Forest Drought Response Index (ForDRI): A New Combined Model to Monitor Forest Drought in the Eastern United States

 ,
,  , ,
, ,  , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
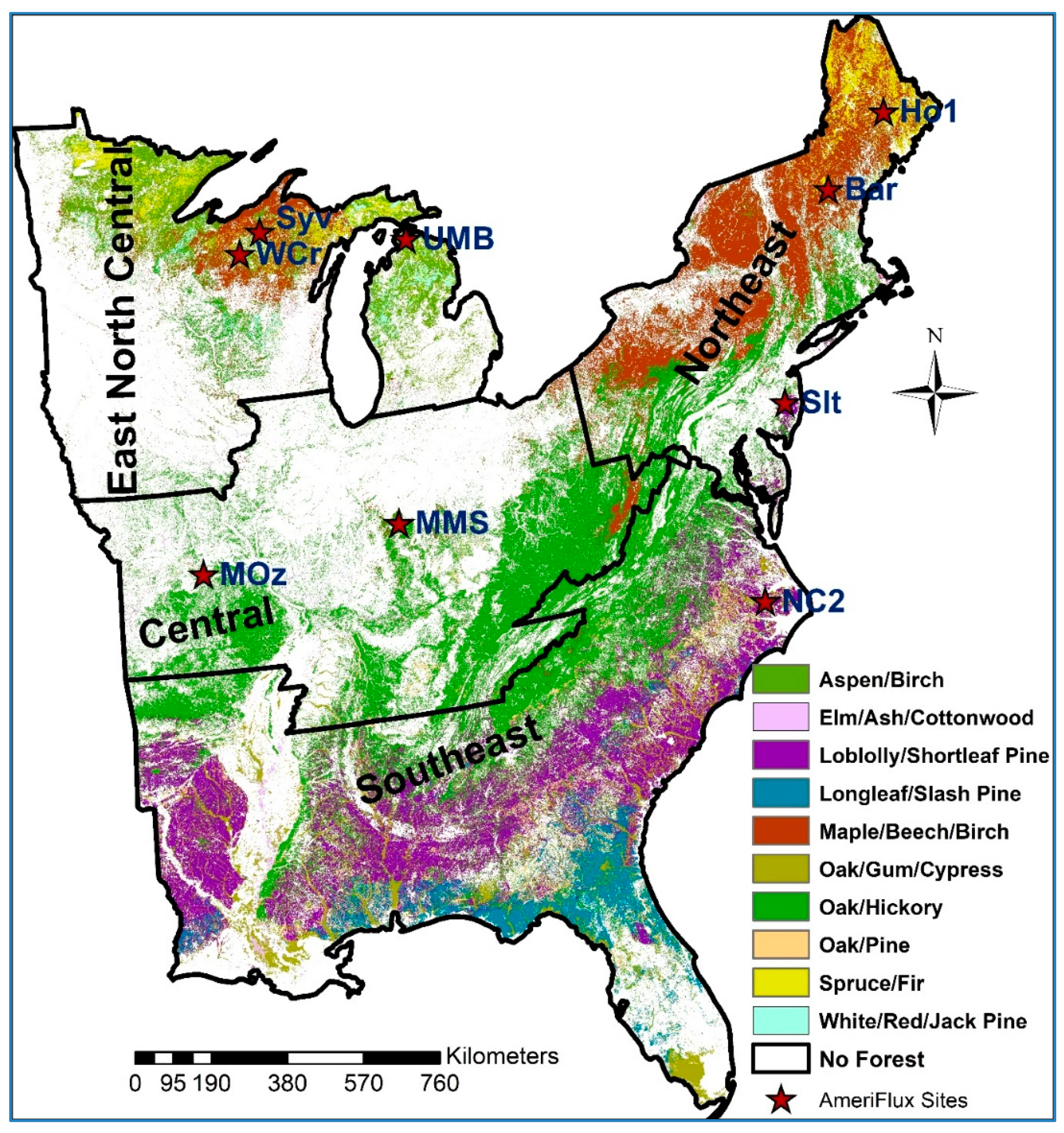
2.1. Study Area Forest Group Type Coverage by Climate Region
2.2. Data Used in ForDRI Model Development
2.2.1. MODIS-based Normalized Difference Vegetation Index (NDVI)
2.2.2. Standardized Precipitation Index (SPI)
2.2.3. Standardized Precipitation Evapotranspiration Index (SPEI)
2.2.4. Evaporative Demand Drought Index (EDDI)
2.2.5. Ground Water Storage (GWS)
2.2.6. Palmer Drought Severity Index (PDSI) and Palmer Z Index (PZI)
2.2.7. Noah Soil Moisture (SM)
2.2.8. Vapor Pressure Deficit
2.2.9. National Forest Groups and Types
2.2.10. Bowen Ratio Data to Compare with ForDRI at Nine AmeriFlux Sites
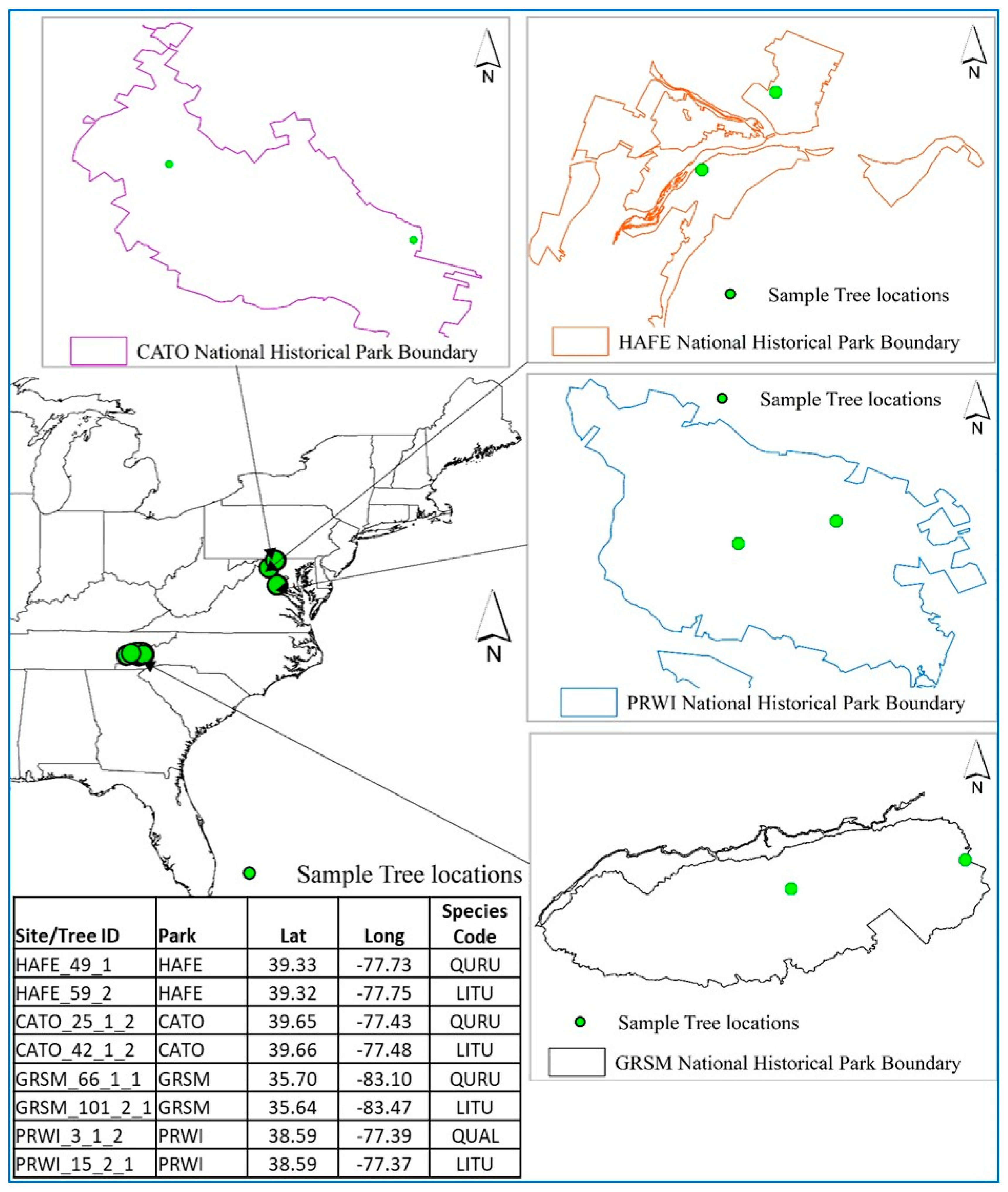
2.2.11. Tree Ring Data for Evaluation
