches/Plant | Genotypes | 2.51 ** |
| G × E | 2.47 ** | |
| Plant Height | Genotypes | 10.07 **** |
| G × E | 0.90 | |
| Days to Maturity | Genotypes | 189.81 *** |
| G × E | 17.63 *** | |
| Plant Weight | Genotypes | 11.55 **** |
| G × E | 3.85 ** | |
| Root Length | Genotypes | 0.82 |
| G × E | 0.53 | |
| Pods/Plant | Genotypes | 3.79 *** |
| G × E | 1.31 | |
| Pod Weight/Plant | Genotypes | 9.06 **** |
| G × E | 3.38 | |
| Pod Length | Genotypes | 8.48 **** |
| G × E | 7.31 **** | |
| Pod Width | Genotypes | 5.28 **** |
| G × E | 5.36 **** | |
| Single Pod Weight | Genotypes | 8.69 **** |
| G × E | 8.07 **** | |
| Seeds/Pod | Genotypes | 3.68 *** |
| G × E | 1.75 * | |
| Seed Length | Genotypes | 13.72 **** |
| G × E | 5.63 **** | |
| Seed Width | Genotypes | 6.79 **** |
| G × E | 3.75 *** | |
| Seed Height | Genotypes | 13.61 **** |
| G × E | 3.41 *** | |
| Seeds/Plant | Genotypes | 4.06 *** |
| G × E | 2.42 * | |
| Yield Per Plant | Genotypes | 9.76 **** |
| G × E | 3.55 ** | |
| 100 Seeds/Weight | Genotypes | 8.42 **** |
| G × E | 0.93 |
| Traits | varE | varU | h2 |
|---|---|---|---|
| DE | 0.678 | 0.012 | 0.052 |
| DF | 0.354 | 0.450 | 0.792 |
| DFtF | 0.230 | 0.298 | 0.795 |
| FPdH | 0.207 | 0.751 | 0.916 |
| PB | 0.525 | 0.426 | 0.709 |
| SB | 0.801 | 0.154 | 0.365 |
| PH | 0.105 | 0.808 | 0.959 |
| DMt | 0.102 | 0.849 | 0.943 |
| PW | 0.354 | 0.587 | 0.833 |
| RL | 0.821 | 0.076 | 0.218 |
| PdPP | 0.505 | 0.454 | 0.729 |
| PdWt | 0.386 | 0.547 | 0.809 |
| PdL | 0.165 | 0.451 | 0.891 |
| PdWd | 0.195 | 0.389 | 0.857 |
| SPdW | 0.314 | 0.385 | 0.786 |
| SPSPd | 0.436 | 0.428 | 0.747 |
| SL | 0.199 | 0.609 | 0.902 |
| SWd | 0.105 | 0.554 | 0.940 |
| SH | 0.293 | 0.659 | 0.871 |
| SPPt | 0.640 | 0.330 | 0.608 |
| YPP | 0.373 | 0.559 | 0.818 |
| 100 SW | 0.278 | 0.723 | 0.886 |
| Traits | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|
| DE | 10 | 15 | 13 | 1.09 |
| DF | 51 | 75 | 61 | 4.39 |
| DFtF | 59 | 80 | 67 | 4.61 |
| FPdH | 6.4 | 68.9 | 23 | 13.53 |
| PB | 1.6 | 5.3 | 3.6 | 0.84 |
