Asymmetry of Daytime and Nighttime Warming in Typical Climatic Zones along the Eastern Coast of China and Its Influence on Vegetation Activities



Abstract
1. Introduction
2. Materials and Methods
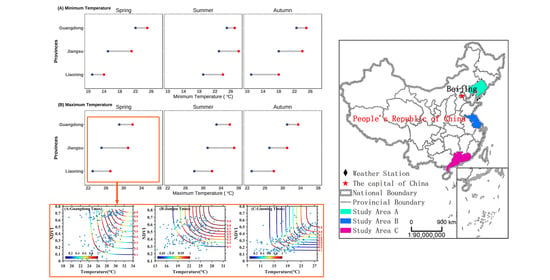
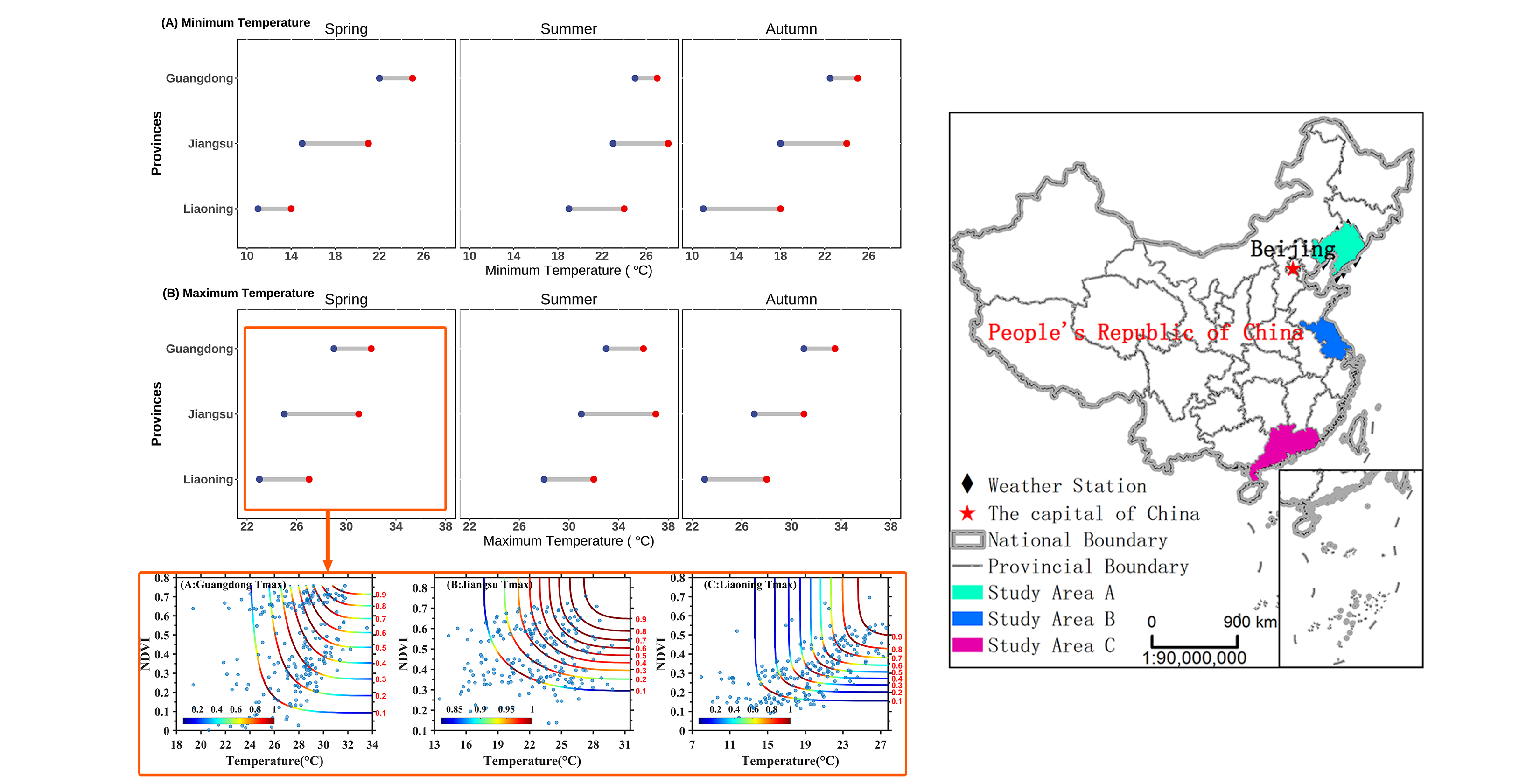
2.1. Research Areas
2.2. Data and Preprocessing
2.2.1. Meteorological Data and Preprocessing
2.2.2. NDVI Data and Preprocessing
2.3. Methods
2.3.1. Maximum Value Composites
2.3.2. Copula Function Theory
Parameter Estimation
Verification and Evaluation
Correlation Analysis and Establishment of Marginal Distribution Function
Joint Probability Distribution
The Return Period of NDVI and Tmax/Tmin
3. Results
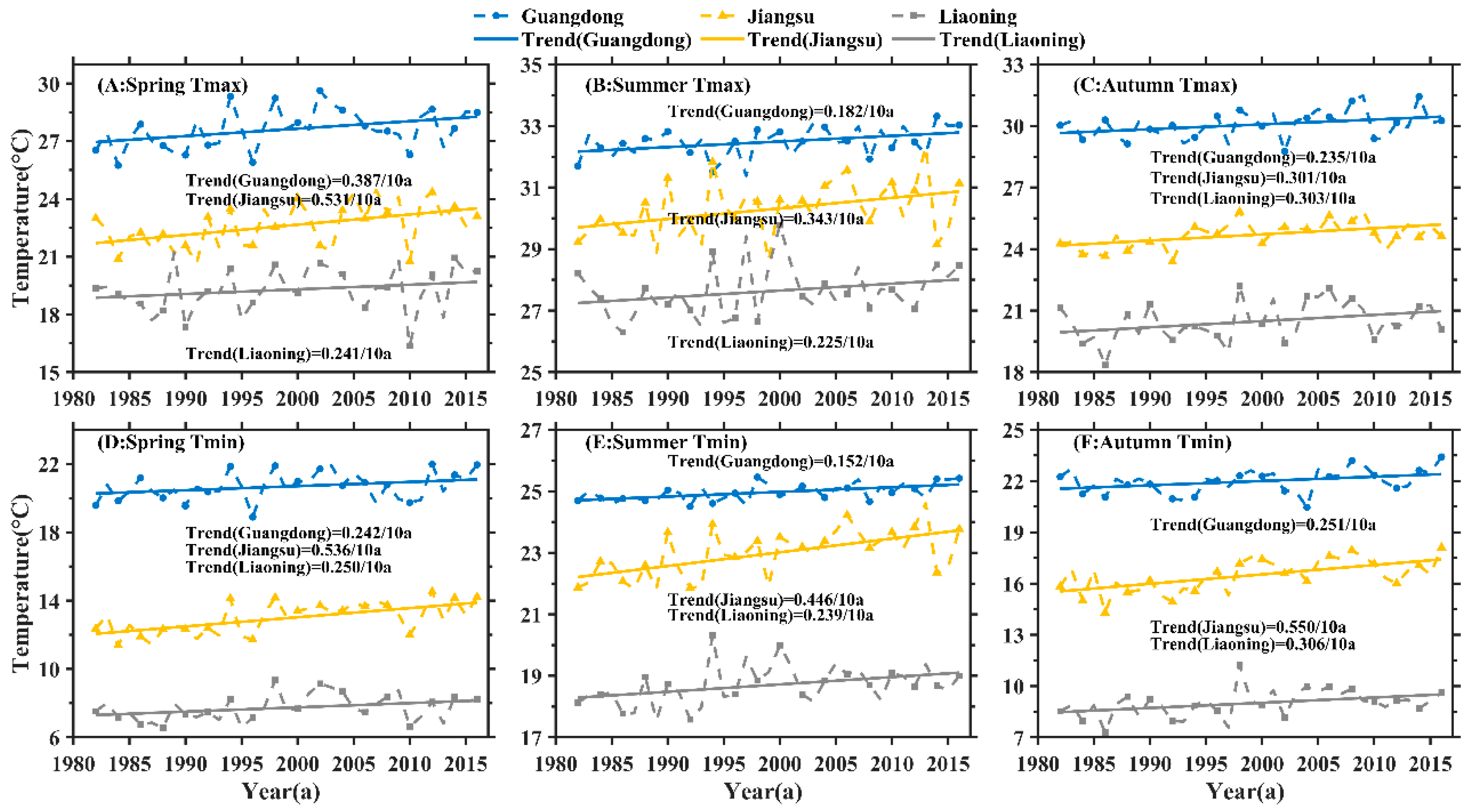
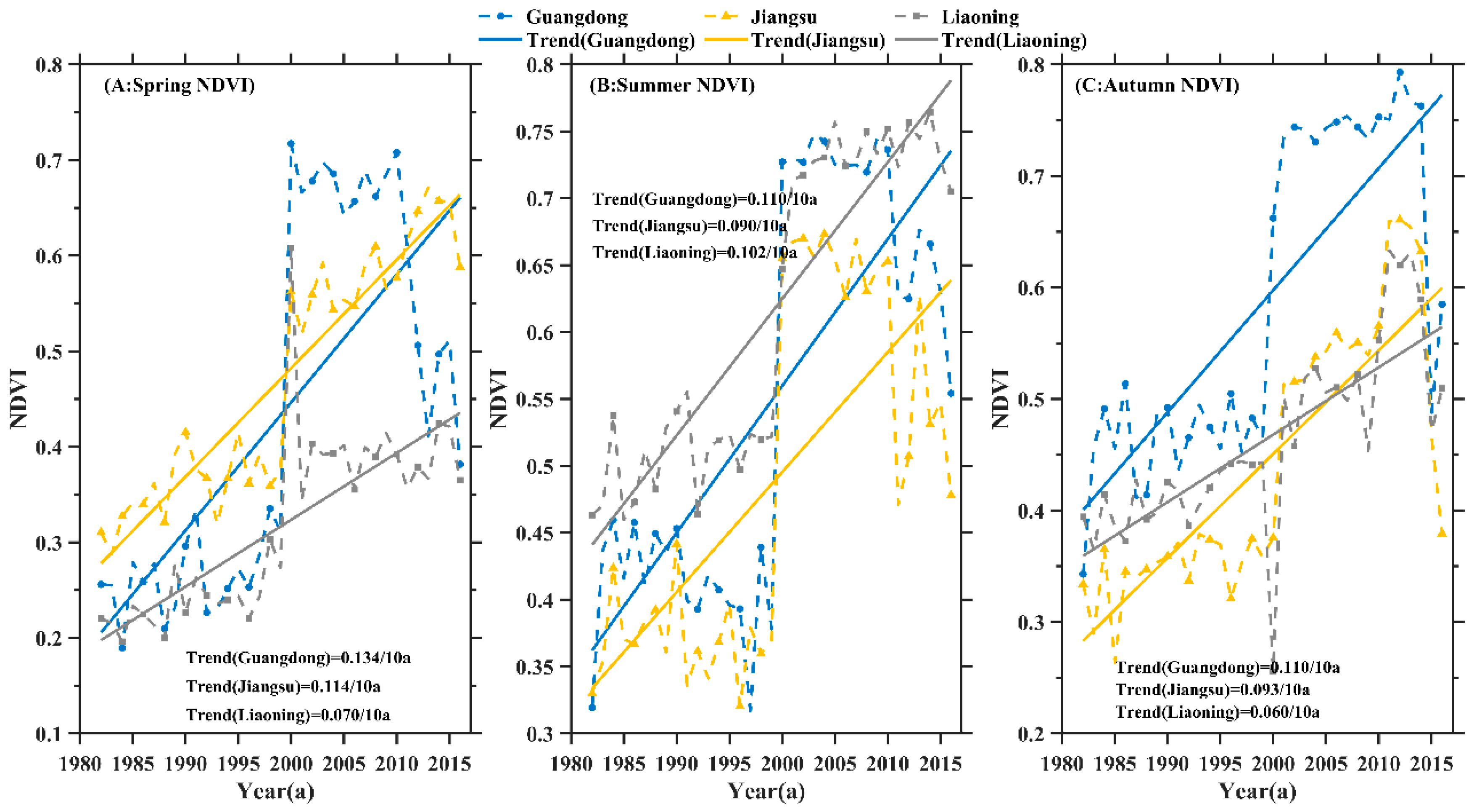
3.1. Trend Analysis of Seasonal Daytime and Nighttime Temperature Increases and NDVI
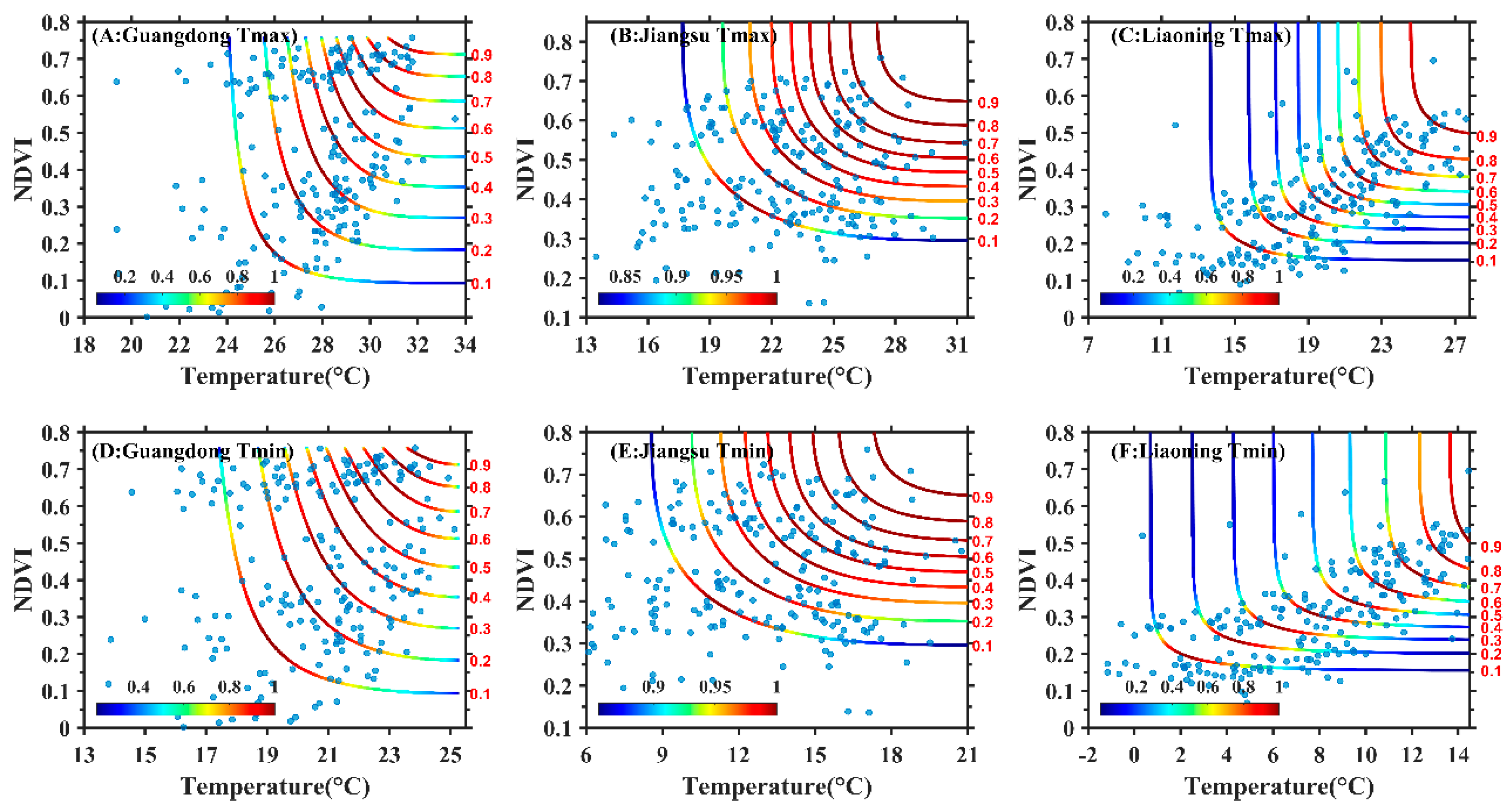
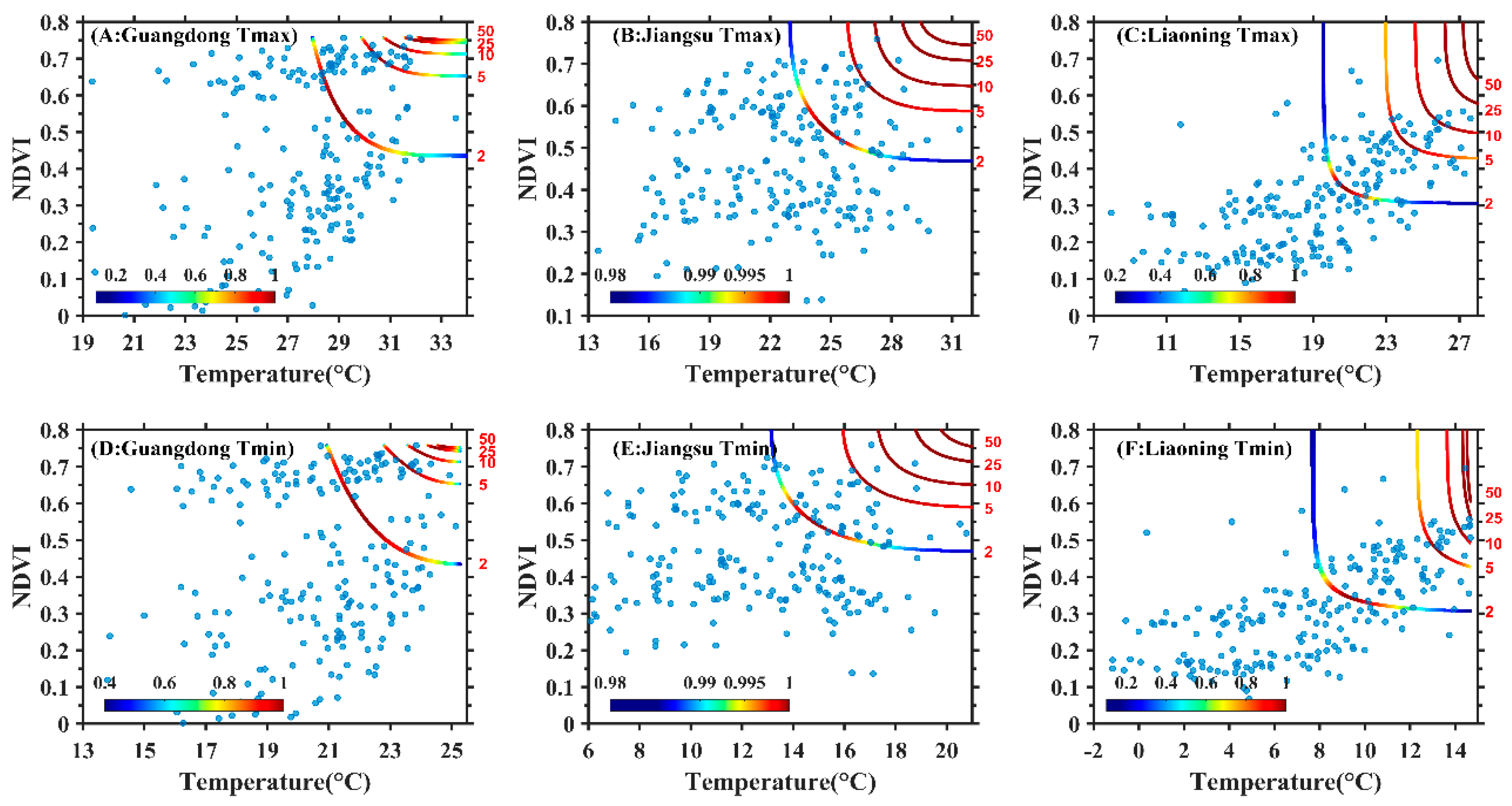
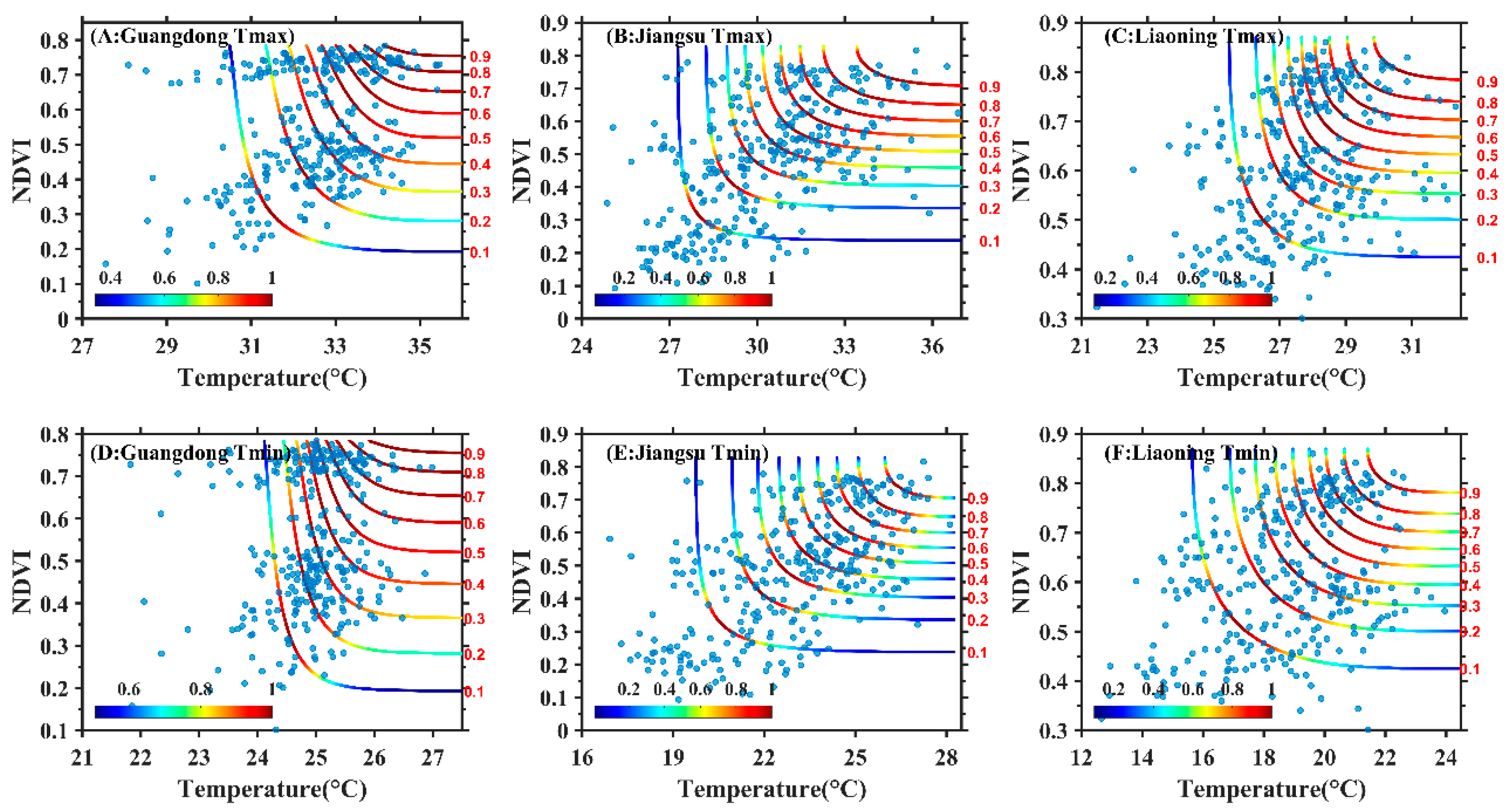
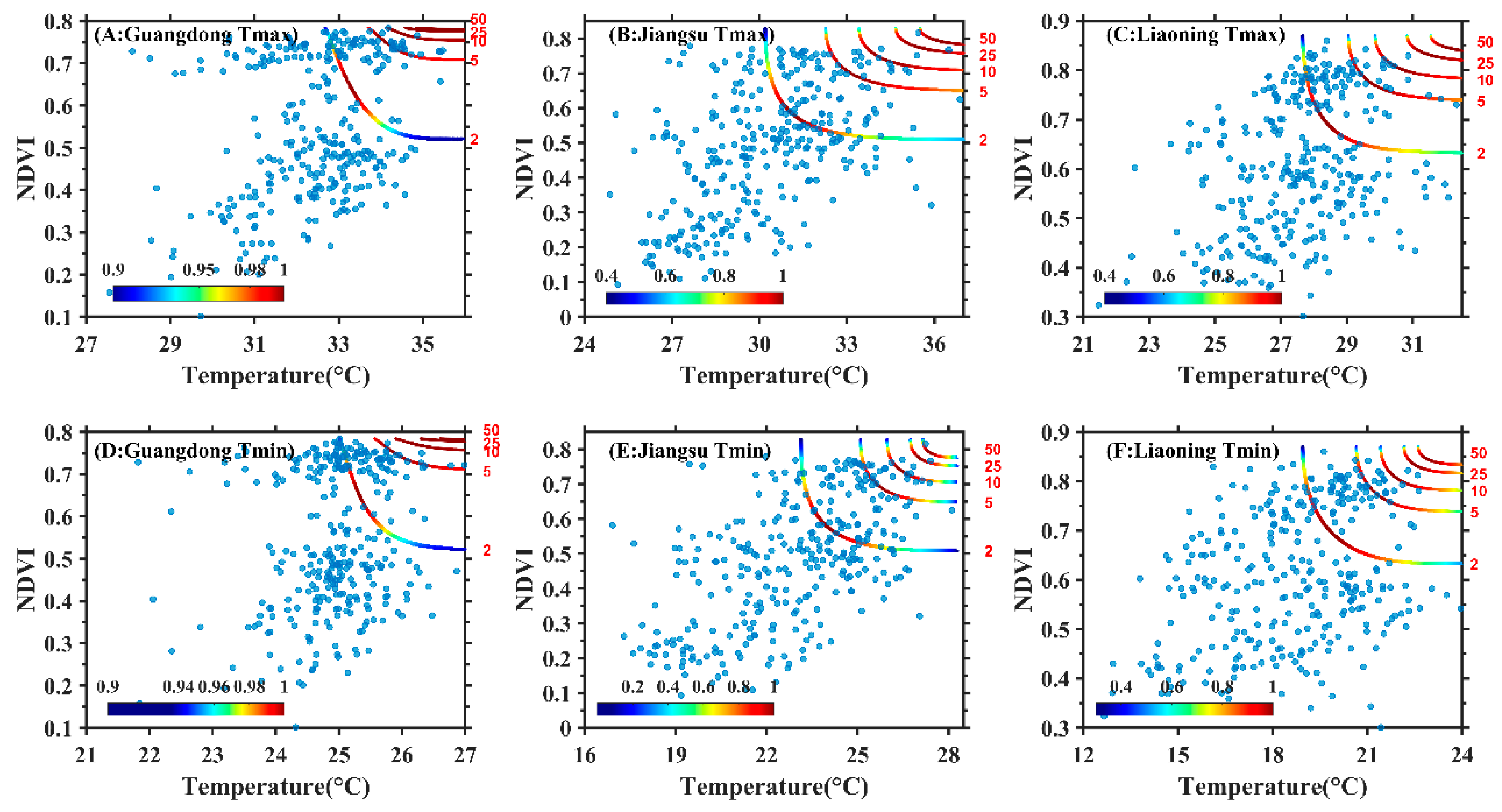
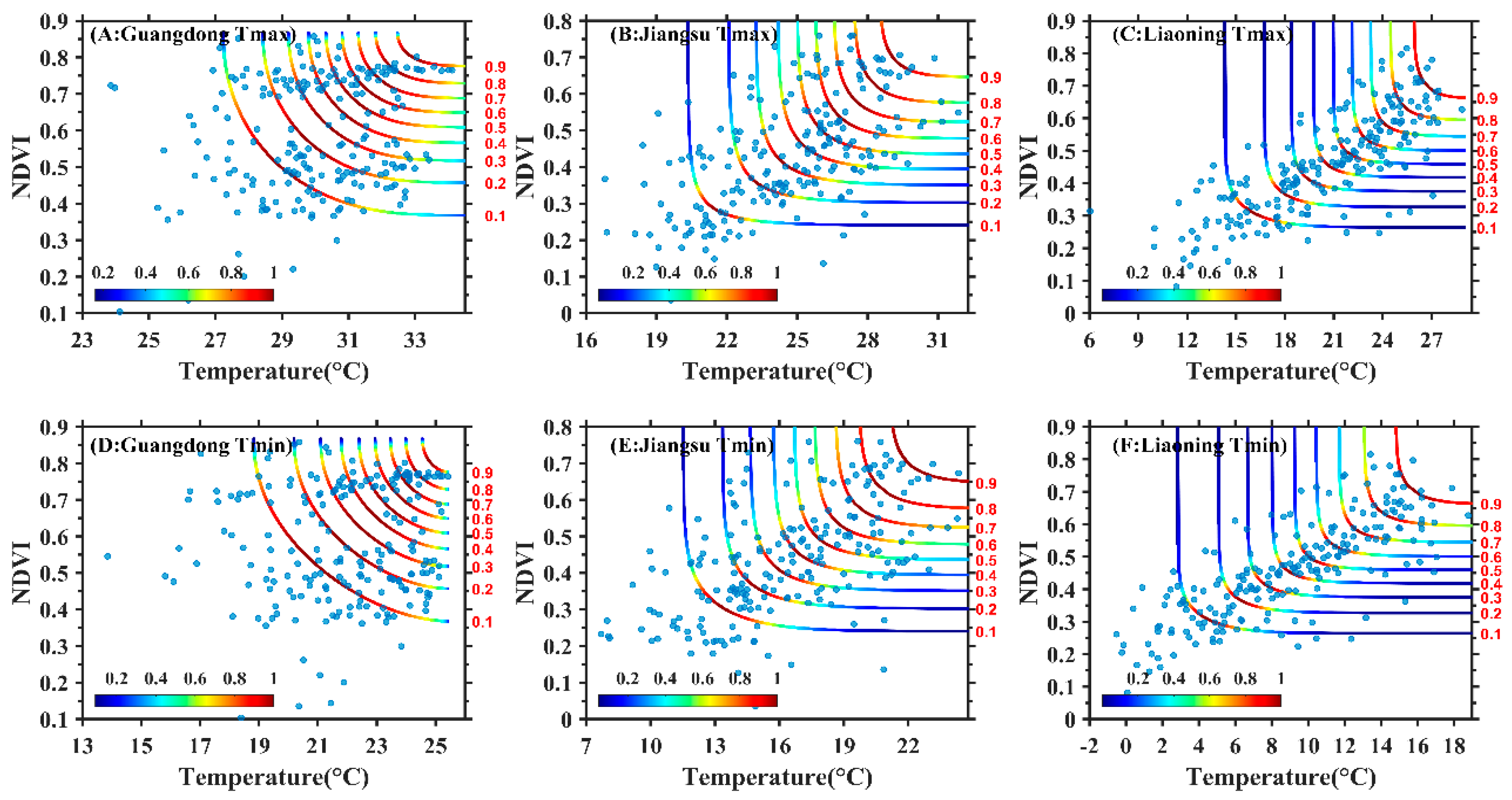
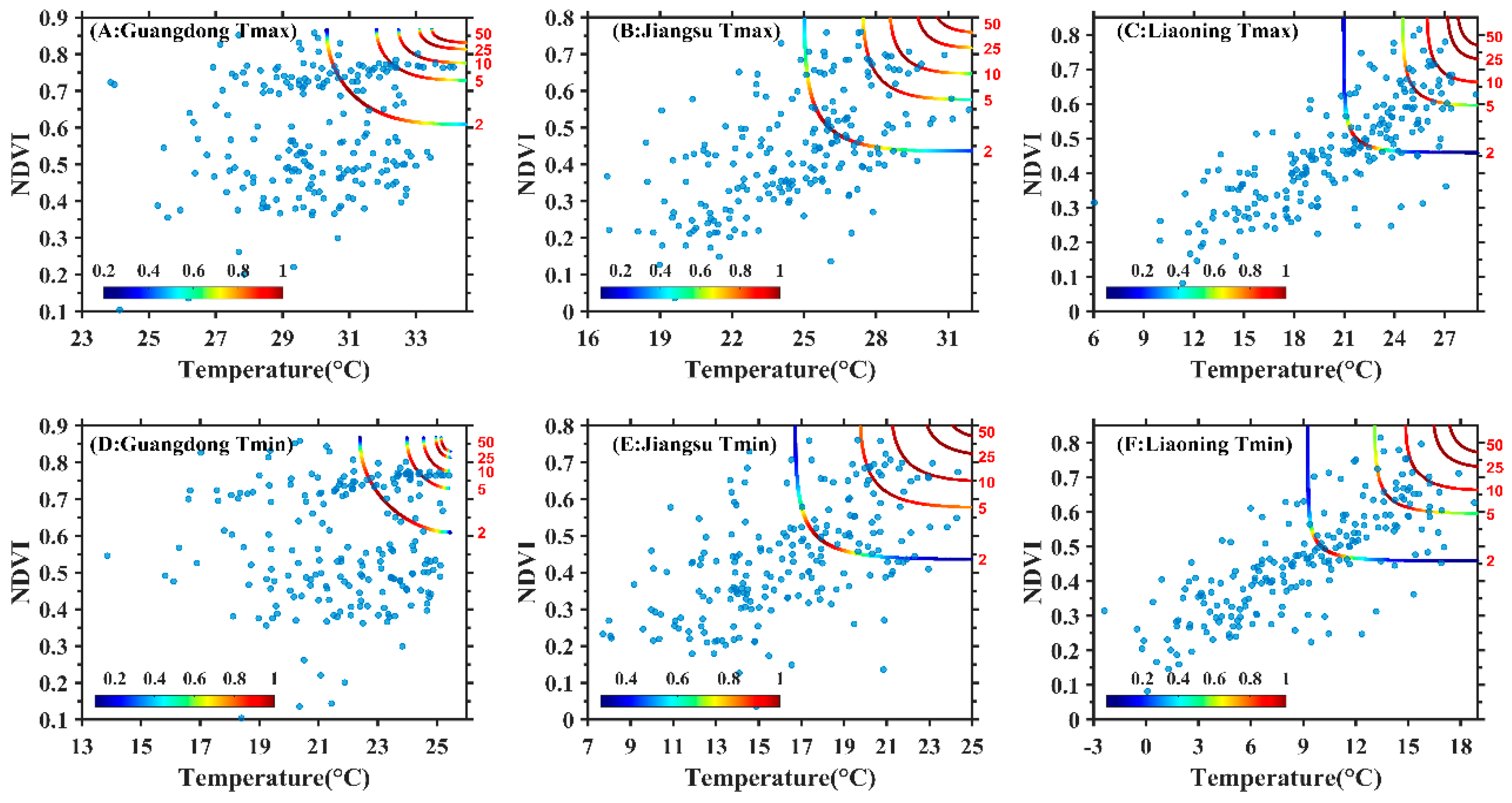
3.2. Construction of Copula Function Cluster of the Maximum Temperature, Minimum Temperature and NDVI
3.3. Joint Probability Distribution Characteristics of Maximum Temperature, Minimum Temperature and NDVI
4. Discussion
4.1. Effects of the Asymmetry of Daytime and Nighttime Temperature Increases on Vegetation Activities
4.2. Effects of Non-Uniform Temperature Increase in Different Seasons on Vegetation Activities
4.3. Exploration of Dynamic Changes of Temperature and NDVI Using Copula Function
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gong, Z.; Zhao, S.; Gu, J. Correlation analysis between vegetation coverage and climate drought conditions in North China during 2001–2013. J. Geogr. Sci. 2017, 27, 143–160. [Google Scholar] [CrossRef]
- Zhang, W.; Döscher, R.; Koenigk, T.; Miller, P.A.; Jansson, C.; Samuelsson, P.; Wu, M.; Smith, B. The Interplay of Recent Vegetation and Sea Ice Dynamics—Results from a Regional Earth System Model Over the Arctic. Geophys. Res. Lett. 2020, 47, e2019GL085982. [Google Scholar] [CrossRef]
- Teixeira, A.H.D.C.; Bastiaanssen, W.G.M.; Ahmad, M.D.; Moura, M.S.B.; Bos, M.G. Analysis of energy fluxes and vegetation-atmosphere parameters in irrigated and natural ecosystems of semi-arid Brazil. J. Hydrol. 2008, 362, 110–127. [Google Scholar] [CrossRef]
- Balzarolo, M.; Vicca, S.; Nguy-Robertson, A.L.; Bonal, D.; Elbers, J.A.; Fu, Y.H.; Grünwald, T.; Horemans, J.A.; Papale, D.; Peñuelas, J.; et al. Matching the phenology of Net Ecosystem Exchange and vegetation indices estimated with MODIS and FLUXNET in-situ observations. Remote Sens. Environ. 2016, 174, 290–300. [Google Scholar] [CrossRef]
