
Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer

 ,
,  ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Analysis of Diffuse- and Intestinal-Type GC
2.2. Network Analysis
2.3. Data Visualization
2.4. Statistical Analysis
3. Results
3.1. Canonical Pathways in Diffuse- and Intestinal-Type GC
3.2. Cell Cycle-Related Canonical Pathways in Diffuse- and Intestinal-Type GC
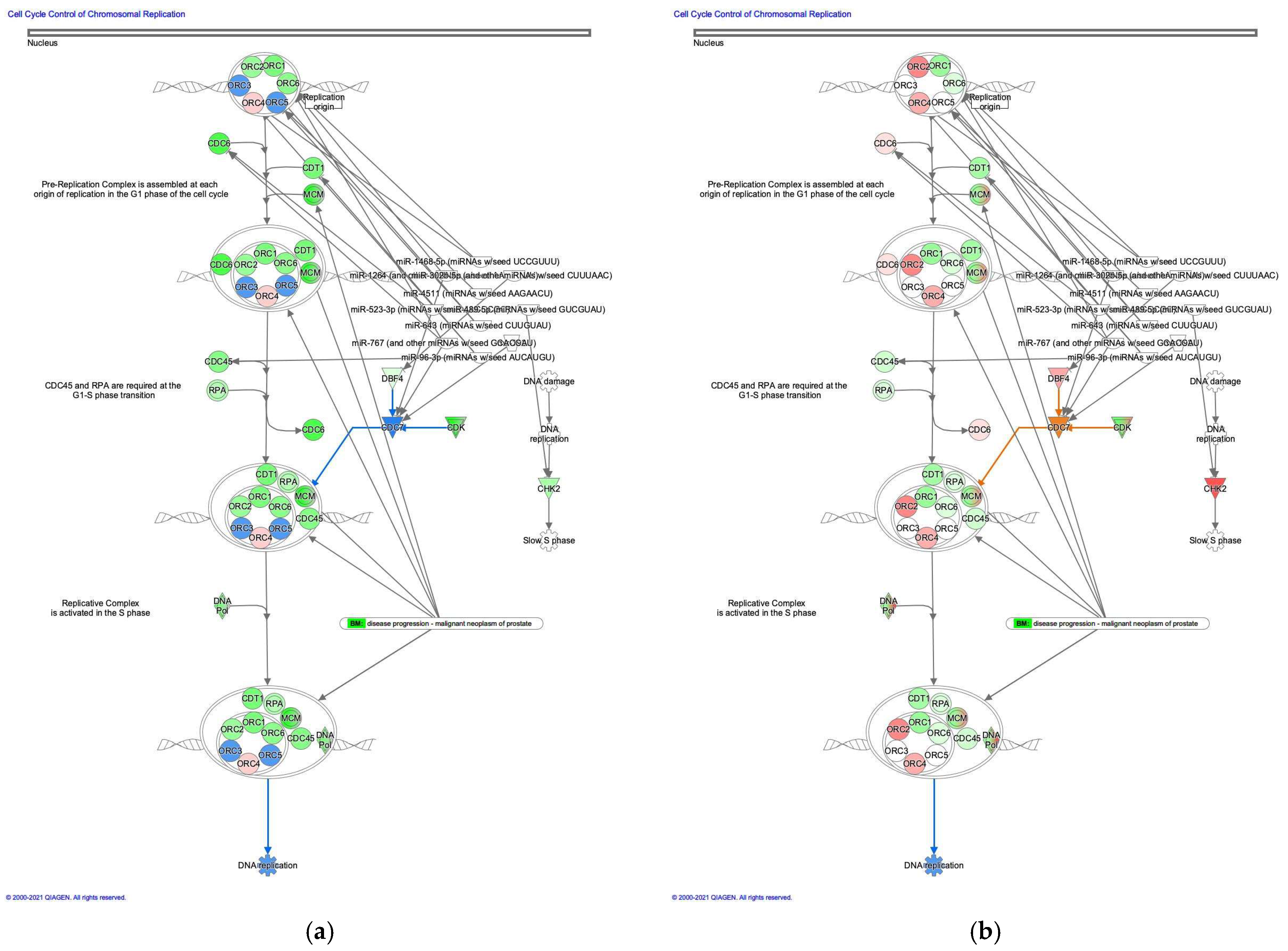
3.2.1. Cell Cycle Control of Chromosomal Replication was Activated in Intestinal-Type GC
3.2.2. The G1/S Cell Cycle Checkpoint Regulation Pathway was Activated in Diffuse-Type GC
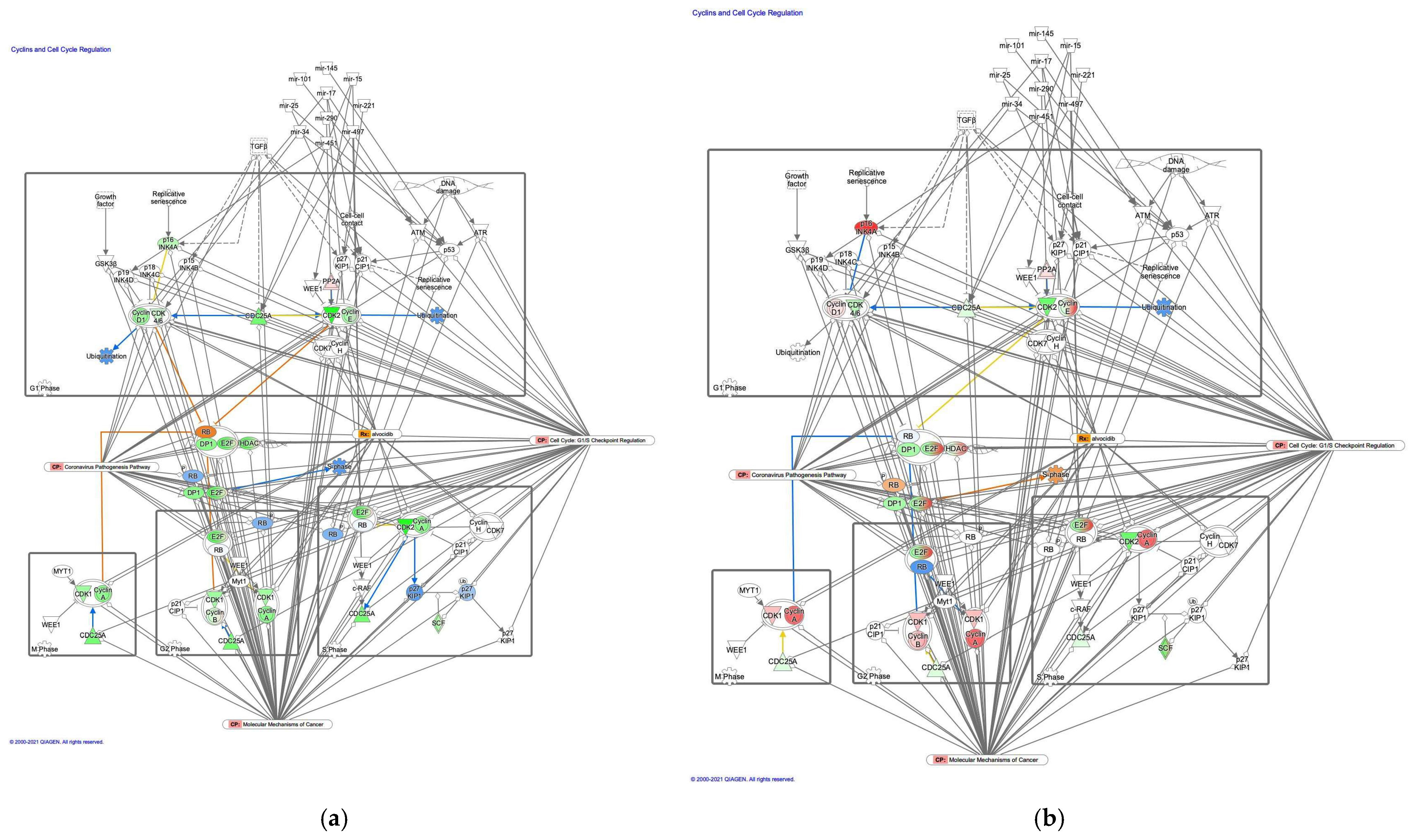
3.2.3. The Cyclin and Cell Cycle Regulation Pathway was Activated in Intestinal-Type GC
3.2.4. Canonical Pathway of the Role of BRCA1 in the DNA Damage Response was Activated in Intestinal-Type GC Compared to Diffuse-Type GC
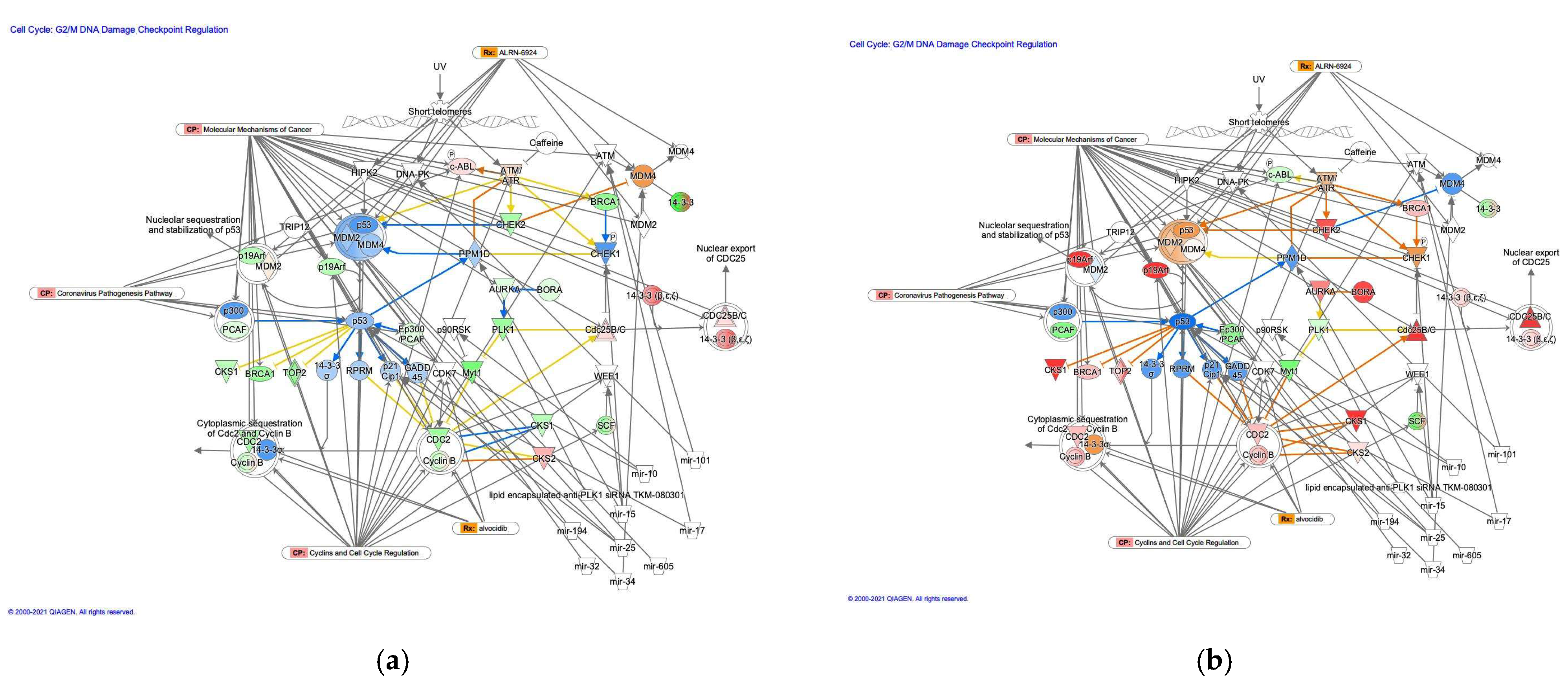
3.2.5. The G2/M DNA Damage Cell Cycle Checkpoint Regulation Pathway in Diffuse- and Intestinal-Type GC
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Yokozaki, H.; Sasaki, H. Molecular Network Profiling in Intestinal- and Diffuse-Type Gastric Cancer. Cancers 2020, 12, 3833. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Quader, S.; Cabral, H.; Ono, R. Interplay of EMT and CSC in Cancer and the Potential Therapeutic Strategies. Front. Pharmacol. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, R.; Shen, Q.; Yang, D.; Peng, H.; Tong, J.; Fu, Q. Overexpression of Circular RNA circ_0013587 Reverses Erlotinib Resistance in Pancreatic Cancer Cells Through Regulating the miR-1227/E-Cadherin Pathway. Front. Oncol. 2021, 11, 754146. [Google Scholar] [CrossRef]
- Chen, S.; Wu, H.; Zhu, L.; Jiang, M.; Wei, S.; Luo, J.; Liu, A. MiR-199b-5p Promotes Gastric Cancer Progression by Regulating HHIP Expression. Front. Oncol. 2021, 11, 728393. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Wang, W.; Zhang, L.; Huang, P. Candidate oncogene circularNOP10 mediates gastric cancer progression by regulating miR-204/SIRT1 pathway. J. Gastrointest. Oncol. 2021, 12, 1428–1443. [Google Scholar] [CrossRef]
