
Mangrove Forest Cover and Phenology with Landsat Dense Time Series in Central Queensland, Australia



Abstract
:
1. Introduction
2. Materials and Methods
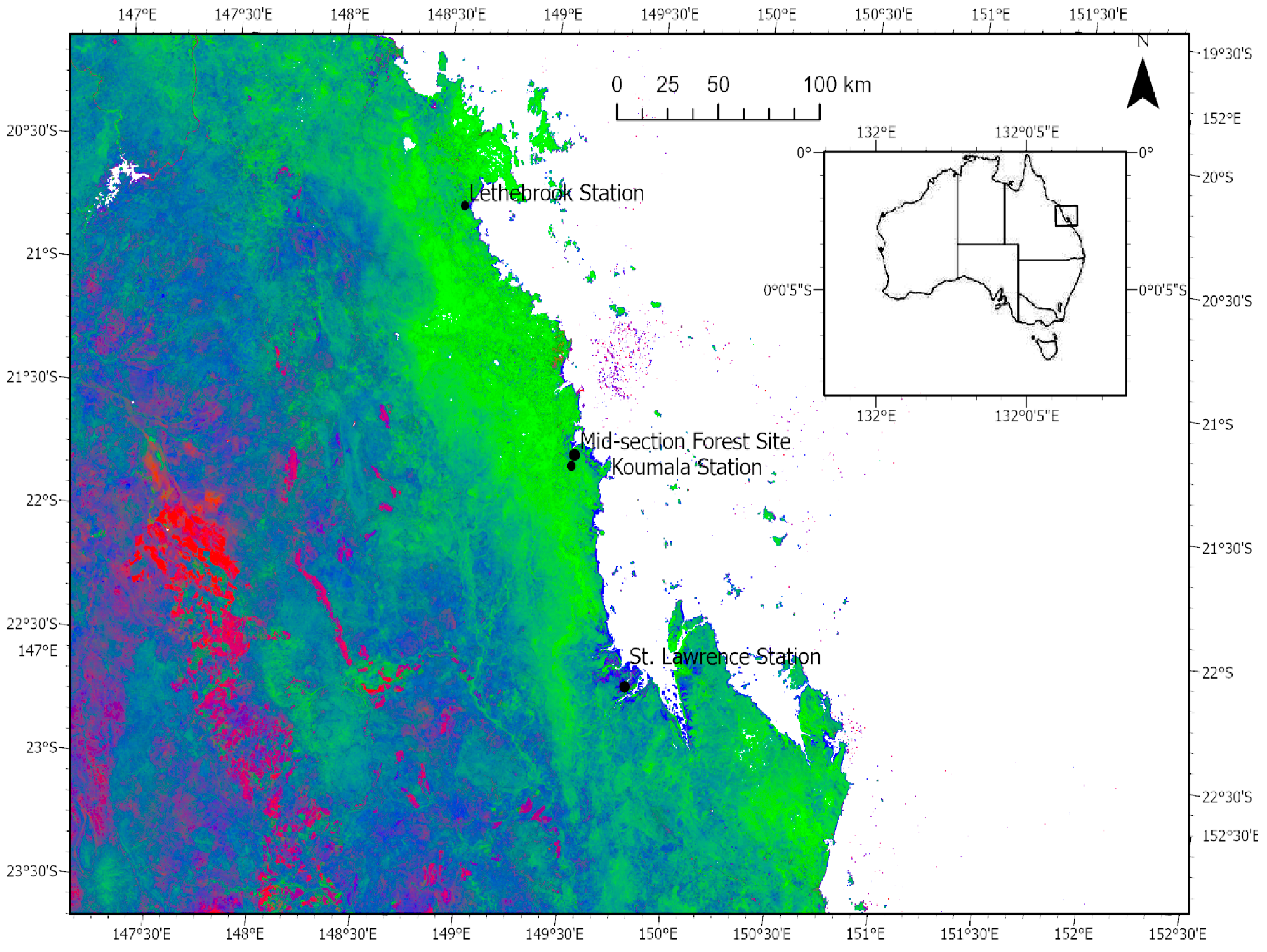
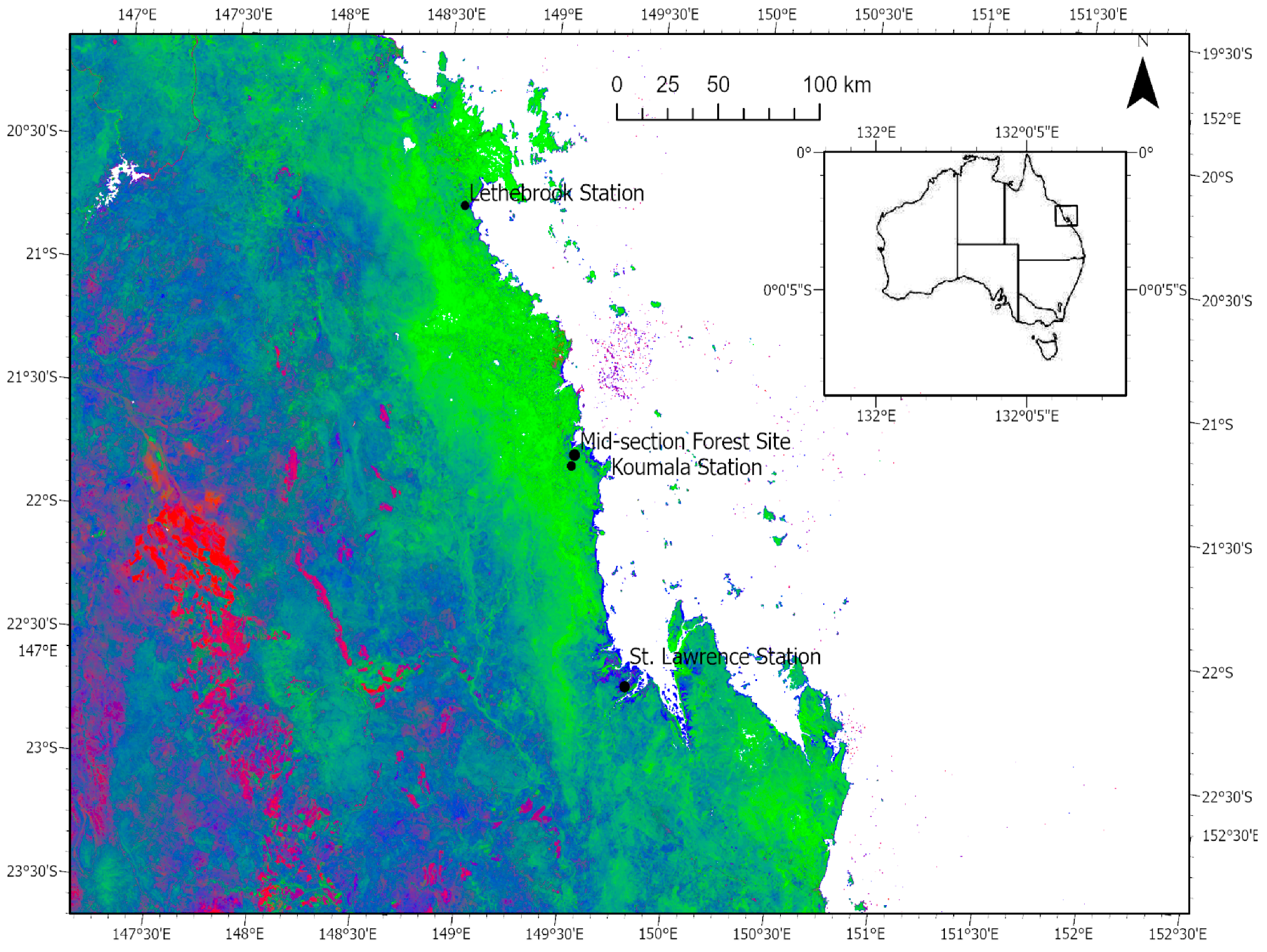
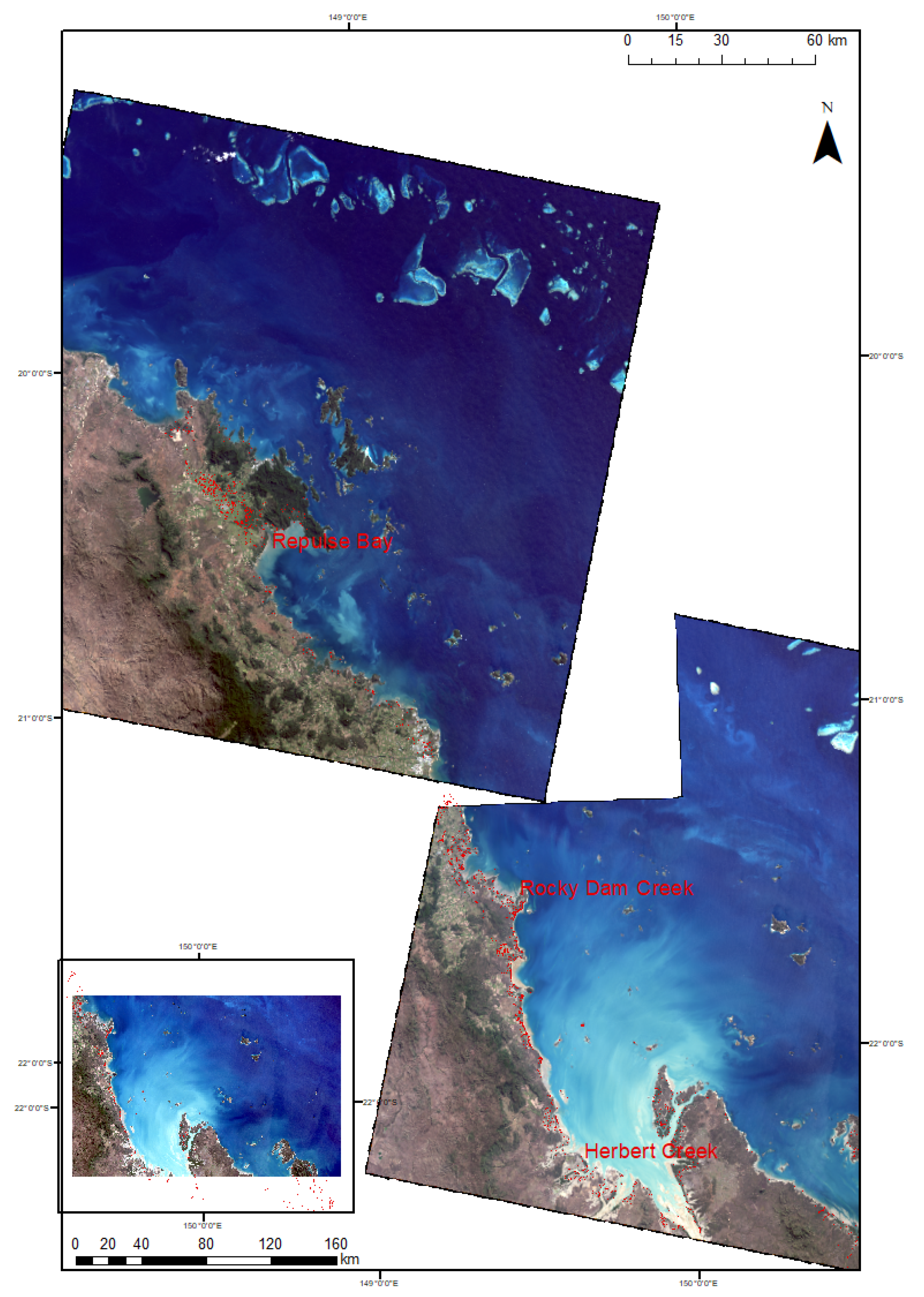
2.1. Study Area
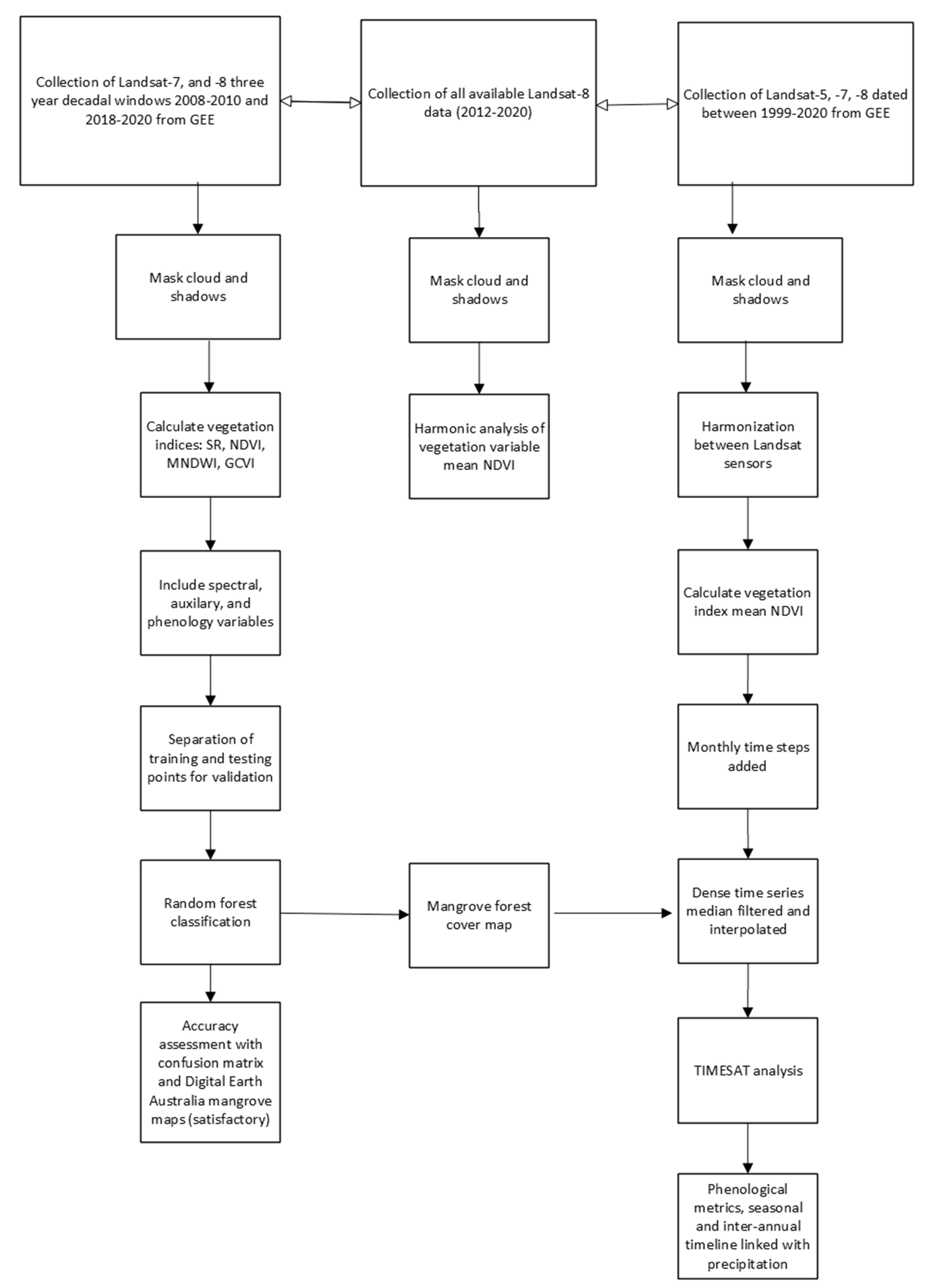
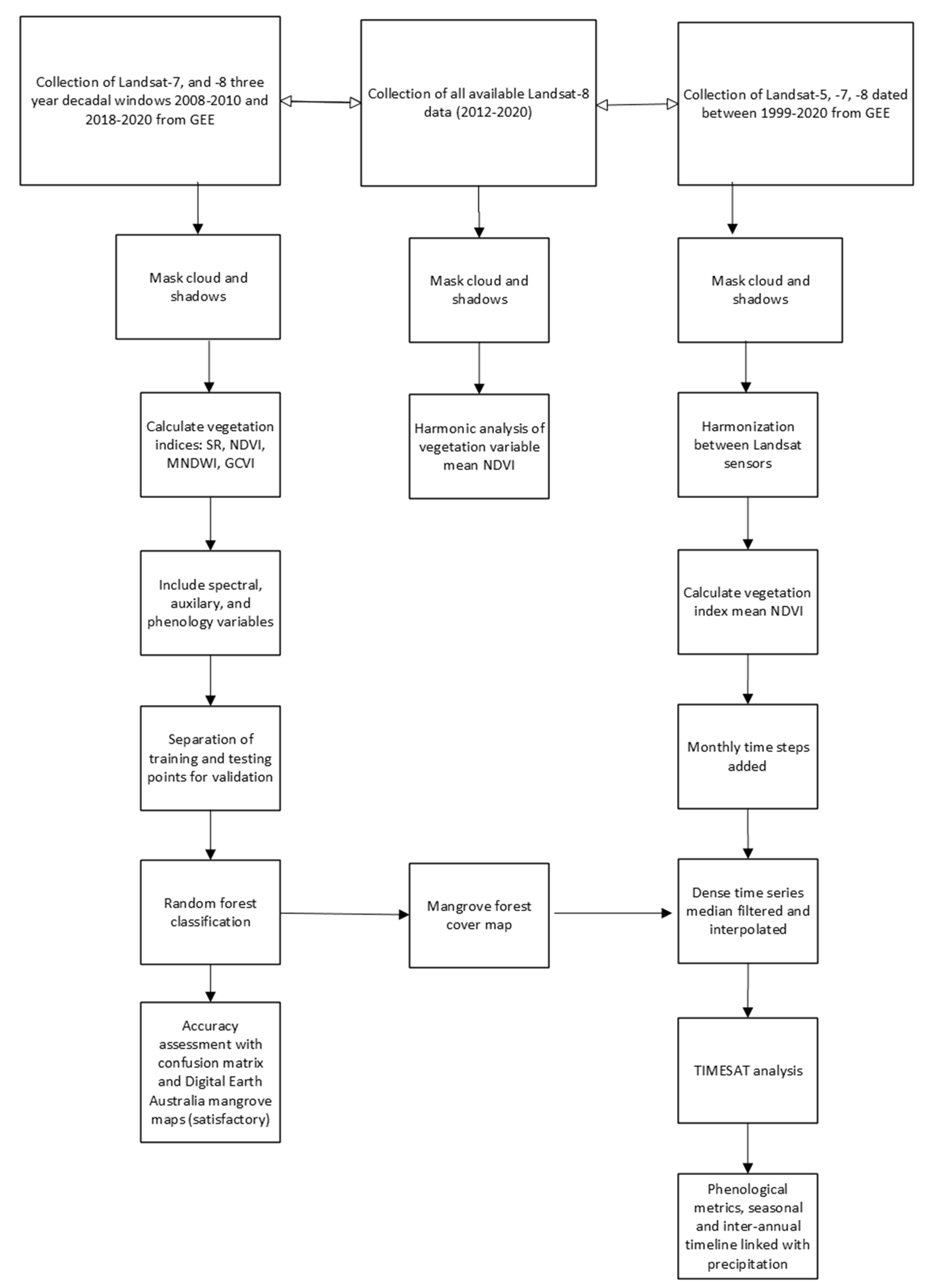
2.2. Image Classification
2.3. Mangrove Phenology
TIMESAT Analysis
3. Results
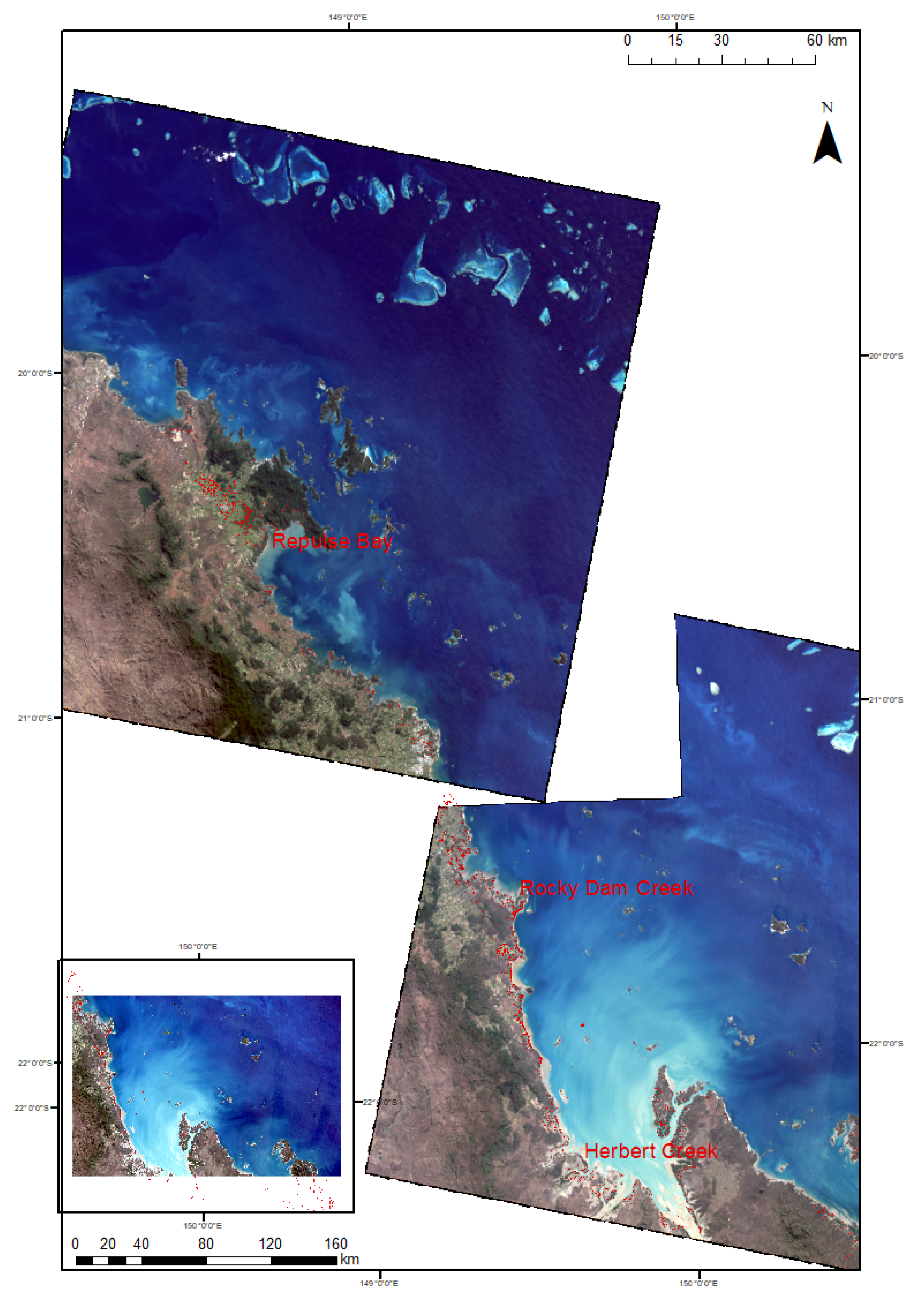
3.1. Image Classification
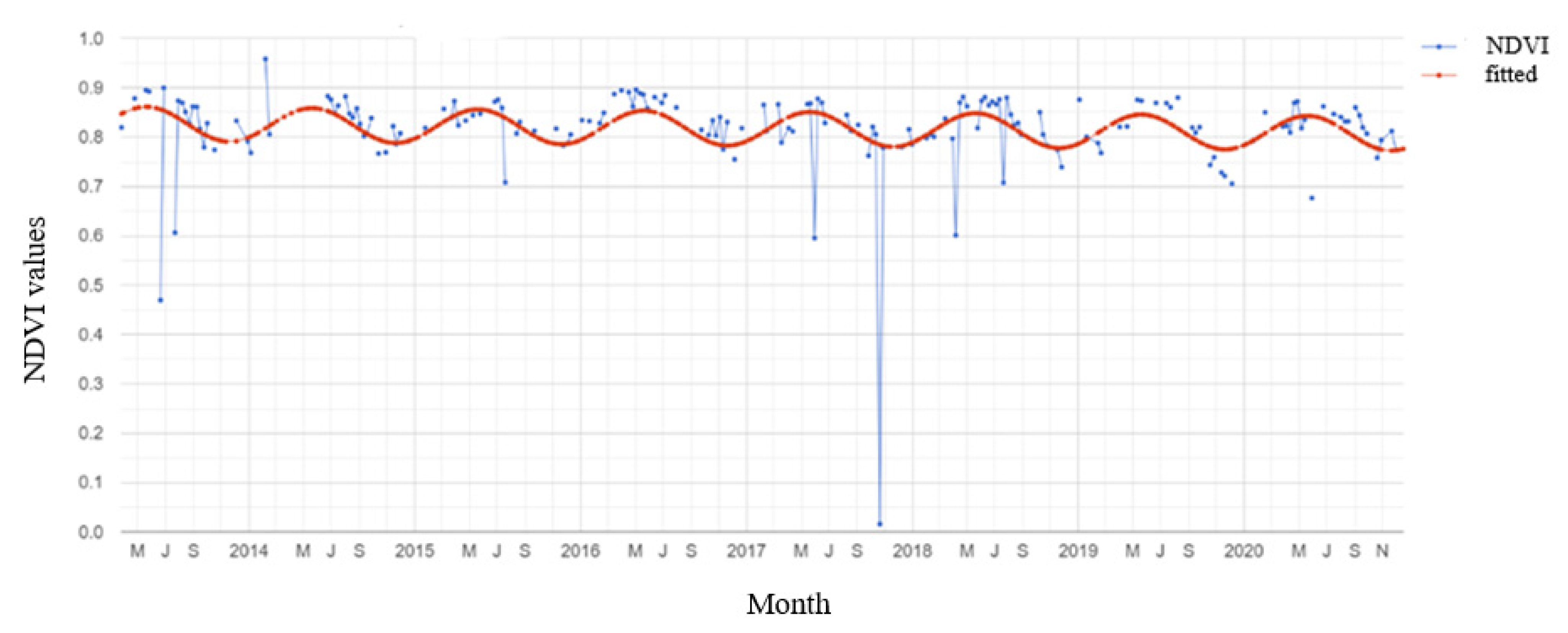
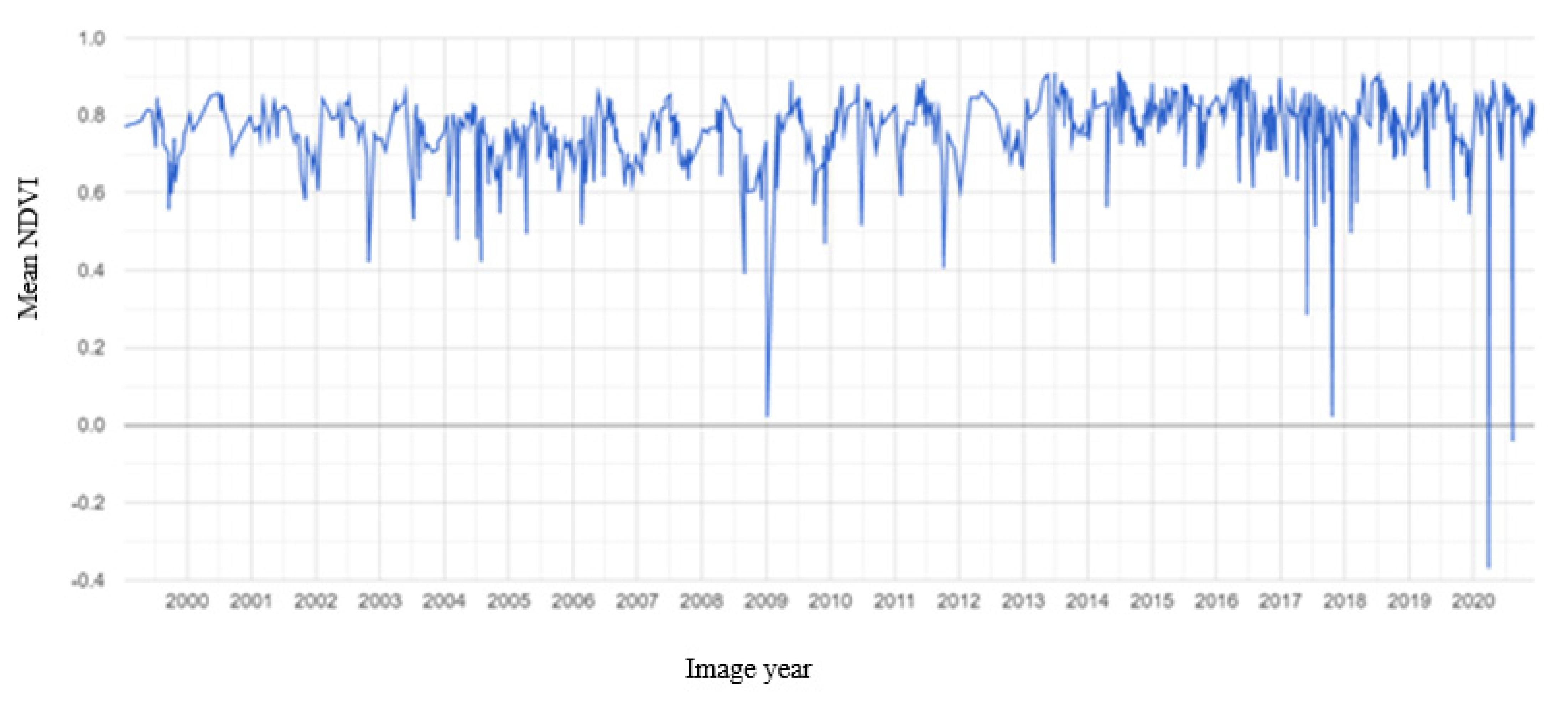
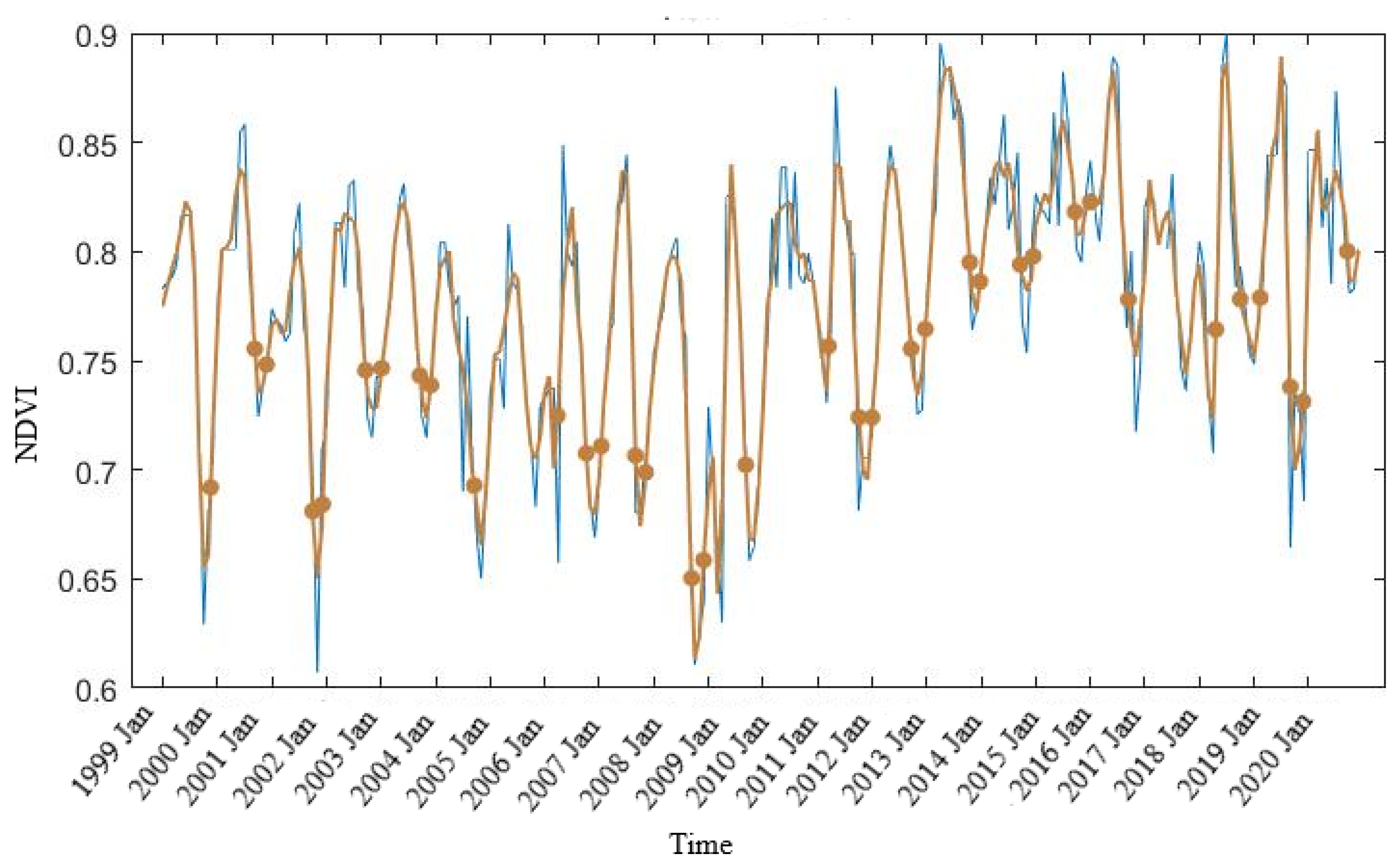
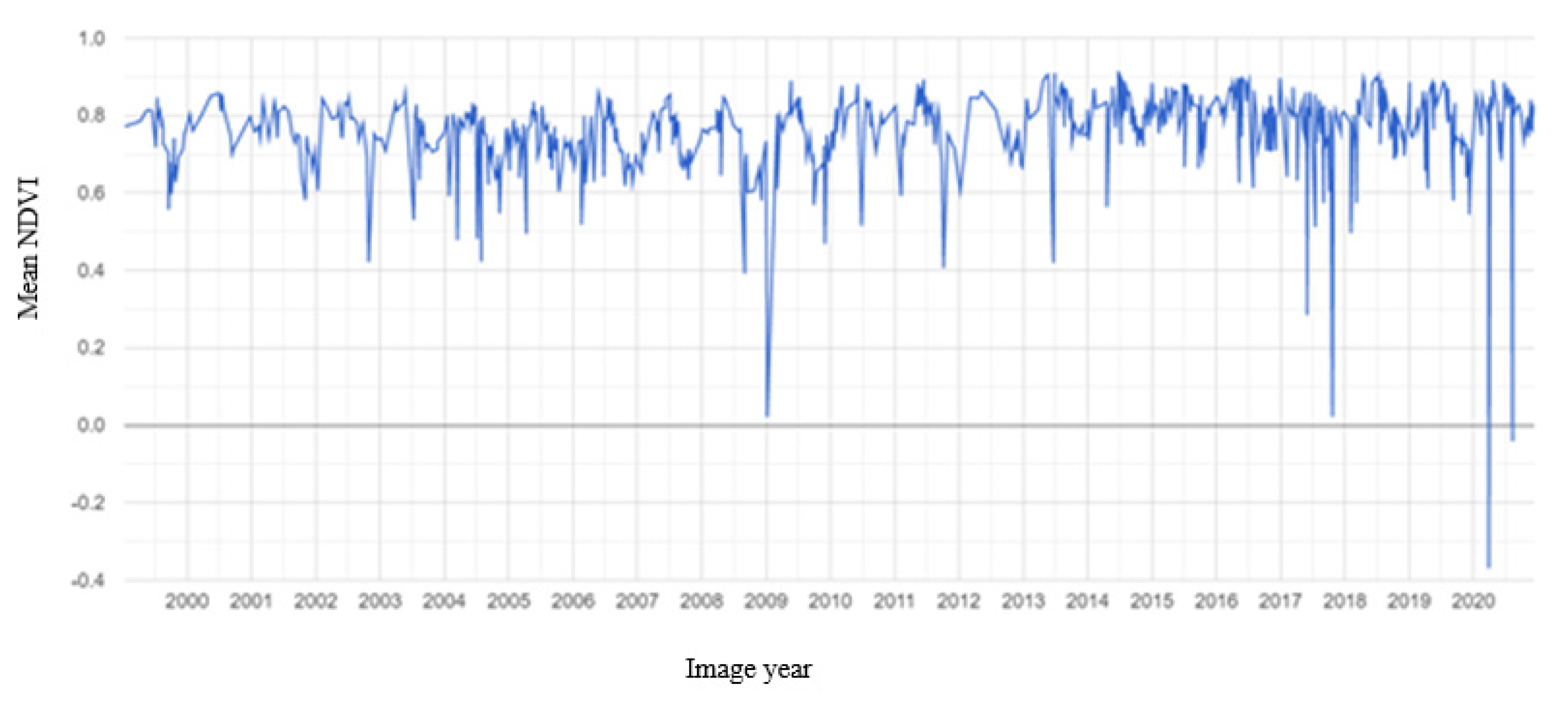
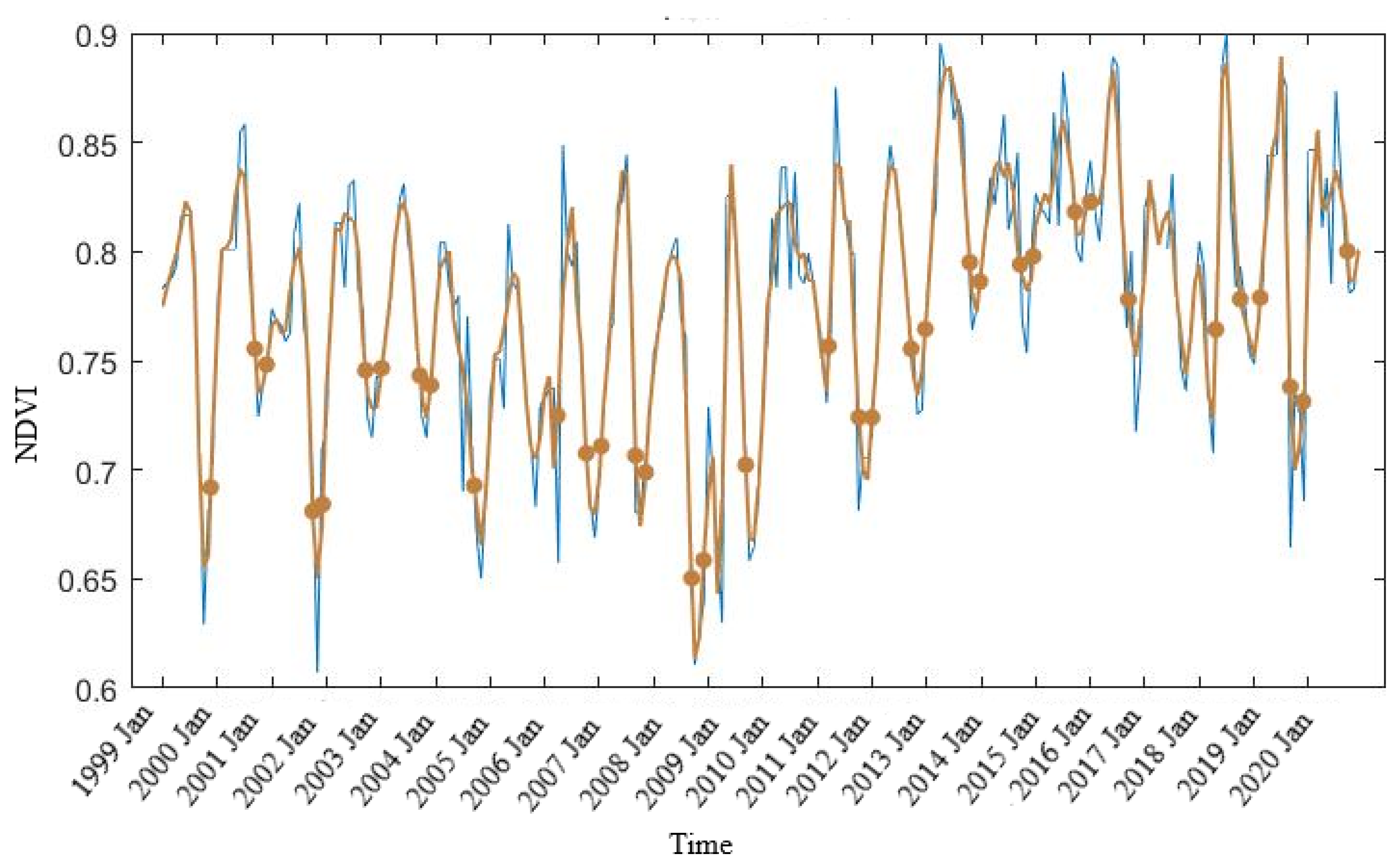
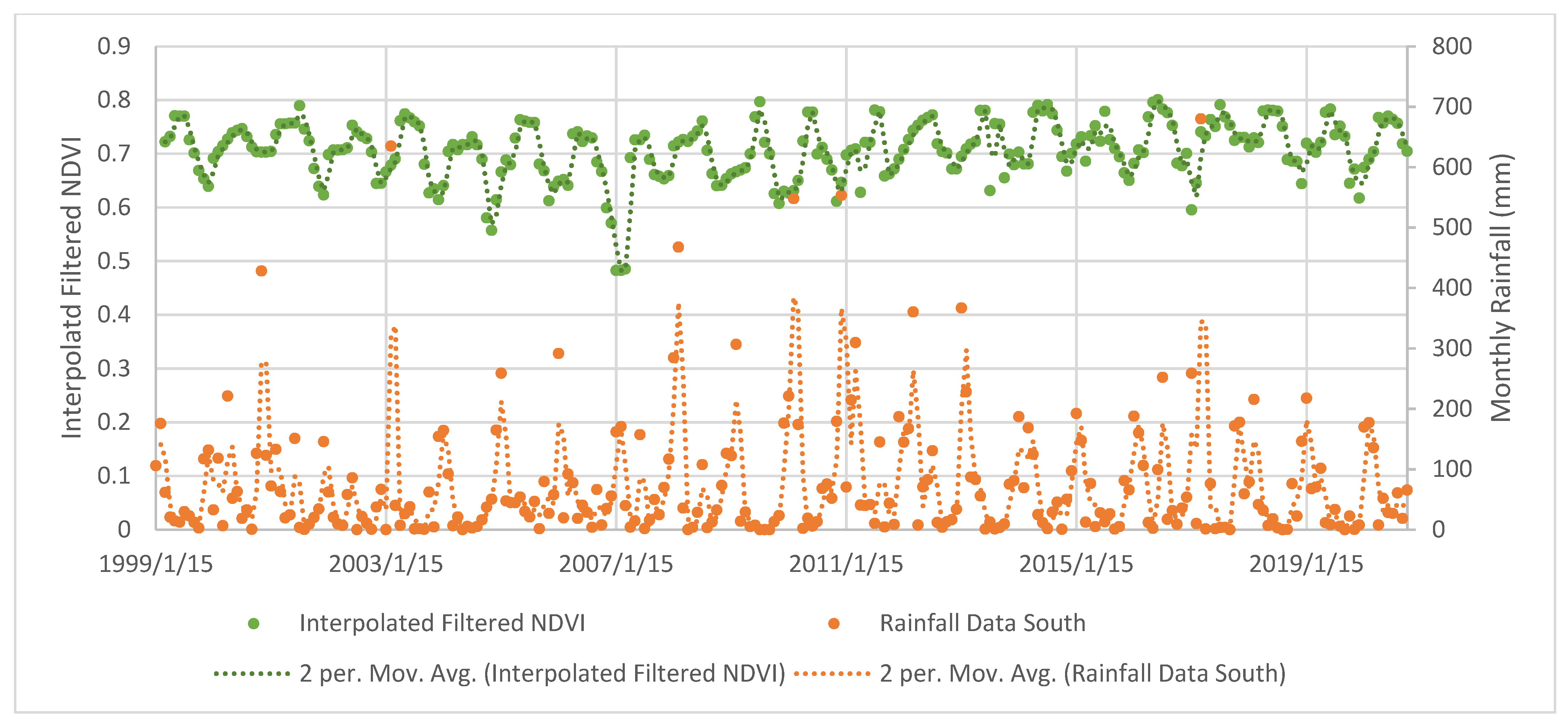
3.2. Mangrove Phenology
TIMESAT Analysis
4. Discussion
4.1. Change Dynamics in Mangrove Forests
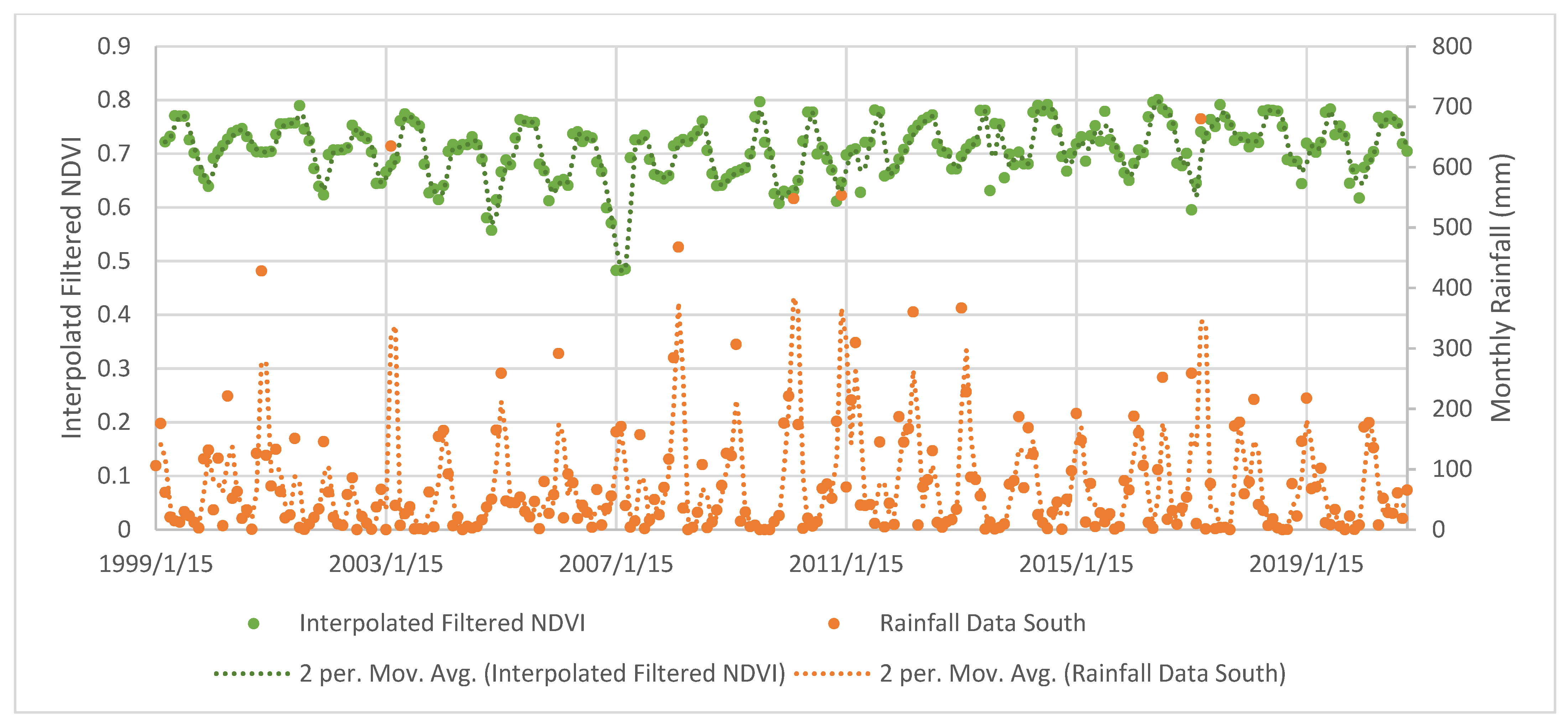
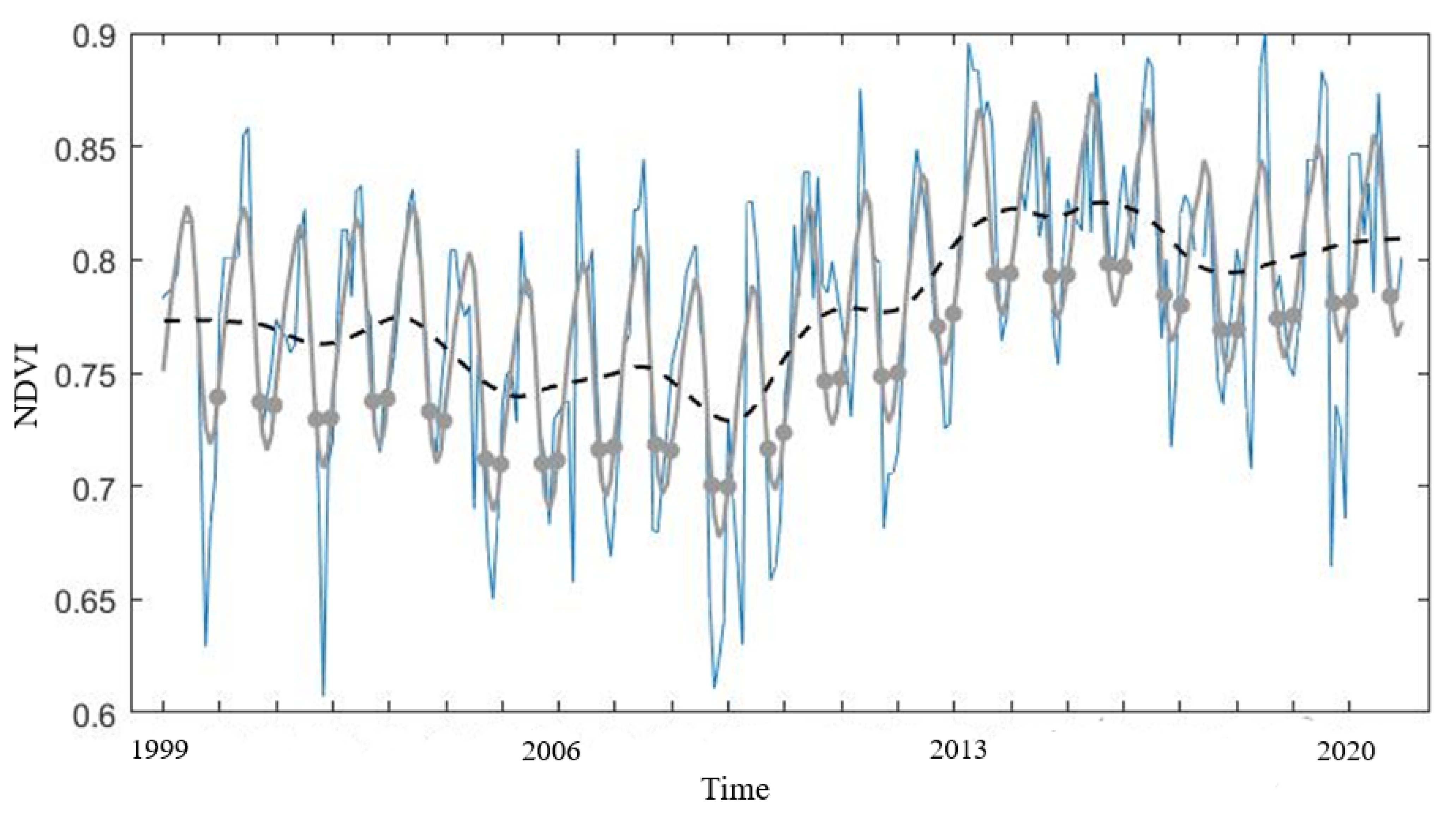
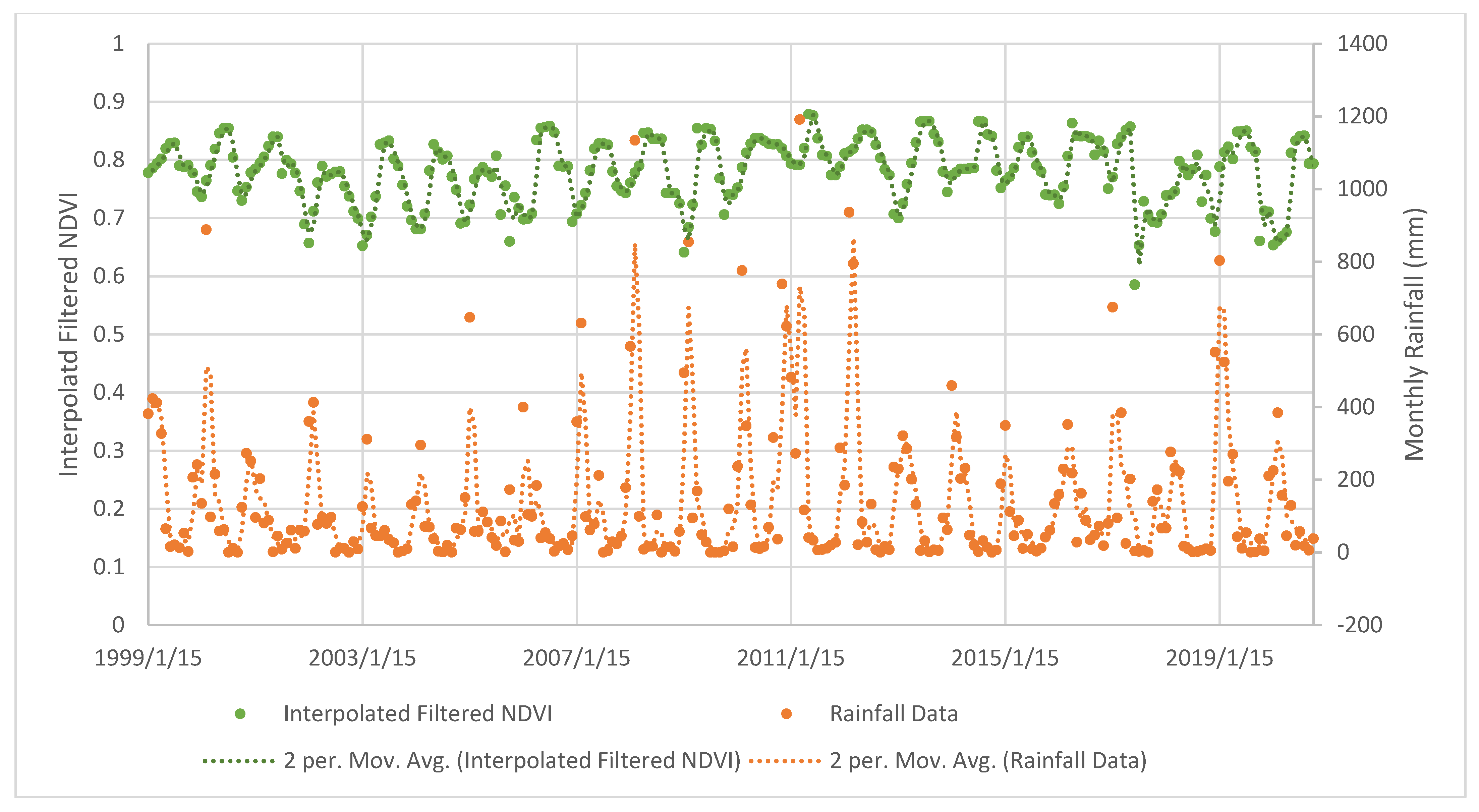
4.2. Mangrove Phenology
TIMESAT Analysis
4.3. Limitations of the Study
4.4. Implications for the Conservation of Mangrove Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological role and services of tropical mangrove ecosystems: A reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Sheaves, M.; Johnston, R.; Baker, R. Use of mangroves by fish: New insights from in-forest videos. Mar. Ecol. Prog. Ser. 2016, 549, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Macreadie, P.I.; Anton, A.; Raven, J.A.; Beaumont, N.; Connolly, R.M.; Friess, D.A.; Kelleway, J.J.; Kennedy, H.; Kuwae, T.; Lavery, P.S.; et al. The future of Blue Carbon science. Nat. Commun. 2019, 10, 3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adame, M.F.; Connolly, R.M.; Turschwell, M.P.; Lovelock, C.E.; Fatoyinbo, T.; Lagomasino, D.; Goldberg, L.A.; Holdorf, J.; Friess, D.A.; Sasmito, S.D.; et al. Future carbon emissions from global mangrove forest loss. Glob. Chang. Biol. 2021. [Google Scholar] [CrossRef]
