Abstract
The aim of our study was to evaluate the discriminative value of gene and protein expression levels of the inflammatory marker (YKL-40) and lysosome-associated membrane protein 1 and 2 (LAMP-1 and LAMP-2) in patients with central nervous system (CNS) infections. Thirty hospitalized patients with CNS infections and undefined etiology, and 22 healthy subjects as a control group, were included in the study. Gene expression levels of YKL-40, LAMP-1 and LAMP-2 were determined by qPCR. Plasma and CSF concentrations of the tree proteins of interest were detected by ELISA. Our results showed that mRNA levels of YKL-40 were significantly downregulated in WBCs of patients compared to controls, while plasma YKL-40 concentrations were higher. LAMP-1 significantly increased in patients’ plasma and CSF was found. Patients were subdivided depending on the confirmed or presumed etiological agent into two subgroups groups—patients with bacterial or with viral neuroinfection. Differences between plasma levels of YKL-40 in the subgroups when matched with controls were detected. The concentrations of the glycoprotein were higher in patients with bacterial infections compared to those with the viral ones. We revealed that LAMP-1 plasma levels were also significantly increased in patients with viral infections in comparison to healthy individuals. We could speculate that YKL-40 plasma levels might serve as a fast discriminative tool to support the presence of viral or bacterial CNS infections.
1. Introduction
Central nervous system (CNS) infections, with meningitis, encephalitis, and brain abscess being the most common presentation, are generally considered rare clinical entities [1,2]. The diverse etiology hampers pathogen recognition and thus the delay in choosing the most appropriate therapy negatively influences patient outcomes [3]. Microorganisms induce a host immune response in the intrathecal space, characterized by the production of various pro- and anti-inflammatory mediators, which considerably affect the disease progression and outcome [4,5,6,7]. Microglia is involved in response to infection with functions involving pathogen recognition, induction and modulation of the immune response, and clearing of microorganisms [8]. Dysregulated immune response during CNS infections may be life-threatening, and treatment alone could be insufficient to control disease progression [9].
The protein YKL-40, also known as CHI3L1, is a part of the glycoside hydrolase family 18, a mammalian chitinase family. In the CNS, YKL-40 is mostly expressed by microglia, especially when responding to acute and chronic inflammation [10]. YKL-40 has shown to be a promising predictive marker for brain injury severity but appears to be nonspecific to CNS infections only. Therefore, interpreting the available literature results needs to be more thoroughly explored. The glycoprotein YKL-40 is intensively investigated as a biomarker involved in inflammatory and tumor processes [11,12]. It is reported to participate in a variety of conditions related to proliferation, angiogenesis, tissue remodeling, and fibrosis [13]. Few studies are showing elevated YKL-40 in serum or cerebrospinal fluid (CSF) in traumatic brain injury [14,15,16]. However, none of them focused on its gene and protein expression nor on the relationship with autophagy in CNS infections.
It was suggested that autophagy during such infections has an essential housekeeping role by eliminating defective proteins and organelles and supporting neuronal plasticity. Dysfunctional autophagy is often associated with uncontrolled inflammatory responses during infection. Lysosomes have a major role in autophagy and lysosome-associated membrane proteins (LAMPs) are an important constituent of the lysosomal membrane. LAMP-1 and LAMP-2 are both type I transmembrane proteins. Each of them has a large highly glycosylated domain and a short C-terminal cytoplasmic tail [17]. They share 37% amino acid sequence identity but are distinct proteins that most likely diverged relatively early in evolution [18]. Both proteins are well characterized in mammalian cells but their importance in CNS infections remains unknown.
Neither YKL-40 nor LAMPs are absolutely cell or tissue-specific as different sources may contribute to their total serum/plasma concentrations. It is unclear if the levels of YKL-40 alone or in combination with LAMPs are associated with disease outcomes, especially in brain pathology. Therefore, new non-invasive biomarkers are explored to help the monitoring of disease progression and outcome.
The aim of the present study was to evaluate how plasma and cerebrospinal levels of YKL-40, independently or together with LAMPs, correlated to the etiology in CNS infections. We present novel data on gene and protein expression of these molecules in patients with bacterial and viral CNS infections to evaluate their discriminative potential.
2. Materials and Methods
2.1. Study Design and Patients’ Selection
We performed a prospective study on 29 patients presenting with both clinical and laboratory abnormalities consistent with an acute CNS infection. All patients were admitted to the Clinic of Infectious Diseases at St. George University Hospital and received standard care following the generally accepted algorithm for the diagnosis and management of CNS infections. Patients were evaluated for the presence of specific clinical signs and symptoms of neuroinfection as well as by standard clinical laboratory and microbiological examination. Clinical laboratory parameters like white blood cell (WBC) count, absolute neutrophil count, absolute lymphocyte count, erythrocyte sedimentation rate (ESR), serum C-reactive protein (CRP), and fibrinogen were tested. The CSF analysis included white blood cell count, total protein content, as well as microbiological tests to determine the etiological agent.