| SB | 0.7 | 5.4 | 2.9 | 1.00 |
| PH | 21.0 | 168.76 | 99.55 | 43.65 |
| DMt | 99 | 161 | 133 | 14.64 |
| PW | 22.4 | 137.5 | 58.6 | 21.93 |
| RL | 2.6 | 9.6 | 5.2 | 1.27 |
| PdPP | 7.01 | 42.7 | 19.5 | 6.71 |
| PdWt | 14.30 | 84.80 | 37.5 | 14.62 |
| PdL | 8.3 | 19.7 | 11.4 | 1.97 |
| PdWd | 5.9 | 14.01 | 9.3 | 1.39 |
| SPdW | 0.9 | 3.7 | 2.0 | 0.53 |
| SPSPd | 1.8 | 8.2 | 3.9 | 0.98 |
| SL | 7.4 | 16.4 | 12.5 | 1.79 |
| SWd | 5.3 | 9.7 | 7.1 | 0.91 |
| SH | 4.02 | 10.9 | 5.7 | 0.88 |
| SPPt | 17.81 | 254.5 | 60.1 | 31.00 |
| YPP | 5.9 | 59.5 | 22.02 | 9.86 |
| 100 SW | 24.97 | 73.88 | 42.21 | 9.04 |
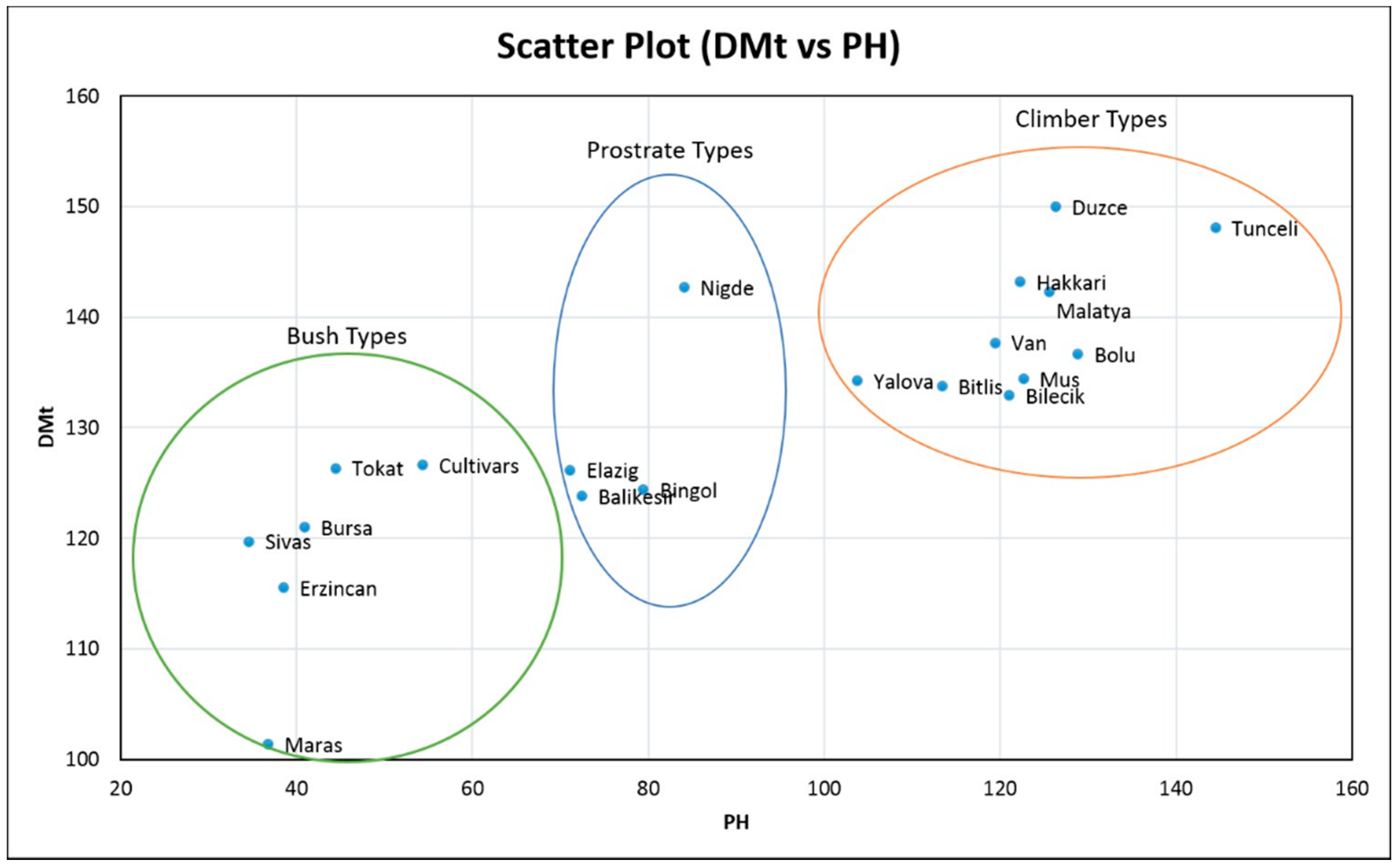
| Provinces | Number of Accessions | DE | DF | DFtF | FPdH | PB | SB | PH | DMt | PW | RL | PdPP |
| Bingol | 18 | 11.92 ± 1.09 | 58.34 ± 2.56 | 64.10 ± 3.56 | 16.04 ± 8.32 | 3.97 ± 0.75 | 2.41 ± 1.52 | 79.44 ± 43.94 | 124.3 ± 10.18 | 63.91 ± 20.28 | 5.75 ± 1.37 | 20.83 ± 8.11 |
| Hakkari | 22 | 12.73 ± 0.95 | 63.366 ± 4.15 | 69.872 ± 5.28 | 287478 ± 9.65 | 3.466 ± 0.81 | 2.26 ± 0.47 | 122.27 ± 17.90 | 139.586 ± 10.27 | 67.35 ± 26.02 | 5.690 ± 1.30 | 18.05 ± 7.87 |
| Tokat | 1 | 13.54 ± 3.22 | 61.85 ± 6.59 | 66.59 ± 7.15 | 14.09 ± 6.09 | 4.32 ± 0.40 | 3.44 ± 0.89 | 44.42 ± 6.95 | 126.3 ± 4.93 | 35.22 ± 7.01 | 3.99 ± 1.25 | 9.18 ± 8.80 |
| Maras | 1 | 13.21 ± 3.10 | 55.85 ± 6.03 | 60.26 ± 5.60 | 10.83 ± 4.55 | 3.89 ± 0.48 | 4.81 ± 2.40 | 36.75 ± 8.08 | 101.3 ± 15.94 | 28.76 ± 5.33 | 6.62 ± 2.03 | 12.81 ± 11.13 |
| Bitlis | 30 | 12.06 ± 0.72 | 61.43 ± 3.57 | 66.81 ± 3.70 | 22.88 ± 9.72 | 3.24 ± 0.75 | 2.62 ± 0.89 | 113.37 ± 34.44 | 133.7 ± 8.42 | 66.07 ± 21.67 | 4.87 ± 1.18 | 22.45 ± 7.65 |
| Malatya | 14 | 11.18 ± 0.54 | 63.01 ± 4.94 | 68.90 ± 5.008 | 31.29 ± 16.77 | 3.33 ± 0.77 | 3.21 ± 0.91 | 125.59 ± 40.22 | 142.3 ± 15.58 | 63.24 ± 21.98 | 5.57 ± 0.63 | 18.84 ± 5.38 |
| Tunceli | 3 | 11.07 ± 0.38 | 64.25 ± 2.85 | 68.23 ± 3.46 | 32.13 ± 16.63 | 2.77 ± 0.35 | 2.60 ± 0.80 | 144.48 ± 15.34 | 148.1 ± 12.03 | 64.77 ± 4.53 | 5.85 ± 0.33 | 18.03 ± 2.48 |