- Tanabe, S.; Aoyagi, K.; Yokozaki, H.; Sasaki, H. Gene expression signatures for identifying diffuse-type gastric cancer associated with epithelial-mesenchymal transition. Int. J. Oncol. 2014, 44, 1955–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazăr, D.; Tăban, S.; Ardeleanu, C.; Dema, A.; Sporea, I.; Cornianu, M.; Lazăr, E.; Vernic, C. The immunohistochemical expression of E-cadherin in gastric cancer; correlations with clinicopathological factors and patients’ survival. Rom. J. Morphol. Embryol. 2008, 49, 459–467. [Google Scholar]
- Wilson, M.M.; Weinberg, R.A.; Lees, J.A.; Guen, V.J. Emerging Mechanisms by which EMT Programs Control Stemness. Trends Cancer 2020, 6, 775–780. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zheng, K.; Huang, Y.; Xiong, H.; Su, J.; Chen, R.; Zou, Y. PARP inhibitors in gastric cancer: Beacon of hope. J. Exp. Clin. Cancer. Res. 2021, 40, 211. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.D.; Rizzo, A.; Brandi, G. DNA damage response alterations in gastric cancer: Knocking down a new wall. Future Oncol. 2021, 17, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Kawabata, T.; Aoyagi, K.; Yokozaki, H.; Sasaki, H. Gene expression and pathway analysis of CTNNB1 in cancer and stem cells. World J. Stem Cells 2016, 8, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 2016, 375, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Lee, J.H.; Berger, J.M. Cell Cycle-Dependent Control and Roles of DNA Topoisomerase II. Genes 2019, 10, 859. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Zheng, H.; Zhan, Y.; Liu, S.; Yang, Y.; Zang, H.; Wen, Q.; Zhang, Y.; Fan, S. Overexpression of P4HA1 associates with poor prognosis and promotes cell proliferation and metastasis of lung adenocarcinoma. J. Cancer 2021, 12, 6685–6694. [Google Scholar] [CrossRef]
- Prakash, V.; Carson, B.B.; Feenstra, J.M.; Dass, R.A.; Sekyrova, P.; Hoshino, A.; Petersen, J.; Guo, Y.; Parks, M.M.; Kurylo, C.M.; et al. Ribosome biogenesis during cell cycle arrest fuels EMT in development and disease. Nat. Commun. 2019, 10, 2110. [Google Scholar] [CrossRef]
