
Extracellular Non-Coding RNAs in Cardiovascular Diseases


Abstract
1. Introduction
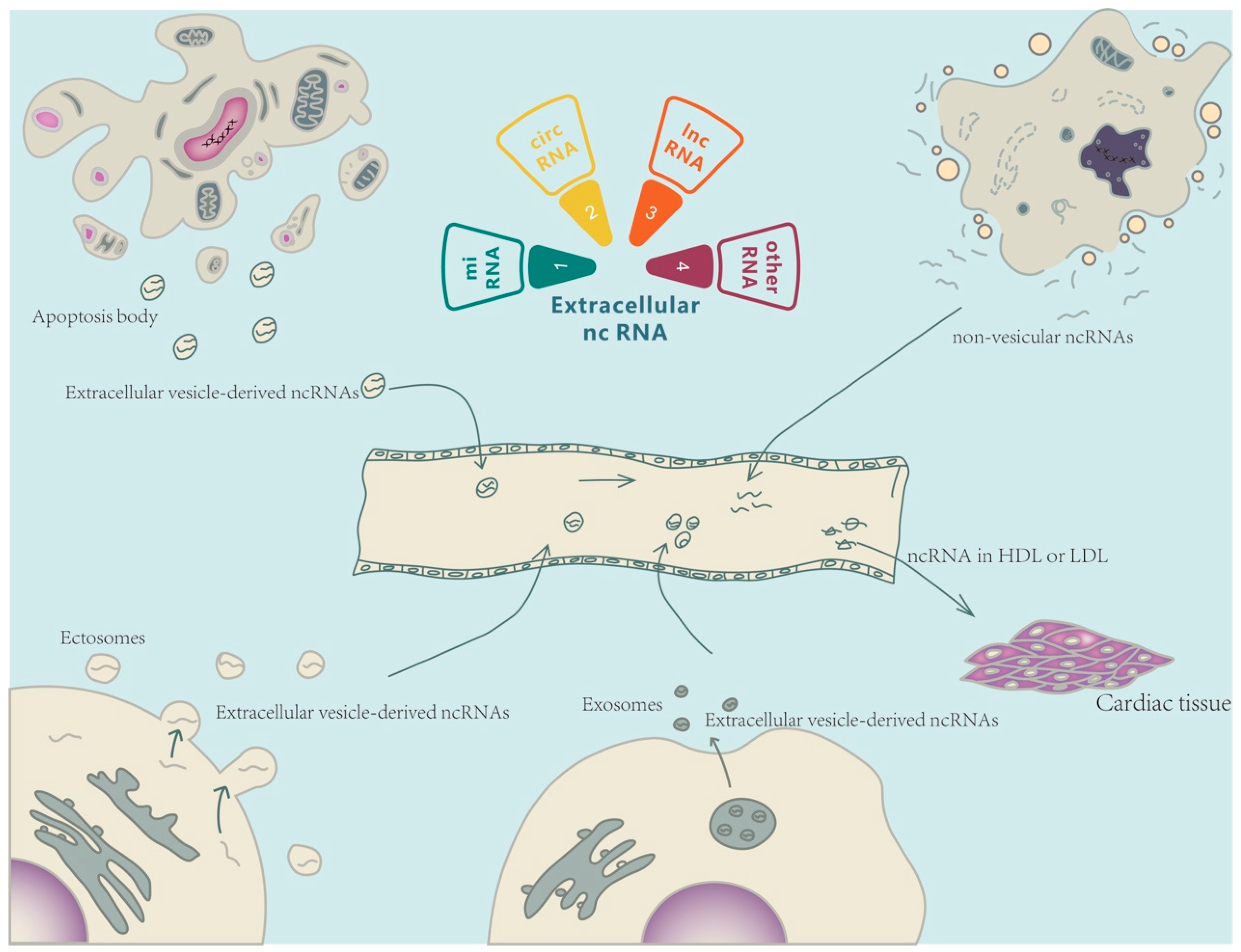
2. Ex-ncRNAs
2.1. Extracellular Small ncRNAs
2.2. Extracellular LncRNAs
2.3. Extracellular CircRNAs
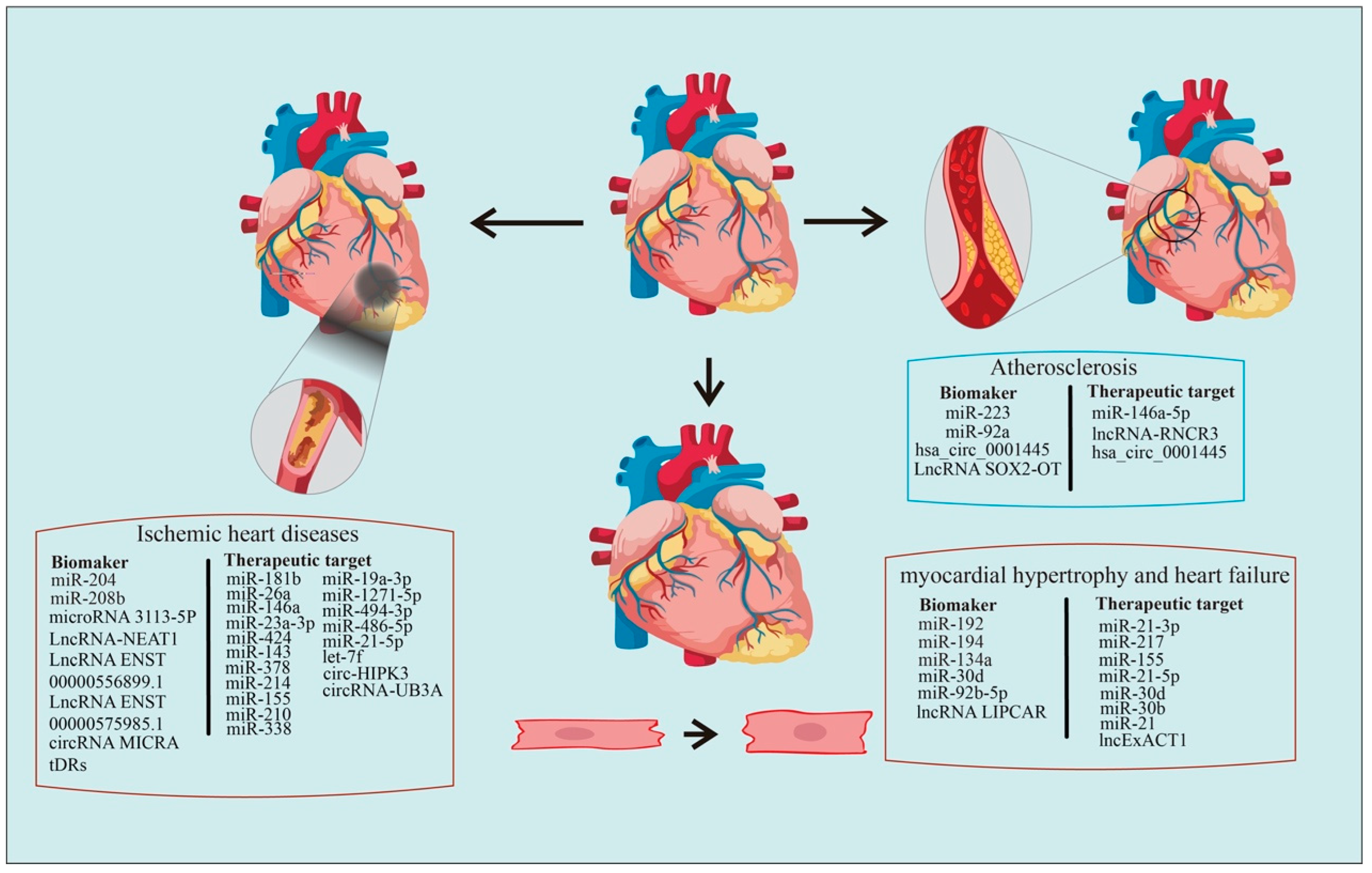
3. Ex-ncRNAs as Biomarkers in CVDs
3.1. Atherosclerosis
3.1.1. Extracellular Small ncRNAs
3.1.2. Extracellular LncRNAs
3.1.3. Extracellular CircRNAs
3.2. Ischemic Heart Disease
3.2.1. Extracellular Small ncRNAs
3.2.2. Extracellular LncRNAs
3.2.3. Extracellular CircRNAs
3.3. Hypertension
Extracellular Small ncRNAs
3.4. Heart Failure
3.4.1. Extracellular Small ncRNAs
3.4.2. Extracellular LncRNAs

{kind=link}
{kind=link}
| Reference | Ex-ncRNA | Carriers | Expression (↑) (↓) | Type of CVDs | |
|---|---|---|---|---|---|
| Kasey et al. [33] | miR-223 | HDL | ↑ | - | Atherosclerosis |
| Huang et al. [35] | miR-92a | - | ↑ | - | Atherosclerosis |
| Tao et al. [37] | LncRNA SOX2-OT | - | ↑ | - | Atherosclerosis |
| Vilades et al. [38] | hsa_circ_0001445 | - | - | ↓ | Atherosclerosis |
| Corsten et al. [46] | miR-208b | - | ↑ | - | Acute myocardial infarction |
| Li et al. [11] | tDR-1:32-His-GTG-1, tDR-37:72-Val-TAC-1, tDR-1:32-Pro-AGG-1-M4, etc. | - | ↑ | Cardiac ischemia/reperfusion | |
| tDR-1:36-Glu-CTC-1, tDR-1:36-Glu-CTC-1-D5G, tDR-1:36-Asp-GTC-2-M2, etc. | - | ↓ | |||
| Chen et al. [47] | LncRNA-NEAT1, miR-204 | EV | ↑ | ↓ | Acute ST-segment elevation myocardial infarction |
| Hildebrandt et al. [48] | miR-370-3p, miR-409-3p | EV | ↑ | - | Coronary heart disease |
| Chen et al. [50] | microRNA 3113-5P | - | ↑ | - | Cardiac ischemia/Reperfusion injury |
| Zheng et al. [53] | lncRNA ENST00000556899.1, lncRNA ENST00000575985.1 | EV | ↑ | - | Acute myocardial infarction |
| Kishore et al. [54] | circRNA MICRA, circRNA-284, hsa-circ-0098964 | - | ↑ | ↓ | Myocardial infarction, Ischemic heart disease, Hypertension |
| Suzuki et al. [61] | miR-133a, miR-27a | - | - | ↓ | Hypertension |
| Wang et al. [62] | miR-92a | - | ↑ | - | Hypertension |
| He et al. [66] | miR-200a-3p | - | ↑ | - | Hypertension |
| Janjusevic et al. [68] | lncRNA LIPCAR, miR-192, miR-134a, miR-194, miR-30d | EV | ↑ | ↓ | Heart failure |
| Li et al. [69] | miR-30d | EV | - | ↓ | Heart failure |
| Wu et al. [70] | miR-92b-5p | EV | ↑ | - | Heart failure |
4. Ex-ncRNAs as Therapeutic Targets in CVDs
4.1. Atherosclerosis
4.1.1. Extracellular Small ncRNA
4.1.2. Extracellular LncRNAs
4.1.3. Extracellular CircRNAs
4.2. Ischemic Heart Disease
4.2.1. Extracellular Small ncRNAs
4.2.2. Extracellular LncRNAs
4.2.3. Extracellular CircRNAs
4.3. Hypertension
Extracellular Small ncRNAs
4.4. Cardiac Hypertrophy
Extracellular Small ncRNAs
4.5. Heart Failure
4.5.1. Extracellular Small ncRNAs
4.5.2. Extracellular LncRNAs
| Reference | Donor Cells | Ex-ncRNAs | Carriers | Expression Quantity (↑) (↓) | Target/Pathway | Type of CVDs | |
|---|---|---|---|---|---|---|---|
| Shimada et al. [82] | - | miR-146a-5p | EV | ↑ | - | TLR7 | Atherosclerosis |
| Shan et al. [84] | Human umbilical vein endothelial cells | lncRNA-RNCR3 | EV | ↑ | - | miR-185-5p/KLF2 | Atherosclerosis |
| Yang et al. [85] | - | hsa_circ_0001445 | - | - | ↓ | mir-208b-5p/ABCG1 | Atherosclerosis |
| Liu et al. [89] | M1-like macrophage | miR-155 | EV | ↑ | - | Sirt1/AMPKα2 and RAC1-PAK2 | Acute myocardial infarction |
| Long et al. [90] | M2-like macrophage | miR-1271-5p | EV | ↑ | - | SOX6 | Acute myocardial infarction |
| Li et al. [69] | Cardiomyocytes | miR-30d | EV | ↑ | - | MAP4K4 and integrin α5 | Ischemic HF |
| Liu et al. [91] | Dendritic cell | miR-494-3p | EV | ↑ | - | VEGF | Myocardial infarction |
| Li et al. [92] | MSC | miR-486-5p | EV | ↑ | - | MMP 19/VEGF | Myocardial infarction |
| Cheng et al. [93] | MSC | miR-210 | EV | ↑ | - | AIFM3/PI3K/AKT and p53 | Myocardial infarction |
| Mao et al. [94] | MSC | miR-224-5p | EV | ↑ | - | TXNIP/GATA4/Bcl-2 | Ischemia-reperfusion injury |
| He et al. [95] | MSC | miR-150-5p | EV | ↑ | - | TXNIP | Ischemia-reperfusion injury |
| Li et al. [96] | BMSC | miR-29c | EV | ↑ | - | PTEN/Akt/mTOR | Ischemia-reperfusion injury |
| Zou et al. [97] | - | miR-149, let-7c | EV | ↑ | - | miR-149/Let-7c/FASLG | Ischemia-reperfusion injury |
| Fu et al. [98] | BMSC | miR-338 | EV | ↑ | - | MAP3K2/JNK axis | Myocardial infarction |
| Walravens et al. [99,100,101,102] | CDC | miR-26a, miR-146a, miR-181b | EV | ↑ | - | Adam17, TLR-NFkB, PKCδ | Myocardial infarction |
| Moghiman et al. [104] | MSC, Cardiomyocyte, EC | miR-23a-3p, miR-424, let-7f, miR-378, miR-214 | EV | ↑ | - | HOXA5, GAX, p38 MAPK, Smad2/3, ATM | Myocardial infarction |
| Gou et al. [105] | Cardiomyocyte | miR-19a-3p | EV | ↑ | - | HIF-1α | Myocardial infarction |
| Geng et al. [106] | Cardiomyocyte | miR-143 | EV | - | ↓ | IGF-IR/NO | Myocardial infarction |
| Liao et al. [107] | CT | miR-21-5p | EV | ↑ | - | Cdip1/caspase-3 | Myocardial infarction |
| Li et al. [108] | BMSC | lncRNA HCP5 | EV | ↑ | - | miR-497/IGF1/PI3K/AKT | Ischemia-reperfusion injury |
| Diao et al. [109] | hUCMSC | lncRNA UCA1 | EV | ↑ | - | miR-143/Bcl-2/Beclin-1 | Ischemia-reperfusion injury |
| Wang et al. [110] | Cardiomyocyte | circ-HIPK3 | EV | ↑ | - | miR-29a/VEGFA | Myocardial infarction |
| Wang et al. [111] | M2-like macrophage | circRNA-UB3A | EV | ↑ | - | miR-138-5p/RhoC | Myocardial fibrosis after myocardial infarction |
| Zhang et al. [112] | - | miR-483 | - | - | ↓ | TGF-β, TGFBR2, β-catenin, CTGF, IL-1β, ET-1 | Hypertension |
