Abstract
The blood–brain barrier acts as a physical barrier that prevents free entry of blood-derived substances, including those intended for therapeutic applications. The development of molecular Trojan horses is a promising drug targeting technology that allows for non-invasive delivery of therapeutics into the brain. This concept relies on the application of natural or genetically engineered proteins or small peptides, capable of specifically ferrying a drug-payload that is either directly coupled or encapsulated in an appropriate nanocarrier, across the blood–brain barrier via receptor-mediated transcytosis. Specifically, in this process the nanocarrier–drug system (“Trojan horse complex”) is transported transcellularly across the brain endothelium, from the blood to the brain interface, essentially trailed by a native receptor. Naturally, only certain properties would favor a receptor to serve as a transporter for nanocarriers, coated with appropriate ligands. Here we briefly discuss brain microvascular endothelial receptors that have been explored until now, highlighting molecular features that govern the efficiency of nanocarrier-mediated drug delivery into the brain.
1. Introduction
Effective delivery of drugs across the blood–brain barrier (BBB) is a major limiting factor in successful therapy of brain diseases. The BBB is constituted by the specialized endothelium of the brain microvessels that preserves brain homeostasis by restricting the influx of a variety of compounds, including biomedicine, and yet enabling the supply of nutrients. Indeed, the potential treatment of many brain-related diseases is severely hampered because of the fact that numerous potential drug candidates are recognized as “brain-hostile”, thus precluding their penetration across the BBB into the brain. To overcome this problem, much research efforts are currently focused on the development and application of safe and efficient delivery devices, capable of promoting drug transport across the BBB. These efforts include the ability to achieve specific delivery by means of targeting to the BBB endothelium, followed by effective transport of the delivery device across the endothelial cell barrier and subsequent release of the drug at appropriate sites within the brain.
Here we will first briefly discuss the anatomical and molecular nature of the BBB. Obviously, such knowledge will be imperative for appreciating the need for development of state-of-the-art molecular devices to enable crossing of the barrier. We will subsequently discuss the presence of distinct receptors on the cell surface of vascular endothelial cells, capable of binding specific ligands and whose mechanism of internalization can be exploited for achieving targeted delivery and passage of a variety of nanodevices across the BBB.
2. Blood–Brain Barrier—Morphology and Function
2.1. The BBB as a Physical Barrier
Anatomically, the organization of the BBB is comprised of endothelial cells, basal lamina, astrocytes’ end-feet and pericytes. Endothelial cells of the BBB, in contrast to the systemic endothelium, are tightly connected to each other by means of tight junctions (TJ) and adherence junctions, and consequently lack fenestrations. Among others, proteins of the claudin family, i.e., claudin 3, 5 and 12, occludin, and junction-adhesion molecules (JAMs), which are expressed at opposing cell surfaces, form the tight junctions by means of homophilic interactions. In this manner TJ strands “zip” adjacent endothelial cells and close the paracellular space for access of blood-borne molecules. The structure is further stabilized by ZO-1, -2 and -3, and MAGUK family proteins that link the TJ with the actin cytoskeleton. Signaling and regulatory proteins include multi-PDZ-protein 1 (MUPP1), the partitioning defective proteins 3 and 6 (PAR3/6), MAGI-1,-2,-3 (membrane-associated guanylate kinase inverted 1–3), ZO-1-associated nucleic acid-binding protein (ZONAB), afadin (AF6), and Regulator of G-protein signaling 5 (RGS5) [1]. Importantly, the presence of the tight junctions divides the plasma membrane of the vascular endothelial cells into two separate domains, that is the apical membrane, which faces the blood, and the basolateral membrane, which faces the brain tissue. The tight junctions preclude randomization of the surface components localized at each of these domains, thus warranting a specific composition of each domain. Basolaterally from the TJ, PECAM and VE-cadherin form the adherence junctions, which are linked to the cytoskeleton via desmoplakin and p120 catenin [1,2,3]. The structure is further supported by the basal lamina, a 30–40 nm thick matrix, composed of collagen type IV, heparin sulfate proteoglycans, fibonectin, laminin and other extracellular matrix proteins [4].
Multiple basal lamina proteins, matrix metalloproteases (MMPs) and their inhibitors, i.e., the tissue inhibitor of metalloproteases (TIMPs), are involved in regulating the dynamics of the integrity of the BBB at physiological and inflammatory conditions [5]. In addition, molecular factors released from glial cells, such as glial-derived neurotrophic factor (GDNF), angiopoietin-1 [6] and angiotensin II [7] also contribute to the robustness of the integrity of the BBB. Furthermore pericytes regulate the developing BBB [8] and control the vascular permeability by restricting transcytosis, a transcellular transport process (see below) through PDGFR-β signaling [9] and regulation of Mfsd2a expression at the BBB [10]. Lastly, stability of the BBB is also provided by interactions with astrocytes, which cover more than 99% of the basal capillary membrane.
2.2. The BBB as a Functional Barrier
Apart from acting as a morphological barrier as such, the endothelial cell surface also contains molecular entities that further effectuate its barrier function. Thus, transporters belonging to the family of the ATP-binding cassette (ABC) transporters are expressed on the apical membrane surface of the BBB and their activity precludes entry and/or facilitates extrusion of many xenobiotics and endogenous compounds. In this manner, the functional activity of ABC transporters contributes to the barrier function of the vascular endothelium.
In an ATP-dependent manner the ABC-transporters actively extrude undesired compounds into the extracellular milieu. P-gp, MRP1 and BCRP are the most relevant ABC transporters at the BBB, while P-gp, consisting of two six transmembrane spanning domains and two ATP-binding sites, is the most active one. Common substrates for P-gp are cationic compounds or compounds with hydrogen bond acceptor groups [11], but also peptides [12]. Other efflux pumps expressed at the apical surface of the BBB endothelium are MRP4 and MRP5, whereas MRP4 is also present on the basolateral membrane.
Given the structural and functional tightness of the barrier, it is apparent that for purposes of drug delivery into the brain, development of sophisticated devices are needed, capable of passing the vascular endothelial cell lining. Potential mechanisms to be exploited are those based on paracellular entry (“between cells”) or via transcytosis (“through cells”). To appreciate the potency of these events we will therefore briefly describe the nature of such transport processes, occurring at (patho) physiological conditions.
3. Passive and Active Mechanisms of Transport across the BBB
Although preventing random influx of molecular compounds into the brain, the BBB does allow selective entry of nutrients and supply of regulatory factors into the brain. Thus the endothelial cells enable transport of molecules in a passive, gradient-driven manner or active, energy-dependent mode.
At physiological conditions, passive movement of water solutes along the aqueous route of the intercellular cleft between endothelial cells via small pores in the tight junctions, i.e., the paracellular pathway, is negligible (Figure 1a). Since the exact molecular nature of paracellular transport is still largely obscure, and neurodegenerative and neuro-oncological diseases at the early stages of development are not necessarily accompanied by defects in BBB integrity, general exploitation of this pathway for transport of drug-loaded nanoparticles into the brain might be virtually negligible. As an alternative, transport via the endothelial cells per se, i.e., by means of transcellular transport or transcytosis should thus be considered. Indeed, transport of small molecules through cells is a common event in polarized cells, including vascular endothelial cells (Figure 1b). Thus hydrophobic molecules, like many psychoactive drugs, with a size less than 500 Da, may diffuse transcellularly-provided they escape from MDR-mediated extrusion-to reach the brain parenchyma from the systemic circulation. However, transport of nutrients and building blocks is more tightly regulated and facilitated by specialized transporters (Figure 1c). For example, glucose uses the glucose transporter (GLUT1), while amino acids are transported by the large neutral amino acid transporter (LAT1). The latter transporter is also involved in transport of drugs like l-dopa, gabapentin, and baclofen [13]. For a detailed review of carrier-mediated transport at the BBB, see [14].
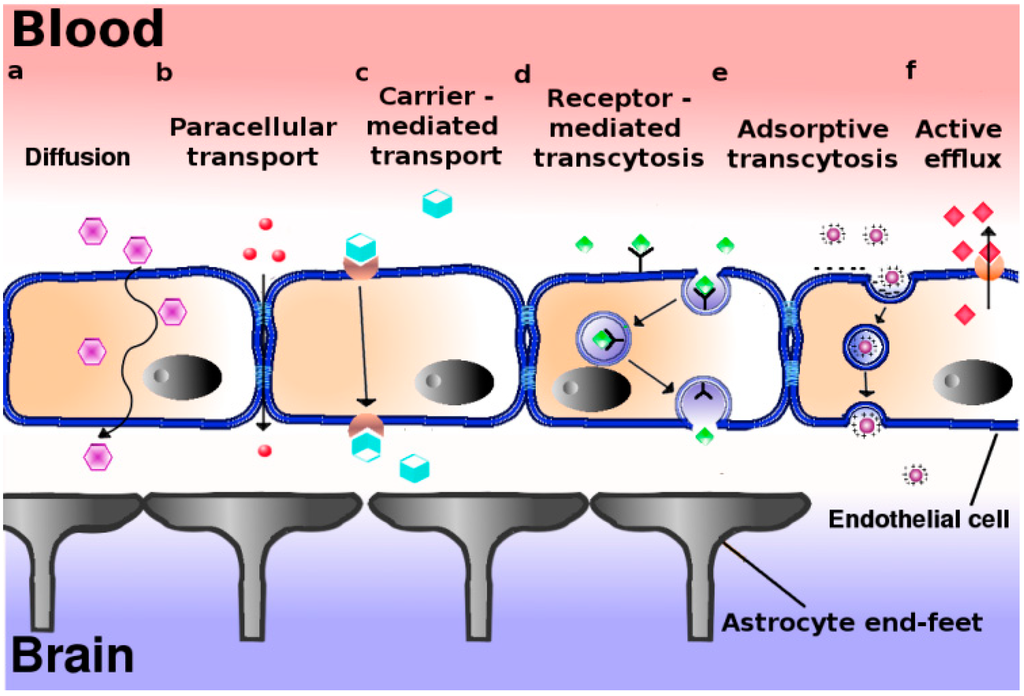
Figure 1.
Proposed mechanisms of transport across the blood–brain barrier. The blood brain barrier consists in essence of a polarized layer of vascular endothelial cells, tightly attached to each other by means of tight junctions, and lined up by astrocytes. A variety of transcellular transport processes can be distinguished: (a) Diffusion, driven by a concentration gradient, mainly involving small hydrophobic molecules. This pathway represents the main entry route into the brain of current therapeutics; (b) Paracellular transport–limited to small water-soluble molecules; (c) Carrier-mediated transport, as occurs for e.g., glucose, amino acids, nucleosides, and therapeutics such as vinca alkaloids, azidothymidine etc.; (d) Receptor-mediated transcytosis for peptidic signaling and regulatory molecules (insulin, leptin, interleukins), nutrients (iron, LDL); (e) Adsorptive transcytosis, presumably relying on transport of positively charged cargo (serum proteins) in a non-specific manner; (f) Proton pump efflux transporters.
Whether these transporters, being primarily involved in molecular translocation of relatively small molecules, might similarly translocate larger sized nanoparticles into and out of the cell, appears less likely. Arguing that the brain depends on an extremely high glucose demand, i.e., nearly 30% of the total body glucose consumption, liposomes prepared with glucose-modified cholesterol have been applied for brain-directed transport. These studies have shown a mild advantage in transporting coumarin-6 into the brain, over conventional liposomes. Although attempts have been undertaken to tie the GLUT1 transport system to transport of glucose-functionalized nanoparticles [15,16] the involvement of GLUT1 was not confirmed, or completely excluded in favor of passive cytoplasmic transfer [17]. Glucose-modified paclitaxel-loaded polymeric nanoparticles were successfully employed in the treatment of intracranial glioma in mice. Here, a role for GLUT1 in the transport of the nanoparticles across the BBB was confirmed [18].