- Dong, L.; Tang, S.; Min, M.; Veroustraete, F. Estimation of Forest Canopy Height in Hilly Areas Using Lidar Waveform Data. IEEE J. Sel. Top. in Appl. Earth Obs. Remote Sens. 2019, 12, 1559–1571. [Google Scholar] [CrossRef]
- Verrelst, J.; Camps-Valls, G.; Muñoz-Marí, J.; Rivera, J.P.; Veroustraete, F.; Clevers, J.G.P.W.; Moreno, J. Optical remote sensing and the retrieval of terrestrial vegetation bio-geophysical properties–A review. ISPRS J. Photogramm. Remote Sens. 2015, 108, 273–290. [Google Scholar] [CrossRef]
- Baldocchi, D.; Finnigan, J.; Wilson, K.; Paw U, K.T.; Falge, E. On Measuring Net Ecosystem Carbon Exchange Over Tall Vegetation on Complex Terrain. Bound. Layer Meteorol. 2000, 96, 257–291. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Mac Arthur, A.; Rossini, M.; Eklundh, L.; Pacheco-Labrador, J.; Anderson, K.; Balzarolo, M.; Martín, M.; Jin, H.; Tomelleri, E.; et al. EUROSPEC: At the interface between remote sensing and ecosystem CO2 flux measurements in Europe. Biogeosciences Discuss. 2015, 12, 13069–13121. [Google Scholar] [CrossRef]
- Balzarolo, M.; Vescovo, L.; Hammerle, A.; Gianelle, D.; Papale, D.; Wohlfahrt, G. On the relationship between ecosystem-scale hyperspectral reflectance and CO2 exchange in European mountain grasslands. Biogeosciences Discuss. 2014, 11, 10323–10363. [Google Scholar] [CrossRef]
- Couwenberg, J.; Thiele, A.; Tanneberger, F.; Augustin, J.; Bärisch, S.; Dubovik, D.; Liashchynskaya, N.; Michaelis, D.; Minke, M.; Skuratovich, A.; et al. Assessing greenhouse gas emissions from peatlands using vegetation as a proxy. Hydrobiologia 2011, 674, 67–89. [Google Scholar] [CrossRef]
- Führer, E. Forest functions, ecosystem stability and management. For. Ecol. Manag. 2000, 132, 29–38. [Google Scholar] [CrossRef]
- Zinnert, J.C.; Shiflett, S.A.; Vick, J.K.; Young, D.R. Woody vegetative cover dynamics in response to recent climate change on an Atlantic coast barrier island: A remote sensing approach. Geocarto Int. 2011, 26, 595–612. [Google Scholar] [CrossRef]
- Sailor, D. Simulated Urban Climate Response to Modifications in Surface Albedo and Vegetative Cover. J. Appl. Meteorol. 1995, 34, 1694–1704. [Google Scholar] [CrossRef]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Peñuelas, J.; Zhang, G.; et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Balzarolo, M.; Penuelas, J.; Veroustraete, F. Influence of Landscape Heterogeneity and Spatial Resolution in Multi-Temporal In Situ and MODIS NDVI Data Proxies for Seasonal GPP Dynamics. Remote Sens. 2019, 11, 1656. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.A.O.; Liu, J.I.E. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Zhao, L.; Dai, A.; Dong, B. Changes in global vegetation activity and its driving factors during 1982–2013. Agric. For. Meteorol. 2018, 249, 198–209. [Google Scholar] [CrossRef]
