Inhibition of MicroRNA 6937 Delays Photoreceptor and Vision Loss in a Mouse Model of Retinitis Pigmentosa

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Animal Handling
2.2. Sample Collection
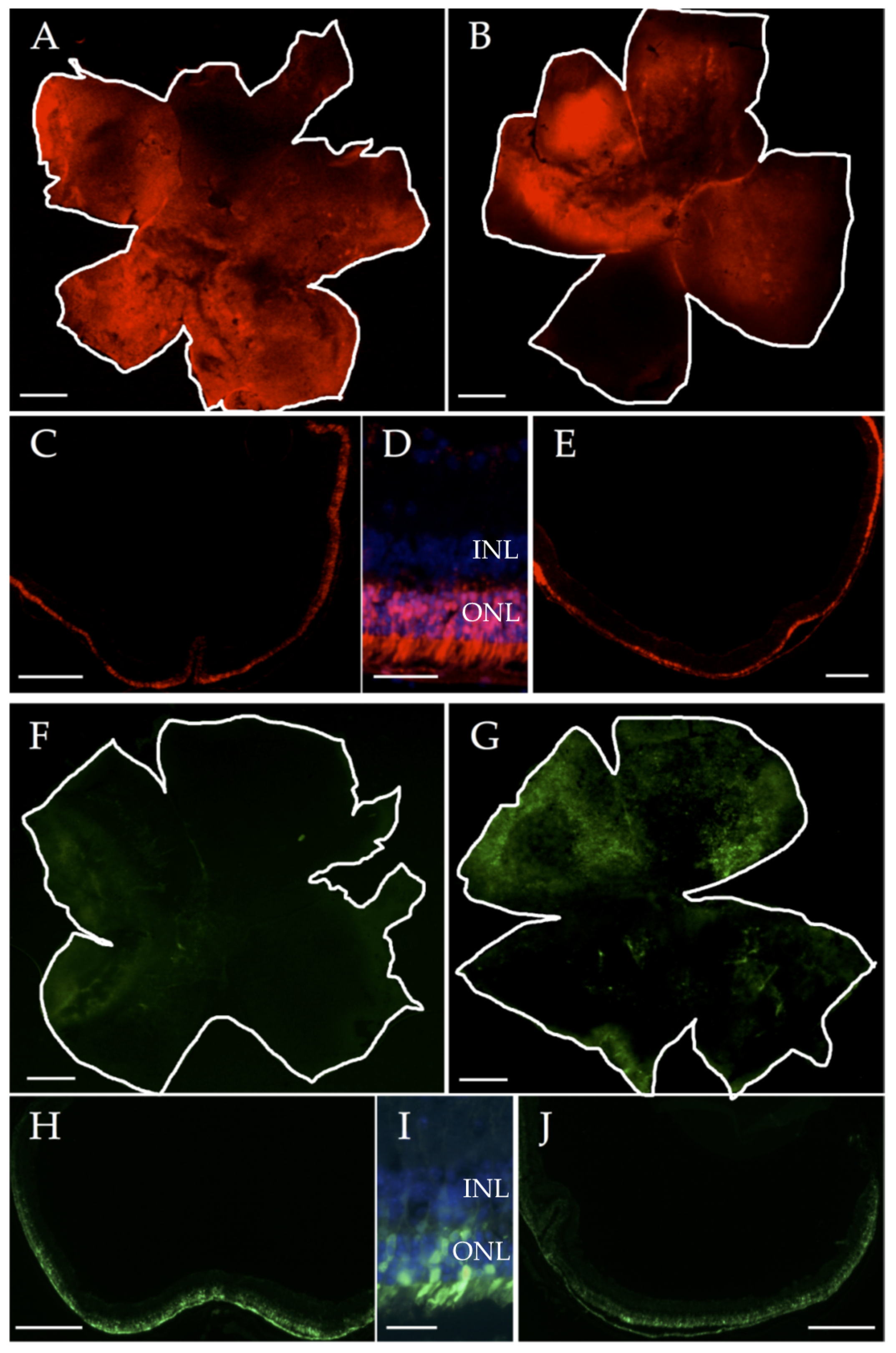
2.3. Generation of Adeno-Associated Viruses (AAVs) Expressing miRNA Modulators
2.4. Sub-Retinal Injections
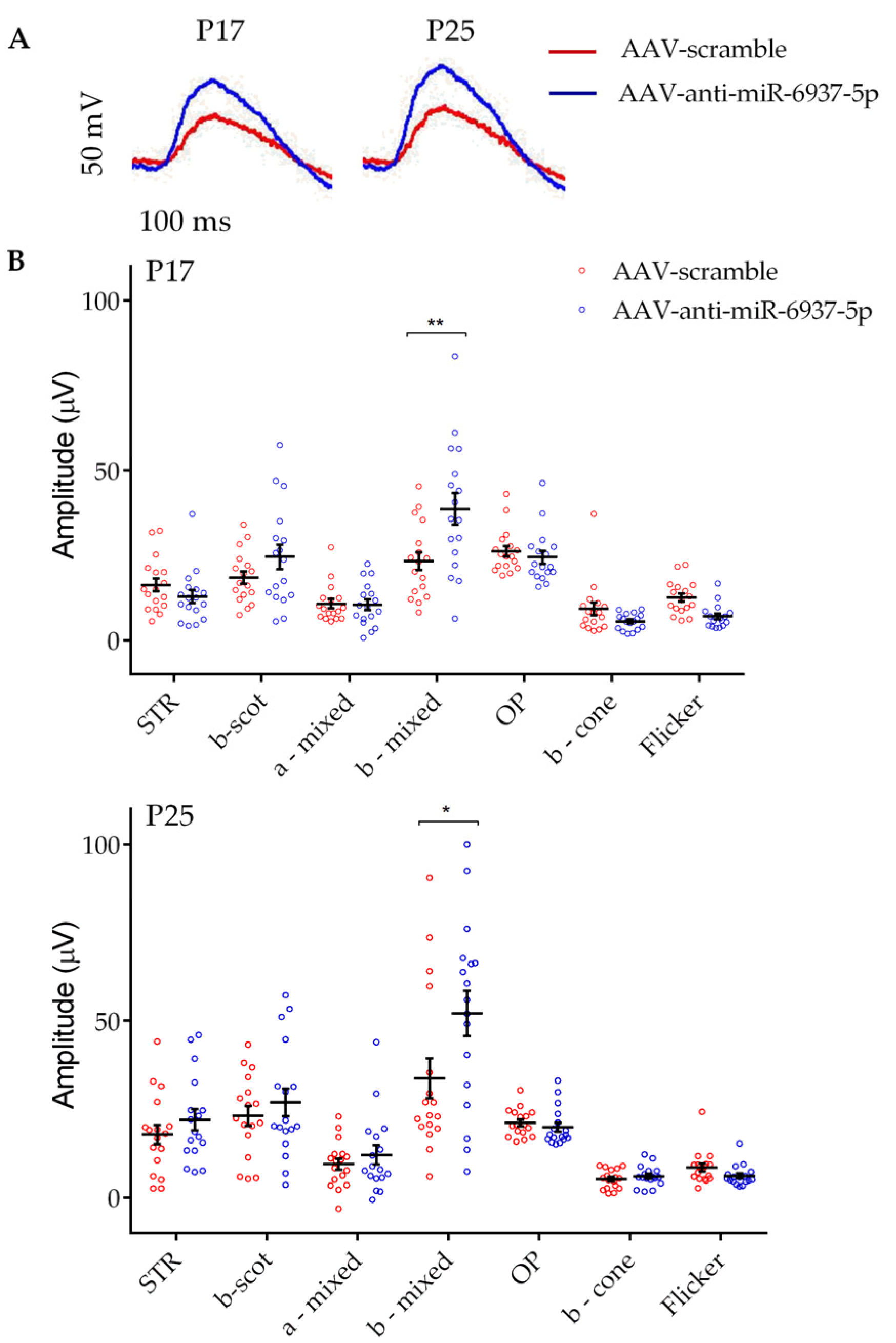
2.5. Electroretinogram Recordings