| Shang et al. [113] | - | miR-483-3p | EV | ↑ | - | TGF-β, CTGF, ACE1, ET-1 | Hypertension |
| Niu et al. [116] | - | miR-634 | - | - | ↓ | Wnt/β-catenin | Hypertension |
| Xiang et al. [119] | Cardiomyocyte | miR-217 | EV | ↑ | - | PTEN | Cardiac hypertrophy |
| Claudia et al. [120] | Cardiac fibroblast | miR-21-3p | EV | ↑ | - | SH3 domain-containing protein 2 and PDZ and LIM domain 5 | Cardiac hypertrophy |
| Thum et al. [121] | - | miR-21 | - | ↑ | - | ERK-MAPK and Spry1 | Cardiac hypertrophy |
| Yu et al. [125] | Macrophage | miR-155 | EV | ↑ | - | Son of Sevenless 1 and Suppressor of Cytokine Signaling 1 | Cardiac hypertrophy |
| Yang et al. [129] | Telocyte | miR-30b | EV | ↑ | - | Runx2/Wnt/β-catenin | Cardiac hypertrophy |
| Li et al. [130] | Cardiac stromal cell | miR-21-5p | EV | ↑ | - | phosphatase and tensin homolog/Akt | HF |
| Li et al. [131,132,133] | - | lncExACT1 | - | ↑ | - | DCHS2 | HF |
5. Clinical Developments and Applications of Ex-ncRNAs in CVDs
5.1. Ex-ncRNAs as Biomarkers
5.2. Ex-ncRNAs as Therapeutics Targets
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, N.R.C.; Ordunez, P.; Giraldo, G.; Rodriguez Morales, Y.A.; Lombardi, C.; Khan, T.; Padwal, R.; Tsuyuki, R.T.; Varghese, C. WHO HEARTS: A Global Program to Reduce Cardiovascular Disease Burden: Experience Implementing in the Americas and Opportunities in Canada. Can. J. Cardiol. 2021, 37, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Şahin, B.; İlgün, G. Risk factors of deaths related to cardiovascular diseases in World Health Organization (WHO) member countries. Health Soc. Care Community 2022, 30, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Dai, X. Emerging Roles of Extracellular Non-Coding RNAs in Vascular Diseases. J. Cardiovasc. Transl. Res. 2022, 15, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.G.; Gray, E.; Heman-Ackah, S.M.; Mäger, I.; Talbot, K.; Andaloussi, S.E.; Wood, M.J.; Turner, M.R. Extracellular vesicles in neurodegenerative disease—Pathogenesis to biomarkers. Nat. Rev. Neurol. 2016, 12, 346–357. [Google Scholar] [CrossRef]
- Zijlstra, C.; Stoorvogel, W. Prostasomes as a source of diagnostic biomarkers for prostate cancer. J. Clin. Investig. 2016, 126, 1144–1151. [Google Scholar] [CrossRef]
- Gruner, H.N.; McManus, M.T. Examining the evidence for extracellular RNA function in mammals. Nat. Rev. Genet. 2021, 22, 448–458. [Google Scholar] [CrossRef]
- Lin, Z.; Lu, F.; Ma, X.; Xia, X.; Zou, F.; Jiang, J. Roles of circular RNAs in the pathogenesis of intervertebral disc degeneration (Review). Exp. Ther. Med. 2021, 22, 1221. [Google Scholar] [CrossRef]
- Sato-Kuwabara, Y.; Melo, S.A.; Soares, F.A.; Calin, G.A. The fusion of two worlds: Non-coding RNAs and extracellular vesicles--diagnostic and therapeutic implications (Review). Int. J. Oncol. 2015, 46, 17–27. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Cheng, X.M.; Wu, N.; Tao, Y.; Wang, X.N. Non-coding RNAs Regulate the Pathogenesis of Aortic Dissection. Front. Cardiovasc. Med. 2022, 9, 890607. [Google Scholar] [CrossRef]
- Li, G.; Manning, A.C.; Bagi, A.; Yang, X.; Gokulnath, P.; Spanos, M.; Howard, J.; Chan, P.P.; Sweeney, T.; Kitchen, R.; et al. Distinct Stress-Dependent Signatures of Cellular and Extracellular tRNA-Derived Small RNAs. Adv. Sci. 2022, 9, e2200829. [Google Scholar] [CrossRef] [PubMed]
- Brandão, B.B.; Guerra, B.A.; Mori, M.A. Shortcuts to a functional adipose tissue: The role of small non-coding RNAs. Redox Biol. 2017, 12, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Videira, R.F.; da Costa Martins, P.A. Non-coding RNAs in Cardiac Intercellular Communication. Front. Physiol. 2020, 11, 738. [Google Scholar] [CrossRef] [PubMed]
- Kondkar, A.A.; Abu-Amero, K.K. Utility of circulating microRNAs as clinical biomarkers for cardiovascular diseases. Biomed. Res. Int. 2015, 2015, 821823. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.J.F.; Biessen, E.A.L.; Hohl, M.; Weber, C.; van der Vorst, E.P.C.; Santovito, D. Small Things Matter: Relevance of MicroRNAs in Cardiovascular Disease. Front. Physiol. 2020, 11, 793. [Google Scholar] [CrossRef]
- Shang, X.; Fang, Y.; Xin, W.; You, H. The Application of Extracellular Vesicles Mediated miRNAs in Osteoarthritis: Current Knowledge and Perspective. J. Inflamm. Res. 2022, 15, 2583–2599. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Xia, W.; Zhu, X.-W.; Mo, X.-B.; Wu, L.-F.; Wu, J.; Guo, Y.-F.; Zeng, K.-Q.; Wang, M.-J.; Lin, X.; Qiu, Y.-H.; et al. Integrative multi-omics analysis revealed SNP-lncRNA-mRNA (SLM) networks in human peripheral blood mononuclear cells. Human Genet. 2017, 136, 451–462. [Google Scholar] [CrossRef]
- Robinson, E.K.; Covarrubias, S.; Carpenter, S. The how and why of lncRNA function: An innate immune perspective. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194419. [Google Scholar] [CrossRef]
- Sun, R.; He, X.Y.; Mei, C.; Ou, C.L. Role of exosomal long non-coding RNAs in colorectal cancer. World J. Gastrointest. Oncol. 2021, 13, 867–878. [Google Scholar] [CrossRef]