The following inclusion criteria for patient selection were applied:
- age > 18 years
- individuals with clinical presentation of acute CNS infection—fever, headache, vomiting, seizures, disturbances of consciousness, signs of meningeal irritation (neck stiffness, Kernig’s and Brudzinski’s signs).
- patients with abnormal CSF findings consistent with acute CNS infections including the presence of CSF pleocytosis, increased protein levels, changes in CSF glucose levels or in the cerebrospinal fluid/serum glucose ratio.
Patients with the following neurological disorders were not included in the current study—stroke, lentiviral encephalitis, traumatic brain injury, amyotrophic lateral sclerosis, multiple sclerosis, and Alzheimer’s disease.
Patients were classified into two groups of observation based on the medical history, measured CSF abnormalities, CSF appearance, clinical course of the neuroinfection, and microbiological result (where present).
Written informed consent was signed by all patients involved in the study or by their relatives. All procedures were performed in accordance with the 1964 Helsinki Declaration and its later amendments. The study was approved by the University Ethics Committee (Protocol No. 1/28.02.2019).
2.2. Control Group
The control group consisted of 22 healthy individuals, based on the following selection criteria—age > 18 years, no present or inherited illnesses, and written informed consent. The mean age of the control group is 37 years, as 70% are females and 30% are males. A physical examination was performed to confirm that they fit the inclusion criteria. Plasma and white blood cells were isolated. In addition, CSF samples were collected from forensic autopsies of 16 adult cadavers (healthy individuals before death). The causes of death were gunshots, car accidents, and hanging. CSF was taken fresh out of the lateral ventricles. All plasma and CSF samples were stored at −80 °C until analysis.
2.3. Laboratory Methods
All biological samples, including both blood and CSF, were collected within several hours after hospital admission of patients presenting with acute CNS infection. CSF samples from patients were collected via lumbar puncture (LP) and blood specimens were taken by venipuncture in EDTA-Vacutainer monovettes (S—Monovette 2.6 mL, Z—Sarstedt). Both biological samples were collected in parallel at the time of hospital admission and stored at −80 °C until to be assayed, but no longer than 6 months.
2.3.1. Microbiological Evaluation
We performed routine microbiological examination of CSF samples including direct microscopy, CSF culture, and identification. Gram and methylene blue stains were used for direct microscopy. The microscope slides were subsequently observed on 100× immersion oil light microscopy. CSF specimens were cultured on two sets of 5% sheep blood agar and Levinthal’s medium, incubated for up to 48 h at 36 ± 1 °C. The first set was placed in a 5–10% CO2 enriched atmosphere, while the other was incubated without CO2 along with eosin-methylene blue agar (Levine’s medium) for Gram (-) negative bacteria. A selective medium for fungi (Sabouraud-dextrose agar) was also inoculated and kept for up to 14 days. The isolates were identified by classical microbiological identification tests and/or by MALDI-TOF (bioMerieux, Lion, France).
In addition, CSF specimens were also subjected to analysis by a multiplex PCR (BioFire, FilmArray, bioMerieux, France) applying the Meningitis/Encephalitis Panel. The method allows for the simultaneous detection of 14 common bacterial, viral, and fungal targets, associated with acute CNS infection—Escherichia coli K1, Haemophilus influenzae, Listeria monocytogenes, Neisseria meningitidis, Streptococcus agalactiae, Streptococcus pneumoniae, Cryptococcus spp. (C. neoformans/C. gattii), Cytomegalovirus (CMV), Enterovirus (EV), Herpes simplex virus 1 (HSV-1), Herpes simplex virus 2 (HSV-2), Human herpesvirus 6 (HHV-6), Human parechovirus (HPeV), and Varicella zoster virus (VZV).
2.3.2. Gene Expression of YKL-40, LAMP-1, LAMP-2
- Isolation of RNA from blood cells—RNA was isolated from WBC using TRIzol (Thermo Fisher Scientific, Waltham, MA, USA, Lot. No. 1559602,) following the manufacturer’s instructions. After total RNA extraction, samples were treated with the TURBO DNA-free kit (Thermo Fisher Scientific, Waltham, MA, USA, Lot. No. AM1907) to remove residual DNA. Extracted RNAs were quantified at 260/280 nm absorbance by NanoDrop Nucleic Acid Quantification (Thermo Fisher Scientific, Waltham, MA, USA).