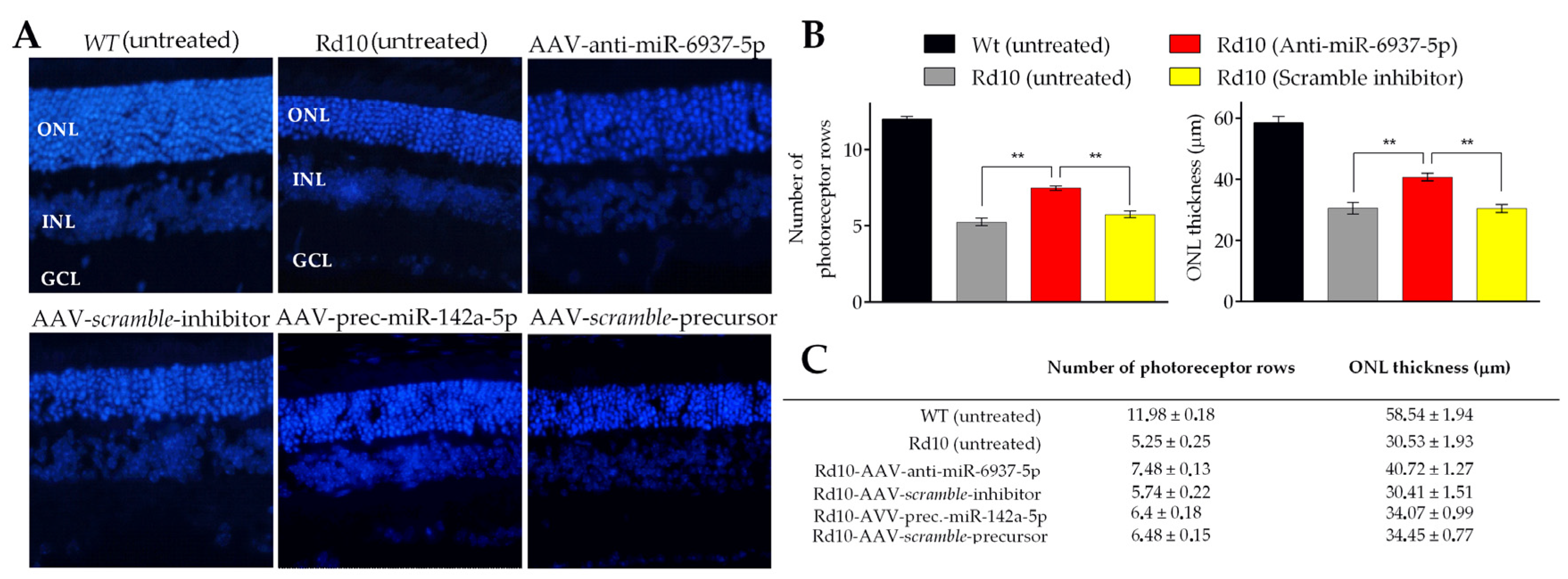
2.6. Histological Evaluation
2.7. Clariom S Arrays-Based Transcriptome Analysis
2.8. Validation of mRNA Expression by Quantitative PCR
2.9. miR-6937-5p In Vitro Modulation
2.10. miR-6937-5p In Vitro Induced Cell Viability and Cytotoxicity
2.11. Statistical Analysis
3. Results
3.1. miR-6937-5p Inhibition Slows Down Visual Loss in rd10 Mice
3.2. miR-6937-5p Inhibition Delays Photoreceptor Cell Loss in rd10 Mice
3.3. In Vivo Inhibition of miR-6937-5p in rd10 Mice Modulates the Expression of 101 Genes
3.4. Increased Cellular Cytotoxicity Induced by Over-Expression of miR-6937-5p in the MU-PH1 Photoreceptor-Like Cell Line
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- RetNet: Summaries. Available online: https://sph.uth.edu/retnet/sum-dis.htm (accessed on 21 September 2020).