Yet, a focus on receptors that internalize ligands by means of vesicular transport appears a more appropriate option since such intracellular transport vesicles can readily accommodate nanoparticles with a size of 100–200 nm. Indeed, transcellular transport in this manner (“transcytosis”) is a common phenomenon in polarized cells (Figure 1d,e).
Transcytosis represents a means of vectorial transport of cargo (including nanoparticles) between the apical and basolateral surface in polarized cells. It operates bidirectionally and three distinct steps in overall transcytotic transport can be distinguished: endocytosis of the cargo/nanoparticle at the plasma membrane (Box 1), followed by intracellular vesicular trafficking towards the opposite surface where exocytosis of the vesicular contents takes place. Transcytosis can rely on receptor-mediated and/or adsorptive, charge-dependent endocytic internalization events (Figure 1d,e). The adsorptive mode has been described to occur in particular for cationized serum albumin. Receptor-mediated transcytosis in brain endothelium may involve ligands like transferrin (iron), deltorphin, enkephalin, insulin, and LDL [5].
Here we will further discuss possibilities for specific targeting of nanoparticles into the brain relying on their specific processing along a transcytotic pathway rather than entry into an endocytosis-mediated degradation pathway (cf. Box 1).
Box 1. Entry of molecules and particles into cells: Modes of internalization.
- Internalization via the endocytic mechanism is subdivided into clathrin-dependent, caveolin-dependent and clathrin- and caveolin-independent pathways (Figure 2). Clearly, the mode of cellular cargo entry is crucial for its eventual fate, as will be discussed below.
- Clathrin-mediated endocytosis involves the assembly of a clathrin coat at membrane regions enriched in receptor-ligand complexes, just underneath the plasma membrane, which subsequently triggers its inward budding. The small GTPase dynamin closes the neck of the invaginating membrane surface, leading to the formation of the clathrin-coated vesicle, which subsequently pinches off [19]. Shortly thereafter clathrin molecules dissociate from the vesicles, which then merge homotypically and/or with preexisting compartments that become enriched in EEA-1, a typical marker of early sorting endosomes [20]. From this compartment recycling may occur either directly to the plasma membrane, the so-called rapid recycling route [21,22] or via a recycling endosome, the slow recycling route [21]. Alternatively, early endosomes deliver their cargo to late endosomes, either via maturation [23] or by means of a vesicular transport mechanism [24] and eventually to lysosomes where its degradation may take place. Along the transition from early to late endosomes, the compartmental pH gradually drops [25].
- A better known but still poorly defined pathway capable of transferring cargo across the endothelial cell lining, constituting the blood-brain barrier, appears to rely on entry via caveolae [26,27]. As an advantage, lysosomal delivery as often occurs for entry along the clathrin-mediated pathway, may be avoided thus promoting secretory transport, rather than capture of cargo in a digestive compartment [28]. Accordingly, targeting specific receptors associated with caveolae may therefore well represent a far more appropriate strategy in facilitating transcellular transport, including that of targeted nanodevices. Caveolae are plasma membrane invaginations, commonly described as structures with a flask-like shape [29]. They are predominantly present at the surface of adipocytes [30], lung epithelium [31] and vascular endothelium [32,33]. A major constituent of caveolae is caveolin-1, which localizes at the inner leaflet of the plasma membrane [34]. Supported by the cavins-1, -2, -3 and -4, caveolae sustain their characteristic morphology, but gradually flatten [35]. Caveolae are considered platforms, where trafficking and signaling events take place [36,37,38].
- Stimulation of caveolar receptors triggers their internalization involving formation of a vesicular structure, named cavicle [39]. Cargo taken up by caveolae might be delivered to the Golgi apparatus or the endoplasmic reticulum, as reported for the cholera and shiga toxin B subunits in epithelium [40].
- A caveolar transcytotic route has been proposed to operate in endothelial cells of the BBB. Specifically, this pathway has been described for LDL particles which, prior to their release at the basolateral surface (brain side), localize at multivesicular bodies [41].
- The molecular organization of caveolae is strongly dependent on the presence of cholesterol, and agents that perturb the cholesterol content and organization, such as cyclodextrin and filipin, frustrate caveolae-mediated internalization [42,43]. The kinase inhibitor genistein is also known for its capacity to block caveolar uptake [44].
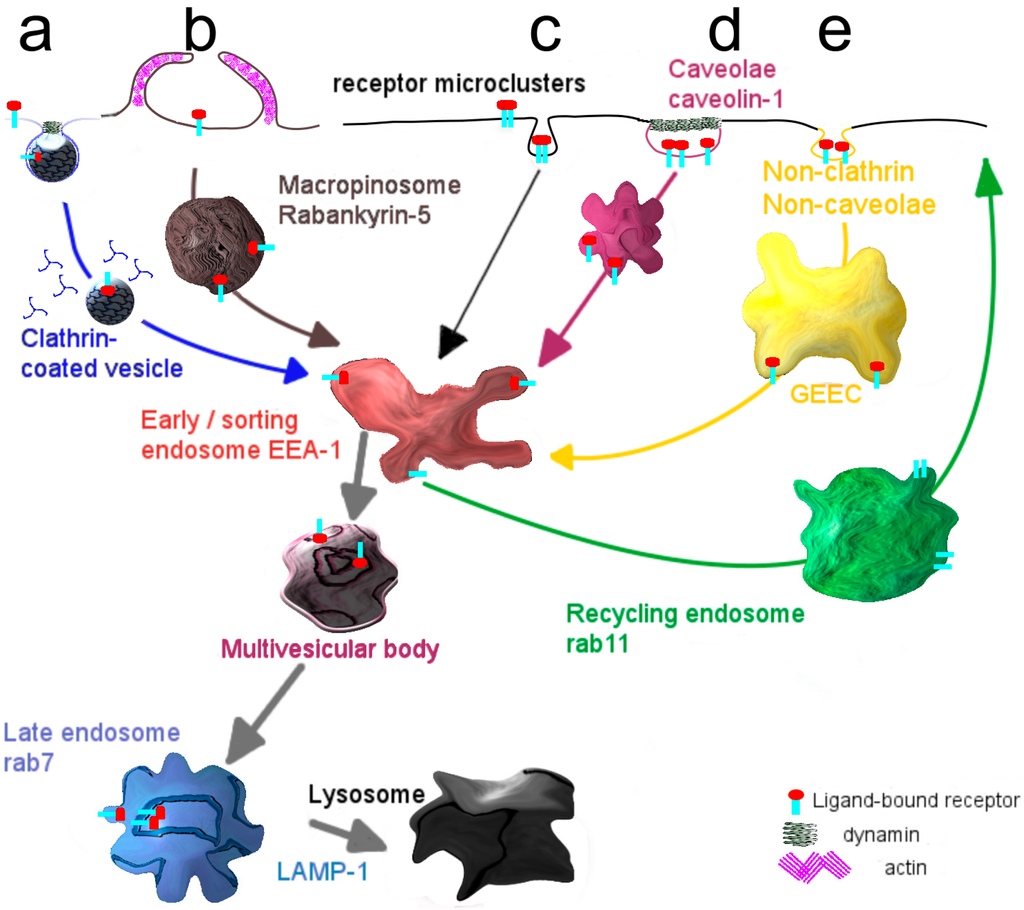
Figure 2.
Mechanisms and transport pathways in endocytosis. (a) Adaptor molecules sense the binding of a ligand to its cognate receptor and cause the formation of a clathrin cage around the plasma membrane that subsequently pinches off as internal vesicle; (b) Caveolin-1, constitutively present at the inner leaflet of the plasma membrane, forms invaginated microdomains, known as caveolae, that mediate transport of receptor-ligand complexes; (c) A non-clathrin, non-caveolae dependent mechanism is mainly responsible for the uptake of glycosylphosphatidylinositol anchored proteins.
4. Nanocarriers and Active Targeting Systems (Ligands)
As depicted in Box 1, for the purpose of nanocarrier-mediated drug delivery into the brain, transcytosis from the blood (luminal side) into the tissue (abluminal side) seems to be the preferred pathway of crossing the BBB, as imposed by the vascular endothelium. Since caveolae have been proposed to be particularly active as transcytotic portal of entry [45], such a strategy would thus require caveolar targeting of the nanocarriers, implying coupling of appropriate ligands to these devices. The concept of coupling ligands directly to the therapeutic agent or to the surface of therapeutics-containing nanocarriers thus aims at (i) tissue specific delivery and (ii) improved penetration via transcytosis. As suggested by Pardridge [46], the Trojan horse constructed in this manner should contain a ligand that meets the necessary characteristics in order to serve as an effective tool in brain targeting. When coupled to the Trojan horse, the ligand should retain its targeting capacity towards a transcytotic receptor, as simple endocytosis might result in trafficking towards lysosomes and hence degradation. Thus, the targeted Trojan horse should bind with high affinity to the correct BBB receptor, and this affinity should be retained after fusion/conjugation with the therapeutic compound. Furthermore an effective internalization of the nanodevice is essential for achieving efficient brain uptake of the complex in vivo (>2% ID/gram brain tissue is required). Such uptake should of course result in a pharmacological effect, an effect that also should be warranted after intravenous administration of the complex. In the following we will briefly discuss a variety of ligands that have been used in optimizing brain delivery. Several of these have acquired a vector status of their own, involving their direct linkage to the drug rather than linkage to a drug-containing device. However, the use of nanocarriers for drug delivery to the brain may be preferred to prevent degradation of the drug during circulation and increase its concentration at the target site, the latter being of critical importance for reaching a therapeutic effect.
4.1. Transferrin
The best-known example of receptor-mediated endocytosis, operating in endothelial cells constituting the BBB, is the transport of iron-loaded transferrin (Tf). The holo-transferrin (iron-bound form) interacts with the transferrin receptor (TfR) at the (blood-facing) apical cell surface and releases its iron in early endosomes, triggered by the (mild acid) pH drop. The apoTf–receptor complex is subsequently recycled to the apical plasma membrane through rab11-positive compartments. The relative abundance of the TfR on the BBB endothelial cells, rationalizes the use of Tf–TfR system as a brain targeting system in a number of studies. Tf-conjugated solid lipid nanoparticles (SLNs) clearly increase the bioavailability of quinine dihydrochloride in total brain (parenchyma and microvessels) of rats compared to the administration of the free drug [47]. However, the particles remain associated with the microvessels, and the effect is therefore unlikely due to the transport of the drug–SLN complex into the brain. Moreover, a high plasma concentration of endogenous Tf balances the saturation of the receptor and discredits the use of Tf as an efficient targeting vector. Alternatively antibodies, with affinity for different epitopes on the TfR were therefore generated and investigated.
The radiolabeled anti-transferrin receptor monoclonal antibody OX26 was recovered in the brain of rats after intravenous injection, suggesting transcytosis across a functional BBB [48]. This observation subsequently accounted for the use of OX26 in many studies as a transport vector. Indeed, antisense oligonucleotides (ODNs) and peptide nucleic acids (PNAs) emerge as therapeutics. When PNA was conjugated to OX26, through biotin-streptavidin coupling, brain uptake, which is negligible for unconjugated drug, equals 0.08 ID%/g (% injected dose per gram tissue) after carotid artery perfusion. According to the authors this dose might be sufficient to elicit an effect with therapeutic significance, alluding to the brain recovery dose of injected morphine (0.1 ID%/g) [49]. Likewise, the mAb OX26 promotes the brain uptake of functionalized polymersomes (PO), suggesting retention of targeting properties when coupled to a nanoparticle [50]. The number of OX26 molecules exposed on the surface of a PO (100 nm) was optimal at 34 per particle, which yielded a brain recovery value of 0.136 ID%/g after i.v. injection in rats. Further increase in the OX26 concentration failed to produce higher brain uptake, possibly due to an increased liver accumulation.