- Reverse transcription and qPCR—2 µg of total RNA was reverse transcribed by RevertAid H minus First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA, Lot. No. 01189716) according to the manufacturer’s instructions. The resulting cDNA was used to quantify YKL-40, LAMP-1, LAMP-2 expression. The cDNA was then used as a template for amplification in a quantitative PCR reaction by Genaxon GreenMasterMix (2×) (Genaxxon bioscience GmbH, Ulm, Germany, Cat. No. M3023.0500) following the manufacturer’s recommendations. The following specific primers were used for RNA transcripts of: YKL-40 (Fw 5′-CTGCTCCAGTGCTGCTCT-3′, Rev 5′-TACAGAGGAAGCGGTCCAAGG-3′), LAMP-1 (Fw 5′-CTCTAATGTCTGCAGCTCAAGG-3′, Rev 5′-TGTACACAGCGCAGAACAGG-3′), LAMP-2 (Fw5′-ACAACAGTGGATCAGACAGTACG-3′, Rev5′-AGCAGCAAGCATCAGTTCTTC-3′).
The obtained expression values were normalized to the levels of GAPDH (Fw 5′-AGG TCCACCACTGACACGTTG-3′, Rw 5′-AGCTGAACGGGATGCTCACT-3′) and hUBC (Fw 5′TCCTCAGGCAGAGGTTGATCTT-3′, Rev 5′-GGACCAAGTGCAGAGTGGACTCTT-3′) (Integrated DNA Technologies, Leuven, Belgium). qPCR reactions were performed on a Rotor-Gene Q 600 (Qiagen, Hilden, Germany) using the comparative 2−ΔΔCt method. All samples were analyzed in duplicate.
2.3.3. Detection of YKL-40, LAMP-1 and LAMP-2 in Plasma and CSF by ELISA
The plasma was transferred to an Eppendorf tube and stored at −80 °C after centrifugation, to be used for measurement of protein levels. Plasma levels for each of YKL-40 and LAMPs were measured using specific individual ELISA kits according to the manufacturer’s recommendations. Stored plasma and CSF were analyzed with the following kits: YKL-40 (YKL-40 Human ELISA Kit, Microvue, YKL-40 cat. No. 8020), LAMP- 1 (AssayGenie, Ireland, cat. No. HUDL01679), LAMP-2 (AssayGenie, Dublin, Ireland, cat. No. HUDL01680).
All samples were measured in duplicate. The assays employ the sandwich-based ELISA method performed on a SUNRISE reader (TECAN Life Sciences, Zurich, Switzerland) at 405 nm wavelength. Data were then averaged for each sample and run for statistical analysis. The ELISA Kit’s sensitivity and detection range was as follows: YKL-40 5.4 ng/mL, 15.6–300 ng/mL; LAMP-1 0.094 ng/mL, 0.154–10 ng/mL; LAMP-2 0.023 ng/mL, 62.5–4000 pg/mL.
2.4. Statistical Analysis
In respect to continuous data distribution parametric or non-parametric statistical tests were applied. The level of significance was set at 0.05.
3. Results
- 1.
- Microbiological results and groups of observation
Based on the microbiological evaluation of CSF specimens we managed to culture the following bacterial pathogens—S. pneumoniae (n = 2), N. meningitidis serogroup C (n = 1), L. monocytogenes (n = 1), and H. influenzae type b. All the bacterial pathogens were culture-positive, confirmed by the mPCR ME Panel, and also identified by the MALDI-TOF MS. Microbial cultures showed no fungal growth on SDA agar plates in all the tested CSF samples. Among the CSF samples, we managed to detect only two viral pathogens by the mPCR ME Panel—VZV and HSV-1. Data are summarized in Table 1.

Table 1.
Demographic and laboratory characteristics of the studied patients.
Based on the clinical course, laboratory findings, and microbiological results, we subdivided the patients into two groups. The first group of verified viral or suspected viral neuroinfections (VN) included 22 patients, of which 64% were males (n = 22). The other group consisted of 7 patients with verified bacterial or suspected bacterial neuroinfections (BN), of which 29% were males (n = 2).
The median levels of CSF WBC and total protein were higher in the group of BN when compared to the patients with VN. The same tendency was observed for blood parameters like WBCs, neutrophils, lymphocytes, fibrinogen, CRP, and erythrocyte sedimentation rate—Table 2.
Table 2.
Demographic and laboratory characteristics of patients with CNS infections by groups of observation.
- 2.
- Gene and protein expression of YKL-40
To gain more detailed information on YKL-40’s involvement in cases with CNS infections we evaluated its gene and protein expression. mRNA levels of YKL-40 were significantly downregulated in WBCs of patients with CNS infections (p = 0.008) compared to controls (Figure 1).
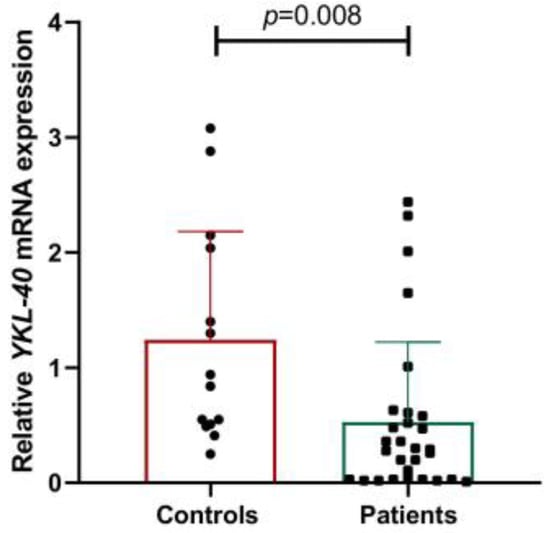
Figure 1.