- Chen, B.; Ma, J.; Li, C.; Wang, Y. Long noncoding RNA KCNQ1OT1 promotes proliferation and epithelial-mesenchymal transition by regulation of SMAD4 expression in lens epithelial cells. Mol. Med. Rep. 2018, 18, 16–24. [Google Scholar] [CrossRef]
- Kaimori, A.; Potter, J.; Kaimori, J.Y.; Wang, C.; Mezey, E.; Koteish, A. Transforming growth factor-beta1 induces an epithelial-to-mesenchymal transition state in mouse hepatocytes in vitro. J. Biol. Chem. 2007, 282, 22089–22101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Baserga, R.; Chen, L.; Wang, C.; Lisanti, M.P.; Pestell, R.G. microRNA, cell cycle, and human breast cancer. Am. J. Pathol. 2010, 176, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, Y.; Zhang, Y.; Cheng, L.; Zhou, X.; Chen, K. SNHG7 accelerates cell migration and invasion through regulating miR-34a-Snail-EMT axis in gastric cancer. Cell Cycle 2020, 19, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.D.; Cheng, J.T.; Chandoo, A.; Sun, X.W.; Zhang, L.; Lu, M.D.; Sun, W.J.; Huang, Y.P. microRNA-33a prevents epithelial-mesenchymal transition, invasion, and metastasis of gastric cancer cells through the Snail/Slug pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G147–G160. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, Z.; Wang, G.H.; Zhou, Y.M.; Deng, J.P.; Feng, Y.; Chen, J.Q.; Tian, L. AURKB Promotes the Metastasis of Gastric Cancer, Possibly by Inducing EMT. Cancer Manag. Res. 2020, 12, 6947–6958. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Cao, W.; Wu, K.; Qin, X.; Wang, X.; Li, Y.; Yu, B.; Zhang, Z.; Wang, X.; Yan, M.; et al. LncRNA LINC00460 promotes EMT in head and neck squamous cell carcinoma by facilitating peroxiredoxin-1 into the nucleus. J. Exp. Clin. Cancer Res. 2019, 38, 365. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Yang, Y.; Ma, Y.; Wang, F.; Xue, A.; Zhu, J.; Yang, H.; Chen, Q.; Chen, M.; Ye, L.; et al. Potential regulatory role of lncRNA-miRNA-mRNA axis in osteosarcoma. Biomed. Pharmacother. 2020, 121, 109627. [Google Scholar] [CrossRef]