- Dragomir, M.; Chen, B.; Calin, G.A. Exosomal lncRNAs as new players in cell-to-cell communication. Transl. Cancer Res. 2018, 7, S243–S252. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Chen, W.; Li, X. Extracellular vesicle-transferred long noncoding RNAs in bladder cancer. Clin. Chim. Acta 2021, 516, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Wang, J.; Pan, S.; Zheng, L.; Wang, Z.-W.; Zhu, X. Nucleic acids and proteins carried by exosomes of different origins as potential biomarkers for gynecologic cancers. Mol. Ther.-Oncolytics 2021, 24, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Xu, Y.; Fang, S.; Ji, Y.; Lu, L.; Xu, W.; Qian, H.; Liang, Z.F. Circular RNA and Its Roles in the Occurrence, Development, Diagnosis of Cancer. Front. Oncol. 2022, 12, 845703. [Google Scholar] [CrossRef]
- Tang, X.; Ren, H.; Guo, M.; Qian, J.; Yang, Y.; Gu, C. Review on circular RNAs and new insights into their roles in cancer. Comput. Struct. Biotechnol. J. 2021, 19, 910–928. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, X.; Zhan, X.; Kang, S.; Liu, H.; Luo, Y.; Lin, L. Advance in circular RNA modulation effects of heart failure. Gene X 2020, 5, 100036. [Google Scholar] [CrossRef]
- Yan, F.; Xie, X.; Huo, Q.; Zhang, W.; Wu, T.; Daniyaer, D.; Shi, L. circ-CCND1 regulates the CCND1/P53/P21 pathway through sponging miR-138-5p in valve interstitial cells to aggravate aortic valve calcification. J. Physiol. Biochem. 2022, 78, 845–854. [Google Scholar] [CrossRef]
- E, S.; Costa, M.C.; Kurc, S.; Drożdż, A.; Cortez-Dias, N.; Enguita, F.J. The circulating non-coding RNA landscape for biomarker research: Lessons and prospects from cardiovascular diseases. Acta Pharmacol. Sin. 2018, 39, 1085–1099. [Google Scholar] [CrossRef]
- Shen, S.; Jiang, H.; Bei, Y.; Xiao, J.; Li, X. Long Non-Coding RNAs in Cardiac Remodeling. Cell. Physiol. Biochem. 2017, 41, 1830–1837. [Google Scholar] [CrossRef]
- Zhang, Y.; Ying, F.; Tian, X.; Lei, Z.; Li, X.; Lo, C.-Y.; Li, J.; Jiang, L.; Yao, X. TRPM2 Promotes Atherosclerotic Progression in a Mouse Model of Atherosclerosis. Cells 2022, 11, 1423. [Google Scholar] [CrossRef]
- Aryal, B.; Suárez, Y. Non-coding RNA regulation of endothelial and macrophage functions during atherosclerosis. Vascul. Pharmacol. 2019, 114, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Lund-Katz, S.; Phillips, M.C. High density lipoprotein structure-function and role in reverse cholesterol transport. Subcell. Biochem. 2010, 51, 183–227. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Buchan, R.J.; Cook, S.A. MicroRNA-223 regulates Glut4 expression and cardiomyocyte glucose metabolism. Cardiovasc. Res. 2010, 86, 410–420. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, S.; Ji-Yan, C.; Huang, C.; Li, J.; Cai, A.P.; Feng, Y.Q. Circulating miR-92a expression level in patients with essential hypertension: A potential marker of atherosclerosis. J. Hum. Hypertens. 2017, 31, 200–205. [Google Scholar] [CrossRef]
- Yang, G.; Lin, C. Long Noncoding RNA SOX2-OT Exacerbates Hypoxia-Induced Cardiomyocytes Injury by Regulating miR-27a-3p/TGFβR1 Axis. Cardiovasc. Ther. 2020, 2020, 2016259. [Google Scholar] [CrossRef]
- Tao, J.; Hu, Y. Diagnostic and prognostic significance of lncRNA SOX2-OT in patients with carotid atherosclerosis. BMC Cardiovasc. Disord. 2022, 22, 211. [Google Scholar] [CrossRef]
- Vilades, D.; Martinez-Camblor, P.; Ferrero-Gregori, A.; Bar, C.; Lu, D.; Xiao, K.; Vea, A.; Nasarre, L.; Sanchez Vega, J.; Leta, R.; et al. Plasma circular RNA hsa_circ_0001445 and coronary artery disease: Performance as a biomarker. FASEB J. 2020, 34, 4403–4414. [Google Scholar] [CrossRef]
- Moroni, F.; Gertz, Z.; Azzalini, L. Relief of Ischemia in Ischemic Cardiomyopathy. Curr. Cardiol. Rep. 2021, 23, 80. [Google Scholar] [CrossRef]
- Han, D.; Kang, S.-H.; Yoon, C.-H.; Youn, T.-J.; Chae, I.-H. Attenuation of ischemia–reperfusion injury by intracoronary chelating agent administration. Sci. Rep. 2022, 12, 2050. [Google Scholar] [CrossRef]
- Rios-Navarro, C.; Daghbouche-Rubio, N.; Gavara, J.; de Dios, E.; Perez, N.; Vila, J.M.; Chorro, F.J.; Ruiz-Sauri, A.; Bodi, V. Ischemia-reperfusion injury to coronary arteries: Comprehensive microscopic study after reperfused myocardial infarction. Ann. Anat. 2021, 238, 151785. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Yiang, G.T.; Liao, W.T.; Tsai, A.P.; Cheng, Y.L.; Cheng, P.W.; Li, C.Y.; Li, C.J. Current Mechanistic Concepts in Ischemia and Reperfusion Injury. Cell. Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Kuster, D.W.; Cardenas-Ospina, A.; Miller, L.; Liebetrau, C.; Troidl, C.; Nef, H.M.; Möllmann, H.; Hamm, C.W.; Pieper, K.S.; Mahaffey, K.W.; et al. Release kinetics of circulating cardiac myosin binding protein-C following cardiac injury. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H547–H556. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, R.; Hirashiki, A.; Cheng, X.W.; Yamada, T.; Shimazu, S.; Shinoda, N.; Okumura, T.; Takeshita, K.; Bando, Y.; Kondo, T.; et al. Usefulness of serum cardiac troponins T and I to predict cardiac molecular changes and cardiac damage in patients with hypertrophic cardiomyopathy. Int. Heart J. 2013, 54, 202–206. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mannu, G.S. The non-cardiac use and significance of cardiac troponins. Scott. Med. J. 2014, 59, 172–178. [Google Scholar] [CrossRef]
- Corsten, M.F.; Dennert, R.; Jochems, S.; Kuznetsova, T.; Devaux, Y.; Hofstra, L.; Wagner, D.R.; Staessen, J.A.; Heymans, S.; Schroen, B. Circulating MicroRNA-208b and MicroRNA-499 reflect myocardial damage in cardiovascular disease. Circ. Cardiovasc. Genet. 2010, 3, 499–506. [Google Scholar] [CrossRef]