- Donato, L.; Scimone, C.; Rinaldi, C.; Aragona, P.; Briuglia, S.; D’Ascola, A.; D’Angelo, R.; Sidoti, A. Stargardt phenotype associated with two ELOVL4 promoter variants and ELOVL4 downregulation: New possible perspective to etiopathogenesis? Investig. Ophthalmol. Vis. Sci. 2018, 59, 843–857. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, R.; Donato, L.; Venza, I.; Scimone, C.; Aragona, P.; Sidoti, A. Possible protective role of the ABCA4 gene c.1268A>G missense variant in Stargardt disease and syndromic retinitis pigmentosa in a Sicilian family: Preliminary data. Int. J. Mol. Med. 2017, 39, 1011–1020. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Scimone, C.; Donato, L.; Esposito, T.; Rinaldi, C.; D’Angelo, R.; Sidoti, A. A novel RLBP1 gene geographical area-related mutation present in a young patient with retinitis punctata albescens. Hum. Genom. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Le Meur, G.; Lebranchu, P.; Billaud, F.; Adjali, O.; Schmitt, S.; Bézieau, S.; Péréon, Y.; Valabregue, R.; Ivan, C.; Darmon, C.; et al. Safety and Long-Term Efficacy of AAV4 Gene Therapy in Patients with RPE65 Leber Congenital Amaurosis. Mol. Ther. 2018, 26, 256–268. [Google Scholar] [CrossRef]
- Wang, X.; Yu, C.; Tzekov, R.T.; Zhu, Y.; Li, W. The effect of human gene therapy for RPE65-associated Leber’s congenital amaurosis on visual function: A systematic review and meta-analysis. Orphanet J. Rare Dis. 2020, 15, 49. [Google Scholar] [CrossRef]
- Mellis, D.; Caporali, A. MicroRNA-based therapeutics in cardiovascular disease: Screening and delivery to the target. Biochem. Soc. Trans. 2018, 46, 11–21. [Google Scholar] [CrossRef]
- Kjaer-Frifeldt, S.; Hansen, T.F.; Nielsen, B.S.; Joergensen, S.; Lindebjerg, J.; Soerensen, F.B.; Depont Christensen, R.; Jakobsen, A. The prognostic importance of miR-21 in stage II colon cancer: A population-based study. Br. J. Cancer 2012, 107, 1169–1174. [Google Scholar] [CrossRef]
- Loginov, V.I.; Rykov, S.V.; Fridman, M.V.; Braga, E.A. Methylation of miRNA genes and oncogenesis. Biochem. 2015, 80, 145–162. [Google Scholar] [CrossRef]
- Bertoli, G.; Cava, C.; Castiglioni, I. MicroRNAs as Biomarkers for Diagnosis, Prognosis and Theranostics in Prostate Cancer. Int. J. Mol. Sci. 2016, 17, 421. [Google Scholar] [CrossRef]
- Cui, Z.; Zheng, X.; Kong, D. Decreased miR-198 expression and its prognostic significance in human gastric cancer. World J. Surg. Oncol. 2016, 14. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Balakathiresan, N.S.; Dalgard, C.; Gutti, U.; Armistead, D.; Jozwik, C.; Srivastava, M.; Pollard, H.B.; Biswas, R. Elevated miR-155 promotes inflammation in cystic fibrosis by driving hyperexpression of interleukin-8. J. Biol. Chem. 2011, 286, 11604–11615. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; Scimone, C.; Alibrandi, S.; Rinaldi, C.; Sidoti, A.; D’angelo, R. Transcriptome analyses of lncrnas in A2E-stressed retinal epithelial cells unveil advanced links between metabolic impairments related to oxidative stress and retinitis pigmentosa. Antioxidants 2020, 9, 318. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, M.; Darweesh, S.K.L.; de Looper, H.W.J.; van Luijn, M.M.; Hofman, A.; Ikram, M.A.; Franco, O.H.; Erkeland, S.J.; Dehghan, A. Genetic Variants in MicroRNAs and Their Binding Sites Are Associated with the Risk of Parkinson Disease. Hum. Mutat. 