Gene expression of YKL-40 in controls and patients with CNS infections.
We detected statistically elevated concentrations of both plasma and CSF YKL-40 in patients compared to the control group (p = 0.0005 and p < 0.03 respectively) (Figure 2a,b). In addition, we observed that YKL-40 levels in the CSF of patients were significantly higher compared to plasma levels (p = 0.0021), supporting the inflammatory process within the CNS (Figure 2c).
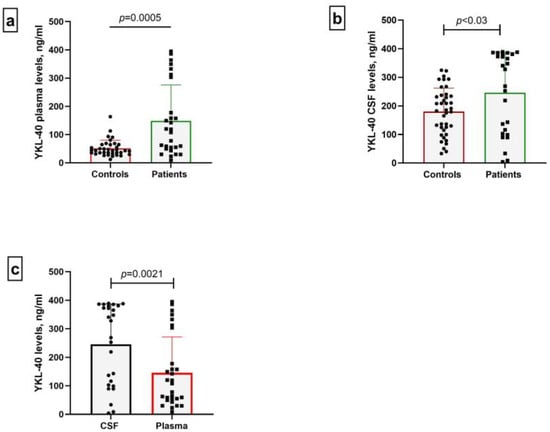
Figure 2.
Plasma (a) and CSF (b) levels of YKL-40 in controls and patients with CNS infections. Glycoprotein levels in both biological specimens were compared. In addition, a parallel group analysis was performed between plasma and CSF samples of patients (c).
Subsequently, after splitting the patients into two subgroups of VN and BN, we found that the plasma YKl-40 levels were higher in BN in comparison with VN.
Higher CSF YKL-40 levels compared to plasma levels in patients with viral infection were detected. There was no change in plasma or CSF levels in bacterial infections. Plasma concentrations of the glycoprotein were higher in patients with viral infections compared to bacterial ones (p = 0.01), while there was no difference between CSF levels of YKL-40 in both patient groups.
- 3.
- Gene and protein expression of LAMPs
Next, we followed a similar approach to determine LAMP-1 and LAMP-2 in CNS infections. We observed a decrease in LAMPs expression in WBCs of patients with CNS infections compared to controls, with a statistically significant difference in the case of LAMP-1 (p = 0.02, Figure 3a), while no change in LAMP-2 was found. However, a positive correlation between LAMP-1 and LAMP-2 FC values in patients was detected (r = 0.499; p = 0.006, Figure 3b). There is a positive linear correlation between LAMP-2 and YKL-40 gene expression in the patients’ group (r = 0.615, p = 0.0004) (Figure 3b).
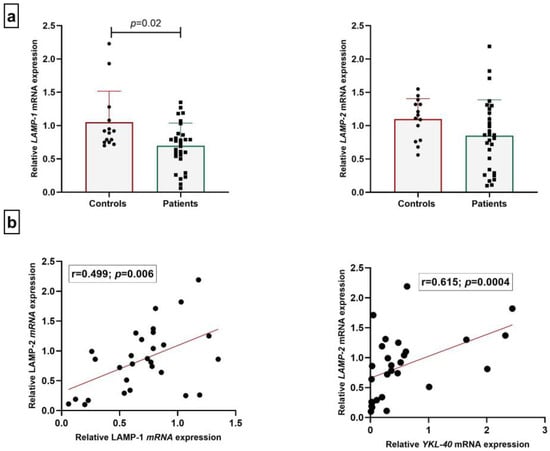
Figure 3.
Relative mRNA expression of LAMPs (a) and correlation analysis (b).
Regarding the protein levels of both glycoproteins, we found that plasma and CSF LAMP-1 concentrations were significantly higher in patients in comparison to the control group (p < 0.0001; p = 0.04 respectively). Values for LAMP-2 in plasma and CSF samples showed different patterns, with those in plasma being lower (p < 0.0001), while the CSF levels were higher in patients compared to the control group, but without statistical significance.
The levels of LAMP-1 and LAMP-2 glycoproteins varied among samples and in some cases, the concentration was under the detection minimum of the commercial ELISA kit. We found that the levels of LAMP-1 and LAMP-2 were dominantly between the limit of quantification (LOQ) and 4.99 ng/mL. Only, in some cases, concentrations higher than 20.00 ng/mL, were observed. Interestingly, there was no variance in plasma LAMP-1 concentrations between viral and bacterial-induced conditions. However, we revealed that LAMP-1 plasma levels significantly increased in patients with viral infections in comparison to healthy individuals (p = 0.0002). No change in plasma or CSF levels of LAMP-2 between the two patient subgroups was observed. Summarized information of samples based on plasma and CSF levels of the glycoproteins is shown in Table 3.
