
Application and Development of Bt Insect Resistance Genes in Rice Breeding


Abstract
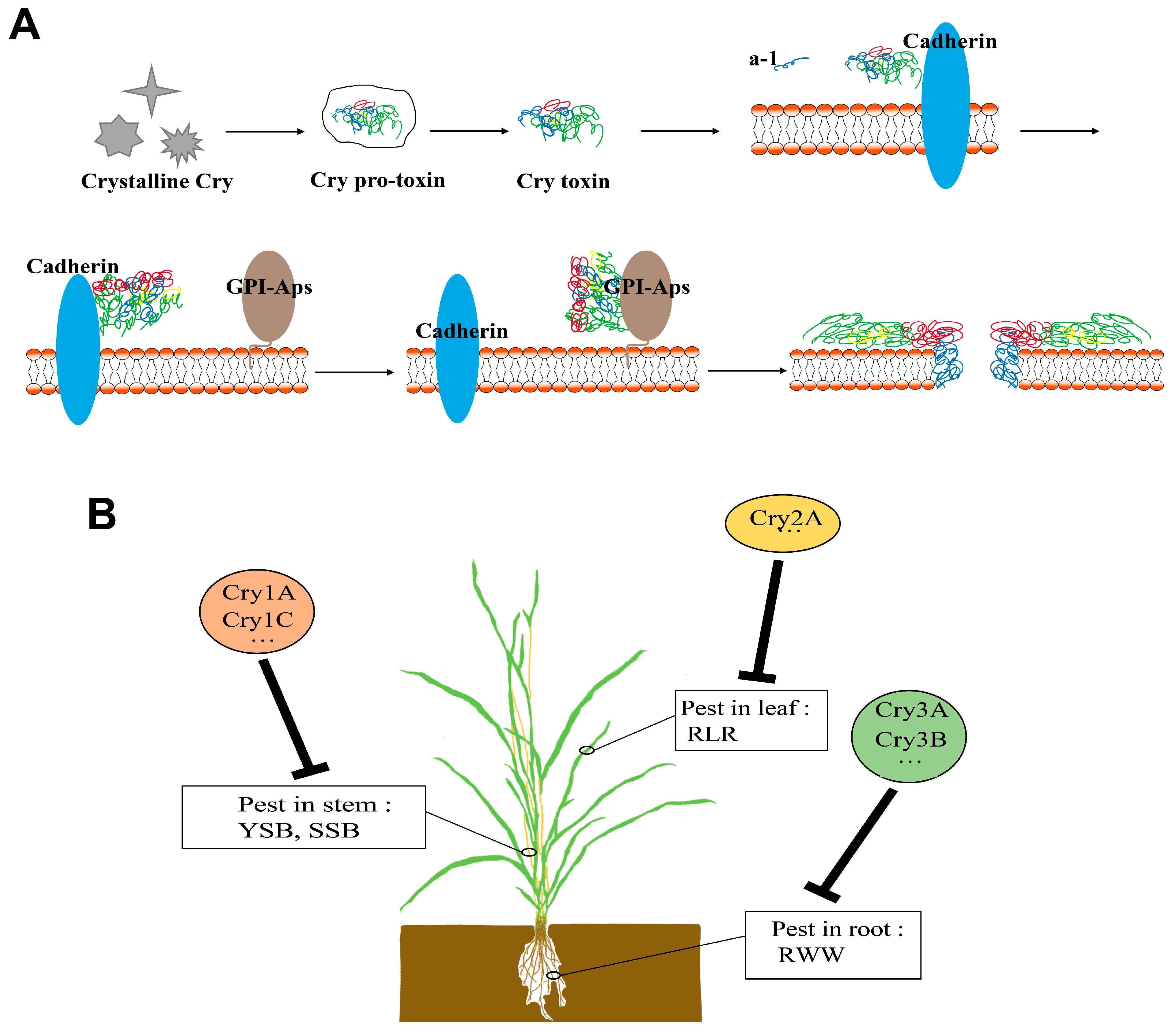
1. Introduction
2. Bt Gene
3. Research Progress of Transgenic Rice That Is Resistant to Lepidoptera Pests
3.1. A Close-To-Commercial Insect-Resistant Rice Strain: Huahui No.1
3.2. Cultivation of Insect-Resistant Transgenic Rice with the New Bt Gene
3.3. Cultivation of Bivalent Bt Insect-Resistant Rice
3.4. The Tissue-Specific Expression of Bt in Insect-Resistant Rice
3.5. Expression of Hybrid Fusion Protein in Transgenic Rice
4. Research Status of the Safety of Transgenic Rice
4.1. Edible Safety
4.2. Environmental Security
4.2.1. Effects on Non-Target Insects
4.2.2. Effects of Bt Rice on the Root Ecosystem
5. Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Sousa, I.G.; Oliveira, J.; Mexia, A.; Barros, G.; Almeida, C.; Brazinha, C.; Vega, A.; Brites, C. Advances in Environmentally Friendly Techniques and Circular Economy Approaches for Insect Infestation Management in Stored Rice Grains. Foods 2023, 12, 511. [Google Scholar] [CrossRef]
- Chen, H.; Lin, Y.; Zhang, Q. Review and prospect of transgenic rice research. Chin. Sci. Bull. 2009, 54, 4049–4068. [Google Scholar]
- Barton, K.A.; Binns, A.N.; Matzke, A.J.; Chilton, M.D. Regeneration of intact tobacco plants containing full length copies of genetically engineered T-DNA, and transmission of T-DNA to R1 progeny. Cell 1983, 32, 1033–1043. [Google Scholar] [CrossRef]
- Abdul, A.M.; Brini, F.; Rouached, H.; Masmoudi, K. Genetically engineered crops for sustainably enhanced food production systems. Front. Plant Sci. 2022, 13, 1027828. [Google Scholar]
- Höfte, H.; Whiteley, H. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [CrossRef]
- Crickmore, N.; Zeigler, D.; Feitelson, J.; Schnepf, E.; Van, R.J.; Lereclus, D.; Baum, J.; Dean, D. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar] [CrossRef]
- Schnepf, H.E.; Whiteley, H. Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli. Proc. Natl. Acad. Sci. USA 1981, 78, 2893–2897. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.; Bonning, B. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2021, 186, 107438. [Google Scholar] [CrossRef]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin. Nat. Biotechnol. 2000, 18, 1101–1104. [Google Scholar] [CrossRef]
- Chen, H.; Tang, W.; Xu, C.; Li, X.; Lin, Y.; Zhang, Q. Transgenic indica rice plants harboring a synthetic cry2A* gene of Bacillus thuringiensis exhibit enhanced resistance against lepidopteran rice pests. Theor. Appl. Genet. 2005, 111, 1330–1337. [Google Scholar] [CrossRef]