2.3. Methods
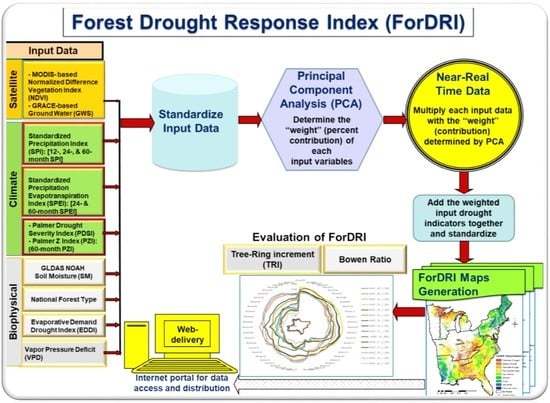
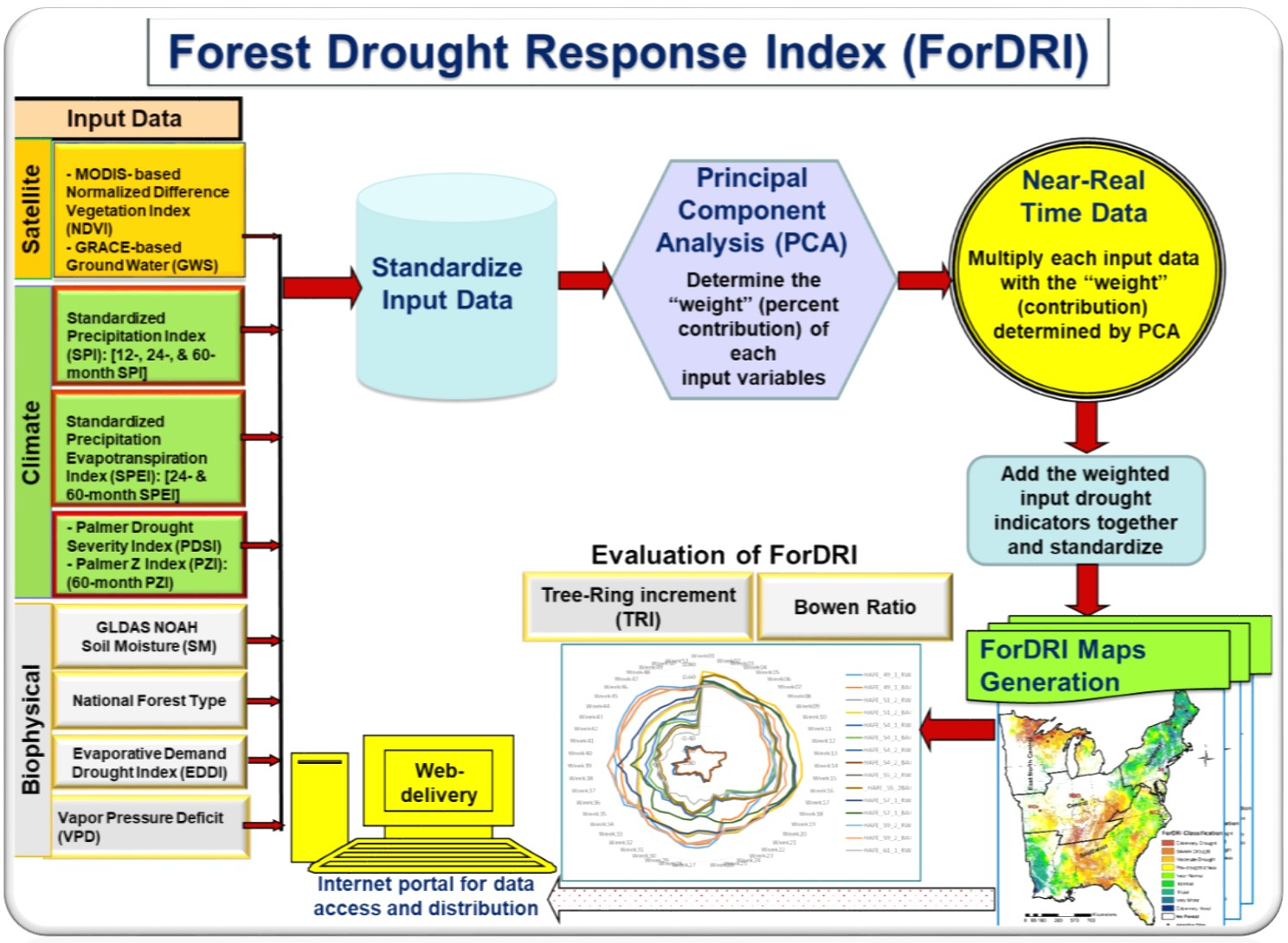
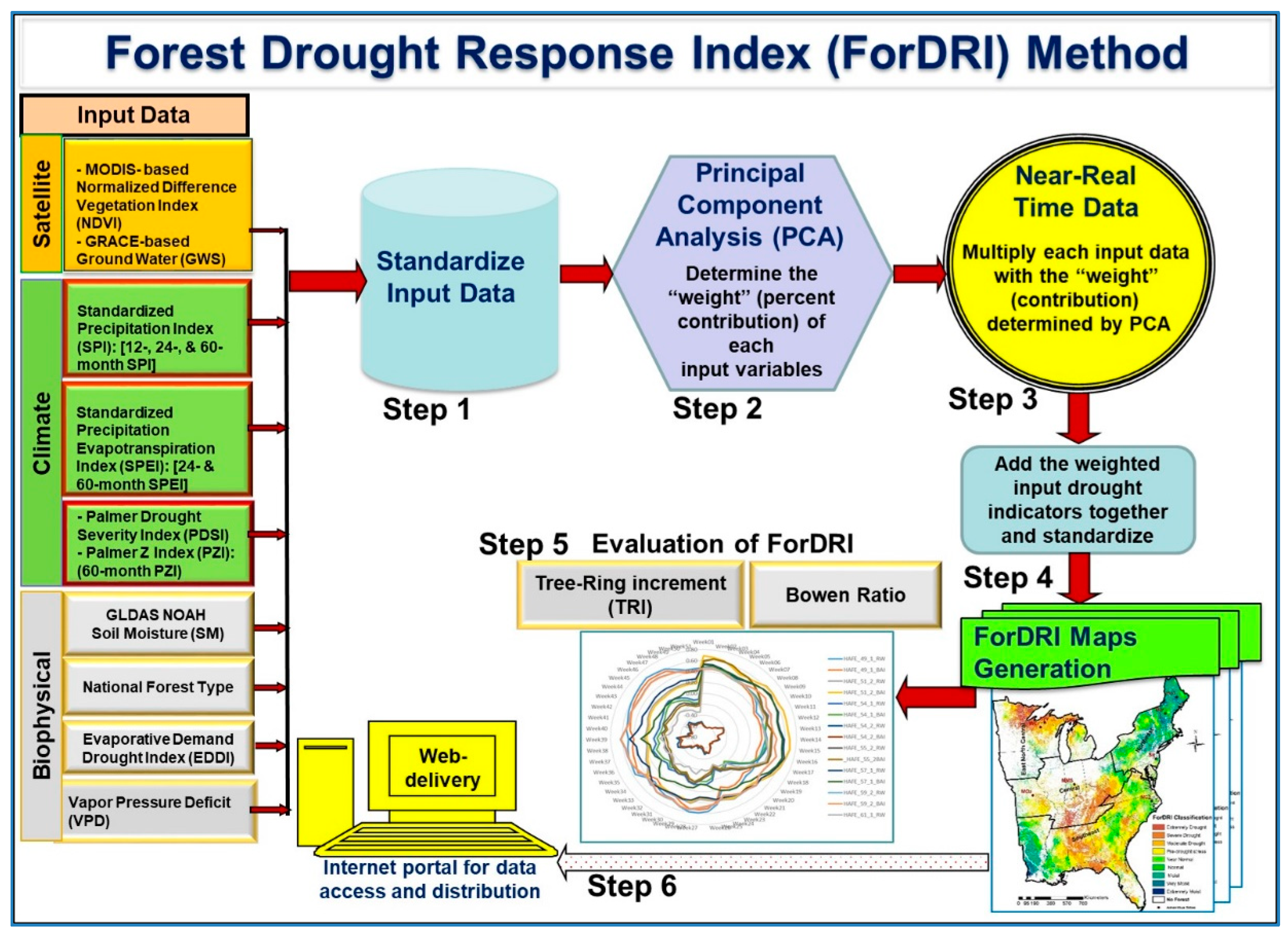
2.3.1. ForDRI Model Development
2.3.2. Evaluation Method/Approaches for ForDRI (Both Qualitative and Quantitative Approaches)
3. Results
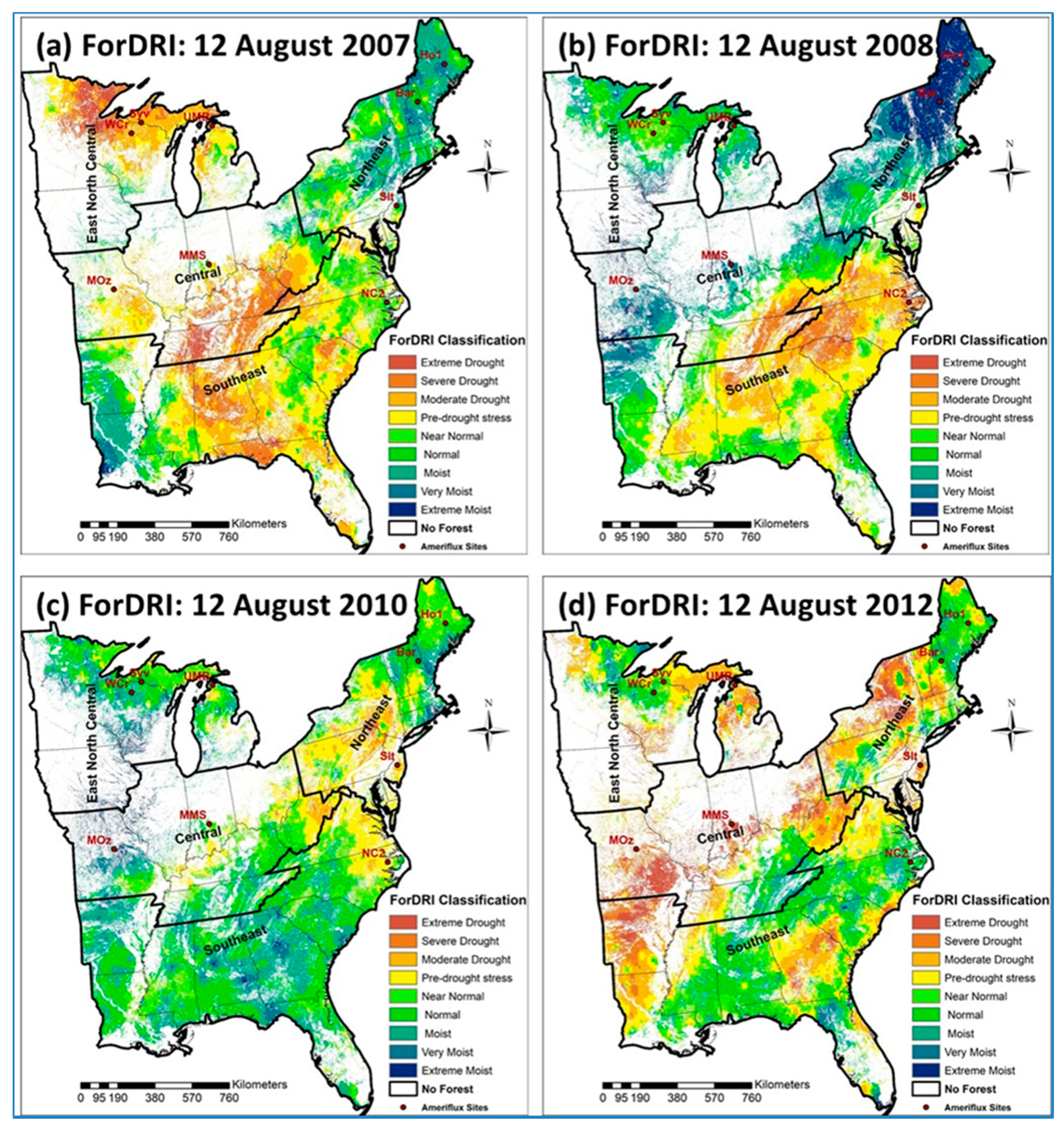
3.1. ForDRI Maps for Selected Drought Years
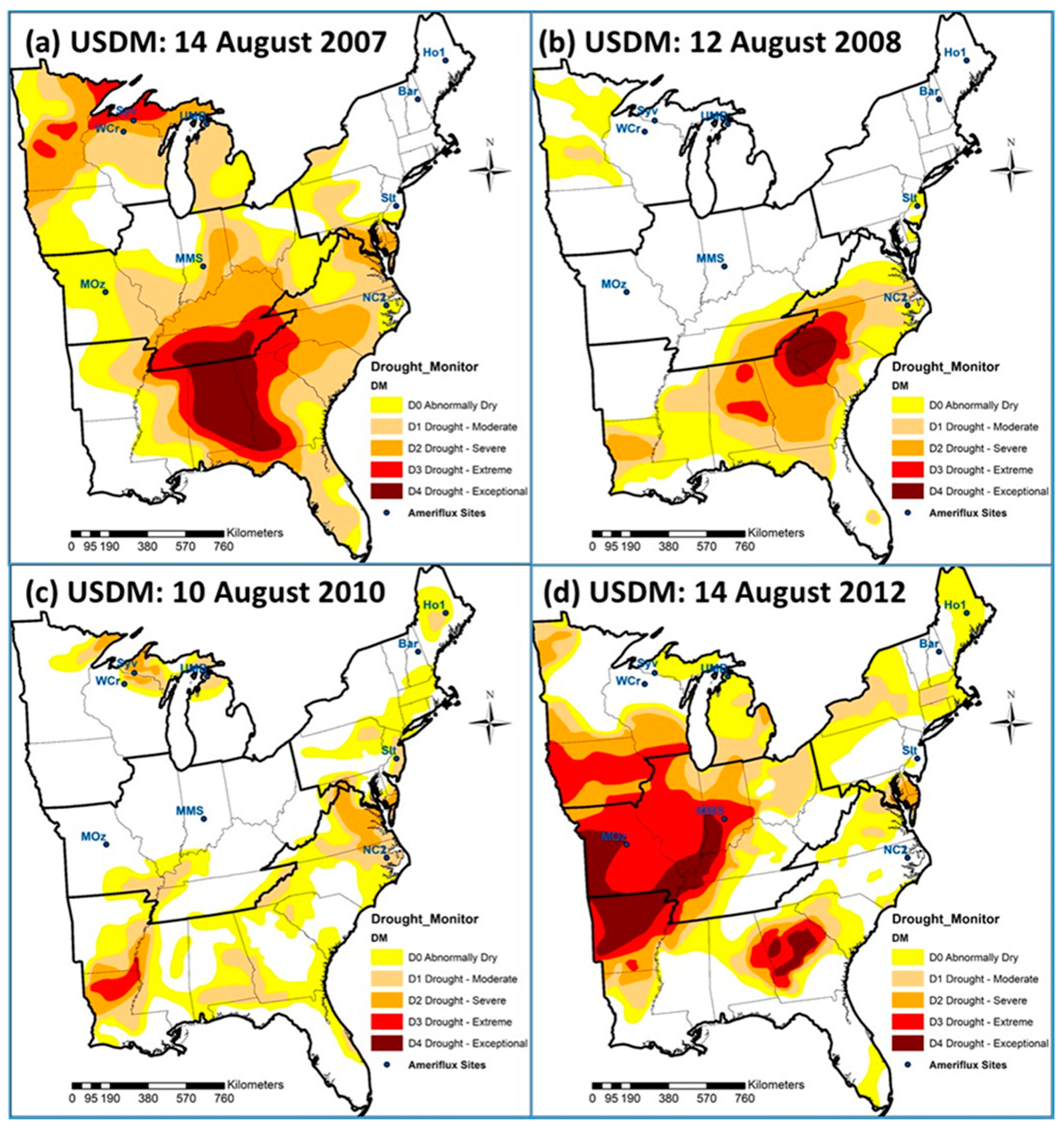
3.2. Comparison of ForDRI with U.S. Drought Monitor (USDM)
3.3. Evaluating ForDRI with Bowen Ratio
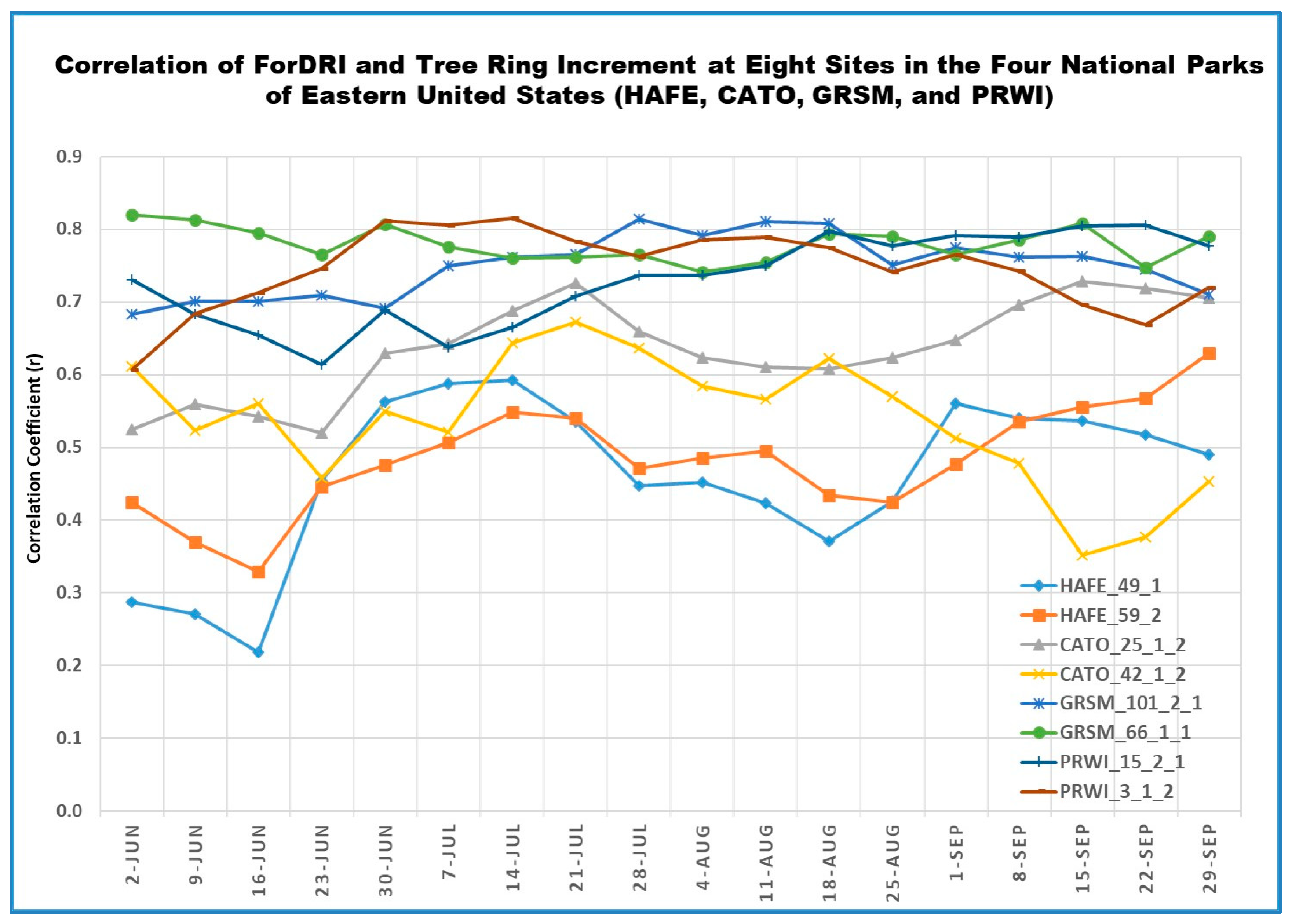
3.4. Evaluating ForDRI with Tree Ring Increments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rigden, A.J.; Mueller, N.D.; Holbrook, N.M.; Pillai, N.; Huybers, P. Combined influence of soil moisture and atmospheric evaporative demand is important for accurately predicting US maize yields. Nat. Food 2020, 1, 127–133. [Google Scholar] [CrossRef]