| Van | 17 | 12.74 ± 1.53 | 64.60 ± 4.19 | 69.70 ± 4.26 | 29.52 ± 14.22 | 3.43 ± 0.94 | 3.06 ± 0.98 | 119.41 ± 35.28 | 137.4 ± 5.14 | 66.93 ± 22.19 | 4.96 ± 1.64 | 20.13 ± 4.73 |
| Elazig | 13 | 13.18 ± 0.54 | 59.52 ± 4.59 | 64.52 ± 4.84 | 14.34 ± 8.66 | 4.06 ± 0.45 | 3.27 ± 0.69 | 71.10 ± 46.87 | 126.1 ± 16.59 | 47.44 ± 18.17 | 4.50 ± 0.92 | 19.61 ± 8.21 |
| Mus | 22 | 13.43 ± 0.55 | 59.25 ± 3.74 | 65.79 ± 2.83 | 25.28 ± 13.90 | 3.25 ± 0.80 | 2.79 ± 0.82 | 122.63 ± 25.61 | 134.4 ± 10.42 | 53.36 ± 14.84 | 4.63 ± 0.71 | 20.59 ± 4.54 |
| Sivas | 12 | 13.24 ± 0.61 | 57.05 ± 1.98 | 62.92 ± 2.01 | 10.53 ± 2.88 | 4.28 ± 0.68 | 3.16 ± 1.040 | 34.60 ± 7.20 | 119.7 ± 11.31 | 40.48 ± 11.45 | 5.58 ± 1.109 | 14.69 ± 3.35 |
| Alikesir | 6 | 12.97 ± 0.68 | 57.35 ± 2.36 | 65.11 ± 5.85 | 18.42 ± 11.06 | 3.59 ± 0.98 | 3.17 ± 1.09 | 72.43 ± 47.91 | 123.8 ± 15.65 | 51.40 ± 17.21 | 4.75 ± 1.94 | 17.15 ± 8.46 |
| Bilecik | 7 | 13.37 ± 0.38 | 61.37 ± 4.19 | 67.65 ± 4.90 | 41.11 ± 2.01 | 3.09 ± 0.54 | 2,13 ± 0.05 | 121.03 ± 6.69 | 134.4 ± 13.9 | 79.47 ± 34,04 | 4.10 ± 1.47 | 24.83 ± 5.27 |
| Duzce | 2 | 12.13 ± 0.23 | 68.42 ± 6.36 | 72.58 ± 2.35 | 39.88 ± 22.22 | 2.95 ± 1.64 | 2.94 ± 0.30 | 126.30 ± 2.52 | 150.0 ± 8.95 | 42.39 ± 13.35 | 4.40 ± 1.32 | 14.80 ± 3.18 |
| Yalova | 3 | 12.63 ± 0.33 | 62.36 ± 5.66 | 69.03 ± 6.18 | 32.31 ± 17.76 | 2.76 ± 1.13 | 3.51 ± 0.64 | 103.74 ± 52.57 | 134.2 ± 11.40 | 57.80 ± 15.06 | 5.08 ± 0.15 | 20.67 ± 1.16 |
| Erzincan | 4 | 12.55 ± 0.32 | 56.92 ± 2.32 | 63.42 ± 3.55 | 11.83 ± 2.55 | 4.33 ± 0.49 | 4.57 ± 0.68 | 38.49 ± 5.91 | 115.5 ± 3.59 | 38.74 ± 11.83 | 6.83 ± 0.70 | 15.14 ± 2.99 |
| Bursa | 2 | 12.47 ± 1.65 | 56.42 ± 1.17 | 62.75 ± 0.23 | 12.12 ± 2.29 | 4.10 ± 0.20 | 3.84 ± 0.44 | 40.90 ± 1.71 | 121.0 ± 6.60 | 39.34 ± 4.72 | 5.87 ± 0.79 | 15.05 ± 0.37 |
| Nigde | 2 | 11.97 ± 0.94 | 62.42 ± 0.24 | 67.08 ± 0.23 | 15.15 ± 0.12 | 3.36 ± 0.57 | 3.26 ± 0.19 | 84.04 ± 4.55 | 142.6 ± 2.35 | 44.09 ± 18.12 | 5.44 ± 0.29 | 20.62 ± 6.03 |
| Bolu | 1 | 12.63 ± 2.41 | 63.58 ± 4.71 | 67.92 ± 8.50 | 25.91 ± 2.9 | 2.86 ± 0.99 | 3.23 ± 1.07 | 128.85 ± 22.3 | 128.4 ± 8.41 | 47.3 ± 17.4 | 4.81 ± 2.17 | 14.98 ± 4.87 |