- Tang, W.; Chen, H.; Xu, C.; Li, X.; Lin, Y.; Zhang, Q. Development of insect-resistant transgenic indica rice with a synthetic cry1C* gene. Mol. Breed. 2006, 18, 1–10. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, H.; Tang, W.; Hua, H.; Lin, Y. Development and characterisation of transgenic rice expressing two Bacillus thuringiensis genes. Pest Manag. Sci. 2011, 67, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Lou, J.; Gao, N.; Yang, X.; Deng, J. Production and Insect-Resistant Characterization of Hanhui 3T a Derivative Line of Huahui No.1. J. Nucl. Agric. Sci. 2022, 36, 1–6. [Google Scholar]
- Qiu, C.; Sangha, J.S.; Song, F.; Zhou, Z.; Yin, A.; Gu, K.; Tian, D.; Yang, J.; Yin, Z. Production of marker-free transgenic rice expressing tissue-specific Bt gene. Plant Cell Rep. 2010, 29, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Huang, H.; Yang, Z.; Chen, T.; Liu, L.; Li, X.; Chen, H.; Lin, Y. Development of insect-resistant transgenic rice with Cry1C*-free endosperm. Pest Manag. Sci. 2009, 65, 1015–1020. [Google Scholar] [CrossRef]
- Qi, Y.; Ye, S.; Lu, Y.; Jin, Q.; Zhang, X. Development of Marker-Free Transgenic Cry1Ab Rice with Lepidopteran Pest Resistance by Agrobacterium Mixture-Mediated Co-transformation. Rice Sci. 2009, 16, 181–186. [Google Scholar] [CrossRef]
- Fujimoto, H.; Itoh, K.; Yamamoto, M.; Kyozuka, J.; Shimamoto, K. Insect resistant rice generated by introduction of a modified δ-endotoxin gene of Bacillus thuringiensis. Nat. Biotechnol. 1993, 11, 1151–1155. [Google Scholar] [CrossRef]
- Wünn, J.; Klöti, A.; Burkhardt, P.K.; Biswas, G.C.G.; Launis, K.; Iglesias, V.A.; Potrykus, I. Transgenic indica rice breeding line IR58 expressing a synthetic crylA (b) gene from Bacillus thuringiensis provides effective insect pest control. Nat. Biotechnol. 1996, 14, 171–176. [Google Scholar] [CrossRef]
- Ghareyazie, B.; Alinia, F.; Menguito, C.A.; Rubia, L.G.; de Palma, J.M.; Liwanag, E.A.; Cohen, M.B.; Khush, G.S.; Bennett, J. Enhanced resistance to two stem borers in an aromatic rice containing a synthetic cryIA (b) gene. Mol. Breed. 1997, 3, 401–414. [Google Scholar] [CrossRef]
- Nayak, P.; Basu, D.; Das, S.; Basu, A.; Ghosh, D.; Ramakrishnan, N.A.; Ghosh, M.; Sen, S.K. Transgenic elite indica rice plants expressing CryIAc ∂-endotoxin of Bacillus thuringiensis are resistant against yellow stem borer (Scirpophaga incertulas). Proc. Natl. Acad. Sci. USA 1997, 94, 2111–2116. [Google Scholar] [CrossRef]
- Wu, C.; Fan, Y.; Zhang, C.; Oliva, N.; Datta, S. Transgenic fertile japonica rice plants expressing a modified cryIA (b) gene resistant to yellow stem borer. Plant Cell Rep. 1997, 17, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Sardana, R.; Kaplan, H.; Altosaar, I. Agrobacterium-transformed rice plants expressing synthetic cryIA (b) and cryIA (c) genes are highly toxic to striped stem borer and yellow stem borer. Proc. Natl. Acad. Sci. USA 1998, 95, 2767–2772. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.Y.; Shu, Q.Y.; Yao, H.W.; Cui, H.R.; Cheng, X.Y.; Hu, C.; Xia, Y.W.; Gao, M.W.; Altosaar, I. Field evaluation of resistance of transgenic rice containing a synthetic cry1Ab gene from Bacillus thuringiensis Berliner to two stem borers. J. Econ. Entomol. 