- Manzoni, S.; Katul, G.; Porporato, A. A dynamical system perspective on plant hydraulic failure. Water Resour. Res. 2014, 50, 5170–5183. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest growth responses to drought at short-and long-term scales in Spain: Squeezing the stress memory from tree rings. Front. Ecol. Evol. 2018, 6, 1–11. [Google Scholar] [CrossRef]
- Yin, J.; Bauerle, T.L. A global analysis of plant recovery performance from water stress. Oikos 2017, 126, 1377–1388. [Google Scholar] [CrossRef]
- Matheny, A.M.; Fiorella, R.P.; Bohrer, G.; Poulsen, C.J.; Morin, T.H.; Wunderlich, A.; Vogel, C.S.; Curtis, P.S. Contrasting strategies of hydraulic control in two codominant temperate tree species. Ecohydrology 2017, 10, e1815. [Google Scholar] [CrossRef]
- Roman, D.T.; Novick, K.A.; Brzostek, E.R.; Dragoni, D.; Rahman, F.; Phillips, R.P. The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought. Oecologia 2015, 179, 641–654. [Google Scholar] [CrossRef]
- Plaut, J.A.; Yepez, E.A.; Hill, J.; Pangle, R.; Sperry, J.S.; Pockman, W.T.; Mcdowell, N.G. Hydraulic limits preceding mortality in a piñon–juniper woodland under experimental drought. PlantCell Environ. 2012, 35, 1601–1617. [Google Scholar] [CrossRef]
- Sanchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, A.; Vila-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sanguesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Cailleret, M.; Jansen, S.; Robert, E.M.; Desoto, L.; Aakala, T.; Antos, J.A.; Beikircher, B.; Bigler, C.; Bugmann, H.; Caccianiga, M.; et al. A synthesis of radial growth patterns preceding tree mortality. Glob. Chang. Biol. 2017, 23, 1675–1690. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef]
- Ruffault, J.; Martin-StPaul, N.; Pimont, F.; Dupuy, J.L. How well do meteorological drought indices predict live fuel moisture content (LFMC)? An assessment for wildfire research and operations in Mediterranean ecosystems. Agric. For. Meteorol. 2018, 262, 391–401. [Google Scholar] [CrossRef]
- McKee, T.B.; Doesken, N.J.; Kleist, J. Drought Monitoring with Multiple Time Scales. In Proceedings of the 9th Conference on Applied Climatology, Dallas, TX, USA, 15–20 January 1995; American Meteorological Society: Boston, MA, USA. [Google Scholar]
- Wells, N.; Goddard, S.; Hayes, M.J. A self-calibrating Palmer drought severity index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological Drought; Research Paper, No. 45; US Department of Commerce, Weather Bureau: Washington, DC, USA, 1965; Volume 30, p. 58.
- Keetch, J.J.; Byram, G.M. A Drought Index for Forest Fire Control; US Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1968; Volume 38.