- Chen, Z.; Yan, Y.; Wu, J.; Qi, C.; Liu, J.; Wang, J. Expression level and diagnostic value of exosomal NEAT1/miR-204/MMP-9 in acute ST-segment elevation myocardial infarction. IUBMB Life 2020, 72, 2499–2507. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Kirchner, B.; Meidert, A.S.; Brandes, F.; Lindemann, A.; Doose, G.; Doege, A.; Weidenhagen, R.; Reithmair, M.; Schelling, G.; et al. Detection of Atherosclerosis by Small RNA-Sequencing Analysis of Extracellular Vesicle Enriched Serum Samples. Front. Cell. Dev. Biol. 2021, 9, 729061. [Google Scholar] [CrossRef]
- Crouser, E.D.; Julian, M.W.; Bicer, S.; Ghai, V.; Kim, T.K.; Maier, L.A.; Gillespie, M.; Hamzeh, N.Y.; Wang, K. Circulating exosomal microRNA expression patterns distinguish cardiac sarcoidosis from myocardial ischemia. PLoS ONE 2021, 16, e0246083. [Google Scholar] [CrossRef]
- Chen, Y.; Ye, X.; Yan, F. MicroRNA 3113-5p is a novel marker for early cardiac ischemia/reperfusion injury. Diagn. Pathol. 2019, 14, 121. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Deng, Y.; Li, H. MicroRNA-223 Regulates Cardiac Fibrosis After Myocardial Infarction by Targeting RASA1. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1439–1454. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; You, X.; Xu, T.; Liu, Y.; Ren, Y.; Liu, S.; Wu, F.; Xu, Z.; Zou, L.; Wang, G. Screening and Function Analysis of MicroRNAs Involved in Exercise Preconditioning-Attenuating Pathological Cardiac Hypertrophy. Int. Heart J. 2018, 59, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.L.; Liu, X.Y.; Han, R.J.; Yuan, W.; Sun, K.; Zhong, J.C.; Yang, X.C. Circulating exosomal long non-coding RNAs in patients with acute myocardial infarction. J. Cell. Mol. Med. 2020, 24, 9388–9396. [Google Scholar] [CrossRef]
- Kishore, R.; Garikipati, V.N.S.; Gonzalez, C. Role of Circular RNAs in Cardiovascular Disease. J. Cardiovasc. Pharmacol. 2020, 76, 128–137. [Google Scholar] [CrossRef] [PubMed]
- DeGuire, J.; Clarke, J.; Rouleau, K.; Roy, J.; Bushnik, T. Blood pressure and hypertension. Health Rep. 2019, 30, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.N. High Blood Pressure. JAMA 2020, 324, 1254–1255. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.C.; Lang, N.N.; Touyz, R.M. Drug Treatment of Hypertension: Focus on Vascular Health. Drugs 2016, 76, 1529–1550. [Google Scholar] [CrossRef]
- Besler, C.; Urban, D.; Watzka, S.; Lang, D.; Rommel, K.P.; Kandolf, R.; Klingel, K.; Thiele, H.; Linke, A.; Schuler, G.; et al. Endomyocardial miR-133a levels correlate with myocardial inflammation, improved left ventricular function, and clinical outcome in patients with inflammatory cardiomyopathy. Eur. J. Heart Fail. 2016, 18, 1442–1451. [Google Scholar] [CrossRef]
- Cengiz, M.; Yavuzer, S.; Kılıçkıran Avcı, B.; Yürüyen, M.; Yavuzer, H.; Dikici, S.A.; Karataş, Ö.F.; Özen, M.; Özen, M.; Uzun, H.; et al. Circulating miR-21 and eNOS in subclinical atherosclerosis in patients with hypertension. Clin. Exp. Hypertens. 2015, 37, 643–649. [Google Scholar] [CrossRef]
- Zou, X.; Wang, J.; Chen, C.; Tan, X.; Huang, Y.; Jose, P.A.; Yang, J.; Zeng, C. Secreted Monocyte miR-27a, via Mesenteric Arterial Mas Receptor-eNOS Pathway, Causes Hypertension. Am. J. Hypertens. 2020, 33, 31–42. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamada, H.; Fujii, R.; Munetsuna, E.; Yamazaki, M.; Ando, Y.; Ohashi, K.; Ishikawa, H.; Mizuno, G.; Tsuboi, Y.; et al. Circulating microRNA-27a and -133a are negatively associated with incident hypertension: A five-year longitudinal population-based study. Biomarkers 2022, 27, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, H.; Xing, Y.; Ye, Y.; He, F.; Yin, Q.; Li, Y.; Shang, F.; Shyy, J.Y.; Yuan, Z.Y. Endothelial-derived extracellular microRNA-92a promotes arterial stiffness by regulating phenotype changes of vascular smooth muscle cells. Sci. Rep. 2022, 12, 344. [Google Scholar] [CrossRef]
- Corsello, S.M.; Paragliola, R.M. Evaluation and Management of Endocrine Hypertension During Pregnancy. Endocrinol. Metab. Clin. N. Am. 2019, 48, 829–842. [Google Scholar] [CrossRef]
- Shah, S.; Gupta, A. Hypertensive Disorders of Pregnancy. Cardiol. Clin. 2019, 37, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Cui, R.; Liu, C.; Zhang, Z. MicroRNA regulation of transthyretin in trophoblast biofunction and preeclampsia. Arch. Biochem. Biophys. 2019, 676, 108129. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ding, D. High miR-200a-3p expression has high diagnostic values for hypertensive disorders complicating pregnancy and predicts adverse pregnancy outcomes. BMC Pregnancy Childbirth 2022, 22, 490. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Coats, A.J.S.; Tsutsui, H.; Abdelhamid, C.M.; Adamopoulos, S.; Albert, N.; Anker, S.D.; Atherton, J.; Böhm, M.; Butler, J.; et al. Universal definition and classification of heart failure: A report of the Heart Failure Society of America, Heart Failure Association of the European Society of Cardiology, Japanese Heart Failure Society and Writing Committee of the Universal Definition of Heart Failure. Eur. J. Heart Fail. 2021, 23, 352–380. [Google Scholar] [CrossRef]
- Janjusevic, M.; Fluca, A.L.; Ferro, F.; Gagno, G.; D’Alessandra, Y.; Beltrami, A.P.; Sinagra, G.; Aleksova, A. Traditional and Emerging Biomarkers in Asymptomatic Left Ventricular Dysfunction-Promising Non-Coding RNAs and Exosomes as Biomarkers in Early Phases of Cardiac Damage. Int. J. Mol. Sci. 2021, 22, 4937. [Google Scholar] [CrossRef]