2001, 94, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Maqbool, S.B.; Riazuddin, S.; Sticklen, M.B. Expression of synthetic cry1Ab and cry1Ac genes in basmati rice (Oryza sativa L.) variety 370 via Agrobacterium-mediated transformation for the control of the European corn borer (Ostrinia nubilalis). Vitr. Cell. Dev. Biol. Plant 2002, 38, 213–220. [Google Scholar] [CrossRef]
- Khanna, H.K.; Raina, S.K. Elite Indica transgenic rice plants expressing modified Cry1Ac endotoxin of Bacillus thuringiensis show enhanced resistance to yellow stem borer (Scirpophaga incertulas). Transgenic. Res. 2002, 11, 411–423. [Google Scholar] [CrossRef]
- Ramesh, S.; Nagadhara, D.; Pasalu, I.; Kumari, A.P.; Sarma, N.; Reddy, V.; Rao, K. Development of stem borer resistant transgenic parental lines involved in the production of hybrid rice. J. Biotechnol. 2004, 111, 131–141. [Google Scholar] [CrossRef]
- Ballester, V.; Granero, F.; Tabashnik, B.E.; Malvar, T.; Ferré, J. Integrative model for binding of Bacillus thuringiensis toxins in susceptible and resistant larvae of the diamondback moth (Plutella xylostella). Appl. Environ. Microb. 1999, 65, 1413–1419. [Google Scholar] [CrossRef]
- Li, H.; González-Cabrera, J.; Oppert, B.; Ferré, J.; Higgins, R.A.; Buschman, L.L.; Radke, G.A.; Zhu, K.Y.; Huang, F. Binding analyses of Cry1Ab and Cry1Ac with membrane vesicles from Bacillus thuringiensis resistant and susceptible Ostrinia nubilalis. Biochem. Biophys. Res. Commun. 2004, 323, 52–57. [Google Scholar]
- Zhao, J.Z.; Li, Y.X.; Collins, H.L.; Cao, J.; Earle, E.D.; Shelton, A.M. Different cross-resistance patterns in the diamondback moth (Lepidoptera: Plutellidae) resistant to Bacillus thuringiensis toxin Cry1C. J. Econ. Entomol. 2001, 94, 1547–1552. [Google Scholar] [CrossRef]
- Luo, S.; Wu, K.; Tian, Y.; Liang, G.; Feng, X.; Zhang, J.; Guo, Y. Cross-resistance studies of Cry1Ac-resistant strains of Helicoverpa armigera (Lepidoptera: Noctuidae) to Cry2Ab. J. Econ. Entomol. 2007, 100, 909–915. [Google Scholar] [CrossRef]
- Caccia, S.; Hernández-Rodríguez, C.S.; Mahon, R.J.; Downes, S.; James, W.; Bautsoens, N.; Van, R.J.; Ferre, J. Binding site alteration is responsible for field-isolated resistance to Bacillus thuringiensis Cry2A insecticidal proteins in two Helicoverpa species. PLoS ONE 2010, 5, e9975. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carrière, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Van Rensburg, J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar] [CrossRef]
- Xu, C.; Cheng, J.; Lin, H.; Gao, J.; Shen, Z. Characterization of transgenic rice expressing fusion protein Cry1Ab/Vip3A for insect resistance. Sci. Rep. 2018, 8, 15788. [Google Scholar] [CrossRef] [PubMed]