As noted above, an important criterion to be met for effective targeting is the ability to accomplish a pharmacological effect, following delivery of the drug-containing device. Interestingly, such an effect was indeed obtained for the OX26-targeted polymersomes, loaded with the NC-1900 peptide, known to ameliorate spatial memory deficits. Thus, in scopolamine provoked rats, subcutaneous injection of the peptide per se and intravenous injection of its OX26 polymersomal formulation both improved the performance of the animals in water-maze tests. However, when encapsulated in OX26-conjugated polymersomes, a dose of NC-1900 lower than the free drug sufficed to produce a comparable effect, suggesting a more efficient brain delivery by means of polymersome-mediated transport [50]. Similarly, the effect of loperamide to produce analgesia was demonstrated. The drug is BBB impermeable and hence analgesic activity is measurable only after effective brain penetration. Tf-, OX26-, and RI7217 (mouse equivalent of OX26)-targeted albumin nanoparticles (160–185 nm) produced significant pain relief compared to mock-targeted (IgG2a) nanoparticles [51]. However, these antibodies lack the capacity to mediate actual crossing of the endothelial cell layer, as was determined in vivo using capillary depletion and morphological examination, resulting in capture of the antibodies and antibody–drug/nanocarrier conjugates within the brain vasculature [52,53,54]. A biphasic behavior of antibody–drug/nanocarrier conjugates with firstly, actual accumulation of the complex in endothelial blood capillaries, that has been documented, and secondly a passive release of the drug from the basal site might explain their efficacy in producing a pharmacological effect. Then, a low-affinity antibody would serve an improved delivery, as it will allow for easier release of the drug from the complex, once inside the cell [55]. In addition, carrier systems that will degrade within the brain endothelial cells may improve the subsequent transport of cargo further into the brain. It has also been postulated that novel ligands that prevent the dimerization of Tf receptors at the cell surface, thereby influencing the mode of internalization and subsequent intracellular trafficking, promote actual transcytosis [56]. Overall, these data support the concept that targeted brain delivery of nanodevices appears a realistic goal, and that the application of using an antibody against the transferrin receptor as brain vector, could serve that purpose.
4.2. Melanotransferrin
Melanotransferrin (p97) is a GPI-anchored protein that is highly expressed in melanomas. The protein, similar to Tf, is an iron-binding protein [57], and compared to Tf shows a 14-fold enrichment in the brain following injection of the recombinant form. Rather than the Tf-receptor, evidence supports a role of the low-density lipoprotein-receptor related protein (LRP) in the transport of p97 across the endothelium [58]. The targeting properties of p97 were demonstrated when covalently linked to paclitaxel or adriamycin. The total accumulation of the p97-drug complex in the brain reached a 10-fold higher level than that of the free drug. Furthermore, when linked to p97, the pharmacological activity of adriamycin was preserved and, when compared to non-targeted adriamycin, significantly decreased the progression of intracranial gliomas and mammary tumors [59]. After 24 h, p97-conjugates outperformed other, similarly designed vector-drug conjugates [49] and reached delivery levels of 1%–2% of the injected dose, which is equivalent to the ratio of brain to body weight. Hence, the p97 was thus claimed as the first carrier system to approximate this biological feature. Thus, melanotransferrin is preferentially transported from blood into brain tissue across the BBB, i.e., in an apical to basolateral direction with a kinetic profile that reflects a low-affinity binding to a receptor with a high transport capacity. Moreover, in contrast to the plasma concentration of transferrin, the plasma concentration of endogenous p97 is relatively low, thus assuring ready access of melanotransferrin to available binding sites. Accordingly, relative to the TfR system, the melanotransferrin-receptor system emerges as a preferred targeting vector for drug transport into the brain.
4.3. Insulin
Insulin is transported to brain tissue from the circulation by means of a transcytotic mechanism, involving the insulin receptor present at the vascular endothelial cell surface [60]. The human anti-insulin receptor monoclonal antibody (HIRMAb) was shown to shuttle a TNFα decoy receptor (TNFR), which would counteract the effects of TNFα in inflamed brain regions, into the brain of Rhesus monkeys following i.v. administration. Although the detected values remain low, the concentration of the HIRMAb-TNFR fusion protein in the brain parenchyma was calculated as sufficient to neutralize the effect of TNFα in inflammatory conditions [61]. In the same study the inability of the fusion protein between TNFR and an antibody against the microvascular FcRn receptor (involved in the transcytosis of IgG) to cross the BBB, was explained by the predominant efflux of the FcRn receptor from the brain to the blood, i.e., by its reverse transcytosis [62]. Although the HIRMAb was engineered to bind with high affinity to a different receptor epitope than the endogenous insulin, sequestration of the HIR from the plasma membrane by means of HIRMAB-drug conjugates would make it unavailable for its endogenous ligand–insulin, and vice versa. The competition between endogenous ligands and HIRMAb-modified drug conjugates for the same receptor may result in lack in efficacy of the nanoformulation and/or changes in receptor activity, of which the latter may have negative consequences for glucose metabolism [63].
4.4. LDL
The binding of low-density lipoproteins to the endothelial LDL-receptor (LDLR; [27]) or scavenger receptor-BI (SR-BI) initiates their transcytosis across the BBB. Remarkably, two endocytic pathways may be involved in the processing of the LDLR, which appears to depend on cell development. Thus, in non-differentiated, proliferating endothelial cells the LDLR is processed along the classical clathrin-mediated pathway (Box 1). LDL particles, when bound to the LDLR are taken up and delivered to late endosomes/lysosomes to meet the cell’s nutritional needs in terms of cholesterol supply. In this process, the receptor per se recycles back to the cell surface after dissociation of the ligand in early endosomes. However, in fully differentiated endothelial cells, constituting the BBB, the LDLR switches from a recycling to a transcytotic pathway. In this case, the receptor is captured by lateral diffusion in caveolae which, following their internalization, deliver intact LDL particles from the blood into brain tissue via intermediate multivesicular bodies (MVBs) (Box 1). Collectively, these data thus rationalize the use of LDLR as an appropriate transport system for drug delivery into the brain.
The first evidence to support this hypothesis comes from a study of Alyautdin et al. [64], which reported an analgesic effect after intravenous injection of dalargin (a hexapeptide analog of Leu-enkephalin, containing d-Ala in the second position and an additional C-terminal arginine), absorbed onto polysorbate 80-coated poly(butyl cyanoacrylate) (PBCA) nanoparticles. A series of follow-up studies [65,66,67] subsequently supported the enhanced drug delivery, as accomplished by using polysorbate 80-coated PBCA nanoparticles. Initially, the translocating effect across the endothelial cells was attributed to a general toxic effect of polysorbate 80 on the cells by causing a disruption of the tight junctions [68]. However, more recent data [69,70], in conjunction with the observation that serum apolipoproteins could adsorb on the surface of surfactant-coated nanoparticles [71] thereby promoting PBCA nanoparticles-endothelial cell interactions, support the currently prevailing hypothesis of receptor-mediated transcytotic transport of PBCA nanoparticles across BBB endothelial cells. Thus after intravenous injection, polysorbate 80 facilitates adsorption of the LDLR ligands, serum ApoB and/or ApoE onto the nanoparticles. As a consequence, the nanoparticles are targeted towards the LDL receptor on the brain capillary endothelial cells and are internalized, mimicking transcytosis of natural lipoproteins. Obviously, these observations subsequently raised an interest in surfactant-modified nanoformulations for brain delivery. Indeed, a nanosuspension of Amphotericin B prepared with Tween-80 (trade name of polysorbate 80) showed an enhanced adsorption of ApoE and ApoJ, and caused a moderate survival effect in Balamuthia mandrillaris encephalitis [72]. PBCA nanoparticles (35–40 nm) also proved to promote the CNS bioavailability of rivastigmine and tacrine as applied in the treatment of Alzheimer disease [73]. However, the PBCA nanoparticles also show several disadvantages, including a limited drug loading capacity, a low rate of in vivo biodegradation, and the release of toxic formaldehyde residues. Accordingly, physiologically better tolerable solid lipid nanoparticles (SLNs) have been suggested as suitable alternatives, relying on polysorbate 80 SLN preparations for further in vitro/in vivo studies [74]. Although polysorbate 80 is generally recognized as a safe emulsifier and included in many pharmaceuticals, care should be taken when a formulation is intended to be used for long-term parenteral application. For example, when treatment of a given disease will require repeated drug administration over periods of months, rather than a single shot, a higher toxicological burden caused by surfactant-nanoformulations might be a limiting factor, particularly in the treatment of chronic diseases.
Nevertheless, the concept of targeting the LDL-receptor to piggyback PBCA nanoparticles across the BBB, proved successful. Interestingly, essential for the targeting properties is not the surfactant itself, but rather, the apolipoproteins it attracts from the serum. Therefore, a variety of other apolipoprotein species were tested in order to verify their potency in targeting LDLR. Thus it has been shown that ApoE, covalently attached to human serum albumin nanoparticles (ApoE–HSA, size of approx. 250 nm), causes a relatively fast uptake of such particles (within 15–30 min) by brain capillary endothelial cells in vivo [75], and their processing by transcytosis was inferred from ultrastructural analyses. As a control, the authors demonstrated that pegylated nanoparticles (size of approx. 210 nm) do not associate with BBB endothelium, and consequently were not observed in brain parenchyma. Pegylation is a modification, usually carried out to prolong the circulation half-life of drugs and/or nanoparticles [76]. In this particular case, the control experiment may not have been an appropriate control given the rapid uptake of the non-pegylated particles, and hence questions the time span of the control experiment. Nevertheless, ApoE–HSA nanoparticles are transcellularly transported across the BBB and reach the brain intact, enabling brain delivery of drugs that usually cannot cross the BBB. The intracellular trafficking of ApoE–HSA was studied in an in vitro BBB model, i.e., bEnd3 cells. After a 2 h incubation many ApoE–HSA particles localize in intracellular compartments that were similar in appearance to lysosomes. Considering some recent advances in the mechanism of LDL transcytosis [41], revealing LAMP-positive multivesicular bodies as an intermediate step in the process of transcytosis of LDL, the observed localization of the ApoE-HSA in lysosome-like compartments could thus reflect a very similar transcytotic trafficking route.
Comparable to Apo-E, other apolipoproteins, e.g., ApoA-I and ApoB-100, may also act as vectors for transport of loperamide-HSA nanoparticles into the brain [77]. ApoA-I is mainly associated with HDL particles, mediating internalization via the SR-BI, suggesting that SR-BI may function as an entry promoting receptor for ApoA-I–HSA particles. When scored by a fast antinociceptive response (15 min after injection), ApoE–HSA particles produce the highest effect, followed by ApoA-I–HSA particles, loaded with the same concentration of loperamide. Consistently, ApoA-I-HSA particles have been visualized in different brain regions [78].
The expression level of LDLR on the luminal surface of the brain capillary ECs is synchronized with the demand for brain lipid requirement, and strictly controlled by the underlying astrocytes [79]. Thus when their cholesterol level is limited, astrocytes will transmit signals that lead to an up-regulation of luminal endothelial LDLR. A limited expression of LDLR will consequently result in a saturable transport capacity, thereby potentially limiting its use in brain delivery of cargo-containing nanoparticles. This implicates that drug delivery efficiency will likely be influenced by body rhythms, such as eating rhythm. Moreover, similarly to the transferrin receptor, LDLR is also expressed in various other tissues with the highest functionality in hepatocytes, implying a lack of brain capillary endothelium specificity.