| Cultivars | 3 | 10.55 ± 0.57 | 60.82 ± 2.61 | 66.04 ± 2.59 | 13.25 ± 0.41 | 4.33 ± 0.30 | 3.82 ± 0.44 | 54.33 ± 13.0 | 126.6 ± 4.30 | 40.48 ± 6.26 | 5.98 ± 0.86 | 19.86 ± 3.11 |
| Provinces | PdWt | PdL | PdWd | SPdW | SPSPd | SL | SWd | SH | SPPt | TSW | 100Swt | |
| Bingol | 18 | 42.72 ± 14.80 | 10.23 ± 1.01 | 9.19 ± 0.71 | 2.18 ± 0.56 | 3.89 ± 0.92 | 12.25 ± 2.10 | 6.72 ± 0.68 | 5.47 ± 0.59 | 64.74 ± 53.57 | 22.47 ± 9.08 | 46.93 ± 11.61 |
| Hakkari | 22 | 42.42 ± 14.3 | 12.43 ± 2.71 | 10.39 ± 1.73 | 2.01 ± 0.62 | 3.99 ± 1.16 | 12.54 ± 1.71 | 7.09 ± 0.85 | 5.51 ± 0.76 | 63.40 ± 28.25 | 23.65 ± 9.98 | 41.16 ± 11.62 |
| Tokat | 1 | 25.01 ± 9.23 | 14.79 ± 0.70 | 12.62 ± 2.18 | 2.49 ± 0.67 | 3.07 ± 0.32 | 15.73 ± 1.25 | 8.44 ± 0.99 | 7.90 ± 0.53 | 25.41 ± 11.04 | 17.32 ± 4.23 | 51.78 ± 2.50 |
| Maras | 1 | 16.81 ± 3.97 | 9.17 ± 1.96 | 8.75 ± 2.25 | 1.69 ± 0.12 | 3.50 ± 0.63 | 12.71 ± 0.85 | 7.09 ± 1.16 | 5.87 ± 0.26 | 33.78 ± 16.67 | 9.83 ± 6.43 | 48.21 ± 12.91 |
| Bitlis | 30 | 44.24 ± 15.60 | 10.85 ± 1.46 | 9.07 ± 1.35 | 2.31 ± 0.60 | 4.27 ± 1.11 | 12.17 ± 1.67 | 6.99 ± 0.90 | 5.57 ± 0.70 | 70.44 ± 30.75 | 25.06 ± 11.64 | 40.99 ± 6.91 |
| Malatya | 14 | 38.49 ± 14.09 | 12.45 ± 2.83 | 9.33 ± 1.75 | 1.98 ± 0.67 | 4.42 ± 0.70 | 12.28 ± 1.30 | 7.15 ± 1.28 | 5.96 ± 1.29 | 66.30 ± 34.92 | 22.80 ± 9.75 | 38.42 ± 6.56 |
| Tunceli | 3 | 42.31 ± 2.35 | 11.16 ± 1.15 | 11.46 ± 1.08 | 2.28 ± 0.37 | 4.07 ± 0.39 | 12.94 ± 1.35 | 8.08 ± 0.54 | 6.80 ± 0.33 | 55.29 ± 9.53 | 25.20 ± 2.42 | 44.28 ± 1.74 |
| Van | 17 | 43.32 ± 13.53 | 13.15 ± 2.14 | 9.53 ± 1.34 | 1.93 ± 0.48 | 3.84 ± 1.27 | 13.11 ± 1.70 | 7.18 ± 0.97 | 5.26 ± 0.72 | 63.32 ± 20.86 | 26.79 ± 9.39 | 43.564 ± 10.59 |
| Elazig | 13 | 31.17 ± 14.24 | 11.43 ± 1.24 | 8.03 ± 0.55 | 1.76 ± 0.21 | 3.49 ± 0.58 | 12.03 ± 1.13 | 6.24 ± 0.73 | 5.23 ± 0.69 | 59.69 ± 24.009 | 21.240 ± 10.58 | 37.02 ± 5.56 |
| Mus | 22 | 34.35 ± 7.81 | 11.18 ± 1.65 | 9.12 ± 1.005 | 1.85 ± 0.39 | 3.95 ± 0.82 | 12.38 ± 1.73 | 7.50 ± 0.82 | 5.89 ± 0.66 | 64.66 ± 19.38 | 22.50 ± 6.03 | 40.12 ± 7.85 |