Table 3.
Subgroup analysis of plasma and CSF LAMP-1 and LAMP-2 levels in patients with CNS infection.
- 4.
- Discriminative power of YKL-40 and LAMPs
To determine whether plasma YKL-40 could serve as a potential biomarker to distinguish patients from healthy controls, a ROC curve was constructed. The high discriminative power of YKL-40 plasma levels (AUC = 0.746; p = 0.0007) was detected (Figure 4). Our results revealed that YKL-40 plasma levels could be used as potential marker in CNS infections. AUC/ROC analysis showed no statistical significance when applied to patient subgroups of VN and BN. In addition, we evaluated the discriminative power of both LAMPs, but there was no significance in the values.
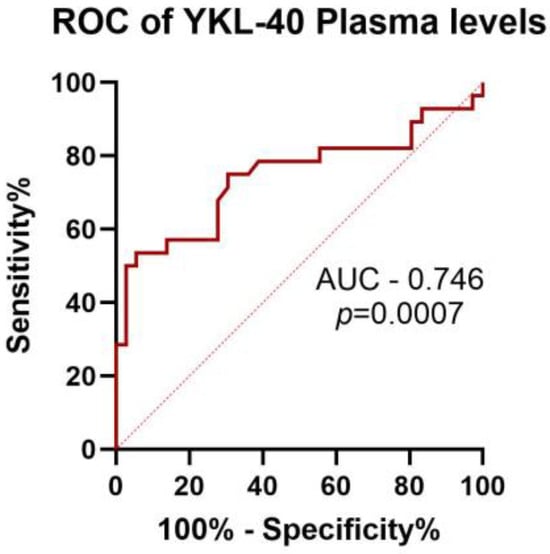
Figure 4.
ROC curves of YKL-40 plasma levels to discriminate between neuroinfections and healthy controls.
4. Discussion
There are several major challenges that meet the diagnosis and epidemiological surveillance of CNS infections. The identification of unknown pathogens that may cause or be involved in the development of these diseases reduces the application of adequate treatment. In addition, outbreaks triggered by emerging vector-borne pathogens that spread into new geographical regions not previously affected [19], cause difficulties in engaging and proper handling of the epidemics. Third, the rise of microorganisms that are resistant to antibiotics limits the options for disease management and fast curing of patients. Both inflammation and autophagy could be triggered by pathogens and their interaction continues during the host response [20].
Autophagy is an essential process by which cytoplasmic molecules are delivered to the lysosome for degradation and recycling [21]. It is induced under stressful conditions such as the absence of growth factors, low oxygen levels, or nutrients necessary for cell survival. It is also required for development, differentiation, homeostasis and, in specific conditions, for cell death [22]. We aimed to find a possible relationship between the etiological agent in CNS infections and the gene or protein expression of autophagy-associated molecules LAMP-1 and LAMP-2. Our results showed that only LAMP-1 plasma and CSF concentrations were increased in patients compared to the control group. No change in protein or gene expression of LAMP-2 was observed. It was shown that several herpes viruses negatively regulate autophagy. Orvedahl et al. determined that the herpes simplex virus type 1 (HSV-1) protein can inhibit autophagy via two pathways: the dephosphorylation of translation initiation factor (eIF2D) and the binding to Beclin-1 [23]. Researchers concluded that in an ongoing race for survival, viruses have acquired the potent ability to repurpose autophagy for their benefit. They proved that LAMP-2 is involved in the viral cycle of HSV-1 [23]. Although autophagy is intensively investigated in CNS infection, studies on LAMP-1 and LAMP-2 proteins are insufficient. As data for such analysis are lacking in the literature, the interpretation and explanation of these results are quite difficult. Our previous experience shows that these molecules change their cellular localization in tumor cell lines and their membrane expression promotes binding to extracellular matrix molecules and thus facilitates tumor progression [24]. It was detected that the LAMP-1 gene was overexpressed in high-grade glioma [25]. Furthermore, it was found that LAMP-2 deficiency significantly attenuates the neurodegenerative effects of increased phosphorylated tau and inhibition of Aβ secretion induced by HSV-1 in chronic conditions [26]
The interplay between autophagy and inflammation is essential for maintaining cellular homeostasis and health. Both processes are the two sides of the same coin and their interaction is required for keeping the balance in healthy status. Dysfunctional autophagy often leads to uncontrolled inflammatory responses triggered by infection, resulting in severe host damage [27]. It is still unclear under which circumstances enhanced autophagy plays a role in cell death or represents a mechanism with protective effects [28].