- Li, J.; Salvador, A.M.; Li, G.; Valkov, N.; Ziegler, O.; Yeri, A.; Yang Xiao, C.; Meechoovet, B.; Alsop, E.; Rodosthenous, R.S.; et al. Mir-30d Regulates Cardiac Remodeling by Intracellular and Paracrine Signaling. Circ. Res. 2021, 128, e1–e23. [Google Scholar] [CrossRef]
- Wu, T.; Chen, Y.; Du, Y.; Tao, J.; Zhou, Z.; Yang, Z. Serum Exosomal MiR-92b-5p as a Potential Biomarker for Acute Heart Failure Caused by Dilated Cardiomyopathy. Cell. Physiol. Biochem. 2018, 46, 1939–1950. [Google Scholar] [CrossRef]
- Huang, Z.P.; Ding, Y.; Chen, J.; Wu, G.; Kataoka, M.; Hu, Y.; Yang, J.H.; Liu, J.; Drakos, S.G.; Selzman, C.H.; et al. Long non-coding RNAs link extracellular matrix gene expression to ischemic cardiomyopathy. Cardiovasc. Res. 2016, 112, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Ounzain, S.; Micheletti, R.; Beckmann, T.; Schroen, B.; Alexanian, M.; Pezzuto, I.; Crippa, S.; Nemir, M.; Sarre, A.; Johnson, R.; et al. Genome-wide profiling of the cardiac transcriptome after myocardial infarction identifies novel heart-specific long non-coding RNAs. Eur. Heart J. 2015, 36, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Kumarswamy, R.; Bauters, C.; Volkmann, I.; Maury, F.; Fetisch, J.; Holzmann, A.; Lemesle, G.; de Groote, P.; Pinet, F.; Thum, T. Circulating long noncoding RNA, LIPCAR, predicts survival in patients with heart failure. Circ. Res. 2014, 114, 1569–1575. [Google Scholar] [CrossRef]
- Han, P.; Li, W.; Lin, C.H.; Yang, J.; Shang, C.; Nuernberg, S.T.; Jin, K.K.; Xu, W.; Lin, C.Y.; Lin, C.J.; et al. A long noncoding RNA protects the heart from pathological hypertrophy. Nature 2014, 514, 102–106. [Google Scholar] [CrossRef]
- Piersma, B.; Bank, R.A.; Boersema, M. Signaling in Fibrosis: TGF-β, WNT, and YAP/TAZ Converge. Front. Med. 2015, 2, 59. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics-challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Rohner, E.; Yang, R.; Foo, K.S.; Goedel, A.; Chien, K.R. Unlocking the promise of mRNA therapeutics. Nat. Biotechnol. 2022, 40, 1586–1600. [Google Scholar] [CrossRef]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting circular RNAs as a therapeutic approach: Current strategies and challenges. Signal Transduct. Target. Ther. 2021, 6, 185. [Google Scholar] [CrossRef]
- Das, S.; Shah, R.; Dimmeler, S.; Freedman, J.E.; Holley, C.; Lee, J.M.; Moore, K.; Musunuru, K.; Wang, D.Z.; Xiao, J.; et al. Noncoding RNAs in Cardiovascular Disease: Current Knowledge, Tools and Technologies for Investigation, and Future Directions: A Scientific Statement from the American Heart Association. Circ. Genom. Precis. Med. 2020, 13, e000062. [Google Scholar] [CrossRef]
- Burger, F.; Baptista, D.; Roth, A.; da Silva, R.F.; Montecucco, F.; Mach, F.; Brandt, K.J.; Miteva, K. NLRP3 Inflammasome Activation Controls Vascular Smooth Muscle Cells Phenotypic Switch in Atherosclerosis. Int. J. Mol. Sci. 2021, 23, 340. [Google Scholar] [CrossRef]
- Cao, R.Y.; Zhang, Y.; Feng, Z.; Liu, S.; Liu, Y.; Zheng, H.; Yang, J. The Effective Role of Natural Product Berberine in Modulating Oxidative Stress and Inflammation Related Atherosclerosis: Novel Insights Into the Gut-Heart Axis Evidenced by Genetic Sequencing Analysis. Front. Pharmacol. 2021, 12, 764994. [Google Scholar] [CrossRef]
- Shimada, B.K.; Yang, Y.; Zhu, J.; Wang, S.; Suen, A.; Kronstadt, S.M.; Jeyaram, A.; Jay, S.M.; Zou, L.; Chao, W. Extracellular miR-146a-5p Induces Cardiac Innate Immune Response and Cardiomyocyte Dysfunction. Immunohorizons 2020, 4, 561–572. [Google Scholar] [CrossRef]
- Preissner, K.T.; Fischer, S.; Deindl, E. Extracellular RNA as a Versatile DAMP and Alarm Signal That Influences Leukocyte Recruitment in Inflammation and Infection. Front. Cell. Dev. Biol. 2020, 8, 619221. [Google Scholar] [CrossRef]
- Shan, K.; Jiang, Q.; Wang, X.Q.; Wang, Y.N.; Yang, H.; Yao, M.D.; Liu, C.; Li, X.M.; Yao, J.; Liu, B.; et al. Role of long non-coding RNA-RNCR3 in atherosclerosis-related vascular dysfunction. Cell Death Dis. 2016, 7, e2248. [Google Scholar] [CrossRef]
- Yang, Z.; Liang, X.; Yang, L. Circular RNA circ_0001445 alleviates the ox-LDL-induced endothelial injury in human primary aortic endothelial cells through regulating ABCG1 via acting as a sponge of miR-208b-5p. Gen. Thorac. Cardiovasc. Surg. 2022, 70, 779–792. [Google Scholar] [CrossRef]
- Gao, X.F.; Wang, Z.M.; Chen, A.Q.; Wang, F.; Luo, S.; Gu, Y.; Kong, X.Q.; Zuo, G.F.; Jiang, X.M.; Ding, G.W.; et al. Plasma Small Extracellular Vesicle-Carried miRNA-501-5p Promotes Vascular Smooth Muscle Cell Phenotypic Modulation-Mediated In-Stent Restenosis. Oxidative Med. Cell. Longev. 2021, 2021, 6644970. [Google Scholar] [CrossRef]
- Simsekyilmaz, S.; Cabrera-Fuentes, H.A.; Meiler, S.; Kostin, S.; Baumer, Y.; Liehn, E.A.; Weber, C.; Boisvert, W.A.; Preissner, K.T.; Zernecke, A. Role of extracellular RNA in atherosclerotic plaque formation in mice. Circulation 2014, 129, 598–606. [Google Scholar] [CrossRef]
- Chiou, N.T.; Kageyama, R.; Ansel, K.M. Selective Export into Extracellular Vesicles and Function of tRNA Fragments during T Cell Activation. Cell Rep. 2018, 25, 3356–3370.e4. [Google Scholar] [CrossRef]
- Liu, S.; Chen, J.; Shi, J.; Zhou, W.; Wang, L.; Fang, W.; Zhong, Y.; Chen, X.; Chen, Y.; Sabri, A.; et al. M1-like macrophage-derived exosomes suppress angiogenesis and exacerbate cardiac dysfunction in a myocardial infarction microenvironment. Basic Res. Cardiol. 2020, 115, 22. [Google Scholar] [CrossRef]