- Boddupally, D.; Tamirisa, S.; Gundra, S.; Vudem, D.; Khareedu, V. Expression of hybrid fusion protein (Cry1Ac::ASAL) in transgenic rice plants imparts resistance against multiple insect pests. Sci. Rep. 2018, 8, 8458. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Guo, S.; Chen, X.; Fan, Y. Transgenic rice plants produced by direct uptake of δ-endotoxin protein gene from Bacillus thuringiensis into rice protoplasts. Sci. Agric. Sin. 1989, 22, 605–611. [Google Scholar]
- Wu, G.; Cui, H.; Ye, G.; Xia, Y.; Sardana, R.; Cheng, X.; Li, Y.; Altosaar, I.; Shu, Q. Inheritance and expression of the cry1Ab gene in Bt (Bacillus thuringiensis) transgenic rice. Theor. Appl. Genet. 2002, 104, 727–734. [Google Scholar] [CrossRef]
- Han, L.; Wu, K.; Peng, Y.; Wang, F.; Guo, Y. Efficacy of transgenic rice expressing Cry1Ac and CpTI against the rice leaffolder, Cnaphalocrocis medinalis (Guenee). J. Invertebr. Pathol. 2007, 96, 71–79. [Google Scholar] [CrossRef]
- Wu, D.; Shu, Q.; Ye, Q.; Lei, Z.; Ma, C.; Xia, Y. Comparative studies on major nutritional components and physicochemical properties of the transgenic rice with a synthetic cry1Ab gene from Bacillus thuringiensis. J. Food Biochem. 2003, 27, 295–308. [Google Scholar] [CrossRef]
- Li, X.; Huang, K.; He, X.; Zhu, B.; Liang, Z.; Li, H.; Luo, Y. Comparison of nutritional quality between Chinese indica rice with sck and cry1Ac genes and its nontransgenic counterpart. J. Food Sci. 2007, 72, S420–S424. [Google Scholar] [CrossRef]
- Cao, S.; Xu, W.; Luo, Y.; He, X.; Yuan, Y.; Ran, W.; Liang, L.; Huang, K. Metabonomics study of transgenic Bacillus thuringiensis rice (T2A-1) meal in a 90-day dietary toxicity study in rats. Mol. Biosyst. 2011, 7, 2304–2310. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; He, X.; Xu, W.; Luo, Y.; Yuan, Y.; Liu, P.; Cao, B.; Shi, H.; Huang, K. Safety assessment of transgenic Bacillus thuringiensis rice T1c-19 in Sprague–Dawley rats from metabonomics and bacterial profile perspectives. IUBMB Life 2012, 64, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xu, W.; Luo, Y.; Liu, H.; Lu, J.; Su, C.; Huang, K. Effects of genetically modified T2A-1 rice on faecal microflora of rats during 90 day supplementation. J. Sci. Food Agric. 2011, 91, 2066–2072. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xu, W.; He, X.; Liu, H.; Cao, S.; Qi, X.; Huang, K.; Luo, Y. Effects of genetically modified T2A-1 rice on the GI health of rats after 90-day supplement. Sci. Rep. 2013, 3, 1962. [Google Scholar]
- Wang, Z.H.; Wang, Y.; Cui, H.r.; Xia, Y.W.; Altosaar, I.; Shu, Q.Y. Toxicological evaluation of transgenic rice flour with a synthetic cry1Ab gene from Bacillus thuringiensis. J. Sci. Food Agric. 2002, 82, 738–744. [Google Scholar] [CrossRef]