4.5. Angiopeps
Low-density lipoprotein receptor-related protein (LRP) 1 (alpha-2-macroglobulin receptor) and 2 (Megalin) are other members of the LDL receptor family that are widely studied for their ability to transport multiple ligands across the BBB. Well-characterized are the LRP-dependent endothelial transport of lactoferrin [80], receptor-associated protein (RAP) [81], tissue plasminogen activator (tPA) [82], and secreted amyloid precursor protein (APP) [83,84]. Particularly in the latter case, when the Kunitz protease inhibitor (KPI) fragment was sequestered from the secreted APP, the LRP-dependent uptake and consequent degradation of APP was reduced [84]. Additionally, the transcytosis rate of 125I-aprotinin, another example of a KPI domain containing protein, across endothelium in an in vitro BBB coculture model or in situ brain perfusion, was higher when compared to holo-transferrin, used here as positive control. These observations were taken to suggest the involvement of the Kunitz domain in ligand recognition and subsequent LRP-mediated endocytosis. Accordingly, these findings rationalized the development of designing KPI-based peptides as vectors for brain delivery, operating through the LRP transport system [85]. Alignment of the amino acid sequence of aprotinin with that of bikunin, amyloid β A4 protein precursor, and the Kunitz inhibitor-1 precursor (all LRP ligands) led to the design of 96 peptides, commonly referred to as angiopeps. After in vitro and in vivo postselection, a peptide, designated as Angiopep-2 showed the highest transcytotic potential, and was used for further studies. The hypothetical role of LRP1 as a potential receptor for Angiopep-2 was examined by means of competitive inhibition experiments and colocalization studies, as carried out by fluorescence microscopy. Apical to basal transport of α2-macroglobulin (LRP1 specific ligand) across bovine brain capillary endothelial cells (BBCEC) was partially inhibited in the presence of Angiopep-2, and vice versa [86]. Furthermore, fluorescently-labeled Angiopep-2 colocalized in BBCEC to a certain extent with LRP1 in intracellular vesicles, pointing towards at least a partial LRP1-mediated transport of Angiopep-2. The potency of Angiopep-2 in brain delivery became evident when used as a conjugate, consisting of three molecules of paclitaxel, covalently attached to one Angiopep-2, also known as ANG1005. Paclitaxel preserves its cytotoxic effect in this form, and after intracerebral implantation of primary or metastatic carcinomas, administration by intraperitoneal injection of ANG1005 increases the median survival rate and prolongs the life span of mice, compared to vehicle, i.e., Angiopep-2 without paclitaxel treated animals [87]. Currently, ANG1005 is undergoing phase I/II clinical studies [88,89].
However, the potency of Angiopep-2 as a brain delivery vector is less pronounced when applied in conjunction with nanoparticle-mediated delivery. Thus when Angiopep-2 was grafted onto the surface of liposomes, the recovery of the targeted particles in the brain was negligible, and even less than that of liposomes grafted with RI7217 (the mouse analog of the transferrin antibody OX26, discussed above) [90]. Accordingly, these data would suggest that the transport pathway and/or transport efficiency of Angiopep-2 as target entity may very much depend on the attached cargo (drug versus nanoparticle). It is finally interesting to note that in BBB endothelial cells, newly synthesized LRP1 is sorted to the basolateral surface (i.e., facing brain tissue) [91]. However, since LRP1 apparently operates as transporter from the basolateral to apical surface in endothelial cells [83], these observations would thus imply that the transient appearance of LRP1 at the apical, i.e., blood facing surface, suffices to allow for an interaction with ligands like Angiopep-2 thus allowing in this manner brain directed transport via the opposite transcytotic pathway.
4.6. Leptin
Leptin is a regulator of body weight by controlling the food intake. Post alimentary, leptin is secreted from the cells in the gastrointestinal tract, and is subsequently transported by the system circulation to the brain endothelium. A specific receptor ObR on the luminal side of the brain capillary ensures leptin receptor-mediated transcytosis and its translocation to the brain stem to relay the satiety signal. Hence, these observations would support the potential of the ObR receptor to serve as a transport system in brain delivery. Liposomes with a size of approx. 180 nm, and decorated with a leptin-derived peptide showed a significantly enhanced uptake in brain endothelium, compared to liposomes, devoid of the leptin-peptide [92]. The internalization was sensitive to amiloride, but not filipin III or chlorpromazine, interfering with caveolae- and clathrin-mediated endocytosis, respectively. These data would thus suggest that these latter pathways as mechanisms for cellular entry of the leptin-targeted liposomes can be excluded. Rather, sensitivity toward amiloride would favor macropinocytosis as the mechanism of entry, and this pathway indeed has been linked to transcytosis in epithelia [93,94]. Of further interest, the tissue distribution of the ObR receptor is restricted, with a high expression at the BBB endothelial cells [95], while the receptor is occupied only after meals. These features are of particular interest and merit further investigations as to the development of the leptin–ObR system as a potential target in brain delivery. Notably, obesity in humans is thought to be caused by an impaired transport of leptin across the blood–brain barrier (BBB) [96], making leptin-targeted drugs potentially inappropriate for use in obese individuals.
4.7. Thiamine
Thiamine or vitamine B1 is a water-soluble nutrient with many physiological implications. At the BBB, the expression of the thiamine transporter facilitates its transport into the brain. The transport process of thiamine-functionalized nanoparticles has been investigated by Lockman et al. [97]. Thus, (3H) thiamine was exposed on the surface of solid nanoparticles (67 ± 8.2 nm), prepared from oil-in-water microemulsion, via incorporation of a distearoyl phosphatidylethanolamine (DSPE) anchor. Detailed kinetic profiling of the thiamine-tagged nanoparticles, after in situ rat brain perfusion, revealed a passive brain permeation of the particles, following an association with the BBB thiamine carrier. However, after systemic injection, the brain targeting specificity of thiamine-tagged nanoparticles is lost. Likely, similar to other targeting vectors, thiamine-coated nanoparticles might compete for binding sites with endogenous thiamine or associate with thiamine transporters, localized at other tissues. In summary, the current status and insight concerning the efficiency of transport across and expression of the thiamine receptor on endothelial cells, does not seem to justify high expectations concerning its exploitation as a suitable target for brain-directed delivery of drugs.
4.8. Glutathione
Glutathione, a natural anti-oxidant, is found at high levels in the brain, and its receptor is abundantly expressed at the blood–brain barrier. Hence, liposomes coated with glutathione-conjugated PEG (G-Technology®) proved to mediate safe targeting and enhanced delivery of drugs into the brain [98,99,100]. Currently, clinical trials with patients with brain metastasis, originating from breast cancer, are ongoing with the lead product 2B3-101, which is doxorubicin formulated with the same technology.
4.9. Synthetic Opioid Peptides
Enkephalins are neuroactive peptides that are transported bidirectionally across the BBB in a saturable and nonsaturable mode. The saturable mode likely arises from a specialized Peptide Transport System(s) (PTS; [101]). The PTS1 operates in a brain-to-blood direction and is responsible for the efficient clearance of enkephalins from the brain [102]. The opposite transport-brain influx-of enkephalins, although documented after in situ perfusion [103], is not sufficient to account for therapeutically meaningful amounts for systemic use [104]. Lipidation [105] or glycosylation compensates for the negative net influx, as demonstrated with peripherally administered glycoenkephalins that show antinociceptive activity comparable to that of morphine [106]. An effect that was correlated to glycoenkephalins enhanced blood-to-brain transport across the BBB and bioavailability. In subsequent work, the nature of brain influx of glyconeuropeptides was attributed to their amphipathic structure, and adsorptive endocytosis was proposed as main entry in the BBB endothelium [107]. Based on these findings and in an attempt to simulate similar transport mechanism across the BBB, a short seven amino acids peptide sequence was proposed to suffice for brain delivery [108]. The peptide, derived from the synthetic opioid peptide MMP-2200, which is an enkephalin analog, was deprived from “unnecessary” pharmacological activity and glycosylated at the C-terminal serine with sugar residues (d-glucose, d-galactose, d-xylose or d-lactose). Copolymer nanoparticles (200 nm) of poly (d,l-lactide-co-glycolide) (PLGA), assembled from peptide-modified PLGA copolymer, were administered by in situ perfusion. Confocal images of brain slices, obtained after perfusion, showed localization of the nanoparticles in the cerebral parenchyma for the glucose, galactose and the lactose variants, with higher accumulation of the glucose variant. Further, nanoparticles with the glucose-functionalized peptide were shown to accumulate in brain tissue, also after systemic administration. Proof of principle for therapeutic activity was obtained in several studies [109,110]. PLGA-nanoparticles, functionalized with the same glucose-bound peptide, were loaded with the model drug loperamide. The highly effective transport of loperamide (~13% of the injected dose) across the BBB following i.v. administration of the nanoparticle formulation resulted in a pharmacological effect. Moreover, total brain biodistribution studies, revealing that ~15% of the injected dose of the (fluorescently labeled) nanoparticles were transported into the brain, suggest the transport of drug-nanoparticle conjugates across the BBB. The macropinocytosis-like mechanism, which appears to depend on the Biousian structure of the peptide, is likely the mechanism by which these nanoparticles translocate across the BBB [111].
4.10. RVG Peptide
Rabies virus is a pathogen with a high neurotropism in vivo. The rabies virus glycoprotein (RVG) specifically interacts with the N-acetylcholine receptor on neuronal membranes, which mediates virus entry. A modified RVG peptide with 9 arginine residues that binds siRNA, was designed as a vector for transvascular delivery [112]. After intravenous injection into mice, the FITC-labeled siRNA, attached to the RVG-9R targeting entity, was detected in the brain, but not in spleen or liver. Functionality was demonstrated with anti-GFP siRNA, following intravenous injection. This construct silenced the GFP brain expression in GFP transgenic mice, with no significant effect on the GFP levels in spleen and liver. The therapeutic potential was further explored in mice with fatal flaviviral encephalitis. RVG-9R bound to antiviral siRNA rescued 80% of the infected animals. The acetylcholine receptor (AchR) is highly abundant in the CNS, including brain capillary endothelial cells. Binding of RVG to the alfa7 subunit of AchR likely promotes receptor-mediated transcytosis of RVG-9R–siRNA in brain microvessels. Hence, this vector emerges as a very promising tool when acetylcholinergic neurons are the desired target, as in the case of most senile dementias. However, RVG-9R–siRNA elicits IL-6 mediated immune responses. This effect was neutralized when employing exosomes, produced by self-derived dendritic cells [113], engineered to express Lamp2b, an exosomal membrane protein, fused to the RVG peptide, loaded with siRNA. The therapeutic potential of exosome-mediated siRNA delivery was demonstrated by a significant decrease in beta-secretase 1(BACE1) mRNA (60%) and protein (62%) levels in wild-type mice [113].