2016, 37, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Bhattacharyya, S.; Peters, K.W.; Glover, M.L.; Sen, A.; Cox, R.T.; Kundu, S.; Caohuy, H.; Frizzell, R.A.; Pollard, H.B.; et al. MIR-16 rescues F508del-CFTR function in native cystic fibrosis epithelial cells. Gene Ther. 2015, 22, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Taniguchi, F.; Nyuya, A.; Umeda, Y.; Mori, Y.; Fujiwara, T.; Tanioka, H.; Tsuruta, A.; Yamaguchi, Y.; Nagasaka, T. Upregulation of microRNA-31 is associated with poor prognosis in patients with advanced colorectal cancer. Oncol. Lett. 2020, 19, 2685–2694. [Google Scholar] [CrossRef]
- Luly, F.R.; Lévêque, M.; Licursi, V.; Cimino, G.; Martin-Chouly, C.; Théret, N.; Negri, R.; Cavinato, L.; Ascenzioni, F.; Del Porto, P. MiR-146a is over-expressed and controls IL-6 production in cystic fibrosis macrophages. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Muñoz-Culla, M.; Irizar, H.; Sáenz-Cuesta, M.; Castillo-Triviño, T.; Osorio-Querejeta, I.; Sepúlveda, L.; López De Munain, A.; Olascoaga, J.; Otaegui, D. SncRNA (microRNA &snoRNA) opposite expression pattern found in multiple sclerosis relapse and remission is sex dependent. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- van Rooij, E.; Marshall, W.S.; Olson, E.N. Toward microRNA-based therapeutics for heart disease: The sense in antisense. Circ. Res. 2008, 103, 919–928. [Google Scholar] [CrossRef]
- Xu, P.; Zhu, Y.; Sun, B.; Xiao, Z. Colorectal cancer characterization and therapeutic target prediction based on microRNA expression profile. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Sun, L.; Chen, X.; Jin, Z. Emerging roles of noncoding RNAs in retinal diseases: A review. Clin. Exp. Ophthalmol. 2020. [Google Scholar] [CrossRef]
- Mortuza, R.; Feng, B.; Chakrabarti, S. MiR-195 regulates SIRT1-mediated changes in diabetic retinopathy. Diabetologia 2014, 57, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Kaidonis, G.; Gillies, M.C.; Abhary, S.; Liu, E.; Essex, R.W.; Chang, J.H.; Pal, B.; Sivaprasad, S.; Pefkianaki, M.; Daniell, M.; et al. A single-nucleotide polymorphism in the MicroRNA-146a gene is associated with diabetic nephropathy and sight-threatening diabetic retinopathy in Caucasian patients. Acta Diabetol. 2016, 53, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J.; Surjyadipta, B.; Dua, P.; Alexandrov, P.N. Common micro RNAs (miRNAs) target complement factor H (CFH) regulation in Alzheimer’s disease (AD) and in age-related macular degeneration (AMD). Int. J. Biochem. Mol. Biol. 2012, 3, 105–116. [Google Scholar] [PubMed]
- Bhattacharjee, S.; Zhao, Y.; Dua, P.; Rogaev, E.I.; Lukiw, W.J. MicroRNA-34α-mediated down-regulation of the microglial-enriched triggering receptor and phagocytosis-sensor TREM2 in age-related macular degeneration. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Bai, S.; Tian, B.; Li, A.; Yao, Q.; Zhang, G.; Li, F. MicroRNA-125b promotes tumor growth and suppresses apoptosis by targeting DRAM2 in retinoblastoma. Eye 2016, 30, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Loscher, C.J.; Hokamp, K.; Wilson, J.H.; Li, T.; Humphries, P.; Farrar, G.J.; Palfi, A. A common microRNA signature in mouse models of retinal degeneration. Exp. Eye Res. 2008, 87, 529–534. [Google Scholar] [CrossRef]