| Sivas | 12 | 23.69 ± 5.69 | 10.72 ± 1.378 | 10.13 ± 1.15 | 1.79 ± 0.28 | 2.98 ± 0.56 | 12.57 ± 2.002 | 7.14 ± 0.75 | 6.80 ± 1.28 | 28.89 ± 5.72 | 12.17 ± 3.006 | 42.79 ± 8.14 |
| Balikesir | 6 | 29.75 ± 14.63 | 10.83 ± 1.28 | 8.71 ± 0.59 | 1.78 ± 0.19 | 3.64 ± 0.72 | 12.83 ± 2.10 | 6.99 ± 0.79 | 5.95 ± 0.37 | 43.56 ± 30.87 | 15.58 ± 8.46 | 43.52 ± 7.14 |
| Bilecik | 7 | 59.74 ± 19.65 | 10.38 ± 0.84 | 9.15 ± 1.43 | 2.03 ± 0.67 | 4.27 ± 1.21 | 11.65 ± 2.22 | 7.38 ± 1.10 | 6.15 ± 0.61 | 77.22 ± 38.79 | 31.06 ± 15.28 | 39.10 ± 8.33 |
| Duzce | 2 | 25.21 ± 8.64 | 12.99 ± 2.53 | 9.10 ± 0.60 | 2.60 ± 0.21 | 5.37 ± 0.29 | 12.31 ± 1.31 | 7.65 ± 0.55 | 5.90 ± 0.47 | 51.74 ± 15.28 | 18.52 ± 6.61 | 47.34 ± 1.08 |
| Yalova | 3 | 38.39 ± 11.51 | 11.97 ± 1.47 | 8.50 ± 0.91 | 2.17 ± 0.02 | 4.49 ± 1.004 | 12.66 ± 2.82 | 7.30 ± 0.15 | 6.42 ± 0.31 | 63.54 ± 29.35 | 21.41 ± 9.36 | 46.68 ± 8.96 |
| Erzincan | 4 | 20.68 ± 5.39 | 11.21 ± 0.38 | 8.15 ± 0.46 | 1.66 ± 0.24 | 3.47 ± 0.31 | 13.67 ± 0.91 | 6.44 ± 0.29 | 5.41 ± 0.32 | 35.13 ± 6.63 | 12.67 ± 2.82 | 48.25 ± 5.73 |
| Bursa | 2 | 22.38 ± 2.65 | 11.005 ± 0.27 | 8.27 ± 0.65 | 1.65 ± 0.24 | 3.62 ± 0.40 | 13.94 ± 0.67 | 6.58 ± 0.04 | 5.54 ± 0.01 | 38.97 ± 3.27 | 13.86 ± 2.34 | 47.95 ± 2.02 |
| Nigde | 2 | 33.10 ± 13.29 | 11.66 ± 0.59 | 9.97 ± 1.20 | 1.66 ± 0.25 | 3.39 ± 0.60 | 14.13 ± 3.02 | 7.91 ± 1.08 | 5.06 ± 0.25 | 52.83 ± 10.46 | 22.02 ± 10.84 | 54.69 ± 17.59 |
| Bolu | 1 | 27.99 ± 9.96 | 11.06 ± 1.19 | 8.64 ± 2.85 | 1.81 ± 0.69 | 4.04 ± 0.32 | 10.08 ± 2.15 | 7.31 ± 1.79 | 6.99 ± 0.85 | 53.26 ± 14.59 | 17.70 ± 4.79 | 59.89 ± 12.86 |
| Cultivars | 3 | 25.33 ± 6.02 | 9.93 ± 0.87 | 8.14 ± 0.04 | 1.34 ± 0.09 | 3.78 ± 1.02 | 10.20 ± 3.15 | 6.11 ± 0.69 | 5.17 ± 0.53 | 58.36 ± 32.90 | 15.63 ± 3.22 | 38.71 ± 5.44 |
| DE | DF | DFtF | FPdH | PB | SB | PH | DMt | PW | RL | PdPP | PdWt | PdL | PdWd | SPdW | SPSPd | SL | SWd | SH | SPPt | YPP | 100 SWt | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DE | 1 | −0.153 * | −0.133 | −0.031 | 0.028 | 0.062 | −0.152 * | −0.191 ** | −0.206 ** | −0.161 * | −0.174 * | −0.199 ** | −0.01 | 0.059 | −0.062 | −0.217 ** | 0.114 | 0.165 * | 0.106 | −0.207 ** | −0.109 | 0.071 |