- Chu, H.; Venevsky, S.; Wu, C.; Wang, M. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P.; Swann, A.S.; Cook, B.I.; Scheff, J. Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2. Proc. Natl. Acad. Sci. USA 2018, 115, 4093. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33, L23402. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Q.; Singh, V.P.; Shi, P. Seasonal vegetation response to climate change in the Northern Hemisphere (1982–2013). Glob. Planet. Chang. 2017, 148, 1–8. [Google Scholar] [CrossRef]
- Wu, X.; Liu, H.; Li, X.; Liang, E.; Beck, P.S.A.; Huang, Y. Seasonal divergence in the interannual responses of Northern Hemisphere vegetation activity to variations in diurnal climate. Sci. Rep. 2016, 6, 19000. [Google Scholar] [CrossRef] [PubMed]
- Park, T.; Chen, C.; Macias-Fauria, M.; Tømmervik, H.; Choi, S.; Winkler, A.; Bhatt, U.S.; Walker, D.A.; Piao, S.; Brovkin, V.; et al. Changes in timing of seasonal peak photosynthetic activity in northern ecosystems. Glob. Chang. Biol. 2019, 25, 2382–2395. [Google Scholar] [CrossRef]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Rich, R.L.; Hobbie, S.E.; Montgomery, R.A. Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture. Nature 2018, 562, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5, 5018. [Google Scholar] [CrossRef]
- Chen, B.-M.; Gao, Y.; Liao, H.-X.; Peng, S.-L. Differential responses of invasive and native plants to warming with simulated changes in diurnal temperature ranges. AoB Plants 2017, 9. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef]
- Arora, V.; Saran, R.K.; Kumar, R.; Yadav, S. Separation and sequestration of CO2 in geological formations. Mater. Sci. Energy Technol. 2019, 2, 647–656. [Google Scholar] [CrossRef]
- Xia, J.; Chen, J.; Piao, S.; Ciais, P.; Luo, Y.; Wan, S. Terrestrial carbon cycle affected by non-uniform climate warming. Nat. Geosci. 2014, 7, 173–180. [Google Scholar] [CrossRef]
- Karl, T.R.; Jones, P.; Knight, R.; Kukla, G.; Plummer, N.; Razuvayev, V.; Gallo, K.; Lindseay, J.; Charlson, R.; Peterson, T. A New Perspective on Recent Global Warming: Asymmetric Trends of Daily Maximum and Minimum Temperature. Bull. Am. Meteorol. Soc. 1993, 74, 1007–1024. [Google Scholar] [CrossRef]
- Easterling, D.R.; Horton, B.; Jones, P.D.; Peterson, T.C.; Karl, T.R.; Parker, D.E.; Salinger, M.J.; Razuvayev, V.; Plummer, N.; Jamason, P.; et al. Maximum and Minimum Temperature Trends for the Globe. Science 1997, 277, 364. [Google Scholar] [CrossRef]