- Schrøder, M.; Poulsen, M.; Wilcks, A.; Kroghsbo, S.; Miller, A.; Frenzel, T.; Danier, J.; Rychlik, M.; Emami, K.; Gatehouse, A. A 90-day safety study of genetically modified rice expressing Cry1Ab protein (Bacillus thuringiensis toxin) in Wistar rats. Food Chem. Toxicol. 2007, 45, 339–349. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, Y.; Li, Y.; Wang, J.; Ding, J.; Chen, X.; Peng, Y. A 90 Day Safety Assessment of Genetically Modified Rice Expressing Cry1Ab/1Ac Protein using an Aquatic Animal Model. J. Agric. Food Chem. 2015, 63, 3627–3633. [Google Scholar] [CrossRef]
- Wang, E.H.; Yu, Z.; Hu, J.; Xu, H.B. Effects of 90-day feeding of transgenic Bt rice TT51 on the reproductive system in male rats. Food Chem. Toxicol. 2013, 62, 390–396. [Google Scholar] [CrossRef]
- Wang, E.H.; Yu, Z.; Hu, J.; Jia, X.D.; Xu, H.B. A two-generation reproduction study with transgenic Bt rice TT51 in Wistar rats. Food Chem. Toxicol. 2014, 65, 312–320. [Google Scholar]
- Zhang, M.; Zhuo, Q.; Tian, Y.; Piao, J.; Yang, X. Long-term toxicity study on transgenic rice with Cry1Ac and sck genes. Food Chem. Toxicol. 2014, 63, 76–83. [Google Scholar] [CrossRef]
- Bai, Y.; Jiang, M.; Cheng, J.; Wang, D. Effects of Cry1Ab toxin on Propylea japonica (Thunberg) (Coleoptera: Coccinellidae) through its prey, Nilaparvata lugens Stål (Homoptera: Delphacidae), feeding on transgenic Bt rice. Environ. Entomol. 2006, 35, 1130–1136. [Google Scholar] [CrossRef]
- Chen, M.; Ye, G.; Liu, Z.; Yao, H.; Chen, X.; Shen, Z.; Hu, C.; Datta, S. Field assessment of the effects of transgenic rice expressing a fused gene of cry1Ab and cry1Ac from Bacillus thuringiensis Berliner on nontarget planthopper and leafhopper populations. Environ. Entomol. 2006, 35, 127–134. [Google Scholar] [CrossRef]
- Chen, M.; Liu, Z.; Ye, G.; Shen, Z.; Hu, C.; Peng, Y.; Altosaar, I.; Shelton, A.M. Impacts of transgenic cry1Ab rice on non-target planthoppers and their main predator Cyrtorhinus lividipennis (Hemiptera: Miridae)—A case study of the compatibility of Bt rice with biological control. Biol. Control 2007, 42, 242–250. [Google Scholar] [CrossRef]
- Lu, Z.; Tian, J.; Wang, W.; Xu, H.; Hu, C.; Guo, Y.; Peng, Y.; Ye, G. Impacts of Bt rice expressing Cry1C or Cry2A protein on the performance of nontarget leafhopper, Nephotettix cincticeps (Hemiptera: Cicadellidae), under laboratory and field conditions. Environ. Entomol. 2014, 43, 209–217. [Google Scholar] [CrossRef]
- Lu, Z.; Han, N.; Tian, J.; Peng, Y.; Cui, H.; Guo, Y.; Shen, Z.; Ye, G. Transgenic cry1Ab/vip3H+epsps rice with insect and herbicide resistance acted no adverse impacts on the population growth of a non-target herbivore, the white-backed planthopper, under laboratory and field conditions. J. Integr. Agric. 2014, 13, 2678–2689. [Google Scholar] [CrossRef]
- Han, Y.; Chen, J.; Wang, H.; Zhao, J.; He, Y.; Hua, H. Prey-mediated effects of transgenic cry2Aa rice on the spider Hylyphantes graminicola, a generalist predator of Nilapavarta lugens. Biol. Control 2015, 60, 251–261. [Google Scholar] [CrossRef]