- Liu, D.; Xu, X.; Wen, J.; Xie, L.; Zhang, J.; Shen, Y.; Jiang, G.; Chen, J.; Fan, M. Integrated Genome-Wide Analysis of Gene Expression and DNA Copy Number Variations Highlights Stem Cell-Related Pathways in Small Cell Esophageal Carcinoma. Stem Cells Int. 2018, 2018, 3481783. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Wang, K.; Duan, Y.; Chen, X.; Chu, X.; Hu, P.; Xiong, B. Combined Analysis of RNA Sequence and Microarray Data Reveals a Competing Endogenous RNA Network as Novel Prognostic Markers in Malignant Pleural Mesothelioma. Front. Oncol. 2021, 11, 615234. [Google Scholar] [CrossRef]
- Fei, T.; Chen, Y.; Xiao, T.; Li, W.; Cato, L.; Zhang, P.; Cotter, M.B.; Bowden, M.; Lis, R.T.; Zhao, S.G.; et al. Genome-wide CRISPR screen identifies HNRNPL as a prostate cancer dependency regulating RNA splicing. Proc. Natl. Acad. Sci. USA 2017, 114, E5207–E5215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Roulis, M.; Kaklamanos, A.; Schernthanner, M.; Bielecki, P.; Zhao, J.; Kaffe, E.; Frommelt, L.S.; Qu, R.; Knapp, M.S.; Henriques, A.; et al. Paracrine orchestration of intestinal tumorigenesis by a mesenchymal niche. Nature 2020, 580, 524–529. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canonical Pathways | Diffuse-Type GC | Intestinal-Type GC |
|---|---|---|
| Cell Cycle Control of Chromosomal Replication | −4.811 | 0.962 |
| Kinetochore Metaphase Signaling Pathway | −2.271 | 2.92 |
| Estrogen-mediated S-phase Entry | −3.5 | 1.5 |
| Relaxin Signaling | 1.5 | −3 |
| tRNA Charging | −3 | −1.5 |
| Spliceosomal Cycle | −3.207 | 1.069 |
| Cell Cycle: G1/S Checkpoint Regulation | 2.982 | −1.147 |
| Cyclins and Cell Cycle Regulation | −3.13 | 0.894 |
| Ovarian Cancer Signaling | 0.632 | −3.162 |
| Sumoylation Pathway | 2.673 | 1.069 |
| Coronavirus Pathogenesis Pathway | 2.335 | −1.257 |
| Inhibition of ARE-Mediated mRNA Degradation Pathway | 3.13 | −0.447 |
| Aldosterone Signaling in Epithelial Cells | −2.309 | −1.155 |
| RAN Signaling | −3 | −0.333 |
| Methionine Degradation I (to Homocysteine) | −1.633 | 1.633 |
| Reelin Signaling in Neurons | −0.218 | −2.837 |
| Dolichyl-diphosphooligosaccharide Biosynthesis | −2.646 | 0.378 |
| Pentose Phosphate Pathway | −2 | −1 |
| Ferroptosis Signaling Pathway | 1.706 | −1.279 |
| Glioblastoma Multiforme Signaling | 0.243 | −2.668 |
| Cell Cycle Regulation by BTG Family Proteins | −1.732 | 1.155 |