- The International Council on Clean Transportation. Vision 2050: A Strategy to Decarbonize the Global Transport Sector by Mid-Century; The International Council on Clean Transportation: Washington, DC, USA, 2020; pp. 1–25. [Google Scholar]
- Friess, D.A.; Yando, E.S.; Abuchahla, G.M.O.; Adams, J.B.; Cannicci, S.; Canty, S.W.J.; Cavanaugh, K.C.; Connolly, R.M.; Cormier, N.; Dahdouh-Guebas, F.; et al. Mangroves give cause for conservation optimism, for now. Curr. Biol. 2020, 30, R153–R154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saintilan, N.; Rogers, K.; Kelleway, J.J.; Ens, E.; Sloane, D.R. Climate change impacts on the coastal wetlands of Australia. Wetlands 2018, 38, 1145–1154. [Google Scholar] [CrossRef]
- Spalding, M.D.; Ruffo, S.; Lacambra, C.; Meliane, I.; Hale, L.Z.; Shepard, C.C.; Beck, M.W. The role of ecosystems in coastal protection: Adapting to climate change and coastal hazards. Ocean Coast. Manag. 2014, 90, 50–57. [Google Scholar] [CrossRef]
- Fang, X.; Hou, X.; Li, X.; Hou, W.; Nakaoka, M.; Yu, X. Ecological connectivity between land and sea: A review. Ecol. Res. 2018, 33, 51–61. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove dieback during fluctuating sea levels. Sci. Rep. 2017, 7, 1680. [Google Scholar] [CrossRef]
- UNGA: United Nations General Assembly. Work of the Statistical Commission Pertaining to the 2030 Agenda for Sustainable Development: Resolution/Adopted by the General Assembly (A/RES/71/313); UNGA: New York, NY, USA, 2017. [Google Scholar]
- de Beurs, K.M.; Henebry, G.M. Spatio-temporal statistical methods for modelling land surface phenology. In Phenological Research: Methods for Environmental and Climate Change Analysis; Hudson, I.L., Keatley, M.R., Eds.; Springer: New York, NY, USA, 2010; pp. 177–208. [Google Scholar]
- Hostert, P.; Griffiths, P.; van der Linden, S.; Pflugmacher, D. Time series analyses in a new era of optical satellite data. In Remote Sensing Time Series; Springer International Publishing: Cham, Switzerland, 2015; pp. 25–41. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; El-Askary, H.; Qurban, M.A.; Li, J.; ManiKandan, K.P.; Piechota, T. Using multi-indices approach to quantify mangrove changes over the Western Arabian Gulf along Saudi Arabia coast. Ecol. Indic. 2019, 102, 734–745. [Google Scholar] [CrossRef]