| DF | 1 | 0.883 ** | 0.672 ** | −0.476 ** | −0.265 ** | 0.652 ** | 0.646 ** | 0.456 ** | −0.071 | 0.166 * | 0.411 ** | 0.441 ** | 0.271 ** | 0.285 ** | 0.366 ** | −0.177 * | 0.182 * | 0.064 | 0.330 ** | 0.425 ** | −0.073 | |
| DFtF | 1 | 0.716 ** | −0.505 ** | −0.274 ** | 0.688 ** | 0.682 ** | 0.465 ** | −0.061 | 0.191 ** | 0.406 ** | 0.388 ** | 0.239 ** | 0.262 ** | 0.382 ** | −0.181 * | 0.233 ** | 0.112 | 0.331 ** | 0.423 ** | −0.067 | ||
| FPdH | 1 | −0.619 ** | −0.314 ** | 0.691 ** | 0.640 ** | 0.533 ** | −0.033 | 0.114 | 0.398 ** | 0.328 ** | 0.241 ** | 0.388 ** | 0.427 ** | −0.187 * | 0.404 ** | 0.264 ** | 0.322 ** | 0.452 ** | −0.001 | |||
| PB | 1 | 0.523 ** | −0.649 ** | −0.531 ** | −0.385 ** | 0.180 * | −0.111 | −0.311 ** | −0.169 * | −0.161 * | −0.423 ** | −0.406 ** | 0.244 ** | −0.374 ** | −0.234 ** | −0.253 ** | −0.366 ** | 0.027 | ||||
| SB | 1 | −0.570 ** | −0.390 ** | −0.377 ** | 0.296 ** | −0.254 ** | −0.380 ** | −0.076 | −0.235 ** | −0.286 ** | −0.269 ** | 0.243 ** | −0.174 * | 0.003 | −0.284 ** | −0.367 ** | 0.099 | |||||
| PH | 1 | 0.744 ** | 0.611 ** | −0.236 ** | 0.400 ** | 0.583 ** | 0.276 ** | 0.259 ** | 0.324 ** | 0.491 ** | −0.266 ** | 0.350 ** | 0.039 | 0.500 ** | 0.603 ** | −0.145 * | ||||||
| DMt | 1 | 0.494 ** | −0.114 | 0.242 ** | 0.447 ** | 0.188 * | 0.336 ** | 0.207 ** | 0.289 ** | −0.233 ** | 0.399 ** | 0.115 | 0.354 ** | 0.481 ** | 0.012 | |||||||
| PW | 1 | 0.057 | 0.521 ** | 0.888 ** | 0.190 * | 0.291 ** | 0.446 ** | 0.407 ** | −0.185 * | 0.301 ** | 0.161 * | 0.584 ** | 0.790 ** | 0.048 | ||||||||
| RL | 1 | −0.274 ** | −0.074 | 0.112 | 0.065 | 0.043 | −0.097 | 0.084 | −0.100 | 0.011 | −0.129 | −0.169 * | 0.126 | |||||||||
| PdPP | 1 | 0.668 ** | −0.228 ** | −0.094 | 0.019 | 0.332 ** | −0.358 ** | −0.051 | −0.183 * | 0.633 ** | 0.607 ** | −0.282 ** | ||||||||||