| Small-Cell Lung Cancer Signaling | −1.732 | −1.155 |
| ATM Signaling | −1.46 | 1.043 |
| Mitotic Roles of Polo-Like Kinase | −1.5 | 1 |
| Cholesterol Biosynthesis III (via Desmosterol) | 0 | 2.449 |
| Cholesterol Biosynthesis I | 0 | 2.449 |
| Cholesterol Biosynthesis II (via 24,25-dihydrolanosterol) | 0 | 2.449 |
| Sperm Motility | 0.728 | −1.698 |
| Cell Cycle: G2/M DNA Damage Checkpoint Regulation | 0.655 | −1.528 |
| Senescence Pathway | 1.234 | −0.926 |
| Superpathway of Inositol Phosphate Compounds | −1.976 | −0.18 |
| Pyrimidine Ribonucleotides Interconversion | −1.414 | 0.707 |
| NER (Nucleotide Excision Repair, Enhanced Pathway) | −1.877 | 0.209 |
| Role of BRCA1 in DNA Damage Response | −1.225 | 0.816 |
| Unfolded Protein Response | −1.667 | −0.333 |
| Purine Nucleotides De Novo Biosynthesis II | −2 | 0 |
| HIPPO signaling | 1 | −1 |
| Pyrimidine Ribonucleotides De Novo Biosynthesis | −1.667 | 0.333 |
| Hypoxia Signaling in the Cardiovascular System | −2 | 0 |
| 3-Phosphoinositide Biosynthesis | −1.671 | 0.186 |
| IL-15 Production | −1.46 | 0.209 |
| Leukocyte Extravasation Signaling | 0.408 | −1.225 |
| Regulation of the Epithelial–Mesenchymal Transition in Development Pathway | 0.535 | −1.069 |
| Role of CHK Proteins in Cell Cycle Checkpoint Control | 1 | −0.5 |
| Glioma Signaling | −0.333 | −1 |
| Basal Cell Carcinoma Signaling | 0 | −1.155 |
| Huntington’s Disease Signaling | 0 | −1 |
| miRNA | Targeted Molecules | Entrez Gene Name |
|---|---|---|
| miR-1264 (and other miRNAs with the seed AAGUCUU) | CDC7 | cell division cycle 7 |
| ORC6 | origin recognition complex subunit 6 | |
| miR-1468-5p (miRNAs with the seed UCCGUUU) | ORC4 | origin recognition complex subunit 4 |
| ORC6 | origin recognition complex subunit 6 | |
| mir-192 | CDC7 | cell division cycle 7 |
| miR-302b-5p (and other miRNAs with the seed CUUUAAC) | DBF4 | DBF4 zinc finger |
| miR-4511 (miRNAs with the seed AAGAACU) | ORC5 | origin recognition complex subunit 5 |
| miR-489-5p (miRNAs with the seed GUCGUAU) | CHK2 | checkpoint kinase 2 |
| miR-523-3p (miRNAs with the seed AACGCGC) | CDC6 | cell division cycle 6 |