- Wu, N.; Shi, R.; Zhuo, W.; Zhang, C.; Zhou, B.; Xia, Z.; Tao, Z.; Gao, W.; Tian, B. A classification of tidal flat wetland vegetation combining phenological features with Google Earth Engine. Remote Sens. 2021, 13, 443. [Google Scholar] [CrossRef]
- Keatley, M.R.; Hudson, I.L. Introduction and overview. In Phenological Research: Methods for Environmental and Climate Change Analysis; Hudson, I.L., Keatley, M.R., Eds.; Springer: New York, NY, USA, 2010; pp. 1–22. [Google Scholar] [CrossRef]
- Younes, N.; Joyce, K.E.; Maier, S.W. All models of satellite-derived phenology are wrong, but some are useful: A case study from northern Australia. Int. J. Appl. Earth Obs. Geoinf. 2021, 97, 102285. [Google Scholar] [CrossRef]
- Jimenez Galo, A.J. Monitoring of tropical forest cover with remote sensing. In Tropical Forestry Handbook; Pancel, L., Köhl, M., Eds.; Springer: Berlin, Germany, 2016; pp. 1–19. [Google Scholar] [CrossRef]
- DeVries, B.; Pratihast, A.K.; Verbesselt, J.; Kooistra, L.; Herold, M. Characterizing forest change using community-based monitoring data and Landsat time series. PLoS ONE 2016, 11, e0147121. [Google Scholar] [CrossRef] [PubMed]
- Duarte, E.; Barrera, J.A.; Dube, F.; Casco, F.; Hernández, A.J.; Zagal, E. Monitoring approach for tropical coniferous forest degradation using remote sensing and field data. Remote Sens. 2020, 12, 2531. [Google Scholar] [CrossRef]
- Souza, C.M.; Shimbo, J.Z.; Rosa, M.R.; Parente, L.L.; Alencar, A.A.; Rudorff, B.F.T.; Hasenack, H.; Matsumoto, M.; Ferreira, G.L.; Souza-Filho, P.W.M.; et al. Reconstructing three decades of land use and land cover changes in Brazilian biomes with Landsat archive and Earth Engine. Remote Sens. 2020, 12, 2735. [Google Scholar] [CrossRef]