- Koch, F.H.; Smith, W.D.; Coulston, J.W. An improved method for standardized mapping of drought conditions. In Forest Health Monitoring: National Status, Trends, and Analysis 2010. Gen. Tech. Rep. SRS-GTR-176; Potter, K.M., Conkling, B.L., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2013; pp. 67–83. [Google Scholar]
- Koch, F.H.; Smith, W.D.; Coulston, J.W. Drought patterns in the conterminous United States and Hawaii. In Forest Health Monitoring: National Status, Trends, and Analysis 2012. Gen. Tech. Rep. SRSGTR-198; Potter, K.M., Conkling, B.L., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2014; pp. 49–72. [Google Scholar]
- Koch, F.H.; Smith, W.D.; Coulston, J.W. Drought patterns in the conterminous United States, 2012. In Forest Health Monitoring: National Status, Trends, and Analysis 2013. Gen. Tech. Rep. SRSGTR-207; Potter, K.M., Conkling, B.L., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2015; pp. 55–69. [Google Scholar]
- Saleska, S.R.; Didan, K.; Huete, A.R.; da Rocha, H.R. Amazon forests green-up during 2005 drought. Science 2007, 318, 612. [Google Scholar] [CrossRef]
- Anderson, M.C.; Hain, C.; Wardlow, B.; Pimstein, A.; Mecikalski, J.R.; Kustas, W.P. Evaluation of drought indices based on thermal remote sensing of evapotranspiration over the continental United States. J. Clim. 2011, 24, 2025–2044. [Google Scholar] [CrossRef]
- Asner, G.P.; Alencar, A. Drought impacts on the Amazon forest: The remote sensing perspective. New Phytol. 2010, 187, 569–578. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Samanta, A.; Ganguly, S.; Myneni, R.B. MODIS enhanced vegetation index data do not show greening of Amazon forests during the 2005 drought. New Phytol. 2011, 189, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, C.; Li, W.; Fang, X.; Zhang, T.; Zhu, Q.; Chen, H.; Zhao, P. Monitoring and estimating drought-induced impacts on forest structure, growth, function, and ecosystem services using remote-sensing data: Recent progress and future challenges. Environ. Rev. 2013, 21, 103–115. [Google Scholar] [CrossRef]
- AghaKouchak, A.; Farahmand, A.; Melton, F.S.; Teixeira, J.; Anderson, M.C.; Wardlow, B.D.; Hain, C.R. Remote sensing of drought: Progress, challenges and opportunities. Rev. Geophys. 2015, 53, 452–480. [Google Scholar] [CrossRef]
- Norman, S.P.; Koch, F.H.; Hargrove, W.W. Review of broad-scale drought monitoring of forests: Toward an integrated data mining approach. For. Ecol. Manag. 2016, 380, 346–358. [Google Scholar] [CrossRef]
- Svoboda, M.; LeComte, D.; Hayes, M.; Heim, R.; Gleason, K.; Angel, J.; Rippey, B.; Tinker, R.; Palecki, M.; Stooksbury, D.; et al. The drought monitor. Bull. Am. Meteorol. Soc. 2002, 83, 1181–1190. [Google Scholar] [CrossRef]
- NDMC. U.S. Drought Monitor. 2020. Available online: https://droughtmonitor.unl.edu/About.aspx (accessed on 3 September 2020).
- Iverson, L.R.; Prasad, A.M. Predicting abundance of 80 tree species following climate change in the eastern United States. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- USDA Forest Service. National Forest Type Dataset. 2020. Available online: https://data.fs.usda.gov/geodata/rastergateway/forest_type/ (accessed on 3 September 2020).
- Ruefenacht, B.; Finco, M.V.; Nelson, M.D.; Czaplewski, R.; Helmer, E.H.; Blackard, J.A.; Holden, G.R.; Lister, A.J.; Salajanu, D.; Weyermann, D.; et al. Conterminous US and Alaska forest type mapping using forest inventory and analysis data. Photogramm. Eng. Remote Sens. 2008, 74, 1379–1388. [Google Scholar] [CrossRef]
- USGS. EROS Moderate Resolution Imaging Spectroradiometer (eMODIS) Digital Object Identifier (DOI) Number: /10.5066/F7H41PNT). 2020. Available online: https://www.usgs.gov/centers/eros/science/usgs-eros-archive-vegetation-monitoring-eros-moderate-resolution-imaging?qt-science_center_objects=0#qt-science_center_objects (accessed on 3 September 2020).
- Edwards, D.C.; McKee, T.B. “Characteristics of 20th Century Drought in the United States at Multiple Time Scales,” Climatology Report Number 97-2, Department of Atmospheric Science; Colorado State University: Fort Collins, CO, USA, 1997. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Hobbins, M.T.; Wood, A.; McEvoy, D.J.; Huntington, J.L.; Morton, C.; Anderson, M.; Hain, C. The evaporative demand drought index. Part I: Linking drought evolution to variations in evaporative demand. J. Hydrometeorol. 2016, 17, 1745–1761. [Google Scholar] [CrossRef]
- McEvoy, D.J.; Huntington, J.L.; Hobbins, M.T.; Wood, A.; Morton, C.; Anderson, M.; Hain, C. The evaporative demand drought index. Part II: CONUS-wide assessment against common drought indicators. J. Hydrometeorol. 2016, 17, 1763–1779. [Google Scholar] [CrossRef]
- Bhanja, S.N.; Mukherjee, A.; Rodell, M. Groundwater storage change detection from in situ and GRACE-based estimates in major river basins across India. Hydrol. Sci. J. 2020, 65, 650–659. [Google Scholar] [CrossRef]
- Li, B.; Rodell, M.; Kumar, S.; Beaudoing, H.K.; Getirana, A.; Zaitchik, B.F.; de Goncalves, L.G.; Cossetin, C.; Bhanja, S.; Mukherjee, A.; et al. Global GRACE data assimilation for groundwater and drought monitoring: Advances and challenges. Water Resour. Res. 2019, 55, 7564–7586. [Google Scholar] [CrossRef]
- NASA GSFC Hydrological Sciences Laboratory—Nasa Gesdisc Data Archive, 2020. Available online: https://hydro1.gesdisc.eosdis.nasa.gov/data/GLDAS/GLDAS_CLSM025_DA1_D.2.2/ (accessed on 3 September 2020).
- Keyantash, J.; Dracup, J.A. The quantification of drought: An evaluation of drought indices. Bull. Am. Meteorol. Soc. 2002, 83, 1167–1180. [Google Scholar] [CrossRef]
- Nearing, G.S.; Mocko, D.M.; Peters-Lidard, C.D.; Kumar, S.V.; Xia, Y. Benchmarking NLDAS-2 soil moisture and evapotranspiration to separate uncertainty contributions. J. Hydrometeorol. 2016, 17, 745–759. [Google Scholar] [CrossRef]
- Xia, Y.; Hao, Z.; Shi, C.; Li, Y.; Meng, J.; Xu, T.; Wu, X.; Zhang, B. Regional and global land data assimilation systems: Innovations, challenges, and prospects. J. Meteorol. Res. 2019, 33, 159–189. [Google Scholar] [CrossRef]
- Kumar, S.V.; Peters-Lidard, C.D.; Mocko, D.; Reichle, R.; Liu, Y.; Arsenault, K.R.; Xia, Y.; Ek, M.; Riggs, G.; Livneh, B.; et al. Assimilation of remotely sensed soil moisture and snow depth retrievals for drought estimation. J. Hydrometeorol. 2014, 15, 2446–2469. [Google Scholar] [CrossRef]
- Cai, X.; Yang, Z.L.; Xia, Y.; Huang, M.; Wei, H.; Leung, L.R.; Ek, M.B. Assessment of simulated water balance from Noah, Noah-MP, CLM, and VIC over CONUS using the NLDAS test bed. J. Geophys. Res. Atmos. 2014, 119, 13751–13770. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Parinussa, R.M.; Dorigo, W.A.; De Jeu, R.A.; Wagner, W.; Van Dijk, A.I.J.M.; McCabe, M.F.; Evans, J.P. Developing an improved soil moisture dataset by blending passive and active microwave satellite-based retrievals. Hydrol. Earth Syst. Sci. 2011, 15, 425–436. [Google Scholar] [CrossRef]
- NOAA. NLDAS Drought Monitor Soil Moisture. 2020. Available online: https://www.emc.ncep.noaa.gov/mmb/nldas/drought/ (accessed on 3 September 2020).