- Ventura, F.; Rossi Pisa, P.; Ardizzoni, E. Temperature and precipitation trends in Bologna (Italy) from 1952 to 1999. Atmos. Res. 2002, 61, 203–214. [Google Scholar] [CrossRef]
- Price, C.; Michaelides, S.; Pashiardis, S.; Alpert, P. Long term changes in diurnal temperature range in Cyprus. Atmos. Res. 1999, 51, 85–98. [Google Scholar] [CrossRef]
- Davy, R.; Esau, I.; Chernokulsky, A.; Outten, S.; Zilitinkevich, S. Diurnal asymmetry to the observed global warming. Int. J. Climatol. 2017, 37, 79–93. [Google Scholar] [CrossRef]
- Su, H.; Feng, J.; Axmacher, J.C.; Sang, W. Asymmetric warming significantly affects net primary production, but not ecosystem carbon balances of forest and grassland ecosystems in northern China. Sci. Rep. 2015, 5, 9115. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.; van der Velde, M.; Lin, E.; Xiong, W.; Li, Y. The impacts of climate change on agricultural production systems in China. Clim. Chang. 2013, 120, 313–324. [Google Scholar] [CrossRef]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.H.; Ciais, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z.; Jeong, S.-J.; et al. Leaf onset in the northern hemisphere triggered by daytime temperature. Nat. Commun. 2015, 6, 6911. [Google Scholar] [CrossRef]
- Atkin, O.K.; Turnbull, M.H.; Zaragoza-Castells, J.; Fyllas, N.M.; Lloyd, J.; Meir, P.; Griffin, K.L. Light inhibition of leaf respiration as soil fertility declines along a post-glacial chronosequence in New Zealand: An analysis using the Kok method. Plant Soil 2013, 367, 163–182. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of Earth Observation based global long term vegetation trends—Comparing GIMMS and MODIS global NDVI time series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Jiapaer, G.; Liang, S.; Yi, Q.; Liu, J. Vegetation dynamics and responses to recent climate change in Xinjiang using leaf area index as an indicator. Ecol. Indic. 2015, 58, 64–76. [Google Scholar] [CrossRef]
- He, B.; Chen, A.; Jiang, W.; Chen, Z. The response of vegetation growth to shifts in trend of temperature in China. J. Geogr. Sci. 2017, 27, 801–816. [Google Scholar] [CrossRef]
- Zhang, B.; Cui, L.; Shi, J.; Wei, P. Vegetation Dynamics and Their Response to Climatic Variability in China. Adv. Meteorol. 2017, 2017, 8282353. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Yang, W.; Yang, Z.; Zhang, G.; Piao, S. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int. J. Biometeorol. 2017, 61, 1433–1444. [Google Scholar] [CrossRef]
- Xia, H.; Li, A.; Feng, G.; Li, Y.; Qin, Y.; Lei, G.; Cui, Y. The Effects of Asymmetric Diurnal Warming on Vegetation Growth of the Tibetan Plateau over the Past Three Decades. Sustainability 2018, 10, 1103. [Google Scholar] [CrossRef]