- Criminisi, A. Decision Forests: A unified framework for classification, regression, density estimation, manifold learning and semi-supervised learning. Found. Trends Comput. Graph. Vis. 2011, 7, 81–227. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer: New York, NY, USA, 2017. [Google Scholar]
- Pham, L.T.H.; Brabyn, L. Monitoring mangrove biomass change in Vietnam using SPOT images and an object-based approach combined with machine learning algorithms. ISPRS J. Photogramm. Remote Sens. 2017, 128, 86–97. [Google Scholar] [CrossRef]
- Hu, L.; Li, W.; Xu, B. Monitoring mangrove forest change in China from 1990 to 2015 using Landsat-derived spectral-temporal variability metrics. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 88–98. [Google Scholar] [CrossRef]
- Diniz, C.; Cortinhas, L.; Nerino, G.; Rodrigues, J.; Sadeck, L.; Adami, M.; Souza-Filho, P. Brazilian mangrove status: Three decades of satellite data analysis. Remote Sens. 2019, 11, 808. [Google Scholar] [CrossRef] [Green Version]
- Bihamta Toosi, N.; Soffianian, A.R.; Fakheran, S.; Pourmanafi, S.; Ginzler, C.; Waser, T.L. Land cover classification in mangrove ecosystems based on VHR satellite data and machine learning—an upscaling approach. Remote Sens. 2020, 12, 2884. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sens. 2017, 2017, 1353691. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.; Nichol, C.; Disney, M.; Lewis, P.; Quaife, T.; Bowyer, P. Can we measure terrestrial photosynthesis from space directly, using spectral reflectance and fluorescence? Glob. Chang. Biol. 2007, 13, 1484–1497. [Google Scholar] [CrossRef]
- Venkatappa, M.; Sasaki, N.; Shrestha, R.P.; Tripathi, N.K.; Ma, H.O. Determination of vegetation thresholds for assessing land use and land use changes in Cambodia using the Google Earth Engine cloud-computing platform. Remote Sens. 2019, 11, 1514. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Zhang, J.; Yan, H.; Wu, W.; Biradar, C. Land surface phenology. In Phenology of Ecosystem Processes: Applications in Global Change Research; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 247–270. [Google Scholar] [CrossRef]