- Long, R.; Gao, L.; Li, Y.; Li, G.; Qin, P.; Wei, Z.; Li, D.; Qian, C.; Li, J.; Yang, G. M2 macrophage-derived exosomes carry miR-1271-5p to alleviate cardiac injury in acute myocardial infarction through down-regulating SOX6. Mol. Immunol. 2021, 136, 26–35. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Yuan, J.; Gao, W.; Zhong, X.; Yao, K.; Lin, L.; Ge, J. Dendritic cell-derived exosomal miR-494-3p promotes angiogenesis following myocardial infarction. Int. J. Mol. Med. 2021, 47, 315–325, Corrigendum in Int. J. Mol. Med. 2022, 49, 5096. [Google Scholar] [CrossRef]
- Li, Q.; Xu, Y.; Lv, K.; Wang, Y.; Zhong, Z.; Xiao, C.; Zhu, K.; Ni, C.; Wang, K.; Kong, M.; et al. Small extracellular vesicles containing miR-486-5p promote angiogenesis after myocardial infarction in mice and nonhuman primates. Sci. Transl. Med. 2021, 13, eabb0202. [Google Scholar] [CrossRef]
- Cheng, H.; Chang, S.; Xu, R.; Chen, L.; Song, X.; Wu, J.; Qian, J.; Zou, Y.; Ma, J. Hypoxia-challenged MSC-derived exosomes deliver miR-210 to attenuate post-infarction cardiac apoptosis. Stem Cell Res. Ther. 2020, 11, 224. [Google Scholar] [CrossRef]
- Mao, C.; Li, D.; Zhou, E.; Gao, E.; Zhang, T.; Sun, S.; Gao, L.; Fan, Y.; Wang, C. Extracellular vesicles from anoxia preconditioned mesenchymal stem cells alleviate myocardial ischemia/reperfusion injury. Aging 2021, 13, 6156–6170. [Google Scholar] [CrossRef]
- Ou, H.; Teng, H.; Qin, Y.; Luo, X.; Yang, P.; Zhang, W.; Chen, W.; Lv, D.; Tang, H. Extracellular vesicles derived from microRNA-150-5p-overexpressing mesenchymal stem cells protect rat hearts against ischemia/reperfusion. Aging 2020, 12, 12669–12683. [Google Scholar] [CrossRef]
- Li, T.; Gu, J.; Yang, O.; Wang, J.; Wang, Y.; Kong, J. Bone Marrow Mesenchymal Stem Cell-Derived Exosomal miRNA-29c Decreases Cardiac Ischemia/Reperfusion Injury Through Inhibition of Excessive Autophagy via the PTEN/Akt/mTOR Signaling Pathway. Circ. J. 2020, 84, 1304–1311. [Google Scholar] [CrossRef]
- Zou, L.; Ma, X.; Wu, B.; Chen, Y.; Xie, D.; Peng, C. Protective effect of bone marrow mesenchymal stem cell-derived exosomes on cardiomyoblast hypoxia-reperfusion injury through the miR-149/let-7c/Faslg axis. Free Radic. Res. 2020, 54, 722–731. [Google Scholar] [CrossRef]
- Fu, D.L.; Jiang, H.; Li, C.Y.; Gao, T.; Liu, M.R.; Li, H.W. MicroRNA-338 in MSCs-derived exosomes inhibits cardiomyocyte apoptosis in myocardial infarction. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10107–10117. [Google Scholar] [CrossRef]
- Walravens, A.S.; Smolgovsky, S.; Li, L.; Kelly, L.; Antes, T.; Peck, K.; Quon, T.; Ibrahim, A.; Marbán, E.; Berman, B.; et al. Mechanistic and therapeutic distinctions between cardiosphere-derived cell and mesenchymal stem cell extracellular vesicle non-coding RNA. Sci. Rep. 2021, 11, 8666. [Google Scholar] [CrossRef]
- de Couto, G.; Gallet, R.; Cambier, L.; Jaghatspanyan, E.; Makkar, N.; Dawkins, J.F.; Berman, B.P.; Marbán, E. Exosomal MicroRNA Transfer Into Macrophages Mediates Cellular Postconditioning. Circulation 2017, 136, 200–214. [Google Scholar] [CrossRef]
- de Couto, G.; Jaghatspanyan, E.; DeBerge, M.; Liu, W.; Luther, K.; Wang, Y.; Tang, J.; Thorp, E.B.; Marbán, E. Mechanism of Enhanced MerTK-Dependent Macrophage Efferocytosis by Extracellular Vesicles. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 2082–2096. [Google Scholar] [CrossRef]
- Ibrahim, A.G.; Cheng, K.; Marbán, E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef]
- Ciullo, A.; Li, C.; Li, L.; Ungerleider, K.C.; Peck, K.; Marbán, E.; Ibrahim, A.G.E. Biodistribution of unmodified cardiosphere-derived cell extracellular vesicles using single RNA tracing. J. Extracell. Vesicles 2022, 11, e12178. [Google Scholar] [CrossRef]
- Moghiman, T.; Barghchi, B.; Esmaeili, S.A.; Shabestari, M.M.; Tabaee, S.S.; Momtazi-Borojeni, A.A. Therapeutic angiogenesis with exosomal microRNAs: An effectual approach for the treatment of myocardial ischemia. Heart Fail. Rev. 2021, 26, 205–213. [Google Scholar] [CrossRef]
- Gou, L.; Xue, C.; Tang, X.; Fang, Z. Inhibition of Exo-miR-19a-3p derived from cardiomyocytes promotes angiogenesis and improves heart function in mice with myocardial infarction via targeting HIF-1α. Aging 2020, 12, 23609–23618. [Google Scholar] [CrossRef]
- Geng, T.; Song, Z.Y.; Xing, J.X.; Wang, B.X.; Dai, S.P.; Xu, Z.S. Exosome Derived from Coronary Serum of Patients with Myocardial Infarction Promotes Angiogenesis through the miRNA-143/IGF-IR Pathway. Int. J. Nanomed. 2020, 15, 2647–2658. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, Y.; Duan, C.; Zhu, K.; Huang, R.; Zhao, H.; Hintze, M.; Pu, Q.; Yuan, Z.; Lv, L.; et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics 2021, 11, 268–291. [Google Scholar] [CrossRef]
- Li, K.S.; Bai, Y.; Li, J.; Li, S.L.; Pan, J.; Cheng, Y.Q.; Li, K.; Wang, Z.G.; Ji, W.J.; Zhou, Q.; et al. LncRNA HCP5 in hBMSC-derived exosomes alleviates myocardial ischemia reperfusion injury by sponging miR-497 to activate IGF1/PI3K/AKT pathway. Int. J. Cardiol. 2021, 342, 72–81. [Google Scholar] [CrossRef]