- Tan, J.; Piao, S.; Chen, A.; Zeng, Z.; Ciais, P.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Peng, S.; Peñuelas, J.; et al. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Glob. Chang. Biol. 2015, 21, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, X.; Jiang, M.; Lu, X. Vegetation Change and Its Response to Climate Change between 2000 and 2016 in Marshes of the Songnen Plain, Northeast China. Sustainability 2020, 12, 3569. [Google Scholar] [CrossRef]
- Rossi, S.; Isabel, N. Bud break responds more strongly to daytime than night-time temperature under asymmetric experimental warming. Glob. Chang. Biol. 2017, 23, 446–454. [Google Scholar] [CrossRef]
- Miller, J.A. Species distribution models: Spatial autocorrelation and non-stationarity. Prog. Phys. Geogr. Earth Environ. 2012, 36, 681–692. [Google Scholar] [CrossRef]
- Xu, L.; Chen, X. Spatial modeling of the Ulmus pumila growing season in China’s temperate zone. Sci. China Earth Sci. 2012, 55, 656–664. [Google Scholar] [CrossRef]
- Dai, J.; Wang, H.; Ge, Q. The spatial pattern of leaf phenology and its response to climate change in China. Int. J. Biometeorol. 2014, 58, 521–528. [Google Scholar] [CrossRef]
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens. 1986, 7, 1417–1434. [Google Scholar] [CrossRef]
- Kundu, A.; Denis, D.M.; Patel, N.R.; Dutta, D. Climate Change, Extreme Events and Disaster Risk Reduction: Towards Sustainable Development Goals; Mal, S., Singh, R.B., Huggel, C., Eds.; Springer International Publishing: Cham, Germany, 2018; pp. 89–99. [Google Scholar]
- Li, D.; Gui, Y.; Li, Y.; Xiong, L. A method for constructing asymmetric pair-copula and its application. Commun. Stat. Theory Methods 2018, 47, 4202–4214. [Google Scholar] [CrossRef]
- Salvadori, G.; De Michele, C. On the Use of Copulas in Hydrology: Theory and Practice. J. Hydrol. Eng. 2007, 12, 369–380. [Google Scholar] [CrossRef]
- Clayton, D.G. A Model for Association in Bivariate Life Tables and Its Application in Epidemiological Studies of Familial Tendency in Chronic Disease Incidence. Biometrika 1978, 65, 141–151. [Google Scholar] [CrossRef]
- Genest, C.; Favre, A.-C. Everything You Always Wanted to Know about Copula Modeling but Were Afraid to Ask. J. Hydrol. Eng. 2007, 12, 347–368. [Google Scholar] [CrossRef]
- Li, C.; Singh, V.P.; Mishra, A.K. A bivariate mixed distribution with a heavy-tailed component and its application to single-site daily rainfall simulation. Water Resour. Res. 2013, 49, 767–789. [Google Scholar] [CrossRef]
- Sraj, M.; Bezak, N.; Brilly, M. Bivariate flood frequency analysis using the copula function: A case study of the Litija station on the Sava River. Hydrol. Process. 2015, 29, 225–238. [Google Scholar] [CrossRef]