| miR-643 (miRNAs with the seed CUUGUAU) | ORC2 | origin recognition complex subunit 2 |
| ORC5 | origin recognition complex subunit 5 | |
| miR-767 (and other miRNAs with the seed GCACCAU) | CDC6 | cell division cycle 6 |
| CDC7 | cell division cycle 7 | |
| ORC6 | origin recognition complex subunit 6 | |
| miR-96-3p (miRNAs with the seed AUCAUGU) | CDC45 | cell division cycle 45 |
| CDT1 | chromatin licensing and DNA replication factor 1 | |
| ORC4 | origin recognition complex subunit 4 |
| Symbol | Entrez Gene Name | Location | Family | Interacting Molecules |
|---|---|---|---|---|
| mir-10 | microRNA 99a | Cytoplasm | microRNA | SMAD4, SUV39H1, p53 |
| mir-17 | microRNA 17 | Cyclin D, RB, p21Cip1 | ||
| mir-19 | microRNA 19a | SMAD4, p21Cip1 | ||
| mir-194 | microRNA 194-1 | MDM2 | ||
| mir-224 | microRNA 224 | SMAD4 | ||
| mir-25 | microRNA 25 | MDM2, p21Cip1, p53 | ||
| mir-34 | microRNA 34a | CDK4/6, c-MYC, p53 | ||
| mir-451 | microRNA 451a | p19INK4 | ||
| mir-605 | microRNA 605 | MDM2 | ||
| MYC-targeting siRNA DCR-MYC | Other | Biologic drug | c-Myc |
| miRNA | Targeted Molecules | Entrez Gene Name |
|---|---|---|
| mir-10 | ATM | ATM serine/threonine kinase |
| mir-145 | p53 | tumor protein p53 |
| mir-15 | CDC25A | cell division cycle 25A |
| WEE1 | WEE1 G2 checkpoint kinase | |
| c-RAF | Raf-1 proto-oncogene, serine/threonine kinase | |
| mir-17 | ATM | ATM serine/threonine kinase |
| CyclinD1 | ||
| RB | RB transcriptional corepressor 1 | |
| p21CIP1 | cyclin dependent kinase inhibitor 1A | |
| mir-221 | p27KIP1 | cyclin dependent kinase inhibitor 1B |
| mir-25 | p21CIP1 | cyclin dependent kinase inhibitor 1A |
| p53 | tumor protein p53 | |
| mir-290 | CDK2 | cyclin dependent kinase 2 |
| mir-34 | CDK4/6 | cyclin dependent kinase 4/6 |
| p53 | tumor protein p53 | |
| mir-451 | p19INK4D | cyclin dependent kinase inhibitor 2D |
| mir-497 | CDC25A | cell division cycle 25A |
| c-RAF | Raf-1 proto-oncogene, serine/threonine kinase |
| miRNA | Targeted Molecules | Entrez Gene Name |
|---|---|---|
| miR-125b-2-3p (and other miRNAs with the seed CAAGUCA) | p53 | tumor protein p53 |
| miR-302b-5p (and other miRNAs with the seed CUUUAAC) | BARD1 | BRCA1 associated RING domain 1 |