- Loscher, C.J.; Hokamp, K.; Kenna, P.F.; Ivens, A.C.; Humphries, P.; Palfi, A.; Farrar, G.J. Altered retinal microRNA expression profile in a mouse model of retinitis pigmentosa. Genome Biol. 2007, 8. [Google Scholar] [CrossRef]
- Donato, L.; Bramanti, P.; Scimone, C.; Rinaldi, C.; D’Angelo, R.; Sidoti, A. miRNA expression profile of retinal pigment epithelial cells under oxidative stress conditions. FEBS Open Bio 2018, 8, 219–233. [Google Scholar] [CrossRef]
- Anasagasti, A.; Ezquerra-Inchausti, M.; Barandika, O.; Culla, M.M.; Caffarel, M.M.; Otaegui, D.; López de Munain, A.; Ruiz-Ederra, J. Expression profiling analysis reveals key Microrna-mRNA interactions in early retinal degeneration in retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2381–2392. [Google Scholar] [CrossRef]
- Ban, E.; Kwon, T.-H.; Kim, A. Delivery of therapeutic miRNA using polymer-based formulation. Drug Deliv. Transl. Res. 2019, 9, 1043–1056. [Google Scholar] [CrossRef]
- Ye, Y.; Li, Z.; Feng, Q.; Chen, Z.; Wu, Z.; Wang, J.; Ye, X.; Zhang, D.; Liu, L.; Gao, W.; et al. Downregulation of microRNA-145 may contribute to liver fibrosis in biliary atresia by targeting ADD3. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.P.; Xiong, Y.; Song, W.T.; Zhang, E.D.; Gao, Z.L.; Yao, F.; Su, T.; Zhou, R.R.; Xia, X.B. MicroRNA-28 potentially regulates the photoreceptor lineage commitment of Müller glia-derived progenitors. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Fernando, N.; Wong, J.H.C.; Das, S.; Dietrich, C.; Aggio-Bruce, R.; Cioanca, A.V.; Wooff, Y.; Chu-Tan, J.A.; Schumann, U.; Ngo, C.; et al. MicroRNA-223 Regulates Retinal Function and Inflammation in the Healthy and Degenerating Retina. Front. Cell Dev. Biol. 2020, 8, 516. [Google Scholar] [CrossRef] [PubMed]
- Murillo, O.; Luqui, D.M.; Gazquez, C.; Martinez-Espartosa, D.; Navarro-Blasco, I.; Monreal, J.I.; Guembe, L.; Moreno-Cermeño, A.; Corrales, F.J.; Prieto, J.; et al. Long-term metabolic correction of Wilson’s disease in a murine model by gene therapy. J. Hepatol. 2016, 64, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, S.; Barhoum, R.; Boya, P.; Arroba, A.I.; Rodríguez-Muela, N.; Gómez-Vicente, V.; Bosch, F.; De Pablo, F.; De La Villa, P.; De La Rosa, E.J. Attenuation of vision loss and delay in apoptosis of photoreceptors induced by proinsulin in a mouse model of retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4188–4194. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Vicente, V.; Flores, A.; Lax, P.; Murciano, C.; Yáñez, A.; Gil, M.L.; Cuenca, N.; Gozalbo, D.; Maneu, V. Characterization of a new murine retinal cell line (mu-ph1) with glial, progenitor and photoreceptor characteristics. Exp. Eye Res. 2013, 110, 125–135. [Google Scholar] [CrossRef]
- Toral-Ojeda, I.; Aldanondo, G.; Lasa-Elgarresta, J.; Lasa-Fernandez, H.; Vesga-Castro, C.; Mouly, V.; de Munain, A.L.; Vallejo-Illarramendi, A. A Novel Functional In Vitro Model that Recapitulates Human Muscle Disorders. In Muscle Cell and Tissue—Current Status of Research Field; InTech: London, UK, 2018. [Google Scholar]
- Peng, B.; Xiao, J.; Wang, K.; So, K.-F.; Tipoe, G.L.; Lin, B. Suppression of microglial activation is neuroprotective in a mouse model of human retinitis pigmentosa. J. Neurosci. 2014, 34, 8139–8150. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Donato, L.; D’Angelo, R.; Alibrandi, S.; Rinaldi, C.; Sidoti, A.; Scimone, C. Effects of A2E-induced oxidative stress on retinal epithelial cells: New insights on differential gene response and retinal dystrophies. Antioxidants 2020, 9, 307. [Google Scholar] [CrossRef]