4.11. Tetanus Toxin, Tet1 and G23
Retrograde transport from distal peripheral axonal projections to the cell body, and transsynaptic spreading is a remarkable route used by the tetanus toxin (TeNT) to invade the CNS. Like other bacterial toxins, TeNT has two distinct functional peptide fragments. The C-terminal heavy chain fragment (TTC) is nontoxic, and piggybacks the light chain, which expresses pathogenic enzymatic properties. TTC per se displays a high affinity for GT1b, the receptor for TeNT, and is also efficiently transported along axons [114]. Indeed, targeting of fusion proteins containing the TTC fragment is readily accomplished. Thus after intramuscular injection of superoxide dismutase SOD–TTC [115] or β-gal–TTC fusion proteins [116] into the mouse tongue, the hybrid proteins follow the TTC retrograde route and are detected in higher order brain motor neurons. Also intraperitoneal injection of TTC protein hybrids into mice results in an enhanced CNS uptake [117]. Inspired by these observations, TTC has also been coupled to (d,l-lactide-co-glycolide) PLGA–PEG nanoparticles (size of approx. 250 nm) in order to accomplish transfer of the targeted nanoparticles to the brain parenchyma via the same transport pathway as the TTC fusion proteins [118]. As determined by flow cytometry a six-fold increase in binding of TTC-tagged nanoparticles was observed when binding to N18-RE-105 neuroblastoma cells was compared to that of HepG2 liver cells, thus reflecting the neuronal tropism of TTC. However, although such in vitro data are of interest, it remains to be determined whether in vivo these nanoparticles might reach with a similar effectiveness the CNS. Retrograde axonal transport of virus particles, which are similar in size as the applied nanoparticles is well documented [119]; it is therefore not a priori unlikely that TTC-nanoparticle transport may occur in a similar manner. Thus, TTC emerges as a highly potent vector in brain delivery. However, due to mass vaccination, its immunogenicity remains a serious concern. A potential solution to this problem has been provided by a phage library approach, biopanned on GT1b [120] in search of peptides displaying a binding efficiency and subsequent cellular processing, analogous to that of TTC. The identified peptide Tet1 fulfilled the required criteria. Importantly, Tet1 is similarly transported in a retrograde mode [121]. More recently, screening a peptide phage library against the ganglioside GM1, which is ubiquitously expressed in caveolae at the apical surface of vascular endothelial cells, we identified a peptide, defined as G23, with a sequence identical to Tet1 [122]. Detailed studies revealed that G23 effectively targets a transcytotic pathway in human brain capillary endothelial cells, and mediates efficient transport of G23-coated polymersomes across an in vitro BBB model [122]. Moreover, these studies also provided evidence for in vivo transfer across the BBB of intact peptide-targeted drug delivery vehicles into brain parenchyma, following intracarotid artery injection in mice. Further work confirmed that although specifically selected for GM1 as its targeting receptor, G23, like Tet1, also binds to the structurally similar ganglioside GT1b, implying that both gangliosides may play a role in transcytotic transport of G23-coupled polymersomes across the BBB. These promising observations warrant further investigations, in particular with regard to the design of G23-coated carriers that effectively mediate the actual delivery of drugs into the brain, i.e., the release of carrier contents, preferably in a programmable manner.
4.12. Diphteria Toxin
The non-toxic analog of the diphteria toxin (DT), CRM197, might be advantageous as carrier system because its receptor HB–EGF (membrane-bound precursor of heparin-binding epidermal growth factor) is specifically expressed on the BBB endothelium, neurons and glia, while the expression is up-regulated in disease-related conditions. Furthermore, the receptor lacks endogenous ligands. HRP-conjugates of CRM197, Tf or BSA were compared as targeting vectors in an in vitro co-culture of bovine BCEC with rat astrocytes [123,124]. CRM197 promoted the translocation of HRP at 37 °C, whereas the process was blocked at 4 °C, suggesting an energy-dependent and presumably receptor-mediated uptake and transport. Histological examinations of saline-perfused guinea pig brains, after bolus injection of CRM197-HRP or Tf-HRP, suggests a preferential extravasation and accumulation of the CRM197-conjugated molecule in the brain. Whether the CRM197 vector is able to trigger effective transcytosis, i.e., as effective as its toxic counterpart DT, remains to be determined.
4.13. TAT peptide
Cell penetrating peptides (CPPs), i.e., short cationic peptides, are able to transduce cargo across membranes. Through non-specific electrostatic interactions, CPPs adsorb strongly to the negatively charged cell surface [125]. Although the mechanism of CPPs translocation is still debated, involving issues, such as the formation of transient pores or inverted micelles, the conjugation of CPPs to cargo is thought to promote the rate and extent of cellular uptake. As an example of a CPP, the TAT (transduction domain of human immunodeficiency virus type-1 (HIV-1)) has been extensively studied. After intraperitoneal injection, the TAT peptide considerably increases the ubiquitous tissue penetration of a TAT–β-galactosidase conjugate beyond the capillaries, including the brain [126]. Based on these observation CPPs have been suggested to potentially act as vectors for enhanced brain delivery [127]. The suggestion was further investigated in a study with TAT-functionalized quantum dots (Qdots) for brain imaging. After intracarotid artery administration, the Qdots signal is detected in the brain parenchyma, although a very high accumulation of the particles was seen in arteries [128]. In contrast, in an in vitro study using a similar approach in which the peptide was linked to the cargo, GFP–TAT conjugates were shown to transmigrate through an intact endothelial monolayer, but to an extent very similar as observed for GFP alone [129]. These results thus question the role of TAT in mediating enhanced transport across the BBB. However, there appears to be no consensus as other data have suggested that the TAT peptide is of interest for nanoparticle-mediated drug delivery. In a study in which TAT-conjugated poly (l-lactide) nanoparticles were employed, loaded with the protease inhibitor ritonavir, prolonged sustained release of the drug in brain parenchyma was reported [130]. However, both TAT-conjugated nanoparticles and those devoid of TAT localized abundantly in the ventricles. Accordingly, it is also possible that penetration of released ritonavir may have occurred via cerebrospinal fluid into the brain, rather than via direct delivery involving nanoparticle transport across the BBB. In another study, self-assembled micelles prepared from TAT-PEG-b-cholesterol, labeled with Qdots, were detected 4 h after i.v. injection in rat hippocampus [131]. Whether the TAT–PEG-b-cholesterol micelles remain in the endothelial cells or do acquire access to brain cells is difficult to determine, without specific labeling. Indeed, data obtained in our laboratory provides evidence for a dramatic increase in association of TAT-coated polymersomes, relative to non-coated control polymersomes, to hCMEC/D3 BBB cells, in agreement with the afore mentioned studies (Figure 3a,c).
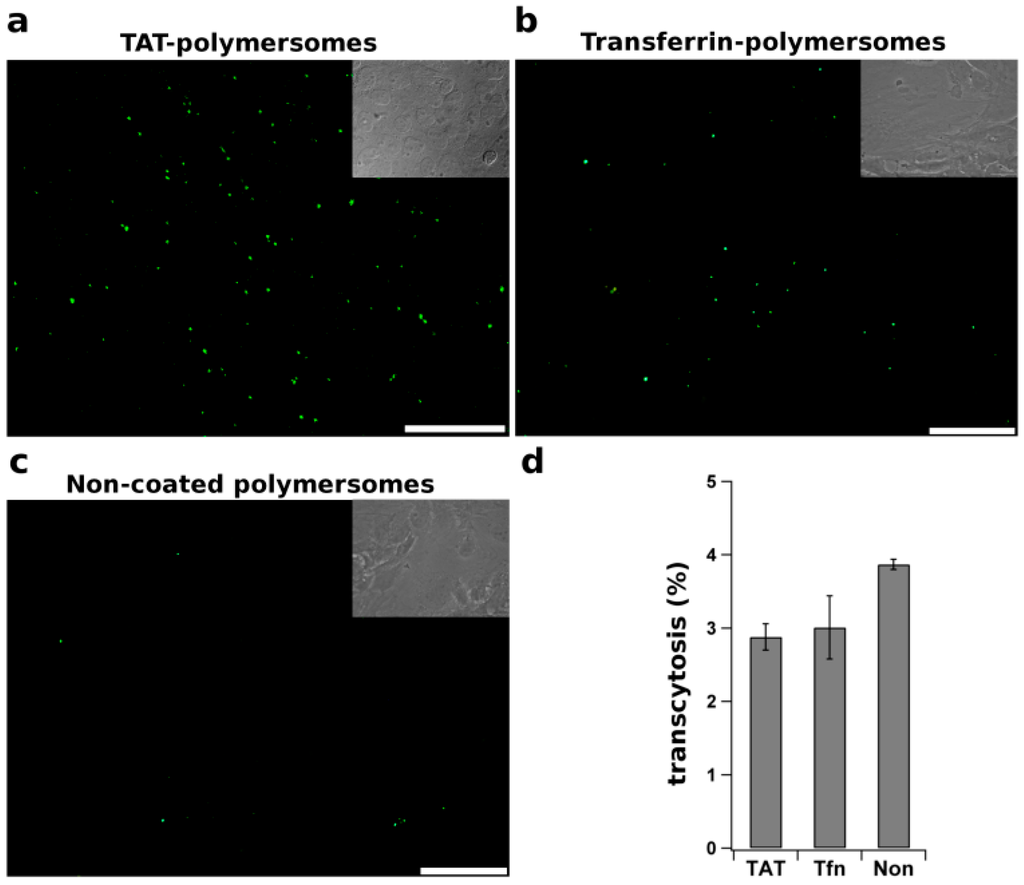
Figure 3.
Internalization and transcytosis of functionalized polymeric nanoparticles. TAT peptide and transferrin were covalently attached to polymersomes and incubated for 18 h at 37 °C with polarized filter-grown endothelial hCMEC/D3 cells, a convenient in vitro model for the BBB (see [132]). After extensive washing to remove non-bound particles, the cells were examined by fluorescence microscopy to visualize the intracellular distribution of (a) TAT-, Transferrin (Tf)- and non-coated polymersomes, as indicated (Scale bar: 20 µm); Alternatively (b), after 18 h of incubation at 37 °C the fractions of polymersomes in the apical and basolateral medium were measured and the fraction of polymersomes, transported by transcytosis across the endothelial cells, was calculated.
The effect, however, did not correlate with an increased transcytosis rate across the BBB endothelium in vitro (Figure 3b), thus questioning the potential of TAT as an effective vector for nanoparticulate brain delivery. Additionally, enhanced ubiquitous tissue accumulation of the cargo-ligand unit remains an essential obstacle of CPPs usability, as organ specificity was not demonstrated thus far.
5. Concluding Remarks
Here, we have discussed the rationale behind the selection of certain receptors, together with the pros and cons of their usability for brain drug delivery. An increasing number of studies on targeted delivery supports the initial hypothesis on a Trojan horse strategy: an appropriate receptor may ferry ligand-coupled cargo/drugs across the BBB. While direct comparison of the listed studies is precluded because of the different protocols used to assess the therapeutic outcome, clearly few vectors are emerging that may be considered as a successful “new generation” of targets. Nevertheless, the glutathione technology, developed for nanoparticle delivery, and the ANG1005 for direct drug-ligand delivery, already entered the stage of clinical evaluation for treatment of brain tumors. Also the tetanus toxin (TTC) and/or its functional competitor (Tet1/G23) and the RVG-9R peptide warrant special attention as very efficient vectors for drug delivery from the system circulation into the brain. The important advantage is that these vectors also hold natural neurotropism with specificity towards certain neuron subpopulations. Tet1 was developed to attack motor neurons and addresses unmet needs in treatment of amyotrophic lateral sclerosis. Similarly, targeting of acetyl cholinergic neurons with RVG-9R-acetylcholinesterase inhibitors would allow for determination of an optimal dosage and hence reduced adverse effects in symptomatic treatment of age-related memory impairments or any neuropathology with disturbed acetylcholine levels.
While researchers are still in search of the ideal vector, gathering more insight into the nature of transcellular transport, combined with identifying novel BBB specific receptors may pave future pathways in crossing the blood–brain barrier. Here, improvement of the current in vitro models of the BBB may further improve the predictive value of in vitro experiments, Cocultures of brain endothelial cells (ECs) with pericytes and astrocytes, thereby reconstituting the properties of the neurovascular unit, may reflect the in vivo properties of the BBB better than monocultures of brain ECs. Moreover, future research may benefit from imaging approaches using drug–carrier conjugates in which the drug and the carrier each carry their own label, allowing to measure the presence of intact conjugates as well as free drugs. This will help to determine the amount of free drug in the brain that is needed to exert a therapeutic effect, as well as provide information on the involvement of nanocarrier transcytosis across the BBB, and is therefore of great value for assessing the significance of the Trojan horse strategy for drug delivery into the brain.