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Sinclair, T.R.; Allen, L.H., Jr. Transpiration responses to vapor pressure deficit in well watered‘slow-wilting’and commercial soybean. Environ. Exp. Bot. 2007, 61, 145–151. [Google Scholar] [CrossRef]
- Li, P.; Omani, N.; Chaubey, I.; Wei, X. Evaluation of Drought Implications on Ecosystem Services: Freshwater Provisioning and Food Provisioning in the Upper Mississippi River Basin. Int. J. Environ. Res. Public Health 2017, 14, 496. [Google Scholar] [CrossRef]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.A. Physiographically-sensitive mapping of temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Daly, C.; Smith, J.I.; Olson, K.V. Mapping atmospheric moisture climatologies across the conterminous United States. PLoS ONE 2015, 10, e0141140. [Google Scholar] [CrossRef]
- PRISM Climate Group; Oregon State University. Available online: http://prism.oregonstate.edu (accessed on 1 July 2020).
- Philip, J.R. Plant water relations: Some physical aspects. Annu. Rev. Plant Physiol. 1966, 17, 245–268. [Google Scholar] [CrossRef]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap pressure in vascular plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Baughn, J.W.; Tanner, C.B. Leaf Water Potential: Comparison of Pressure Chamber and in situ Hygrometer on Five Herbaceous Species 1. Crop Sci. 1976, 16, 181–184. [Google Scholar] [CrossRef]
- Monteith, J.L. Evaporation and Environment. In Symposia of the Society for Experimental Biology 19; Cambridge University Press: Cambridge, UK, 1965; pp. 205–234. [Google Scholar]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1976, 273, 593–610. [Google Scholar]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. Symp. Soc. Exp. Biol. 1977, 31, 471–505. [Google Scholar]
- Ouimette, A.P.; Ollinger, S.V.; Richardson, A.D.; Hollinger, D.Y.; Keenan, T.F.; Lepine, L.C.; Vadeboncoeur, M.A. Carbon fluxes and interannual drivers in a temperate forest ecosystem assessed through comparison of top-down and bottom-up approaches. Agric. For. Meteorol. 2018, 256, 420–430. [Google Scholar] [CrossRef]
- Hollinger, D.Y.; Aber, J.; Dail, B.; Davidson, E.A.; Goltz, S.M.; Hughes, H.; Leclerc, M.Y.; Lee, J.T.; Richardson, A.D.; Rodrigues, C.; et al. Spatial and temporal variability in forest–atmosphere CO2 exchange. Glob. Chang. Biol. 2004, 10, 1689–1706. [Google Scholar] [CrossRef]
- Gu, L.; Pallardy, S.G.; Hosman, K.P.; Sun, Y. Drought-influenced mortality of tree species with different predawn leaf water dynamics in a decade-long study of a central US forest. Biogeosciences 2015, 12, 2831–2845. [Google Scholar] [CrossRef]
- Noormets, A.; Gavazzi, M.J.; McNulty, S.G.; Domec, J.C.; Sun, G.E.; King, J.S.; Chen, J. Response of carbon fluxes to drought in a coastal plain loblolly pine forest. Glob. Chang. Biol. 2010, 16, 272–287. [Google Scholar] [CrossRef]
- Clark, K.L.; Skowronski, N.; Gallagher, M.; Renninger, H.; Schäfer, K. Effects of invasive insects and fire on forest energy exchange and evapotranspiration in the New Jersey pinelands. Agric. For. Meteorol. 2012, 166, 50–61. [Google Scholar] [CrossRef]
- Clark, K.L.; Renninger, H.J.; Skowronski, N.; Gallagher, M.; Schäfer, K.V. Decadal-scale reduction in forest net ecosystem production following insect defoliation contrasts with short-term impacts of prescribed fires. Forests 2018, 9, 145. [Google Scholar] [CrossRef]
- Desai, A.R.; Bolstad, P.V.; Cook, B.D.; Davis, K.J.; Carey, E.V. Comparing net ecosystem exchange of carbon dioxide between an old-growth and mature forest in the upper Midwest, USA. Agric. For. Meteorol. 2005, 128, 33–55. [Google Scholar] [CrossRef]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Su, H.B.; Curtis, P.S. Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric. For. Meteorol. 2008, 148, 158–170. [Google Scholar] [CrossRef]
- Cook, B.D.; Davis, K.J.; Wang, W.; Desai, A.; Berger, B.W.; Teclaw, R.M.; Martin, J.G.; Bolstad, P.V.; Bakwin, P.S.; Yi, C.; et al. Carbon exchange and venting anomalies in an upland deciduous forest in northern Wisconsin, USA. Agric. For. Meteorol. 2004, 126, 271–295. [Google Scholar] [CrossRef]
- Elmore, A.J.; Nelson, D.; Guinn, S.M.; Paulman, R. Landsat-based Phenology and Tree Ring Characterization, Eastern US Forests, 1984–2013; ORNL DAAC: Oak Ridge, TN, USA, 2017. [CrossRef]
- Elmore, A.J.; Nelson, D.M.; Craine, J.M. Earlier springs are causing reduced nitrogen availability in North American eastern deciduous forests. Nat. Plants 2016. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Wardlow, B.D.; Bayissa, Y.A.; Tadesse, T.; Svoboda, M.D.; Gedam, S.S. Developing a Remote Sensing-Based Combined Drought Indicator Approach for Agricultural Drought Monitoring over Marathwada, India. Remote Sens. 2020, 12, 2091. [Google Scholar] [CrossRef]
- Bayissa, Y.A.; Tadesse, T.; Svoboda, M.; Wardlow, B.; Poulsen, C.; Swigart, J.; Van Andel, S.J. Developing a satellite-based combined drought indicator to monitor agricultural drought: A case study for Ethiopia. GIscience Remote Sens. 2019, 56, 718–748. [Google Scholar] [CrossRef]