- Han, Y.; Wang, H.; Chen, J.; Cai, W.; Hua, H. No impact of transgenic cry2Aa rice on Anagrus nilaparvatae, an egg parasitoid of Nilaparvata lugens, in laboratory tests. Biol. Control 2015, 82, 46–51. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Romeis, J.; Liu, Q.; Lin, K.; Chen, X.; Peng, Y. Bt rice expressing Cry2Aa does not cause direct detrimental effects on larvae of Chrysoperla sinica. Ecotoxicology 2013, 22, 1413–1421. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Romeis, J.; Chen, X.; Zhang, J.; Chen, H.; Peng, Y. Consumption of Bt rice pollen expressing Cry2Aa does not cause adverse effects on adult Chrysoperla sinica Tjeder (Neuroptera: Chrysopidae). Biol. Control 2012, 61, 246–251. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Chen, X.; Romeis, J.; Yin, X.; Peng, Y. Consumption of Bt rice pollen containing Cry1C or Cry2A does not pose a risk to Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Sci. Rep. 2015, 5, 7679. [Google Scholar] [CrossRef]
- Sun, X.; Yan, M.; Zhang, A.; Wang, M. Transgenic cry1C gene rough rice line T1C-19 does not change the host preferences of the non-target stored product pest, Rhyzopertha dominica (Fabricius) (Coleoptera: Bostrichidae), and its parasitoid wasp, Anisopteromalus calandrae (Howard) (Hymenoptera: Pteromalidae). Ecotox Environ. Saf. 2015, 120, 449–456. [Google Scholar]
- Ren, S.; Yang, F.; Gao, M.; Pu, D.; Shi, M.; Ye, G.; Shen, Z.; Chen, X. Effects of Transgenic Bt Rice on Nontarget Rhopalosiphum maidis (Homoptera: Aphididae). Environ. Entomol. 2016, 45, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Ye, Q.; Min, H.; Duan, X.; Jin, W. Bt-transgenic rice straw affects the culturable microbiota and dehydrogenase and phosphatase activities in a flooded paddy soil. Soil Biol. Biochem. 2004, 36, 289–295. [Google Scholar]
- Wu, W.; Ye, Q.; Min, H.; Chen, H. Effect of cry1Ab toxin released from straw of Bt-transgenic rice on microflora and enzymatic activities in upland soil. Acta Pedol. Sin. 2003, 40, 606–612. [Google Scholar]
- Wang, J.; Chen, X.; Liang, Y.; Zhu, H.; Ding, J.; Peng, Y. Influence of transgenic rice expressing a fused Cry1Ab/1Ac protein on frogs in paddy fields. Ecotoxicology 2014, 23, 1619–1628. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Q.; Wang, W.; Wu, L.; Wu, W. Cry1Ab protein from Bt transgenic rice does not residue in rhizosphere soil. Environ. Pollut. 2006, 143, 449–455. [Google Scholar] [CrossRef]
- Li, Y.; Wu, K.; Zhang, Y.; Yuan, G. Degradation of Cry1Ac protein within transgenic Bacillus thuringiensis rice tissues under field and laboratory conditions. Environ. Entomol. 2007, 36, 1275–1282. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, J.; Zhang, J.; Jia, R. [Accumulation of Cry proteins in soil released from Bt rice after planting for multiple years]. Ying Yong Sheng Tai Xue Bao 2022, 33, 119–125. [Google Scholar]