Acknowledgments
This work was supported by the Dutch Top Institute Pharma project T5-105.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte-endothelial interactions at the blood–brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Ballabh, P.; Braun, A.; Nedergaard, M. The blood–brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, S.; Tian, F.; Stoeger, T.; Kreyling, W.; de la Fuente, J.M.; Grazú, V.; Borm, P.; Estrada, G.; Ntziachristos, V.; Razansky, D. Multifunctional nanocarriers for diagnostics, drug delivery and targeted treatment across blood–brain barrier: Perspectives on tracking and neuroimaging. Part. Fibre Toxicol. 2010, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Soni, U.; Patravale, V. Nano-interventions for neurodegenerative disorders. Pharmacol. Res. 2010, 62, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W. Metalloproteinases: Mediators of pathology and regeneration in the CNS. Nat. Rev. Neurosci. 2005, 6, 931–944. [Google Scholar] [CrossRef]
- Hori, S.; Ohtsuki, S.; Hosoya, K.; Nakashima, E.; Terasaki, T. A pericyte-derived angiopoietin-1 multimeric complex induces occludin gene expression in brain capillary endothelial cells through Tie-2 activation in vitro. J. Neurochem. 2004, 89, 503–513. [Google Scholar] [CrossRef]
- Wosik, K.; Cayrol, R.; Dodelet-Devillers, A.; Berthelet, F.; Bernard, M.; Moumdjian, R.; Bouthillier, A.; Reudelhuber, T.L.; Prat, A. Angiotensin II controls occludin function and is required for blood brain barrier maintenance: Relevance to multiple sclerosis. J. Neurosci. 2007, 27, 9032–9042. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood–brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef]
- Ben-Zvi, A.; Lacoste, B.; Kur, E.; Andreone, B.J.; Mayshar, Y.; Yan, H.; Gu, C. Mfsd2a is critical for the formation and function of the blood–brain barrier. Nature 2014, 509, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Celia, C.; Cosco, D.; Paolino, D.; Fresta, M. Nanoparticulate devices for brain drug delivery. Med. Res. Rev. 2010, 31, 716–756. [Google Scholar] [PubMed]
- Candela, P.; Gosselet, F.; Saint-Pol, J.; Sevin, E.; Boucau, M.-C.; Boulanger, E.; Cecchelli, R.; Fenart, L. Apical-to-basolateral transport of amyloid-β peptides through blood-brain barrier cells is mediated by the receptor for advanced glycation end-products and is restricted by P-glycoprotein. J. Alzheimers Dis. 2010, 22, 849–859. [Google Scholar] [PubMed]
- Kageyama, T.; Nakamura, M.; Matsuo, A.; Yamasaki, Y.; Takakura, Y.; Hashida, M.; Kanai, Y.; Naito, M.; Tsuruo, T.; Minato, N.; et al. The 4F2hc/LAT1 complex transports l-DOPA across the blood–brain barrier. Brain Res. 2000, 879, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, S.; Terasaki, T. Contribution of carrier-mediated transport systems to the blood-brain barrier as a supporting and protecting interface for the brain; importance for CNS drug discovery and development. Pharm. Res. 2007, 24, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Fan, W.; Chen, H.; Yao, N.; Tang, W.; Tang, J.; Yuan, W.; Kuai, R.; Zhang, Z.; Wu, Y.; et al. In vitro and in vivo investigation of glucose-mediated brain-targeting liposomes. J. Drug Target. 2010, 18, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Yao, N.; Qin, Y.; Zhang, Q.; Chen, H.; Yuan, M.; Tang, J.; Li, X.; Fan, W.; Zhang, Q.; et al. Investigation of glucose-modified liposomes using polyethylene glycols with different chain lengths as the linkers for brain targeting. Int. J. Nanomed. 2012, 7, 163–175. [Google Scholar] [CrossRef]
- Gromnicova, R.; Davies, H.A.; Sreekanthreddy, P.; Romero, I.A.; Lund, T.; Roitt, I.M.; Phillips, J.B.; Male, D.K. Glucose-coated gold nanoparticles transfer across human brain endothelium and enter astrocytes in vitro. PLoS One 2013, 8, e81043. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Xin, H.; Ren, Q.; Gu, J.; Zhu, L.; Du, F.; Feng, C.; Xie, Y.; Sha, X.; Fang, X. Nanoparticles of 2-deoxy-d-glucose functionalized poly(ethylene glycol)-co-poly(trimethylene carbonate) for dual-targeted drug delivery in glioma treatment. Biomaterials 2014, 35, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.L. Clathrin-coated vesicle formation and protein sorting: An integrated process. Annu. Rev. Biochem. 1997, 66, 511–548. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.-T.; Callaghan, J.M.; Steele-Mortimer, O.; Stenmark, H.; Parton, R.G.; Campbell, P.L.; McCluskey, J.; Yeo, J.-P.; Tock, E.P.C.; Toh, B.-H. EEA1, an early endosome-associated protein. EEA1 is a conserved α-helical peripheral membrane protein flanked by cysteine “fingers” and contains a calmodulin-binding IQ motif. J. Biol. Chem. 1995, 270, 13503–13511. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, F.R.; McGraw, T.E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 2004, 5, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Sharma, D.K.; Marks, D.L.; Pagano, R.E. Elevated endosomal cholesterol levels in niemann-pick cells inhibit rab4 and perturb membrane recycling. Mol. Biol. Cell 2004, 15, 4500–4511. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.F. Maturation models for endosome and lysosome biogenesis. Trends Cell Biol. 1991, 1, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.; Gruenberg, J. The arguments for pre-existing early and late endosomes. Trends Cell Biol. 1991, 1, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I. The importance of being acid: The role of acidification in intracellular membrane traffic. J. Exp. Biol. 1992, 172, 39–45. [Google Scholar] [PubMed]
- Schnitzer, J.E.; Oh, P.; Pinney, E.; Allard, J. Filipin-sensitive caveolae-mediated transport in endothelium: Reduced transcytosis, scavenger endocytosis, and capillary permeability of select macromolecules. J. Cell Biol. 1994, 127, 1217–1232. [Google Scholar] [CrossRef] [PubMed]
- Dehouck, B.; Fenart, L.; Dehouck, M.P.; Pierce, A.; Torpier, G.; Cecchelli, R. A new function for the LDL receptor: Transcytosis of LDL across the blood-brain barrier. J. Cell Biol. 1997, 138, 877–889. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, D.P.; Tan, X.-Y.; Oh, P.; Schnitzer, J.E. Targeting endothelium and its dynamic caveolae for tissue-specific transcytosis in vivo: A pathway to overcome cell barriers to drug and gene delivery. Proc. Natl. Acad. Sci. USA 2002, 99, 1996–2001. [Google Scholar] [CrossRef]
- Anderson, R.G.W. The caveolae membrane system. Annu. Rev. Biochem. 1998, 67, 199–225. [Google Scholar] [CrossRef]
- Thorn, H.; Stenkula, K.G.; Karlsson, M.; Örtegren, U.; Nystrom, F.H.; Gustavsson, J. Strålfors, P. Cell surface orifices of caveolae and localization of caveolin to the necks of caveolae in adipocytes. Mol. Biol. Cell 2003, 14, 3967–3976. [Google Scholar] [CrossRef] [PubMed]
- Newman, G.R.; Campbell, L.; von Ruhland, C.; Jasani, B.; Gumbleton, M. Caveolin and its cellular and subcellular immunolocalisation in lung alveolar epithelium: Implications for alveolar epithelial type I cell function. Cell Tissue Res. 1999, 295, 111–120. [Google Scholar] [CrossRef]
- Simionescu, N.; Simionescu, M.; Palade, G.E. Permeability of intestinal capillaries pathway followed by dextrans and glycogens. J. Cell Biol. 1972, 53, 365–392. [Google Scholar] [CrossRef] [PubMed]
- Parton, R.G.; Joggerst, B.; Simons, K. Regulated internalization of caveolae. J. Cell Biol. 1994, 127, 1199–1215. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, K.G.; Heuser, J.E.; Donzell, W.C.; Ying, Y.-S.; Glenney, J.R.; Anderson, R.G.W. Caveolin, a protein component of caveolae membrane coats. Cell 1992, 68, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.G.; Nichols, B.J. Exploring the caves: Cavins, caveolins and caveolae. Trends Cell Biol. 2010, 20, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Shaul, P.W.; Anderson, R.G. Role of plasmalemmal caveolae in signal transduction. Am. J. Physiol. 1998, 275, L843–L851. [Google Scholar]
- Smart, E.J.; Graf, G.A.; McNiven, M.A.; Sessa, W.C.; Engelman, J.A.; Scherer, P.E.; Okamoto, T.; Lisanti, M.P. Caveolins, liquid-ordered domains, and signal transduction. Mol. Cell. Biol. 1999, 19, 7289–7304. [Google Scholar] [PubMed]
- Marx, J. Caveolae: A once-elusive structure gets some respect. Science 2001, 294, 1862–1865. [Google Scholar] [CrossRef] [PubMed]
- Mundy, D.I.; Machleidt, T.; Ying, Y.; Anderson, R.G.W.; Bloom, G.S. Dual control of caveolar membrane traffic by microtubules and the actin cytoskeleton. J. Cell Sci. 2002, 115, 4327–4339. [Google Scholar] [CrossRef] [PubMed]
- Nichols, B.J.; Kenworthy, A.K.; Polishchuk, R.S.; Lodge, R.; Roberts, T.H.; Hirschberg, K.; Phair, R.D.; Lippincott-Schwartz, J. Rapid cycling of lipid raft markers between the cell surface and Golgi complex. J. Cell Biol. 2001, 153, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Candela, P.; Gosselet, F.; Miller, F.; Buee-Scherrer, V.; Torpier, G.; Cecchelli, R.; Fenart, L. Physiological pathway for low-density lipoproteins across the blood–brain barrier: Transcytosis through brain capillary endothelial cells in vitro. Endothelium 2008, 15, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, E.B.; Cooney, A.M.; Pitha, J.; Dawidowicz, E.A.; Dwyer, N.K.; Pentchev, P.G.; Blanchette-Mackie, E.J. Intracellular trafficking of cholesterol monitored with a cyclodextrin. J. Biol. Chem. 1996, 271, 21604–21613. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, P.A.; Fishman, P.H. Filipin-dependent inhibition of cholera toxin: Evidence for toxin internalization and activation through caveolae-like domains. J. Cell Biol. 1998, 141, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Nomura, R.; Fujimoto, T. Tyrosine phosphorylation of caveolin-1 in the endothelium. Exp. Cell Res. 1999, 253, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Simionescu, M.; Popov, D.; Sima, A. Endothelial transcytosis in health and disease. Cell Tissue Res. 2008, 335, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Biopharmaceutical drug targeting to the brain. J. Drug Target. 2010, 18, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Gupta, Y.; Jain, A.; Jain, S.K. Transferrin-conjugated solid lipid nanoparticles for enhanced delivery of quinine dihydrochloride to the brain. J. Pharm. Pharmacol. 2007, 59, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Buciak, J.L.; Friden, P.M. Selective transport of an anti-transferrin receptor antibody through the blood–brain barrier in vivo. J. Pharmacol. Exp. Ther. 1991, 259, 66–70. [Google Scholar]