The expression of YKL-40 is related to inflammatory responses and it is involved in remodeling of the extracellular matrix. Accumulative evidence has highlighted the role of YKL-40 in neuroinflammation [29,30]. We found that protein levels of YKL-40 were significantly different between patients and healthy individuals. In accordance with our data, several studies reported that YKL-40 levels in CSF and serum were highly elevated in different pathologies of the CNS, compared to those of the controls. It was estimated that CSF levels correlated with the severity and prognosis of Anti-leucine-rich glioma inactivated 1(LGI1) encephalitis [31]. Some researchers proposed that CSF YKL-40 might be a candidate biomarker for understanding the pathogenesis of HIV infection in the CNS. The strong correlation between CSF YKL-40 and the neurofilament light protein indicated a pathogenic association between astroglia activation and axonal injury [32]. YKL-40 is a marker largely related to the pathogenesis of diseases, sharing chronic inflammatory features and high cellular activity [33]. On the other hand, C-reactive protein (CRP), an acute-phase protein, is the most studied biomarker of systemic inflammation. Our results revealed higher CRP concentrations in patients supporting inflammation but no correlation between YKL-40 and CRP was detected. Similarly, other authors found increased YKL-40 but not CRP levels in patients with Alzheimer’s disease. where a different inflammatory profile in Alzheimer’s disease was detected [34].
There are no published data on gene or protein expression of YKL-40 in patients with CNS infections depending on the etiological agent. The fact that YKL-40 is increased in various neuroinflammatory and neurodegenerative conditions promotes its possible role in the pathophysiology of CNS diseases. Recently, a research group concluded that YKL-40 discriminates Alzheimer’s disease from controls and may predict the progression from the early preclinical to the late dementia stage. In genetically determined Alzheimer’s disease, YKL-40 was shown to increase decades before the clinical onset [35]. Contrarily, other authors reported no significant differences in YKL-40 levels in CSF in the cases of mild cognitive impairment in patients with Alzheimer’s disease compared with cognitively normal subjects [36]. We suggest that the decreased mRNA expression of YKL-40 and the higher protein levels may result from post-translational regulation of the YKL-40 gene. The results from the ROC curves of YKL-40 plasma levels illustrate the potential of plasma YKl-40 to serve as a discriminative marker. In contrast, the data about LAMPs in CNS infections are unconvincing as they indicate the biased and contradictory role of the glycoproteins. In previous studies, a possible regulatory axis lncRNAs/miR-30e/YKL-40 was shown in systemic sclerosis [37]. A possible regulatory mechanism could be also considered in CNS infections.
Study Limitations
A major limitation of the study is the small number of patients in the target group. Another drawback is the relatively nonspecific tissue expression of the examined glycoproteins. Although this is a clinical observational study with a small sample size, we believe that this investigation provides important evidence for the role of YKL-40 and LAMPs in CNS infections.
5. Conclusions
Based on our results it could be assumed that YKL-40 plasma levels might serve as a fast tool to discriminate viral or bacterial infection in the CNS. LAMPs could reflect the dysregulation of inflammation and autophagy during CNS infections. The downregulated pattern of gene expression in WBCs raises important questions from a biological point of view, which will need further examination.
Author Contributions
M.K. was responsible for the conceptualization and the original draft writing; Y.K. was involved in conceptualization, microbiological evaluation, and interpretation. P.A. was responsible for patient recruitment and selection. V.D. performed qPCR and ELISA experiments. M.K. and Y.K. contributed to funding resources. K.S. performed statistical analysis. V.S. and M.M. provided scientific supervision, study design, reviewing and editing the manuscript. All the authors contributed to the review and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by MU-Plovdiv under project No. 02/2022 (for ELISA kits) and European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project No. BG-RRP-2.004-0007-C01 for Strategic Research and Innovation Program for the Development of MU-Plovdiv (for qPCR and mPCR reagents).