- Priyadarshi, N.; Chowdary, V.M.; Srivastava, Y.K.; Das, I.C.; Jha, C.S. Reconstruction of time series MODIS EVI data using de-noising algorithms. Geocarto Int. 2018, 33, 1095–1113. [Google Scholar] [CrossRef]
- Shamim Hasan Mandal, M.; Kamruzzaman, M.; Hosaka, T. Elucidating the phenology of the Sundarbans mangrove forest using 18-year time series of MODIS vegetation indices. Tropics 2020, 29, 41–55. [Google Scholar] [CrossRef]
- Sadinski, W.; Gallant, A.L.; Roth, M.; Brown, J.; Senay, G.; Brininger, W.; Jones, P.M.; Stoker, J. Multi-year data from satellite- and ground-based sensors show details and scale matter in assessing climate’s effects on wetland surface water, amphibians, and landscape conditions. PLoS ONE 2018, 13, e0201951. [Google Scholar] [CrossRef]
- Kelly, N. Accounting for correlated error structure within phenological data: A case study of trend analysis of snowdrop flowering. In Phenological Research: Methods for Environmental and Climate Change Analysis; Hudson, L.L., Keatley, M.R., Eds.; Springer: New York, NY, USA, 2010; pp. 271–298. [Google Scholar] [CrossRef]
- Adams, B.; Iverson, L.; Matthews, S.; Peters, M.; Prasad, A.; Hix, D.M. Mapping forest composition with Landsat time series: An evaluation of seasonal composites and harmonic regression. Remote Sens. 2020, 12, 610. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Lu, D.; Wu, M.; Shao, X.; Wei, J. Examining land cover and greenness dynamics in Hangzhou Bay in 1985–2016 using Landsat time-series data. Remote Sens. 2017, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Lamb, B.T.; Tzortziou, M.A.; McDonald, K.C. Evaluation of approaches for mapping tidal wetlands of the Chesapeake and Delaware Bays. Remote Sens. 2019, 11, 366. [Google Scholar] [CrossRef] [Green Version]
- Campbell, A.D.; Wang, Y. Salt marsh monitoring along the mid-Atlantic coast by Google Earth Engine enabled time series. PLoS ONE 2020, 15, e0229605. [Google Scholar] [CrossRef] [Green Version]
- Department of Agriculture Water and the Environment. Directory of Important Wetlands in Australia-Information Sheet, Sarina Inlet-Ince Bay Aggregation-QLD053. Available online: https://www.environment.gov.au/cgi-bin/wetlands/report.pl (accessed on 1 March 2021).