| CTIP | RB binding protein 8, endonuclease | |
| GADD45 | growth arrest and DNA damage inducible alpha | |
| miR-6074 (miRNAs with the seed AUAUUCA) | FANCF | FA complementation group F |
| IFNG | interferon gamma | |
| NBS1 | nibrin | |
| mir-101 | ATM | ATM serine/threonine kinase |
| mir-103 | p53 | tumor protein p53 |
| mir-145 | p53 | tumor protein p53 |
| mir-146 | STAT1 | signal transducer and activator of transcription 1 |
| mir-17 | ATM | ATM serine/threonine kinase |
| RB | RB transcriptional corepressor 1 | |
| p21CIP1 | cyclin dependent kinase inhibitor 1A | |
| mir-25 | p21CIP1 | cyclin dependent kinase inhibitor 1A |
| p53 | tumor protein p53 |
| Symbol | Entrez Gene Name | Location | Family | Interacting Molecules |
|---|---|---|---|---|
| Lipid-encapsulated anti-PLK1 siRNA TKM-080301 | Other | Biologic drug | PLK1 | |
| mir-10 | microRNA 99a | Cytoplasm | microRNA | p53, p90RSK |
| mir-101 | microRNA 101-1 | ATM, DNA-PK | ||
| mir-15 | microRNA 15a | CHEK1, PPM1D, WEE1 | ||
| mir-17 | microRNA 17 | ATM, p21Cip1 | ||
| mir-194 | microRNA 194-1 | MDM2 | ||
| mir-25 | microRNA 25 | MDM2, p21Cip1, p53 | ||
| mir-32 | microRNA 32 | MDM2 | ||
| mir-34 | microRNA 34a | MDM4, p53 | ||
| mir-605 | microRNA 605 | MDM2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Yokozaki, H.; Sasaki, H. Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer. Cancers 2021, 13, 5786. https://doi.org/10.3390/cancers13225786
Tanabe S, Quader S, Ono R, Cabral H, Aoyagi K, Hirose A, Yokozaki H, Sasaki H. Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer. Cancers. 2021; 13(22):5786. https://doi.org/10.3390/cancers13225786
Chicago/Turabian StyleTanabe, Shihori, Sabina Quader, Ryuichi Ono, Horacio Cabral, Kazuhiko Aoyagi, Akihiko Hirose, Hiroshi Yokozaki, and Hiroki Sasaki. 2021. "Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer" Cancers 13, no. 22: 5786. https://doi.org/10.3390/cancers13225786
APA StyleTanabe, S., Quader, S., Ono, R., Cabral, H., Aoyagi, K., Hirose, A., Yokozaki, H., & Sasaki, H. (2021). Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer. Cancers, 13(22), 5786. https://doi.org/10.3390/cancers13225786