Institutional Review Board Statement
All procedures were performed in accordance with the 1964 Helsinki Declaration and its later amendments. The study was approved by the University Ethics Committee (Protocol No. 1/28.02.2019).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are available upon a reasonable request by the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CNS | Central nervous system |
| CSF | Cerebrospinal fluid |
| WBCs | White blood cells |
| LAMPs | Lysosome-associated membrane proteins |
| ESR | Erythrocyte sedimentation rate |
| CRP | C-reactive protein |
| LP | Lumbar puncture |
| VN | Viral neuroinfections |
| BN | Bacterial neuroinfections |
| LOQ | Limit of quantification |
| HSV-1 | Herpes simplex virus type 1 |
| eIF2D | Translation initiation factor |
| LGI1 | Anti-leucine-rich glioma inactivated 1 |
References
- Melhuish, F. Viral meningitis and encephalitis. Medicine 2021, 49, 675–680. [Google Scholar] [CrossRef]
- Hasbun, R. Progress and Challenges in Bacterial Meningitis: A Review. JAMA 2022, 328, 2147–2154, Erratum in JAMA 2023, 329, 515. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, M.; Thwaites, G.E. The diagnosis and management of acute bacterial meningitis in resource-poor settings. Lancet Neurol. 2008, 7, 637–648. [Google Scholar] [CrossRef]
- Yekani, M.; Memar, M.Y. Immunologic biomarkers for bacterial meningitis. Clin. Chim. Acta 2023, 548, 117470. [Google Scholar] [CrossRef]
- Pedemonte, G.; Mancardi, D.; Giunti, A.; Corcione, F.; Benvenuto, V.; Pistoia, A. Mechanisms of the adaptive immune response inside the central nervous system during inflammatory and autoimmune diseases. Pharmacol. Ther. 2006, 111, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Kalita, J.; Haldar, R.; Misra, U.K. Blood-CSF-barrier permeability in tuberculous meningitis and its association with clinical, MRI and inflammatory cytokines. J. Neuroimmunol. 2022, 372, 577954. [Google Scholar] [CrossRef]
- Bailey, S.L.; Carpentier, P.A.; McMahon, E.J.; Begolka, W.S.; Miller, S.D. Innate and adaptive immune responses of the central nervous system. Crit. Rev. Immunol. 2006, 26, 149–188. [Google Scholar] [CrossRef]
- Thorsdottir, S.; Henriques-Normark, B.; Iovino, F. The Role of Microglia in Bacterial Meningitis: Inflammatory Response, Experimental Models and New Neuroprotective Therapeutic Strategies. Front. Microbiol. 2019, 10, 576. [Google Scholar] [CrossRef]
- Dorsett, M.; Liang, S.Y. Diagnosis and Treatment of Central Nervous System Infections in the Emergency Department. Emerg. Med. Clin. N. Am. 2016, 34, 917–942. [Google Scholar] [CrossRef]
- Kazakova, M.; Sarafian, V. YKL-40 in health and disease: A challenge for joint inflammation. Biomed. Rev. 2013, 24, 49–56. [Google Scholar] [CrossRef]
- Väänänen, T.; Koskinen, A.; Paukkeri, E.L.; Hämäläinen, M.; Moilanen, T.; Moilanen, E.; Vuolteenaho, K. YKL-40 as a novel factor associated with inflammation and catabolic mechanisms in osteoarthritic joints. Mediat. Inflamm. 2014, 1, 215140. [Google Scholar] [CrossRef]
- Çiledağ, A.; Akın, A.P.; Çelik, G.; Demir, N.; Yüksel, C.; Köycü, G.; Gökmen, Ö.D.; Rad, A.Y.; Kaya, A.; Kutlay, H.; et al. High serum YKL-40 level is associated with poor prognosis in patients with lung cancer. Tuberk Toraks 2018, 66, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Shao, R. YKL-40 acts as an angiogenic factor to promote tumor angiogenesis. Front. Physiol. 2013, 4, 122. [Google Scholar] [CrossRef]
- Bonneh-Barkay, D.; Wang, G.; Starkey, A.; Hamilton, R.L.; Wiley, C.A. In vivo CHI3L1 (YKL-40) expression in astrocytes in acute and chronic neurological diseases. J. Neuroinflamm. 2010, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Shahim, P.; Tegner, Y.; Marklund, N.; Höglund, K.; Portelius, E.; Brody, D.L.; Blennow, K.; Zetterberg, H. Astroglial activation and altered amyloid metabolism in human repetitive concussion. Neurology 2017, 88, 1400–1407. [Google Scholar] [CrossRef]
- Kazakova, M.; Pavlov, G.; Dikov, D.; Simitchiev, K.; Dichev, V.; Stefanov, C.; Sarafian, V. Protein YKL-40 in cerebrospinal fluid in traumatic brain injury. Acta Morphol. Anthropol. 2018, 25, 61–66. [Google Scholar]
- Eskelinen, E.L.; Illert, A.L.; Tanaka, Y.; Schwarzmann, G.; Blanz, J.; Von Figura, K.; Saftig, P. Role of LAMP-2 in lysosome biogenesis and autophagy. Mol. Biol. Cell 2002, 13, 2977–3368. [Google Scholar] [CrossRef]
- Eskelinen, E.L. Roles of LAMP-1 and LAMP-2 in lysosome biogenesis and autophagy. Mol. Asp. Med. 2006, 27, 495–502. [Google Scholar] [CrossRef]