- Genest, C.; Rivest, L.-P. Statistical Inference Procedures for Bivariate Archimedean Copulas. J. Am. Stat. Assoc. 1993, 88, 1034–1043. [Google Scholar] [CrossRef]
- Ben Nasr, I.; Chebana, F. Multivariate L-moment based tests for copula selection, with hydrometeorological applications. J. Hydrol. 2019, 579, 124151. [Google Scholar] [CrossRef]
- Gródek-Szostak, Z.; Malik, G.; Kajrunajtys, D.; Szeląg-Sikora, A.; Sikora, J.; Kuboń, M.; Niemiec, M.; Kapusta-Duch, J. Modeling the Dependency between Extreme Prices of Selected Agricultural Products on the Derivatives Market Using the Linkage Function. Sustainability 2019, 11, 4144. [Google Scholar] [CrossRef]
- Pho, K.-H.; Ly, S.; Ly, S.; Lukusa, T.M. Comparison among Akaike Information Criterion, Bayesian Information Criterion and Vuongs test in Model Selection: A Case Study of Violated Speed Regulation in Taiwan. J. Adv. Eng. Comput. 2019, 3, 293–303. [Google Scholar] [CrossRef]
- Dodangeh, E.; Shahedi, K.; Shiau, J.-T.; MirAkbari, M. Spatial hydrological drought characteristics in Karkheh River basin, southwest Iran using copulas. J. Earth Syst. Sci. 2017, 126, 80. [Google Scholar] [CrossRef]
- Sadegh, M.; Ragno, E.; AghaKouchak, A. Multivariate Copula Analysis Toolbox (MvCAT): Describing dependence and underlying uncertainty using a Bayesian framework. Water Resour. Res. 2017, 53, 5166–5183. [Google Scholar] [CrossRef]
- Sadegh, M.; Moftakhari, H.; Gupta, H.V.; Ragno, E.; Mazdiyasni, O.; Sanders, B.; Matthew, R.; AghaKouchak, A. Multihazard Scenarios for Analysis of Compound Extreme Events. Geophys. Res. Lett. 2018, 45, 5470–5480. [Google Scholar] [CrossRef]
- Chen, H.; Sun, J. Changes in Drought Characteristics over China Using the Standardized Precipitation Evapotranspiration Index. J. Clim. 2015, 28, 5430–5447. [Google Scholar] [CrossRef]
- Singh, V.P.; Zhang, L. Copula–entropy theory for multivariate stochastic modeling in water engineering. Geosci. Lett. 2018, 5, 6. [Google Scholar] [CrossRef]
- Shiau, J.T. Fitting Drought Duration and Severity with Two-Dimensional Copulas. Water Resour. Manag. 2006, 20, 795–815. [Google Scholar] [CrossRef]
- Ma, L.; Qin, F.; Wang, H.; Qin, Y.; Xia, H. Asymmetric seasonal daytime and nighttime warming and its effects on vegetation in the Loess Plateau. PLoS ONE 2019, 14, e0218480. [Google Scholar] [CrossRef]
- Xu, L.; Myneni, R.B.; Chapin Iii, F.S.; Callaghan, T.V.; Pinzon, J.E.; Tucker, C.J.; Zhu, Z.; Bi, J.; Ciais, P.; Tømmervik, H.; et al. Temperature and vegetation seasonality diminishment over northern lands. Nat. Clim. Chang. 2013, 3, 581–586. [Google Scholar] [CrossRef]