- Diao, L.; Zhang, Q. Transfer of lncRNA UCA1 by hUCMSCs-derived exosomes protects against hypoxia/reoxygenation injury through impairing miR-143-targeted degradation of Bcl-2. Aging 2021, 13, 5967–5985. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, R.; Shen, C.; Liu, W.; Yuan, J.; Li, C.; Deng, W.; Wang, Z.; Zhang, W.; Ge, J.; et al. Exosomal CircHIPK3 Released from Hypoxia-Induced Cardiomyocytes Regulates Cardiac Angiogenesis after Myocardial Infarction. Oxidative Med. Cell. Longev. 2020, 2020, 8418407. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Zhao, R.; Qiu, Z.; Shen, C.; Wang, Z.; Liu, W.; Zhang, W.; Ge, J.; Shi, B. CircUbe3a from M2 macrophage-derived small extracellular vesicles mediates myocardial fibrosis after acute myocardial infarction. Theranostics 2021, 11, 6315–6333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, Y.; Yan, X.; Chen, S.; He, M.; Lei, Y.; Zhang, J.; Gongol, B.; Gu, M.; Miao, Y.; et al. MicroRNA-483 amelioration of experimental pulmonary hypertension. EMBO Mol. Med. 2020, 12, e11303. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Guo, X.; Chen, Y.; Wang, C.; Gao, J.; Wen, E.; Lai, B.; Bai, L. Endothelial MicroRNA-483-3p Is Hypertension-Protective. Oxidative Med. Cell. Longev. 2022, 2022, 3698219. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, J.; Zhang, W.; Chen, Y.; Zhang, K.; Popescu, L.M.; Ma, X.; Lau, W.B.; Rong, R.; Yu, X.; et al. Signature microRNA expression profile of essential hypertension and its novel link to human cytomegalovirus infection. Circulation 2011, 124, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Sarrion, I.; Milian, L.; Juan, G.; Ramon, M.; Furest, I.; Carda, C.; Cortijo Gimeno, J.; Mata Roig, M. Role of circulating miRNAs as biomarkers in idiopathic pulmonary arterial hypertension: Possible relevance of miR-23a. Oxidative Med. Cell. Longev. 2015, 2015, 792846. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Sun, N.; Kong, L.; Xu, Y.; Kang, Y. miR-634 inhibits human vascular smooth muscle cell proliferation and migration in hypertension through Wnt4/β-catenin pathway. Front. Biosci. (Landmark. Ed.) 2021, 26, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.K.; Bhullar, S.K.; Elimban, V.; Dhalla, N.S. Oxidative Stress as A Mechanism for Functional Alterations in Cardiac Hypertrophy and Heart Failure. Antioxidants 2021, 10, 931. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Dong, A.; Mueller, P.; Caicedo, J.; Sutton, A.M.; Odetunde, J.; Barrick, C.J.; Klyachkin, Y.M.; Abdel-Latif, A.; Smyth, S.S. Coronary artery remodeling in a model of left ventricular pressure overload is influenced by platelets and inflammatory cells. PLoS ONE 2012, 7, e40196. [Google Scholar] [CrossRef]
- Nie, X.; Fan, J.; Li, H.; Yin, Z.; Zhao, Y.; Dai, B.; Dong, N.; Chen, C.; Wang, D.W. miR-217 Promotes Cardiac Hypertrophy and Dysfunction by Targeting PTEN. Mol. Ther. Nucleic Acids 2018, 12, 254–266. [Google Scholar] [CrossRef]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J. Clin. Investig. 2014, 124, 2136–2146. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Sayed, D.; Rane, S.; Lypowy, J.; He, M.; Chen, I.Y.; Vashistha, H.; Yan, L.; Malhotra, A.; Vatner, D.; Abdellatif, M. MicroRNA-21 targets Sprouty2 and promotes cellular outgrowths. Mol. Biol. Cell. 2008, 19, 3272–3282. [Google Scholar] [CrossRef] [PubMed]
- Tatsuguchi, M.; Seok, H.Y.; Callis, T.E.; Thomson, J.M.; Chen, J.F.; Newman, M.; Rojas, M.; Hammond, S.M.; Wang, D.Z. Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2007, 42, 1137–1141. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Gao, W.; Yao, K.; Ge, J. Roles of Exosomes Derived from Immune Cells in Cardiovascular Diseases. Front. Immunol. 2019, 10, 648. [Google Scholar] [CrossRef]
- Yu, H.; Qin, L.; Peng, Y.; Bai, W.; Wang, Z. Exosomes Derived from Hypertrophic Cardiomyocytes Induce Inflammation in Macrophages via miR-155 Mediated MAPK Pathway. Front. Immunol. 2020, 11, 606045. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, C.; Liu, L.; A, X.; Chen, B.; Li, Y.; Du, J. Macrophage-Derived mir-155-Containing Exosomes Suppress Fibroblast Proliferation and Promote Fibroblast Inflammation during Cardiac Injury. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 192–204. [Google Scholar] [CrossRef]
- Summerhill, V.I.; Moschetta, D.; Orekhov, A.N.; Poggio, P.; Myasoedova, V.A. Sex-Specific Features of Calcific Aortic Valve Disease. Int. J. Mol. Sci. 2020, 21, 5620. [Google Scholar] [CrossRef]
- Duan, C.; Cao, Z.; Tang, F.; Jian, Z.; Liang, C.; Liu, H.; Xiao, Y.; Liu, L.; Ma, R. miRNA-mRNA crosstalk in myocardial ischemia induced by calcified aortic valve stenosis. Aging 2019, 11, 448–466. [Google Scholar] [CrossRef]
- Yang, R.; Tang, Y.; Chen, X.; Yang, Y. Telocytes-derived extracellular vesicles alleviate aortic valve calcification by carrying miR-30b. ESC Heart Fail. 2021, 8, 3935–3946. [Google Scholar] [CrossRef]
- Qiao, L.; Hu, S.; Liu, S.; Zhang, H.; Ma, H.; Huang, K.; Li, Z.; Su, T.; Vandergriff, A.; Tang, J.; et al. microRNA-21-5p dysregulation in exosomes derived from heart failure patients impairs regenerative potential. J. Clin. Investig. 2019, 129, 2237–2250. [Google Scholar] [CrossRef]
- Li, H.; Trager, L.E.; Liu, X.; Hastings, M.H.; Xiao, C.; Guerra, J.; To, S.; Li, G.; Yeri, A.; Rodosthenous, R.; et al. lncExACT1 and DCHS2 Regulate Physiological and Pathological Cardiac Growth. Circulation 2022, 145, 1218–1233. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Lu, Y.W.; Lin, Z.; Huang, Z.P.; Liu, J.; Wang, Y.; Seok, H.Y.; Hu, X.; Ma, Q.; Li, K.; et al. Intercalated disc protein Xinβ is required for Hippo-YAP signaling in the heart. Nat. Commun. 2020, 11, 4666. [Google Scholar] [CrossRef] [PubMed]