- Boado, R.J.; Tsukamoto, H.; Pardridge, W.M. Drug delivery of antisense molecules to the brain for treatment of Alzheimer’s disease and cerebral AIDS. J. Pharm. Sci. 1998, 87, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Lu, W.; Gao, H.; Hu, K.; Chen, J.; Zhang, C.; Gao, X.; Jiang, X.; Zhu, C. Preparation and brain delivery property of biodegradable polymersomes conjugated with OX26. J. Control. Release 2008, 128, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, K.; Hekmatara, T.; Herbert, E.; Kreuter, J. Transferrin- and transferrin-receptor-antibody-modified nanoparticles enable drug delivery across the blood-brain barrier (BBB). Eur. J. Pharm. Biopharm. 2009, 71, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Paris-Robidas, S.; Emond, V.; Tremblay, C.; Soulet, D.; Calon, F. In vivo labeling of brain capillary endothelial cells after intravenous injection of monoclonal antibodies targeting the transferrin receptor. Mol. Pharmacol. 2011, 80, 32–39. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Restricted transport of anti-transferrin receptor antibody (OX26) through the blood–brain barrier in the rat. J. Neurochem. 2001, 79, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Gosk, S.; Vermehren, C.; Storm, G.; Moos, T. Targeting anti-transferrin receptor antibody (OX26) and OX26-conjugated liposomes to brain capillary endothelial cells using in situ perfusion. J. Cereb. Blood Flow Metab. 2004, 24, 1193–1204. [Google Scholar] [CrossRef]
- Yu, Y.J.; Zhang, Y.; Kenrick, M.; Hoyte, K.; Luk, W.; Lu, Y.; Atwal, J.; Elliott, J.M.; Prabhu, S.; Watts, R.J.; et al. Boosting brain uptake of a therapeutic antibody by reducing its affinity for a transcytosis target. Sci. Transl. Med. 2011, 3, 84ra44. [Google Scholar] [CrossRef] [PubMed]
- Niewoehner, J.; Bohrmann, B.; Collin, L.; Urich, E.; Sade, H.; Maier, P.; Rueger, P.; Stracke, J.O.; Lau, W.; Tissot, A.C.; et al. Increased brain penetration and potency of a therapeutic antibody using a monovalent molecular shuttle. Neuron 2014, 81, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Food, M.R.; Rothenberger, S.; Gabathuler, R.; Haidl, I.D.; Reid, G.; Jefferies, W.A. Transport and expression in human melanomas of a transferrin-like glycosylphosphatidylinositol-anchored protein. J. Biol. Chem. 1994, 269, 3034–3040. [Google Scholar] [PubMed]
- Demeule, M.; Poirier, J.; Jodoin, J.; Bertrand, Y.; Desrosiers, R.R.; Dagenais, C.; Nguyen, T.; Lanthier, J.; Gabathuler, R.; Kennard, M.; et al. High transcytosis of melanotransferrin (P97) across the blood–brain barrier. J. Neurochem. 2002, 83, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Karkan, D.; Pfeifer, C.; Vitalis, T.Z.; Arthur, G.; Ujiie, M.; Chen, Q.; Tsai, S.; Koliatis, G.; Gabathuler, R.; Jefferies, W.A. A unique carrier for delivery of therapeutic compounds beyond the blood-brain barrier. PLoS One 2008, 3, e2469. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Eisenberg, J.; Yang, J. Human blood–brain barrier insulin receptor. J. Neurochem. 1985, 44, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Zhou, Q.-H.; Pardridge, W.M. Selective targeting of a TNFR decoy receptor pharmaceutical to the primate brain as a receptor-specific IgG fusion protein. J. Biotechnol. 2010, 146, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Schlachetzki, F.; Zhu, C.; Pardridge, W.M. Expression of the neonatal Fc receptor (FcRn) at the blood-brain barrier. J. Neurochem. 2002, 81, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Ohshima-Hosoyama, S.; Simmons, H.A.; Goecks, N.; Joers, V.; Swanson, C.R.; Bondarenko, V.; Velotta, R.; Brunner, K.; Wood, L.D.; Hruban, R.H.; et al. A monoclonal antibody–GDNF fusion protein is not neuroprotective and is associated with proliferative pancreatic lesions in Parkinsonian monkeys. PLoS One 2012, 7, e39036. [Google Scholar] [PubMed]
- Alyautdin, R.; Gothier, D.; Petrov, V.; Kharkevich, D.; Kreuter, J. Analgesic activity of the hexapeptide dalargin adsorbed on the surface of polysorbate 80-coated poly (butyl cyanoacrylate) nanoparticles. Eur. J. Pharm. Biopharm. 1995, 41, 44–48. [Google Scholar]
- Alyautdin, R.N.; Petrov, V.E.; Langer, K.; Berthold, A.; Kharkevich, D.A.; Kreuter, J. Delivery of loperamide across the blood-brain barrier with polysorbate 80-coated polybutylcyanoacrylate nanoparticles. Pharm. Res. 1997, 14, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Alyautdin, R.N.; Tezikov, E.B.; Ramge, P.; Kharkevich, D.A.; Begley, D.J.; Kreuter, J. Significant entry of tubocurarine into the brain of rats by adsorption to polysorbate 80-coated polybutylcyanoacrylate nanoparticles: An in situ brain perfusion study. J. Microencapsul. 1998, 15, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Gulyaev, A.E.; Gelperina, S.E.; Skidan, I.N.; Antropov, A.S.; Kivman, G.Y.; Kreuter, J. Significant transport of doxorubicin into the brain with polysorbate 80-coated nanoparticles. Pharm. Res. 1999, 16, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Olivier, J.C.; Fenart, L.; Chauvet, R.; Pariat, C.; Cecchelli, R.; Couet, W. Indirect evidence that drug brain targeting using polysorbate 80-coated polybutylcyanoacrylate nanoparticles is related to toxicity. Pharm. Res. 1999, 16, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, J.; Ramge, P.; Petrov, V.; Hamm, S.; Gelperina, S.E.; Engelhardt, B.; Alyautdin, R.; von Briesen, H.; Begley, D.J. Direct evidence that polysorbate-80-coated poly (butylcyanoacrylate) nanoparticles deliver drugs to the CNS via specific mechanisms requiring prior binding of drug to the nanoparticles. Pharm. Res. 2003, 20, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Alyaudtin, R.N.; Reichel, A.; Löbenberg, R.; Ramge, P.; Kreuter, J.; Begley, D.J. Interaction of poly (butylcyanoacrylate) nanoparticles with the blood-brain barrier in vivo and in vitro. J. Drug Target. 2001, 9, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Lück, M.; Schröder, W.; Harnisch, S.; Thode, K.; Blunk, T.; Paulke, B.R.; Kresse, M.; Müller, R.H. Identification of plasma proteins facilitated by enrichment on particulate surfaces: Analysis by two-dimensional electrophoresis and N-terminal microsequencing. Electrophoresis 1997, 18, 2961–2967. [Google Scholar] [CrossRef] [PubMed]
- Lemke, A.; Kiderlen, A.F.; Petri, B.; Kayser, O. Delivery of amphotericin B nanosuspensions to the brain and determination of activity against Balamuthia mandrillaris amebas. Nanomedicine 2010, 6, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Samanta, M.K.; Santhi, K.; Kumar, K.P.S.; Paramakrishnan, N.; Suresh, B. Targeted delivery of tacrine into the brain with polysorbate 80-coated poly (n-butylcyanoacrylate) nanoparticles. Eur. J. Pharm. Biopharm. 2008, 70, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Göppert, T.M.; Müller, R.H. Polysorbate-stabilized solid lipid nanoparticles as colloidal carriers for intravenous targeting of drugs to the brain: Comparison of plasma protein adsorption patterns. J. Drug Target. 2005, 13, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Zensi, A.; Begley, D.; Pontikis, C.; Legros, C.; Mihoreanu, L.; Wagner, S.; Büchel, C.; von Briesen, H.; Kreuter, J. Albumin nanoparticles targeted with Apo E enter the CNS by transcytosis and are delivered to neurones. J. Control. Release 2009, 137, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, M.; Azadi, A.; Rafiei, P. Pharmacokinetic consequences of pegylation. Drug Deliv. 2006, 13, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, J.; Hekmatara, T.; Dreis, S.; Vogel, T.; Gelperina, S.; Langer, K. Covalent attachment of apolipoprotein A-I and apolipoprotein B-100 to albumin nanoparticles enables drug transport into the brain. J. Control. Release 2007, 118, 54–58. [Google Scholar] [PubMed]
- Zensi, A.; Begley, D.; Pontikis, C.; Legros, C.; Mihoreanu, L.; Büchel, C.; Kreuter, J. Human serum albumin nanoparticles modified with apolipoprotein A–I cross the blood–brain barrier and enter the rodent brain. J. Drug Target. 2010, 18, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Dehouck, B.; Dehouck, M.P.; Fruchart, J.C.; Cecchelli, R. Upregulation of the low density lipoprotein receptor at the blood-brain barrier: Intercommunications between brain capillary endothelial cells and astrocytes. J. Cell Biol. 1994, 126, 465–473. [Google Scholar] [PubMed]
- Fillebeen, C.; Descamps, L.; Dehouck, M.P.; Fenart, L.; Benaïssa, M.; Spik, G.; Cecchelli, R.; Pierce, A. Receptor-mediated transcytosis of lactoferrin through the blood-brain barrier. J. Biol. Chem. 1999, 274, 7011–7017. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Kastin, A.J.; Zankel, T.C.; van Kerkhof, P.; Terasaki, T.; Bu, G. Efficient transfer of receptor-associated protein (RAP) across the blood-brain barrier. J. Cell Sci. 2004, 117, 5071–5078. [Google Scholar] [CrossRef] [PubMed]
- Benchenane, K.; Berezowski, V.; Ali, C.; Fernández-Monreal, M.; López-Atalaya, J.P.; Brillault, J.; Chuquet, J.; Nouvelot, A.; MacKenzie, E.T.; Bu, G.; et al. Tissue-type plasminogen activator crosses the intact blood–brain barrier by low-density lipoprotein receptor-related protein-mediated transcytosis. Circulation 2005, 111, 2241–2249. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.D.; Sagare, A.P.; Friedman, A.E.; Bedi, G.S.; Holtzman, D.M.; Deane, R.; Zlokovic, B.V. Transport pathways for clearance of human Alzheimer’s amyloid beta-peptide and apolipoproteins E and J in the mouse central nervous system. J. Cereb. Blood Flow Metab. 2007, 27, 909–918. [Google Scholar]
- Kounnas, M.Z.; Moir, R.D.; Rebeck, G.W.; Bush, A.I.; Argraves, W.S.; Tanzi, R.E.; Hyman, B.T.; Strickland, D.K. LDL receptor-related protein, a multifunctional ApoE receptor, binds secreted beta-amyloid precursor protein and mediates its degradation. Cell 1995, 82, 331–340. [Google Scholar] [PubMed]
- Demeule, M.; Régina, A.; Ché, C.; Poirier, J.; Nguyen, T.; Gabathuler, R.; Castaigne, J.-P.; Béliveau, R. Identification and design of peptides as a new drug delivery system for the brain. J. Pharmacol. Exp. Ther. 2008, 324, 1064–1072. [Google Scholar] [CrossRef] [PubMed]
- Demeule, M.; Currie, J.-C.; Bertrand, Y.; Ché, C.; Nguyen, T.; Régina, A.; Gabathuler, R.; Castaigne, J.-P.; Béliveau, R. Involvement of the low-density lipoprotein receptor-related protein in the transcytosis of the brain delivery vector angiopep-2. J. Neurochem. 2008, 106, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Régina, A.; Demeule, M.; Ché, C.; Lavallée, I.; Poirier, J.; Gabathuler, R.; Béliveau, R.; Castaigne, J.-P. Antitumour activity of ANG1005, a conjugate between paclitaxel and the new brain delivery vector Angiopep-2. Br. J. Pharmacol. 2008, 155, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Gabathuler, R. An engineered peptide compound platform technology incorporating angiopep for crossing the BBB. In Drug Delivery to the Central Nervous System; Jain, K.K., Ed.; Humana Press: Totowa, NJ, USA, 2010; Volume 45, pp. 249–260. [Google Scholar]