- Un Jan Contreras, S.; Gardner, C.M. Environmental fate and behaviour of antibiotic resistance genes and small interference RNAs released from genetically modified crops. J. Appl. Microbiol. 2022, 133, 2877–2892. [Google Scholar] [CrossRef]


{kind=link}
| Nomenclature Gene [5] | Nomenclature Gene [8] | Insect Spectrum | Number of Amino Acids | Applied Crops |
|---|---|---|---|---|
| cryⅠA(a) | cry1Aa | Lepidoptera | 1176 | Rice, Sweet potato |
| cryⅠA(b) | cry1Ab | Lepidoptera | 1155 | Rice, Maize, Cotton |
| cryⅠA(c) | cry1Ac | Lepidoptera | 1178 | Rice, Maize, Cotton |
| cryⅠB | cry1Ba | Lepidoptera | 1207 | Rice, Maize |
| cryⅠC | cry1Ca | Lepidoptera | 1189 | Rice, Maize, Cotton |
| cryⅠD | cry1Da | Lepidoptera | 1165 | Maize |
| cryⅡA | cry2Aa | Lepidoptera/Diptera | 633 | Rice |
| cryⅡB | cry2Ab | Lepidoptera | 633 | Maize, Cotton |
| cryⅢA | cry3Aa | Coleoptera | 644 | Rice, Potato |
| cryⅢB | cry3Ba | Coleoptera | 649 | Rice |
| cryⅣA | cry4Aa | Diptera | 1180 | |
| cryⅣB | cry4Ba | Diptera | 1136 | |
| cryⅣC | cry10Aa | Diptera | 675 | |
| cryⅣD | cry11Aa | Diptera | 643 | |
| cytB | cyt1Aa | Diptera/others | 248 |
| Rice Variety | Rice Type | Gene | Reference |
|---|---|---|---|
| Huahui No.1 | Xian/indica rice | cry1Ab/1Ac | [9] |
| BT Shanyou 63 | Xian/indica rice | cry1Ab/1Ac | |
| Minghui 63 | Xian/indica rice | cry2A*(2A) | [10] |
| Minghui 63 | Xian/indica rice | cry1C*(1C) | [11] |
| Minghui 63 | Xian/indica rice | cry1Ab/1C | [12] |
| Minghui 63 | Xian/indica rice | cry1Ab/2A | |
| Minghui 63 | Xian/indica rice | cry1Ac/1C | |
| Minghui 63 | Xian/indica rice | cry1Ac/2A | |
| Minghui 63 | Xian/indica rice | cry1C/2A | |
| Hanhui 3T | Xian/indica rice | cry1Ab/cry1Ac | [13] |
| Huhan 5A/Hanhui 3T | Hybrid rice | cry1Ab/cry1Ac | |
| Nip | Geng/japonica rice | cry1Ab/1Ac | [14] |
| ZH11 | Geng/japonica rice | cry1C* | [15] |
| Zhejing 22 | Geng/japonica rice | cry1Ab | [16] |
| Zhejing 27 | Geng/japonica rice | cry1Ab | |
| Jiahua 1 | Geng/japonica rice | cry1Ab | |
| Xiushui 63 | Geng/japonica rice | cry1Ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Wang, J.; Ling, F.; You, A. Application and Development of Bt Insect Resistance Genes in Rice Breeding. Sustainability 2023, 15, 9779. https://doi.org/10.3390/su15129779
Li C, Wang J, Ling F, You A. Application and Development of Bt Insect Resistance Genes in Rice Breeding. Sustainability. 2023; 15(12):9779. https://doi.org/10.3390/su15129779
Chicago/Turabian StyleLi, Changyan, Jianyu Wang, Fei Ling, and Aiqing You. 2023. "Application and Development of Bt Insect Resistance Genes in Rice Breeding" Sustainability 15, no. 12: 9779. https://doi.org/10.3390/su15129779
APA StyleLi, C., Wang, J., Ling, F., & You, A. (2023). Application and Development of Bt Insect Resistance Genes in Rice Breeding. Sustainability, 15(12), 9779. https://doi.org/10.3390/su15129779