- Jordi, V.; Jordi, B.; Carmen, M. Molecular diagnosis of the central nervous system (CNS) infections. In Enfermedades Infecciosas y Microbiología Clínica; David, N.O., Jose, P.M., Eds.; Elsevier: Madrid, Spain, 2021; pp. 403–410. [Google Scholar]
- Clemens, D.L.; Horwitz, M.A. Characterization of the Mycobacterium tuberculosis phagosome and evidence that phagosomal maturation is inhibited. J. Exp. Med. 1995, 181, 257–270. [Google Scholar] [CrossRef]
- Kiel, J.A. Autophagy in unicellular eukaryotes. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 819–830. [Google Scholar] [CrossRef]
- Pang, Y.; Wu, L.; Tang, C.; Wang, H.; Wei, Y. Autophagy-Inflammation Interplay During Infection: Balancing Pathogen Clearance and Host Inflammation. Front. Pharmacol. 2022, 13, 832750. [Google Scholar] [CrossRef] [PubMed]
- Orvedahl, A.; Alexander, D.; Tallóczy, Z.; Sun, Q.; Wei, Y.; Zhang, W.; Burns, D.; Leib, D.A.; Levine, B. HSV-1 ICP34.5 confers neurovirulence by targeting the Beclin 1 autophagy protein. Cell Host Microbe 2007, 1, 23–35. [Google Scholar] [CrossRef]
- Sarafian, V.; Jadot, M.; Foidart, J.M.; Letesson, J.J.; Van den Brûle, F.; Castronovo, V.; Wattiaux, R.; Coninck, S.W. Expression of Lamp-1 and Lamp-2 and their interactions with galectin-3 in human tumor cells. Int. J. Cancer 1998, 75, 105–111. [Google Scholar] [CrossRef]
- Sarafian, V.S.; Koev, I.; Mehterov, N.; Kazakova, M.; Dangalov, K. LAMP-1 gene is overexpressed in high grade glioma. Apmis 2018, 126, 657–662. [Google Scholar] [CrossRef]
- Kristen, H.; Sastre, I.; Aljama, S.; Fuentes, M.; Recuero, M.; Frank-García, A.; Martin, A.; Sanchez-Juan, P.; Lage, C.; Bullido, M.J.; et al. LAMP2 deficiency attenuates the neurodegeneration markers induced by HSV-1 infection. Neurochem. Int. 2021, 146, 105032. [Google Scholar] [CrossRef]
- Deretic, V. Autophagy in inflammation, infection, and immunometabolism. Immunity 2021, 54, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef]
- Kirkpatrick, R.B.; Emery, J.G.; Connor, J.R.; Dodds, R.; Lysko, P.G.; Rosenberg, M. Induction and expression of human cartilage glycoprotein 39 in rheumatoid inflammatory and peripheral blood monocyte-derived macrophages. Exp. Cell Res. 1997, 237, 46–54. [Google Scholar] [CrossRef]
- Muszyński, P.; Groblewska, M.; Kulczyńska-Przybik, A.; Kułakowska, A.; Mroczko, B. YKL-40 as a Potential Biomarker and a Possible Target in Therapeutic Strategies of Alzheimer’s Disease. Curr. Neuropharmacol. 2017, 15, 906–917. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Wang, Y.; Zhao, X.; Whang, S.; Li, L. CHI3L1 in the CSF is a potential biomarker for anti-leucine-rich glioma inactivated 1 encephalitis. Front. Immunol. 2022, 13, 1071219. [Google Scholar] [CrossRef]
- Hermansson, L.; Yilmaz, A.; Axelsson, M.; Blennow, K.; Fuchs, D.; Hagberg, L.; Lycke, J.; Zetterberg, H.; Gisslén, M. Cerebrospinal fluid levels of glial marker YKL-40 strongly associated with axonal injury in HIV infection. J. Neuroinflamm. 2019, 16, 1–9. [Google Scholar] [CrossRef]
- Létuvé, S.; Kozhich, A.; Arouche, N.; Grandsaigne, M.; Reed, J.; Dombret, M.C.; Kiener, P.A.; Aubier, M.; Coyle, A.J.; Pretolani, M. YKL-40 is elevated in patients with chronic obstructive pulmonary disease and activates alveolar macrophages. J. Immunol. 2008, 181, 5167–5173. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Palmero, V.A.; Rubio-Fernández, M.; Antequera, D.; Villarejo-Galende, A.; Molina, J.A.; Ferrer, I.; Bartolome, F.; Carro, E. Increased YKL-40 but Not C-Reactive Protein Levels in Patients with Alzheimer’s Disease. Biomedicines 2021, 9, 1094. [Google Scholar] [CrossRef]
- Gispert, J.D.; Monté, G.C.; Falcon, C.; Tucholka, A.; Rojas, S.; Sánchez-Valle, R.; Antonell, A.; Lladó, A.; Rami, L.; Molinuevo, J.L. CSF YKL-40 and pTau181 are related to different cerebral morphometric patterns in early AD. Neurobiol. Aging 2016, 38, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ng, K.P.; Therriault, J.; Kang, M.S.; Pascoal, T.A.; Rosa-Neto, P.; Gauthier, S. Alzheimer’s Disease Neuroimaging Initiative. Cerebrospinal fluid phosphorylated tau, visinin-like protein-1, and chitinase-3-like protein 1 in mild cognitive impairment and Alzheimer’s disease. Transl. Neurodegener. 2018, 7, 23. [Google Scholar] [CrossRef]
- Dichev, V.; Mehterov, N.; Kazakova, M.; Karalilova, R.; Batalov, A.; Sarafian, V. The lncRNAs/miR-30e/CHI3L1 Axis Is Dysregulated in Systemic Sclerosis. Biomedicines 2022, 10, 496. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).