- Chamberlain, D.; Phinn, S.; Possingham, H. Remote sensing of mangroves and estuarine communities in Central Queensland, Australia. Remote Sens. 2020, 12, 197. [Google Scholar] [CrossRef] [Green Version]
- Ronan, M. Ramsar Information Sheet: Shoalwater and Corio Bays Area, Australia; Queensland Department of Environment and Heritage Protection: Brisbane, QLD, Australia, 2018.
- Folkers, A.; Rohde, K.; Delaney, K.; Flett, I. Mackay Whitsunday Water Quality Improvement Plan 2014–2021; Reef Catchments: Mackay, QLD, Australia, 2014. [Google Scholar]
- Reef Catchments Limited. Natural Resource Management Plan; Mackay Whitsunday Isaac: Mackay, QLD, Australia, 2014. [Google Scholar]
- Pascoe, S.; Innes, J.; Tobin, R.; Stoeckle, N.; Paredes, S.; Dauth, K. Beyond GVP: The Value of Inshore Commercial Fisheries to Fishers and Consumers in Regional Communities on Queensland’s East Coast; FRDC Project No 2013-301: Deakin, VIC, Australia, 2016. [Google Scholar]
- Webley, J.; McInnes, K.; Teixeira, D.; Lawson, A.; Quinn, R. Statewide Recreational Fishing Survey 2013-14; Department of Agriculture and Fisheries: Brisbane, QLD, Australia, 2015. [Google Scholar]
- Reef Catchments. State of the Region Report; Mackay Whitsunday Isaac: Mackay, QLD, Australia, 2013. [Google Scholar]
- Duke, N.C. Australia’s Mangroves: The Authoritative Guide to Australia’s Mangrove Plants; University of Queensland: Brisbane, QLD, Australia, 2006. [Google Scholar]
- Duke, N.C.; Larkum, A.W.D. Mangroves and seagrasses. In The Great Barrier Reef: Biology, environment and Management, 2nd ed.; Hutchings, P., Kingsford, M., Hoegh-Guldberg, O., Eds.; CSIRO: Clayton South, VIC, Australia, 2019; pp. 219–228. [Google Scholar]
- AusCover. Seasonal Fractional Vegetation Cover for Queensland Derived from USGS Landsat Images. Available online: http://data.auscover.org.au/xwiki/bin/view/Product+pages/Landsat+Seasonal+Fractional+Cover (accessed on 1 March 2021).