- Moya, I.M.; Halder, G. Hippo-YAP/TAZ signalling in organ regeneration and regenerative medicine. Nat. Rev. Mol. Cell. Biol. 2019, 20, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Bezzerides, V.J.; Platt, C.; Lerchenmüller, C.; Paruchuri, K.; Oh, N.L.; Xiao, C.; Cao, Y.; Mann, N.; Spiegelman, B.M.; Rosenzweig, A. CITED4 induces physiologic hypertrophy and promotes functional recovery after ischemic injury. JCI Insight 2016, 1, 5620. [Google Scholar] [CrossRef]
- Li, K.; Rodosthenous, R.S.; Kashanchi, F.; Gingeras, T.; Gould, S.J.; Kuo, L.S.; Kurre, P.; Lee, H.; Leonard, J.N.; Liu, H.; et al. Advances, challenges, and opportunities in extracellular RNA biology: Insights from the NIH exRNA Strategic Workshop. JCI Insight 2018, 3, e98942. [Google Scholar] [CrossRef]
- Blanco-Dominguez, R.; Sanchez-Diaz, R.; de la Fuente, H.; Jimenez-Borreguero, L.J.; Matesanz-Marin, A.; Relano, M.; Jimenez-Alejandre, R.; Linillos-Pradillo, B.; Tsilingiri, K.; Martin-Mariscal, M.L.; et al. A Novel Circulating MicroRNA for the Detection of Acute Myocarditis. N. Engl. J. Med. 2021, 384, 2014–2027. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef]
- Crooke, S.T. Molecular Mechanisms of Antisense Oligonucleotides. Nucleic Acid Ther. 2017, 27, 70–77. [Google Scholar] [CrossRef]
- Shah, A.M.; Giacca, M. Small non-coding RNA therapeutics for cardiovascular disease. Eur. Heart J. 2022, 43, 4548–4561. [Google Scholar] [CrossRef]
- Haemmig, S.; Feinberg, M.W. Targeting LncRNAs in Cardiovascular Disease: Options and Expeditions. Circ. Res. 2017, 120, 620–623. [Google Scholar] [CrossRef]
- Robinson, E.L.; Port, J.D. Utilization and Potential of RNA-Based Therapies in Cardiovascular Disease. JACC Basic Transl. Sci. 2022, 7, 956–969. [Google Scholar] [CrossRef]
- Lieberman, J. Tapping the RNA world for therapeutics. Nat. Struct. Mol. Biol. 2018, 25, 357–364. [Google Scholar] [CrossRef]
- Egli, M.; Manoharan, M. Re-Engineering RNA Molecules into Therapeutic Agents. Acc. Chem. Res. 2019, 52, 1036–1047. [Google Scholar] [CrossRef]
- Baumann, V.; Winkler, J. miRNA-based therapies: Strategies and delivery platforms for oligonucleotide and non-oligonucleotide agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef]
- Johannes, L.; Lucchino, M. Current Challenges in Delivery and Cytosolic Translocation of Therapeutic RNAs. Nucleic Acid Ther. 2018, 28, 178–193. [Google Scholar] [CrossRef]
- Borden, A.; Kurian, J.; Nickoloff, E.; Yang, Y.; Troupes, C.D.; Ibetti, J.; Lucchese, A.M.; Gao, E.; Mohsin, S.; Koch, W.J.; et al. Transient Introduction of miR-294 in the Heart Promotes Cardiomyocyte Cell Cycle Reentry After Injury. Circ. Res. 2019, 125, 14–25. [Google Scholar] [CrossRef]
- Žarković, M.; Hufsky, F.; Markert, U.R.; Marz, M. The Role of Non-Coding RNAs in the Human Placenta. Cells 2022, 11, 1588. [Google Scholar] [CrossRef]
- Palazzo, C.; D’Alessio, A.; Tamagnone, L. Message in a Bottle: Endothelial Cell Regulation by Extracellular Vesicles. Cancers 2022, 14, 1969. [Google Scholar] [CrossRef]
- Sykaras, A.G.; Christofidis, K.; Politi, E.; Theocharis, S. Exosomes on Endometrial Cancer: A Biomarkers Treasure Trove? Cancers 2022, 14, 1733. [Google Scholar] [CrossRef]
- Samra, M.; Srivastava, K. Non-coding RNA and their potential role in cardiovascular diseases. Gene 2023, 851, 147011. [Google Scholar] [CrossRef]
- Davidson, S.M.; Padró, T.; Bollini, S.; Vilahur, G.; Duncker, D.J.; Evans, P.C.; Guzik, T.; Hoefer, I.E.; Waltenberger, J.; Wojta, J.; et al. Progress in cardiac research: From rebooting cardiac regeneration to a complete cell atlas of the heart. Cardiovasc. Res. 2021, 117, 2161–2174. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Arora, M. RNA Binding Proteins and Non-coding RNA’s in Cardiovascular Diseases. Adv. Exp. Med. Biol. 2020, 1229, 105–118. [Google Scholar] [CrossRef]


| Stage | Drug | Modification and Delivery | Target Gene | Disease | Comments |
|---|---|---|---|---|---|
| Clinical trials | AZD8601 | naked mRNA | VEGF-A | HF | Phase 2 |
| CDR132L | ASO | miR-321 | HF | Phase 2 | |
| IONIS-AGT-LRx | Antisense | Angiotensinogen | HF | Phase 2 | |
| Olezarsen | Antisense | ApoC-III | Lipid disorders | Phase 3 | |
| Olpasiran | siRNA | Lp(a) | Lipid disorders | Phase 2 | |
| Pelacarsen | Antisense | Lp(a) | Lipid disorders | Phase 3 | |
| Vupanorsen | Antisense | ANGPTL3 | Lipid disorders | Phase 2 | |
| FDA/EMA approved | Inclisiran | siRNA | PCSK9 | Lipid disorders | FDA approved |
| Volanesorsen | ASO | APO-C3 | Triglycerides | EMA approved |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiapaer, Z.; Li, C.; Yang, X.; Sun, L.; Chatterjee, E.; Zhang, L.; Lei, J.; Li, G. Extracellular Non-Coding RNAs in Cardiovascular Diseases. Pharmaceutics 2023, 15, 155. https://doi.org/10.3390/pharmaceutics15010155
Jiapaer Z, Li C, Yang X, Sun L, Chatterjee E, Zhang L, Lei J, Li G. Extracellular Non-Coding RNAs in Cardiovascular Diseases. Pharmaceutics. 2023; 15(1):155. https://doi.org/10.3390/pharmaceutics15010155
Chicago/Turabian StyleJiapaer, Zeyidan, Chengyu Li, Xinyu Yang, Lingfei Sun, Emeli Chatterjee, Lingying Zhang, Ji Lei, and Guoping Li. 2023. "Extracellular Non-Coding RNAs in Cardiovascular Diseases" Pharmaceutics 15, no. 1: 155. https://doi.org/10.3390/pharmaceutics15010155
APA StyleJiapaer, Z., Li, C., Yang, X., Sun, L., Chatterjee, E., Zhang, L., Lei, J., & Li, G. (2023). Extracellular Non-Coding RNAs in Cardiovascular Diseases. Pharmaceutics, 15(1), 155. https://doi.org/10.3390/pharmaceutics15010155