| PdWt | 1 | 0.109 | 0.287 ** | 0.464 ** | 0.473 ** | −0.183 * | 0.292 ** | 0.043 | 0.667 ** | 0.872 ** | 0.005 | |||||||||||
| PdL | 1 | 0.265 ** | 0.218 ** | 0.139 | 0.358 ** | 0.040 | −0.077 | 0.001 | 0.137 | 0.128 | ||||||||||||
| PdWd | 1 | 0.283 ** | −0.051 | 0.229 ** | 0.559 ** | 0.316 ** | −0.027 | 0.241 ** | 0.291 ** | |||||||||||||
| SPdW | 1 | 0.555 ** | 0.017 | 0.443 ** | 0.324 ** | 0.257 ** | 0.484 ** | 0.239 ** | ||||||||||||||
| SPSPd | 1 | −0.425 ** | 0.084 | −0.008 | 0.575 ** | 0.546 ** | −0.304 ** | |||||||||||||||
| SL | 1 | 0.203 ** | 0.020 | −0.412 ** | −0.181 * | 0.579 ** | ||||||||||||||||
| SWd | 1 | 0.540 ** | −0.007 | 0.340 ** | 0.446 ** | |||||||||||||||||
| SH | 1 | −0.125 | 0.086 | 0.271 ** | ||||||||||||||||||
| SPPt | 1 | 0.750 ** | −0.243 ** | |||||||||||||||||||
| YPP | 1 | 0.001 | ||||||||||||||||||||
| 100 SWt | 1 |
| Accessions | Traits | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DE | DF | DFtF | FPdH | PB | SB | PH | DMt | PW | RL | PdPP | PdWt | PdL | PdWd | SPdW | SPSPd | SL | SWd | SH | SPPt | YPP | 100SWt | |
| Bingol-53 | 13 | 57 | 61 | 11.6 | 5 | 5 | 25.2 | 113 | 84 | 8.7 | 18.5 | 52 | 10.5 | 2.5 | 9.3 | 3 | 15.3 | 6.3 | 5.9 | 39.3 | 17.7 | 67.1 |
| Bingol-65 | 12 | 56 | 61 | 11.8 | 4 | 4 | 41.2 | 111 | 53.1 | 7.7 | 15.2 | 37.7 | 10.6 | 2 | 8.7 | 2.6 | 14.4 | 6.5 | 5.5 | 32.4 | 17.5 | 57.5 |
| Bitlis-53 | 12 | 57 | 62 | 16 | 4 | 5 | 38.8 | 127 | 47.3 | 9.6 | 14.9 | 26.2 | 10 | 2.3 | 7.7 | 2.8 | 14.4 | 6.5 | 5.4 | 37.1 | 14.2 | 54.4 |
| Van-47 | 14 | 55 | 62 | 12.9 | 4 | 4 | 48.8 | 109 | 37.8 | 8.9 | 14.6 | 23.5 | 10.2 | 2 | 7.5 | 3.3 | 13.4 | 6.1 | 5.4 | 36.7 | 13.8 | 43.6 |
| Elazig-30 | 13 | 57 | 60 | 10 | 5 | 4 | 36.5 | 119 | 43.1 | 4.9 | 16.8 | 31.8 | 11.3 | 2 | 8.1 | 3.3 | 13.6 | 6.2 | 5.5 | 72.2 | 17.1 | 40.7 |
| Sivas-18 | 13 | 59 | 65 | 13.7 | 3 | 2 | 52.1 | 129 | 70.8 | 4.2 | 16 | 39.8 | 12.9 | 2.2 | 10.3 | 2.3 | 15.7 | 8.1 | 6.4 | 34.1 | 20.1 | 59 |
| Sivas-62 | 12 | 56 | 63 | 8 | 5 | 3 | 33.5 | 107 | 33.8 | 6.4 | 13.7 | 21.5 | 11.4 | 1.5 | 8.8 | 1.9 | 14.1 | 7.2 | 6.5 | 23.7 | 12.1 | 52 |