- Hanson, P.J.; Weltzin, J.F. Drought disturbance from climate change: Response of United States forests. Sci. Total Environ. 2000, 262, 205–220. [Google Scholar] [CrossRef]
- Fritts, H. Tree Rings and Climate; Elsevier: Amsterdam, The Netherlands, 2012; p. 582. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Id | Name | Lat. | Long. | Elev. (m) | Veg. | Climate | MAT (°C) | MAP (mm) | Start | End | Site ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| US-Bar | Bartlett Experimental Forest | 44.065 | −71.288 | 272 | DBF | Dfb | 5.61 | 1246 | 2004 | 2017 | [60] |
| US-Ho1 | Howland Forest (main tower) | 45.204 | −68.740 | 60 | ENF | Dfb | 5.27 | 1070 | 1996 | 2018 | [61] |
| US-MMS | Morgan Monroe State Forest | 39.323 | −86.413 | 275 | DBF | Cfa | 10.85 | 1032 | 1999 | 2020 | [6] |
| US-MOz | Missouri Ozark Site | 38.744 | −92.2 | 219 | DBF | Cfa | 12.11 | 986 | 2004 | 2017 | [62] |
| US-NC2 | NC Loblolly Plantation | 35.803 | −76.669 | 5 | ENF | Cfa | 16.6 | 1320 | 2005 | 2019 | [63] |
| US-Slt | Silas Little Forest | 39.914 | −74.596 | 30 | DBF | Dfa | 11.04 | 1138 | 2005 | 2017 | [64,65] |
| US-Syv | Sylvania Wilderness Area | 46.242 | −89.348 | 540 | MF | Dfb | 3.81 | 826 | 2001 | 2020 | [66] |
| US-UMB | Univ. of Mich. Biological Station | 45.560 | −84.714 | 234 | DBF | Dfb | 5.83 | 803 | 2000 | 2019 | [67] |
| US-WCr | Willow Creek | 45.806 | −90.080 | 520 | DBF | Dfb | 4.02 | 787 | 1998 | 2020 | [68] |
| Site | County | State | Year | Dates | Intensity |
|---|---|---|---|---|---|
| MMS | Monroe | Indiana | 2012 | 26 June–4 Sept | D2 |
| 17 July–28 Aug | D3 | ||||
| 24 July–7 Aug | D4 | ||||
| 2010 | 21 Sept–23 November | D2 | |||
| 2007 | 21 Aug–26 Oct | D2 | |||
| MOz | Boone | Missouri | |||
| 2012 | 3 July–end of year | D2 | |||
| 17 July–16 Oct | D3 | ||||
| 14 Aug–28 Aug | D4 | ||||
| 2006 | 8 Aug–22 Aug | D2 | |||
| 2007 | 21 Aug–16 Oct | D1 | |||
| NC2 | Washington | North Carolina | 2011 | 31 May–23 Aug | D2 |
| 20 Nov–4 Mar 2012 | D2 | ||||
| 2008 | 1 Jan–26 Aug | D2 | |||
| 29 Jan–12 Feb, 26 Aug (one week) | D3 | ||||
| 2007 | 4 Sept–23 Oct | D2 | |||
| Slt | Burlington | New Jersey | 2010 | 7 Sept–28 Sept | D2 |
| 2007 | June, Gypsy moth outbreak | none | |||
| UMB | Cheboygan | Michigan | 2011 | 29 Mar–26 Apr | D1 |
| 2010 | 6 April–17 Aug | D1 | |||
| 2007 | 28 Aug–4 Sept | D2 | |||
| 2005 | 19 July–16 Aug | D2 | |||
| 2003 | 7 Jan–1 April, 23 Sept | D1 | |||
| Syv | Gogebic | Michigan | * 2010 | 1-29 June | D3 |
| 13 April–17 Aug | D2 | ||||
| * 2009 | 22 Sept–20 Oct | D2 | |||
| * 2008 | 26 Aug–12 May 2009 | D1 | |||
| * 2007 | 14 Aug–4 Sep | D3 | |||
| 10 July–16 Oct | D2 | ||||
| 2006 | 11 July–25 July | D2 | |||
| WCr | Price | Wisconsin | 2012 | 9–23 Oct | D2 |
| * 2010 | 13 April–22 June | D2 | |||
| * 2009 | 4–18 Aug | D3 | |||
| 25 Jan–Aug | D2 | ||||
| * 2008 | 21 Oct–end of year | D2 | |||
| * 2007 | 12–18 Sept | D2 | |||
| 2005 | 6 Sept–4 Oct | D2 | |||
| 2003 | 18–25 Mar, 22–29 July, 2 Sept–end of year | D1 | |||
| Ho1 | Penobscot | Maine | 2016 | 15 Nov–20 Dec | D2 |
| 2016/17 | 27 Sept–7 Feb 2017 | D1 | |||
| 2010 | 10 Aug–28 Sept | D1 | |||
| Bar | Carrol | New Hampshire | 2016/17 | 27 Sept–7 Feb 2017 | D1 |
| Site/Tree ID | Min | Max | Average |
|---|---|---|---|
| HAFE_49_1 | 0.22 | 0.59 | 0.46 |
| HAFE_59_2 | 0.33 | 0.63 | 0.48 |
| CATO_25_1_2 | 0.52 | 0.73 | 0.64 |
| CATO_42_1_2 | 0.35 | 0.67 | 0.54 |
| GRSM_101_2_1 | 0.68 | 0.81 | 0.75 |
| GRSM_66_1_1 | 0.74 | 0.82 | 0.78 |
| PRWI_15_2_1 | 0.61 | 0.81 | 0.73 |
| PRWI_3_1_2 | 0.61 | 0.81 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadesse, T.; Hollinger, D.Y.; Bayissa, Y.A.; Svoboda, M.; Fuchs, B.; Zhang, B.; Demissie, G.; Wardlow, B.D.; Bohrer, G.; Clark, K.L.; et al. Forest Drought Response Index (ForDRI): A New Combined Model to Monitor Forest Drought in the Eastern United States. Remote Sens. 2020, 12, 3605. https://doi.org/10.3390/rs12213605
Tadesse T, Hollinger DY, Bayissa YA, Svoboda M, Fuchs B, Zhang B, Demissie G, Wardlow BD, Bohrer G, Clark KL, et al. Forest Drought Response Index (ForDRI): A New Combined Model to Monitor Forest Drought in the Eastern United States. Remote Sensing. 2020; 12(21):3605. https://doi.org/10.3390/rs12213605
Chicago/Turabian StyleTadesse, Tsegaye, David Y. Hollinger, Yared A. Bayissa, Mark Svoboda, Brian Fuchs, Beichen Zhang, Getachew Demissie, Brian D. Wardlow, Gil Bohrer, Kenneth L. Clark, and et al. 2020. "Forest Drought Response Index (ForDRI): A New Combined Model to Monitor Forest Drought in the Eastern United States" Remote Sensing 12, no. 21: 3605. https://doi.org/10.3390/rs12213605
APA StyleTadesse, T., Hollinger, D. Y., Bayissa, Y. A., Svoboda, M., Fuchs, B., Zhang, B., Demissie, G., Wardlow, B. D., Bohrer, G., Clark, K. L., Desai, A. R., Gu, L., Noormets, A., Novick, K. A., & Richardson, A. D. (2020). Forest Drought Response Index (ForDRI): A New Combined Model to Monitor Forest Drought in the Eastern United States. Remote Sensing, 12(21), 3605. https://doi.org/10.3390/rs12213605