- Kuenzer, C.; Bluemel, A.; Gebhardt, S.; Quoc, T.V.; Dech, S. Remote sensing of mangrove ecosystems: A review. Remote Sens. 2011, 3, 878–928. [Google Scholar] [CrossRef] [Green Version]
- Vermote, E.; Justice, C.; Claverie, M.; Franch, B. Preliminary analysis of the performance of the Landsat 8/OLI land surface reflectance product. Remote Sens. Environ. 2016, 185, 46–56. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Bunting, P.; Rosenqvist, A.; Lucas, R.; Rebelo, L.-M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, M. The global mangrove watch-a new 2010 global baseline of mangrove extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef] [Green Version]
- Lymburner, L.; Bunting, P.; Lucas, R.; Scarth, P.; Alam, I.; Phillips, C.; Ticehurst, C.; Held, A. Mapping the multi-decadal mangrove dynamics of the Australian coastline. Remote Sens. Environ. 2020, 238, 111185. [Google Scholar] [CrossRef]
- USGS. Landsat Collection 1 Level 1 Product Definition; United States Geological Survey: Vermillion, SD, USA, 2017; pp. 1–24.
- Wu, C.; Niu, Z.; Gao, S. The potential of the satellite derived green chlorophyll index for estimating midday light use efficiency in maize, coniferous forest and grassland. Ecol. Indic. 2012, 14, 66–73. [Google Scholar] [CrossRef]
- Pastor-Guzman, J.; Atkinson, P.; Dash, J.; Rioja-Nieto, R. Spatiotemporal variation in mangrove chlorophyll concentration using Landsat 8. Remote Sens. 2015, 7, 14530–14558. [Google Scholar] [CrossRef] [Green Version]
- Karlson, M.; Ostwald, M.; Reese, H.; Sanou, J.; Tankoano, B.; Mattsson, E. Mapping tree canopy cover and aboveground biomass in Sudano-Sahelian Woodlands using Landsat 8 and Random Forest. Remote Sens. 2015, 7, 10017–10041. [Google Scholar] [CrossRef] [Green Version]
- Berhane, T.M.; Lane, C.R.; Wu, Q.; Autrey, B.C.; Anenkhonov, O.A.; Chepinoga, V.V.; Liu, H. Decision-Tree, rule-based, and Random Forest classification of high-resolution multispectral imagery for wetland mapping and inventory. Remote Sens. 2018, 10, 580. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.D.; Xia, J.; Ha, N.T.; Bui, D.T.; Le, N.N.; Tekeuchi, W. A review of remote sensing approaches for monitoring blue carbon ecosystems: Mangroves, seagrasses and salt marshes during 2010-2018. Sensors 2019, 19, 1933. [Google Scholar] [CrossRef] [Green Version]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Liu, B.; Gao, L.; Li, B.; Marcos-Martinez, R.; Bryan, B.A. Nonparametric machine learning for mapping forest cover and exploring influential factors. Landsc. Ecol. 2020, 35, 1683–1699. [Google Scholar] [CrossRef]
- Pelletier, C.; Valero, S.; Inglada, J.; Champion, N.; Dedieu, G. Assessing the robustness of Random Forests to map land cover with high resolution satellite image time series over large areas. Remote Sens. Environ. 2016, 187, 156–168. [Google Scholar] [CrossRef]
- Maxwell, A.E.; Warner, T.A.; Fang, F. Implementation of machine-learning classification in remote sensing: An applied review. Int. J. Remote Sens. 2018, 39, 2784–2817. [Google Scholar] [CrossRef] [Green Version]
- Eklundh, L.; Jonsson, P. TIMESAT for processing time-series data from satellite sensors for land surface monitoring. In Multitemporal Remote Sensing: Methods and Applications; Ban, Y., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 20, pp. 177–194. [Google Scholar]
- Jönsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Stanimirova, R.; Cai, Z.; Melaas, E.K.; Gray, J.M.; Eklundh, L.; Jönsson, P.; Friedl, M.A. An empirical assessment of the MODIS land cover dynamics and TIMESAT land surface phenology algorithms. Remote Sens. 2019, 11, 2201. [Google Scholar] [CrossRef] [Green Version]
- Valderrama-Landeros, L.; España-Boquera, M.L.; Baret, F.; Sánchez-Vargas, N.; Sáenz-Romero, C. Capacity of phenological data derived from Cyclopes Lai for the year 2000 to distinguish land cover types in the State of MichoacÁn, Mexico. Revista Chapingo Serie Ciencias Forestales Y Del Ambiente 2014, 20, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Hentze, K.; Thonfeld, F.; Menz, G. Evaluating crop area mapping from MODIS time-series as an assessment tool for Zimbabwe’s “Fast Track Land Reform Programme”. PLoS ONE 2016, 11, e0156630. [Google Scholar] [CrossRef]
- Kong, F.; Li, X.; Wang, H.; Xie, D.; Li, X.; Bai, Y. Land cover classification based on fused data from GF-1 and MODIS NDVI time series. Remote Sens. 2016, 8, 741. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.L.; Hoffman, M.T.; Roberts, W. Long-term trends in vegetation phenology and productivity over Namaqualand using the GIMMS AVHRR NDVI3g data from 1982 to 2011. S. Afr. J. Bot. 2017, 111, 76–85. [Google Scholar] [CrossRef]
- Ghosh, S.; Mishra, D. Analyzing the long-term phenological trends of salt marsh ecosystem across Coastal Louisiana. Remote Sens. 2017, 9, 1340. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: https://www.R-project.org/ (accessed on 1 March 2021).
- Bureau of Meteorology. Tropical Cyclone Reports. Available online: http://www.bom.gov.au/cyclone/tropical-cyclone-knowledge-centre/history/past-tropical-cyclones/ (accessed on 1 March 2021).
- Commonwealth of Australia. Reef 2050 Long Term Sustainability Plan 2018; Commonwealth of Australia: Canberra, Australia, 2018. [Google Scholar]
- Bahuguna, A.; Chauhan, H.B.; Sen Sarma, K.; Bhattacharya, S.; Ashutosh, S.; Pandey, C.N.; Thangaradjou, T.; Gnanppazham, L.; Selvam, V.; Nayak, S.R. Mangrove inventory of India at community level. Natl. Acad. Sci. Lett. 2013, 36, 67–77. [Google Scholar] [CrossRef]
- Semeniuk, V.; Cresswell, I.D. Australian mangroves: Anthropogenic impacts by industry, agriculture, ports, and urbanisation. In Threats to Mangrove Forests: Hazards, Vulnerability, and Management; Makowski, C., Finkl, C.W., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 173–197. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Sippo, J.Z.; Lovelock, C.E.; Santos, I.R.; Sanders, C.J.; Maher, D.T. Mangrove mortality in a changing climate: An overview. Estuar. Coast. Shelf Sci. 2018, 215, 241–249. [Google Scholar] [CrossRef]
- Hamunyela, E.; Verbesselt, J.; Herold, M. Using spatial context to improve early detection of deforestation from Landsat time series. Remote Sens. Environ. 2016, 172, 126–138. [Google Scholar] [CrossRef]
- Hirschmugl, M.; Gallaun, H.; Dees, M.; Datta, P.; Deutscher, J.; Koutsias, N.; Schardt, M. Methods for mapping forest disturbance and degradation from optical Earth observation data: A review. Curr. For. Rep. 2017, 3, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Pimple, U.; Simonetti, D.; Sitthi, A.; Pungkul, S.; Leadprathom, K.; Skupek, H.; Som-ard, J.; Gond, V.; Towprayoon, S. Google Earth Engine based three decadal Landsat imagery analysis for mapping of mangrove forests and its surroundings in the Trat Province of Thailand. J. Commun. 2018, 6, 247–264. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Xu, N.; Liang, J.; Li, Z.; Chen, L.; Zhao, F. Advancing the mapping of mangrove forests at national-scale using Sentinel-1 and Sentinel-2 time-series data with Google Earth Engine: A case study in China. Remote Sens. 2020, 12, 3120. [Google Scholar] [CrossRef]
- Glenn, E.P.; Huete, A.R.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: What vegetation indices can and cannot tell us about landscape. Sensors 2008, 8, 2136–2160. [Google Scholar] [CrossRef] [Green Version]
- Elmahdy, S.I.; Ali, T.A.; Mohamed, M.M.; Howari, F.M.; Abouleish, M.; Simonet, D. Spatiotemporal mapping and monitoring of mangrove forests changes from 1990 to 2019 in the Northern Emirates, UAE using Random Forest, Kernel Logistic Regression and Naive Bayes Tree Models. Front. Environ. Sci. 2020, 8, 102. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Peng, Y.; Masek, J.G.; Rundquist, D.C.; Verma, S.; Suyker, A.; Baker, J.M.; Hatfield, J.L.; Meyers, T. Remote estimation of crop gross primary production with Landsat data. Remote Sens. Environ. 2012, 121, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Adame, M.F.; Bennion, V.; Hayes, M.; Reef, R.; Santini, N.; Cahoon, D.R. Sea level and turbidity controls on mangrove soil surface elevation change. Estuar. Coast. Shelf Sci. 2015, 153, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Carrao, H.; Gonalves, P.; Caetano, M. A nonlinear harmonic model for fitting satellite image time series: Analysis and prediction of land cover dynamics. IEEE Trans. Geosci. Remote Sens. 2010, 48, 1919–1930. [Google Scholar] [CrossRef]
- Bi, J.; Knyazikhin, Y.; Choi, S.; Park, T.; Barichivich, J.; Ciais, P.; Fu, R.; Ganguly, S.; Hall, F.; Hilker, T.; et al. Sunlight mediated seasonality in canopy structure and photosynthetic activity of Amazonian rainforests. Environ. Res. Lett. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Albert, L.P.; Lopes, A.P.; Restrepo-Coupe, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. Science 2016, 351, 972–976. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.I.; Dixon, P.; Zagorskis, I. Phenology and litter production in the mangrove genus Xylocarpus along rainfall and temperature gradients in tropical Australia. Mar. Freshw. Res. 2021, 72, 551–562. [Google Scholar] [CrossRef]
- Younes, N.; Northfield, T.D.; Joyce, K.E.; Maier, S.W.; Duke, N.C.; Lymburner, L. A novel approach to modelling mangrove phenology from satellite images: A case study from Northern Australia. Remote Sens. 2020, 12, 4008. [Google Scholar] [CrossRef]