| Sivas-69 | 13 | 55 | 61 | 10 | 4 | 2 | 29.1 | 107 | 39 | 6.2 | 9.8 | 22.5 | 11.6 | 1.9 | 9.1 | 2.7 | 13.9 | 7.2 | 6.4 | 23.8 | 12.5 | 49.1 |
| Balikesir-18 | 14 | 56 | 62 | 11 | 5 | 2 | 38.6 | 109 | 37 | 5.4 | 15.8 | 21.6 | 10.4 | 1.7 | 8.3 | 3 | 14.9 | 6.6 | 5.7 | 27.5 | 10 | 48.1 |
| Erzincan-3 | 12. | 59 | 68 | 15.5 | 5 | 5 | 45.1 | 119 | 55.3 | 7.3 | 14.9 | 26.5 | 11.3 | 1.7 | 8.5 | 3.6 | 12.9 | 6.7 | 5.3 | 39.5 | 16.1 | 55.6 |
| Erzincan-5 | 12. | 55 | 61 | 10.1 | 4 | 4 | 40.3 | 110 | 37.3 | 5.8 | 16.5 | 23.5 | 10.8 | 1.8 | 7.6 | 3.2 | 12.9 | 6 | 5 | 34.8 | 12.8 | 44.3 |
| Bursa-1 | 14 | 55 | 63 | 13.7 | 4 | 4 | 39.6 | 116 | 42.6 | 6.4 | 15.3 | 24.2 | 11.2 | 1.8 | 7.8 | 3.9 | 14.4 | 6.6 | 5.5 | 41.2 | 15.5 | 49.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadeem, M.A.; Karaköy, T.; Yeken, M.Z.; Habyarimana, E.; Hatipoğlu, R.; Çiftçi, V.; Nawaz, M.A.; Sönmez, F.; Shahid, M.Q.; Yang, S.H.; et al. Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes. Agronomy 2020, 10, 272. https://doi.org/10.3390/agronomy10020272
Nadeem MA, Karaköy T, Yeken MZ, Habyarimana E, Hatipoğlu R, Çiftçi V, Nawaz MA, Sönmez F, Shahid MQ, Yang SH, et al. Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes. Agronomy. 2020; 10(2):272. https://doi.org/10.3390/agronomy10020272
Chicago/Turabian StyleNadeem, Muhammad Azhar, Tolga Karaköy, Mehmet Zahit Yeken, Ephrem Habyarimana, Ruştu Hatipoğlu, Vahdettin Çiftçi, Muhammad Amjad Nawaz, Ferit Sönmez, Muhammad Qasim Shahid, Seung Hwan Yang, and et al. 2020. "Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes" Agronomy 10, no. 2: 272. https://doi.org/10.3390/agronomy10020272
APA StyleNadeem, M. A., Karaköy, T., Yeken, M. Z., Habyarimana, E., Hatipoğlu, R., Çiftçi, V., Nawaz, M. A., Sönmez, F., Shahid, M. Q., Yang, S. H., Chung, G., & Baloch, F. S. (2020). Phenotypic Characterization of 183 Turkish Common Bean Accessions for Agronomic, Trading, and Consumer-Preferred Plant Characteristics for Breeding Purposes. Agronomy, 10(2), 272. https://doi.org/10.3390/agronomy10020272