- Yang, Z.; Shen, M.; Jia, S.; Guo, L.; Yang, W.; Wang, C.; Chen, X.; Chen, J. Asymmetric Responses of the End of Growing Season to Daily Maximum and Minimum Temperatures on the Tibetan Plateau. J. Geophys. Res. Atmos. 2017, 122, 13–278. [Google Scholar] [CrossRef]
- Jing, P.; Wang, D.; Zhu, C.; Chen, J. Plant Physiological, Morphological and Yield-Related Responses to Night Temperature Changes across Different Species and Plant Functional Types. Front. Plant Sci. 2016, 7, 1774. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Y.; Shafeeque, M. Interpretation of vegetation phenology changes using daytime and night-time temperatures across the Yellow River Basin, China. Sci. Total Environ. 2019, 693, 133553. [Google Scholar] [CrossRef]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971. [Google Scholar] [CrossRef]
- Alward, R.D.; Detling, J.K.; Milchunas, D.G. Grassland Vegetation Changes and Nocturnal Global Warming. Science 1999, 283, 229. [Google Scholar] [CrossRef] [PubMed]
- Morita, S.; Yonemaru, J.-I.; Takanashi, J.-I. Grain Growth and Endosperm Cell Size under High Night Temperatures in Rice (Oryza sativa L.). Ann. Bot. 2005, 95, 695–701. [Google Scholar] [CrossRef]
- Todisco, F.; Vergni, L. Climatic changes in Central Italy and their potential effects on corn water consumption. Agric. For. Meteorol. 2008, 148, 1–11. [Google Scholar] [CrossRef]
- Nicholls, N. Increased Australian wheat yield due to recent climate trends. Nature 1997, 387, 484–485. [Google Scholar] [CrossRef]
- Turnbull, M.H.; Murthy, R.; Griffin, K.L. The relative impacts of daytime and night-time warming on photosynthetic capacity in Populus deltoides. Plant. Cell Environ. 2002, 25, 1729–1737. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Richardson, A.D.; Andy Black, T.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Copula Function Name | Mathematical Description |
|---|---|
| BB1 [62] | |
| Clayton [57] | |
| Frank [59] | |
| Gaussian [59] | |
| Gumbel [59] | |
| Joe [59] | |
| t [59] | |
| Tawn [63] |
| Province | Season | Kendall | p Value | Spearman | p Value | Pearson | p Value |
|---|---|---|---|---|---|---|---|
| Guangdong | spring | 0.284 | 0.000 | 0.401 | 0.000 | 0.374 | 0.000 |
| Jiangsu | spring | 0.056 | 0.227 | 0.083 | 0.230 | 0.116 | 0.095 |
| Liaoning | spring | 0.468 | 0.000 | 0.660 | 0.000 | 0.619 | 0.000 |
| Guangdong | summer | 0.224 | 0.000 | 0.327 | 0.000 | 0.331 | 0.000 |
| Jiangsu | summer | 0.366 | 0.000 | 0.522 | 0.000 | 0.532 | 0.000 |
| Liaoning | summer | 0.250 | 0.000 | 0.370 | 0.000 | 0.365 | 0.000 |
| Guangdong | autumn | 0.150 | 0.001 | 0.219 | 0.001 | 0.228 | 0.001 |
| Jiangsu | autumn | 0.418 | 0.000 | 0.591 | 0.000 | 0.585 | 0.000 |
| Liaoning | autumn | 0.598 | 0.000 | 0.789 | 0.000 | 0.771 | 0.000 |
| Province | Season | Kendall | p Value | Spearman | p Value | Pearson | p Value |