- Pastor-Guzman, J.; Dash, J.; Atkinson, P.M. Remote sensing of mangrove forest phenology and its environmental drivers. Remote Sens. Environ. 2018, 205, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Songsom, V.; Koedsin, W.; Ritchie, R.J.; Huete, A. Mangrove phenology and environmental drivers derived from remote sensing in Southern Thailand. Remote Sens. 2019, 11, 955. [Google Scholar] [CrossRef] [Green Version]
- Duke, N.C.; Field, C.; Mackenzie, J.R.; Meynecke, J.-O.; Wood, A.L. Rainfall and its possible hysteresis effect on the proportional cover of tropical tidal-wetland mangroves and saltmarsh–saltpans. Mar. Freshw. Res. 2019, 70, 1047–1055. [Google Scholar] [CrossRef]
- Santini, N.S.; Reef, R.; Lockington, D.A.; Lovelock, C.E. The use of fresh and saline water sources by the mangrove Avicennia marina. Hydrobiologia 2014, 745, 59–68. [Google Scholar] [CrossRef]
- Eklundh, L.; Jönsson, P. TIMESAT: A software package for time-series processing and assessment of vegetation dynamics. In Remote Sensing Time Series: Revealing Land Surface Dynamics; Kuenzer, C., Dech, S., Wagner, W., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 141–158. [Google Scholar] [CrossRef]
- Coupland, G.T.; Paling, E.I.; McGuinness, K.A. Vegetative and reproductive phenologies of four mangrove species from northern Australia. Aust. J. Bot. 2005, 53, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Morton, D.C.; Nagol, J.; Carabajal, C.C.; Rosette, J.; Palace, M.; Cook, B.D.; Vermote, E.F.; Harding, D.J.; North, P.R. Amazon forests maintain consistent canopy structure and greenness during the dry season. Nature 2014, 506, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Enwright, N.M.; Day, R.H.; Gabler, C.A.; Stagg, C.L.; Grace, J.B. Beyond just sea-level rise: Considering macroclimatic drivers within coastal wetland vulnerability assessments to climate change. Glob. Chang. Biol. 2016, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Negrón-Juárez, R.I.; Chambers, J.Q. Remote sensing and statistical analysis of the effects of hurricane María on the forests of Puerto Rico. Remote Sens. Environ. 2020, 247, 111940. [Google Scholar] [CrossRef]
- Das, C.S.; Mallick, D.; Mandal, R.N. Mangrove forests in changing climate: A global overview. J. Indian Soc. Coast. Agric. Res. 2020, 38, 104–124. [Google Scholar]
- Rossi, S.; Soares, M.d.O. Effects of El NiÑo on the coastal ecosystems and their related services. Mercator 2017, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Asbridge, E.; Lucas, R.; Ticehurst, C.; Bunting, P. Mangrove response to environmental change in Australia’s Gulf of Carpentaria. Ecol. Evol. 2016, 6, 3523–3539. [Google Scholar] [CrossRef] [Green Version]
- Hoque, M.A.-A.; Phinn, S.; Roelfsema, C. A systematic review of tropical cyclone disaster management research using remote sensing and spatial analysis. Ocean Coast. Manag. 2017, 146, 109–120. [Google Scholar] [CrossRef]
- Foody, G.M. Assessing the accuracy of land cover change with imperfect ground reference data. Remote Sens. Environ. 2010, 114, 2271–2285. [Google Scholar] [CrossRef] [Green Version]
- Younes Cárdenas, N.; Joyce, K.E.; Maier, S.W. Monitoring mangrove forests: Are we taking full advantage of technology? Int. J. Appl. Earth Obs. Geoinf. 2017, 63, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bush, E.R.; Bunnefeld, N.; Dimoto, E.; Dikangadissi, J.-T.; Jeffery, K.; Tutin, C.; White, L.; Abernethy, K.A. Towards effective monitoring of tropical phenology: Maximizing returns and reducing uncertainty in long-term studies. Biotropica 2018, 50, 455–464. [Google Scholar] [CrossRef]
- Bush, E.R.; Abernethy, K.A.; Jeffery, K.; Tutin, C.; White, L.; Dimoto, E.; Dikangadissi, J.T.; Jump, A.S.; Bunnefeld, N.; Freckleton, R. Fourier analysis to detect phenological cycles using long-term tropical field data and simulations. Methods Ecol. Evol. 2017, 8, 530–540. [Google Scholar] [CrossRef]
- Cresswell, I.D.; Semeniuk, V. Australian mangroves: Their distribution and protection. In Threats to Mangrove Forests: Hazards, Vulnerability, and Management; Makowski, C., Finkl, C.W., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 3–22. [Google Scholar] [CrossRef]
- Friess, D.A.; Rogers, K.; Lovelock, C.E.; Krauss, K.W.; Hamilton, S.E.; Lee, S.Y.; Lucas, R.; Primavera, J.; Rajkaran, A.; Shi, S. The state of the world’s mangrove forests: Past, present, and future. Annu. Rev. Environ. Resour. 2019, 44, 89–115. [Google Scholar] [CrossRef] [Green Version]
- Ramsar Convention on Wetlands. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Ramsar Convention Secretariat: Gland, Switzerland, 2018. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mangrove Extent 2009 (Ha) | Mangrove Extent 2019 (Ha) | Decrease from 2009 to 2019 (Ha) | Percent Area Decrease from 2009 to 2019 (%) |
|---|---|---|---|
| 63,990 | 62,510 | −1480 | −2.31 |
| Land Class Name | Producer’s Accuracy (%) | User’s Accuracy (%) | ||
|---|---|---|---|---|
| 2009 | 2019 | 2009 | 2019 | |
| Mangrove forest | 0.99 | 0.96 | 0.98 | 0.95 |
| Non-Mangrove Forest | 0.95 | 0.99 | 0.96 | 0.99 |
| Overall accuracy | 0.98 | 0.98 | ||
| Kappa coefficient | 0.95 | 0.95 | ||
| Seas. | Startt. | Endt. | Length | Baseval. | Peakt. | Peakval. | Ampl. | L.deriv. | R.deriv. | L.integral | S.integral | Startval. | Endval. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 11.9 | 21.12 | 9.222 | 0.721 | 17.34 | 0.8539 | 0.1329 | 0.03167 | 0.03633 | 9.555 | 0.9028 | 0.7173 | 0.7779 |
| 2 | 26.35 | 33.71 | 7.36 | 0.7283 | 30.53 | 0.8131 | 0.08484 | 0.01231 | 0.04226 | 7.002 | 0.4479 | 0.7697 | 0.7208 |
| 3 | 36.6 | 46 | 9.405 | 0.7184 | 41.26 | 0.8337 | 0.1153 | 0.0316 | 0.02131 | 9.411 | 0.7901 | 0.7249 | 0.7581 |
| 4 | 49.65 | 57.38 | 7.728 | 0.7438 | 53.56 | 0.8246 | 0.08079 | 0.02252 | 0.02102 | 7.876 | 0.438 | 0.7563 | 0.7637 |
| 5 | 59.89 | 69.77 | 9.878 | 0.7227 | 63.97 | 0.8039 | 0.08119 | 0.01741 | 0.01763 | 9.234 | 0.5616 | 0.7596 | 0.7184 |
| 6 | 72.56 | 84.11 | 11.55 | 0.7069 | 77.04 | 0.7975 | 0.0906 | 0.02542 | 0.01493 | 10.56 | 0.6589 | 0.7171 | 0.733 |
| 7 | 86 | 94.3 | 8.3 | 0.701 | 90.26 | 0.8121 | 0.1111 | 0.02122 | 0.03094 | 8.354 | 0.6427 | 0.7359 | 0.7106 |
| 8 | 97.41 | 105.2 | 7.776 | 0.6941 | 102.1 | 0.8428 | 0.1487 | 0.02885 | 0.04959 | 7.722 | 0.7812 | 0.7167 | 0.7309 |
| 9 | 108.1 | 117.4 | 9.345 | 0.6692 | 113.2 | 0.8055 | 0.1363 | 0.02137 | 0.04906 | 8.29 | 0.9292 | 0.7234 | 0.6694 |
| 10 | 119.7 | 129.4 | 9.732 | 0.6548 | 126.1 | 0.8275 | 0.1727 | 0.02239 | 0.0459 | 8.826 | 0.9689 | 0.6738 | 0.7048 |
| 11 | 132.3 | 144 | 11.73 | 0.7202 | 137.4 | 0.8444 | 0.1243 | 0.03738 | 0.01137 | 10.41 | 1.043 | 0.7082 | 0.7818 |
| 12 | 146.9 | 154.2 | 7.284 | 0.7394 | 150.4 | 0.8503 | 0.111 | 0.02247 | 0.03217 | 7.939 | 0.5453 | 0.783 | 0.7401 |
| 13 | 157.4 | 164.9 | 7.516 | 0.7331 | 161.3 | 0.8385 | 0.1054 | 0.03491 | 0.02762 | 7.17 | 0.5718 | 0.7378 | 0.7706 |
| 14 | 169.2 | 178.5 | 9.321 | 0.7749 | 173.6 | 0.8965 | 0.1216 | 0.0348 | 0.02082 | 9.346 | 0.8217 | 0.7822 | 0.8163 |
| 15 | 181.6 | 190.5 | 8.946 | 0.8042 | 186.1 | 0.8529 | 0.04878 | 0.01479 | 0.01081 | 9.162 | 0.3164 | 0.8076 | 0.8202 |
| 16 | 193.8 | 201.7 | 7.847 | 0.8168 | 198.1 | 0.8744 | 0.05757 | 0.01329 | 0.01511 | 8.472 | 0.3035 | 0.8245 | 0.8322 |
| 17 | 205.6 | 213.2 | 7.618 | 0.8043 | 209.8 | 0.8898 | 0.08543 | 0.01478 | 0.03051 | 8.493 | 0.4493 | 0.8352 | 0.8076 |
| 18 | 215.2 | 224.5 | 9.297 | 0.7786 | 220.8 | 0.8359 | 0.05734 | 0.007241 | 0.01802 | 8.956 | 0.3909 | 0.7968 | 0.7833 |
| 19 | 227.8 | 238.2 | 10.41 | 0.7684 | 235 | 0.8755 | 0.1071 | 0.03052 | 0.03727 | 10.58 | 0.5864 | 0.7912 | 0.7884 |
| 20 | 242.6 | 249.4 | 6.795 | 0.7457 | 246.2 | 0.9123 | 0.1666 | 0.03448 | 0.05107 | 7.449 | 0.7378 | 0.7958 | 0.7623 |
| 21 | 251.6 | 261.5 | 9.971 | 0.7633 | 254.9 | 0.8783 | 0.115 | 0.03907 | 0.008088 | 9.942 | 0.7833 | 0.7555 | 0.817 |
| Response Variable | Explanatory Variable Pearson Correlation Coefficient r | Adjusted R2 |
|---|---|---|
| Season (year) | Length of season −0.91 | 0.818 |
| Season (year) | Seasonal amplitude −0.78 | 0.597 |
| Season (year) | Rate of senescence −0.9 | 0.793 |
| Season (year) | Peak photosynthetic activity 0.7 | 0.462 |
| Season (year) | Seasonal productivity −0.88 | 0.775 |
| Length of season | Period of peak −0.91 photosynthetic activity | 0.812 |
| Length of season | Rate of senescence −0.9 | 0.721 |
| Growth rate | Seasonal amplitude 0.87 | 0.755 |
| Seasonal productivity | Rate of senescence 0.89 | 0.771 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamberlain, D.A.; Phinn, S.R.; Possingham, H.P. Mangrove Forest Cover and Phenology with Landsat Dense Time Series in Central Queensland, Australia. Remote Sens. 2021, 13, 3032. https://doi.org/10.3390/rs13153032
Chamberlain DA, Phinn SR, Possingham HP. Mangrove Forest Cover and Phenology with Landsat Dense Time Series in Central Queensland, Australia. Remote Sensing. 2021; 13(15):3032. https://doi.org/10.3390/rs13153032
Chicago/Turabian StyleChamberlain, Debbie A., Stuart R. Phinn, and Hugh P. Possingham. 2021. "Mangrove Forest Cover and Phenology with Landsat Dense Time Series in Central Queensland, Australia" Remote Sensing 13, no. 15: 3032. https://doi.org/10.3390/rs13153032
APA StyleChamberlain, D. A., Phinn, S. R., & Possingham, H. P. (2021). Mangrove Forest Cover and Phenology with Landsat Dense Time Series in Central Queensland, Australia. Remote Sensing, 13(15), 3032. https://doi.org/10.3390/rs13153032