- Lin, N.U.; Schwartzberg, L.; Kesari, S.; Elias, A.; Anders, C.; Raizer, J.; Kozloff, M.; Amiri-Kordestani, L. Abstract B76: A phase II study of ANG1005, a novel, brain-penetrant taxane derivative, in breast cancer patients with brain metastases. Mol. Cancer Ther. 2013, 12, B76–B76. [Google Scholar] [CrossRef]
- Van Rooy, I.; Mastrobattista, E.; Storm, G.; Hennink, W.E.; Schiffelers, R.M. Comparison of five different targeting ligands to enhance accumulation of liposomes into the brain. J. Control. Release 2011, 150, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Neuwelt, E.A.; Bauer, B.; Fahlke, C.; Fricker, G.; Iadecola, C.; Janigro, D.; Leybaert, L.; Molnár, Z.; O’Donnell, M.E.; Povlishock, J.T.; et al. Engaging neuroscience to advance translational research in brain barrier biology. Nat. Rev. Neurosci. 2011, 12, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, M.; Akita, H.; Fujiwara, T.; Kajimoto, K.; Harashima, H. Leptin-derived peptide, a targeting ligand for mouse brain-derived endothelial cells via macropinocytosis. Biochem. Biophys. Res. Commun. 2010, 394, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Malyukova, I.; Murray, K.F.; Zhu, C.; Boedeker, E.; Kane, A.; Patterson, K.; Peterson, J.R.; Donowitz, M.; Kovbasnjuk, O. Macropinocytosis in Shiga toxin 1 uptake by human intestinal epithelial cells and transcellular transcytosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G78–G92. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsson, E.G.E.; Schoultz, I.; Gullberg, E.; Carlsson, A.H.; Tafazoli, F.; Lerm, M.; Magnusson, K.-E.; Söderholm, J.D.; Artursson, P. Yersinia pseudotuberculosis induces transcytosis of nanoparticles across human intestinal villus epithelium via invasin-dependent macropinocytosis. Lab. Investig. 2008, 88, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Bjørbaek, C.; Elmquist, J.K.; Michl, P.; Ahima, R.S.; van Bueren, A.; McCall, A.L.; Flier, J.S. Expression of leptin receptor isoforms in rat brain microvessels. Endocrinology 1998, 139, 3485–3491. [Google Scholar] [CrossRef] [PubMed]
- Van Heek, M.; Compton, D.S.; France, C.F.; Tedesco, R.P.; Fawzi, A.B.; Graziano, M.P.; Sybertz, E.J.; Strader, C.D.; Davis, H.R., Jr. Diet-induced obese mice develop peripheral, but not central, resistance to leptin. J. Clin. Investig. 1997, 99, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Lockman, P.R.; Oyewumi, M.O.; Koziara, J.M.; Roder, K.E.; Mumper, R.J.; Allen, D.D. Brain uptake of thiamine-coated nanoparticles. J. Control. Release 2003, 93, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, P.J.; de Boer, A.G. 2B-Trans technology: Targeted drug delivery across the blood–brain barrier. Methods Mol. Biol. 2008, 437, 161–175. [Google Scholar] [PubMed]
- Gaillard, P.J.; Appeldoorn, C.C.M.; Dorland, R.; van Kregten, J.; Manca, F.; Vugts, D.J.; Windhorst, B.; van Dongen, G.A.M.S.; de Vries, H.E.; Maussang, D.; et al. Pharmacokinetics, brain delivery, and efficacy in brain tumor-bearing mice of glutathione pegylated liposomal doxorubicin (2B3–101). PLoS One 2014, 9, e82331. [Google Scholar] [CrossRef] [PubMed]
- Rip, J.; Chen, L.; Hartman, R.; van den Heuvel, A.; Reijerkerk, A.; van Kregten, J.; van der Boom, B.; Appeldoorn, C.; de Boer, M.; Maussang, D.; et al. Glutathione PEGylated liposomes: Pharmacokinetics and delivery of cargo across the blood–brain barrier in rats. J. Drug Target. 2014, 22, 460–467. [Google Scholar] [PubMed]
- Banks, W.A.; Kastin, A.J. Peptide transport systems for opiates across the blood–brain barrier. Am. J. Physiol. 1990, 259, E1–E10. [Google Scholar] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V.; Mackic, J.B.; Djuricic, B.; Davson, H. Kinetic analysis of leucine-enkephalin cellular uptake at the luminal side of the blood–brain barrier of an in situ perfused guinea-pig brain. J. Neurochem. 1989, 53, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, C. Improved brain uptake and pharmacological activity of dalargin using a peptide-vector-mediated strategy. J. Pharmacol. Exp. Ther. 2003, 306, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, N.; Hama, T.; Hibi, T.; Konishi, R.; Futaki, S.; Kitagawa, K. Adamantane as a brain-directed drug carrier for poorly absorbed drug: Antinociceptive effects of (D-Ala2) Leu-enkephalin derivatives conjugated with the 1-adamantane moiety. Biochem. Pharmacol. 1991, 41, R5–R8. [Google Scholar] [CrossRef] [PubMed]
- Polt, D.R.; Palian, M.M. Glycopeptide analgesics. Drugs Future 2001, 26, 561. [Google Scholar] [CrossRef]
- Egleton, R.D.; Bilsky, E.J.; Tollin, G.; Dhanasekaran, M.; Lowery, J.; Alves, I.; Davis, P.; Porreca, F.; Yamamura, H.I.; Yeomans, L.; et al. Biousian glycopeptides penetrate the blood–brain barrier. Tetrahedron Asymmetry 2005, 16, 65–75. [Google Scholar] [CrossRef]
- Costantino, L.; Gandolfi, F.; Tosi, G.; Rivasi, F.; Vandelli, M.A.; Forni, F. Peptide-derivatized biodegradable nanoparticles able to cross the blood–brain barrier. J. Control. Release 2005, 108, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Tosi, G.; Costantino, L.; Rivasi, F.; Ruozi, B.; Leo, E.; Vergoni, A.V.; Tacchi, R.; Bertolini, A.; Vandelli, M.A.; Forni, F. Targeting the central nervous system: In vivo experiments with peptide-derivatized nanoparticles loaded with Loperamide and Rhodamine-123. J. Control. Release 2007, 122, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vergoni, A.V.; Tosi, G.; Tacchi, R.; Vandelli, M.A.; Bertolini, A.; Costantino, L. Nanoparticles as drug delivery agents specific for CNS: In vivo biodistribution. Nanomed. Nanotechnol. Biol. Med. 2009, 5, 369–377. [Google Scholar] [CrossRef]
- Tosi, G.; Fano, R.A.; Bondioli, L.; Badiali, L.; Benassi, R.; Rivasi, F.; Ruozi, B.; Forni, F.; Vandelli, M.A. Investigation on mechanisms of glycopeptide nanoparticles for drug delivery across the blood–brain barrier. Nanomedicine 2011, 6, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Wu, H.; McBride, J.L.; Jung, K.-E.; Kim, M.H.; Davidson, B.L.; Lee, S.K.; Shankar, P.; Manjunath, N. Transvascular delivery of small interfering RNA to the central nervous system. Nature 2007, 448, 39–43. [Google Scholar] [PubMed]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [PubMed]
- Sinha, K.; Box, M.; Lalli, G.; Schiavo, G.; Schneider, H.; Groves, M.; Siligardi, G.; Fairweather, N. Analysis of mutants of tetanus toxin Hc fragment: Ganglioside binding, cell binding and retrograde axonal transport properties. Mol. Microbiol. 2000, 37, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.M.; Hallewell, R.A.; Chen, L.L.; Fairweather, N.F.; Dougan, G.; Savitt, J.M.; Parks, D.A.; Fishman, P.S. Delivery of recombinant tetanus-superoxide dismutase proteins to central nervous system neurons by retrograde axonal transport. Exp. Neurol. 1997, 145, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Coen, L.; Osta, R.; Maury, M.; Brûlet, P. Construction of hybrid proteins that migrate retrogradely and transynaptically into the central nervous system. Proc. Natl. Acad. Sci. USA 1997, 94, 9400–9405. [Google Scholar] [CrossRef]
- Fishman, P.S.; Savitt, J.M.; Farrand, D.A. Enhanced CNS uptake of systemically administered proteins through conjugation with tetanus C-fragment. J. Neurol. Sci. 1990, 98, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Townsend, S.A.; Evrony, G.D.; Gu, F.X.; Schulz, M.P.; Brown, R.H., Jr.; Langer, R. Tetanus toxin C fragment-conjugated nanoparticles for targeted drug delivery to neurons. Biomaterials 2007, 28, 5176–5184. [Google Scholar] [CrossRef] [PubMed]
- Bearer, E.L.; Breakefield, X.O.; Schuback, D.; Reese, T.S.; LaVail, J.H. Retrograde axonal transport of herpes simplex virus: Evidence for a single mechanism and a role for tegument. Proc. Natl. Acad. Sci. USA 2000, 97, 8146–8150. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.; Teng, Q.; Garrity-Moses, M.; Federici, T.; Tanase, D.; Imperiale, M.J.; Boulis, N.M. A novel peptide defined through phage display for therapeutic protein and vector neuronal targeting. Neurobiol. Dis. 2005, 19, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Federici, T.; Liu, J.K.; Teng, Q.; Yang, J.; Boulis, N.M. A means for targeting therapeutics to peripheral nervous system neurons with axonal damage. Neurosurgery 2007, 60, 911–918. [Google Scholar]
- Georgieva, J.V.; Brinkhuis, R.P.; Stojanov, K.; Weijers, C.A.G.M.; Zuilhof, H.; Rutjes, F.P.J.T.; Hoekstra, D.; van Hest, J.C.M.; Zuhorn, I.S. Peptide-mediated blood–brain barrier transport of polymersomes. Angew. Chem. Int. Ed. 2012, 51, 8339–8342. [Google Scholar] [CrossRef]
- Gaillard, P.; Brink, A.; Deboer, A. Diphtheria toxin receptor-targeted brain drug delivery. Int. Congr. Ser. 2005, 1277, 185–198. [Google Scholar] [CrossRef]
- Gaillard, P.J.; Visser, C.C.; de Boer, A.G. Targeted delivery across the blood-brain barrier. Expert Opin. Drug Deliv. 2005, 2, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Patel, L.N.; Zaro, J.L.; Shen, W.-C. Cell penetrating peptides: Intracellular pathways and pharmaceutical perspectives. Pharm. Res. 2007, 24, 1977–1992. [Google Scholar] [PubMed]
- Schwarze, S.R. In vivo protein transduction: Delivery of a biologically active protein into the mouse. Science 1999, 285, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, M.; Hällbrink, M.; Prochiantz, A.; Langel, U. Cell-penetrating peptides. Trends Pharmacol. Sci. 2000, 21, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Santra, S.; Yang, H.; Stanley, J.T.; Holloway, P.H.; Moudgil, B.M.; Walter, G.; Mericle, R.A. Rapid and effective labeling of brain tissue using TAT-conjugated CdS: Mn/ZnS quantum dots. Chem. Commun. Camb. 2005, 3144–3146. [Google Scholar] [CrossRef]
- Simon, M.J.; Kang, W.H.; Gao, S.; Banta, S.; Morrison, B. Evaluation of the cell-penetrating peptide TAT as a trans-blood-brain barrier delivery vehicle. In Bioengineering Conference, Proceedings of the 2010 IEEE 36th Annual Northeast, New York, NY, USA, 26–28 March 2010; pp. 1–2.
- Rao, K.S.; Reddy, M.K.; Horning, J.L.; Labhasetwar, V. TAT-conjugated nanoparticles for the CNS delivery of anti-HIV drugs. Biomaterials 2008, 29, 4429–4438. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, K.; Lu, J.; Venkatraman, S.S.; Luo, D.; Ng, K.C.; Ling, E.-A.; Moochhala, S.; Yang, Y.-Y. Biologically active core/shell nanoparticles self-assembled from cholesterol-terminated PEG-TAT for drug delivery across the blood-brain barrier. Biomaterials 2008, 29, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, J.V.; Kalicharan, D.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Hoekstra, D.; Zuhorn, I.S. Surface characteristics of nanoparticles determine their intracellular fate in and processing by human blood-brain barrier endothelial cells in vitro. Mol. Ther. 2011, 19, 318–325. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).