|---|---|---|---|---|---|---|---|
| Guangdong | spring | 0.177 | 0.000 | 0.258 | 0.000 | 0.234 | 0.001 |
| Jiangsu | spring | 0.039 | 0.402 | 0.062 | 0.372 | 0.086 | 0.213 |
| Liaoning | spring | 0.501 | 0.000 | 0.696 | 0.000 | 0.662 | 0.000 |
| Guangdong | summer | 0.148 | 0.000 | 0.219 | 0.000 | 0.212 | 0.000 |
| Jiangsu | summer | 0.397 | 0.000 | 0.572 | 0.000 | 0.585 | 0.000 |
| Liaoning | summer | 0.233 | 0.000 | 0.335 | 0.000 | 0.359 | 0.000 |
| Guangdong | autumn | 0.090 | 0.052 | 0.128 | 0.064 | 0.087 | 0.209 |
| Jiangsu | autumn | 0.409 | 0.000 | 0.581 | 0.000 | 0.568 | 0.000 |
| Liaoning | autumn | 0.595 | 0.000 | 0.787 | 0.000 | 0.771 | 0.000 |
| PROVINCE | Guangdong | Jiangsu | Liaoning | |
|---|---|---|---|---|
| Copula Function | BB1 | BB1 | Frank | |
| Spring | AIC | −177.71 | −193.27 | −190.29 |
| Spring | BIC | −177.04 | −192.60 | −189.96 |
| Spring | RMSE | 0.21 | 0.14 | 0.16 |
| Summer | AIC | −277.29 | −315.40 | −271.40 |
| Summer | BIC | −276.54 | −314.65 | −271.03 |
| Summer | RMSE | 0.22 | 0.12 | 0.24 |
| Autumn | AIC | −193.76 | −198.09 | −204.55 |
| Autumn | BIC | −193.09 | −197.43 | −204.22 |
| Autumn | RMSE | 0.14 | 0.13 | 0.11 |
| PROVINCE | Guangdong | Jiangsu | Liaoning | |
|---|---|---|---|---|
| Copula Function | BB1 | BB1 | Frank | |
| Spring | AIC | −177.71 | −193.27 | −190.29 |
| Spring | BIC | −177.04 | −192.60 | −189.96 |
| Spring | RMSE | 0.21 | 0.14 | 0.16 |
| Summer | AIC | −277.29 | −315.40 | −271.40 |
| Summer | BIC | −276.54 | −314.65 | −271.03 |
| Summer | RMSE | 0.22 | 0.12 | 0.24 |
| Autumn | AIC | −193.76 | −198.09 | −204.55 |
| Autumn | BIC | −193.09 | −197.43 | −204.22 |
| Autumn | RMSE | 0.14 | 0.13 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, G.; Li, Z. Asymmetry of Daytime and Nighttime Warming in Typical Climatic Zones along the Eastern Coast of China and Its Influence on Vegetation Activities. Remote Sens. 2020, 12, 3604. https://doi.org/10.3390/rs12213604
He G, Li Z. Asymmetry of Daytime and Nighttime Warming in Typical Climatic Zones along the Eastern Coast of China and Its Influence on Vegetation Activities. Remote Sensing. 2020; 12(21):3604. https://doi.org/10.3390/rs12213604
Chicago/Turabian StyleHe, Guangxin, and Zhongliang Li. 2020. "Asymmetry of Daytime and Nighttime Warming in Typical Climatic Zones along the Eastern Coast of China and Its Influence on Vegetation Activities" Remote Sensing 12, no. 21: 3604. https://doi.org/10.3390/rs12213604
APA StyleHe, G., & Li, Z. (2020). Asymmetry of Daytime and Nighttime Warming in Typical Climatic Zones along the Eastern Coast of China and Its Influence on Vegetation Activities. Remote Sensing, 12(21), 3604. https://doi.org/10.3390/rs12213604