- Donato, L.; Scimone, C.; Alibrandi, S.; Nicocia, G.; Rinaldi, C.; Sidoti, A.; D’Angelo, R. Discovery of glo1 new related genes and pathways by rna-seq on a2e-stressed retinal epithelial cells could improve knowledge on retinitis pigmentosa. Antioxidants 2020, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Palfi, A.; Hokamp, K.; Hauck, S.M.; Vencken, S.; Millington-Ward, S.; Chadderton, N.; Carrigan, M.; Kortvely, E.; Greene, C.M.; Kenna, P.F.; et al. MicroRNA regulatory circuits in a mouse model of inherited retinal degeneration. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fent, K. Progestins alter photo-transduction cascade and circadian rhythm network in eyes of zebrafish (Danio rerio). Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Zinn, E.; Pacouret, S.; Khaychuk, V.; Turunen, H.T.; Carvalho, L.S.; Andres-Mateos, E.; Shah, S.; Shelke, R.; Maurer, A.C.; Maurer, E.; et al. In silico reconstruction of the viral evolutionary lineage yields a potent gene therapy vector. Cell Rep. 2015, 12, 1056–1068. [Google Scholar] [CrossRef]
- Adighibe, O.; Pezzella, F. The Role of JMY in p53 Regulation. Cancers 2018, 10, 173. [Google Scholar] [CrossRef]
- Kajkowski, E.M.; Lo, C.F.; Ning, X.; Walker, S.; Sofia, H.J.; Wang, W.; Edris, W.; Chanda, P.; Wagner, E.; Vile, S.; et al. β-Amyloid Peptide-induced Apoptosis Regulated by a Novel Protein Containing a G Protein Activation Module. J. Biol. Chem. 2001, 276, 18748–18756. [Google Scholar] [CrossRef]
- Lee, Y.; Chang, D.J.; Lee, Y.S.; Chang, K.A.; Kim, H.; Yoon, J.S.; Lee, S.; Suh, Y.H.; Kaang, B.K. β-amyloid peptide binding protein does not couple to G protein in a heterologous Xenopus expression system. J. Neurosci. Res. 2003, 73, 255–259. [Google Scholar] [CrossRef]
- Chen, H.A.; Kuo, T.C.; Tseng, C.F.; Ma, J.T.; Yang, S.T.; Yen, C.J.; Yang, C.Y.; Sung, S.Y.; Su, J.L. Angiopoietin-like protein 1 antagonizes MET receptor activity to repress sorafenib resistance and cancer stemness in hepatocellular carcinoma. Hepatology 2016, 64, 1637–1651. [Google Scholar] [CrossRef]
- Gu, H.; Cui, M.; Bai, Y.; Chen, F.; Ma, K.; Zhou, C.; Guo, L. Angiopoietin-1/Tie2 signaling pathway inhibits lipopolysaccharide-induced activation of RAW264.7 macrophage cells. Biochem. Biophys. Res. Commun. 2010, 392, 178–182. [Google Scholar] [CrossRef]
- Yan, Q.; Jiang, L.; Liu, M.; Yu, D.; Zhang, Y.; Li, Y.; Fang, S.; Li, Y.; Zhu, Y.H.; Yuan, Y.F.; et al. ANGPTL1 interacts with integrin α1β1 to suppress HCC angiogenesis and metastasis by inhibiting JAK2/STAT3 signaling. Cancer Res. 2017, 77, 5831–5845. [Google Scholar] [CrossRef]
- Cartagena, C.M.; Schmid, K.E.; Phillips, K.L.; Tortella, F.C.; Dave, J.R. Changes in apoptotic mechanisms following penetrating ballistic-like brain injury. J. Mol. Neurosci. 2013, 49, 301–311. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ssAAV-Anc-CmiR0001 (scramble precursor) | 2.35 × 1012 p/mL |
| ssAAV-Anc-MmiR3437 (precursor miR-142a-5p) | 6.96 × 1011 vp/mL |
| ssAAV-Anc-CmiRAN0001 (scramble inhibitor) | 9.41 × 1011 vp/mL |
| ssAAV-Anc-MmiRAN2804 (miR-6240 inhibitor) | 7.90 × 1011 vp/mL |
| ssAAV-Anc-MmiRAN3970 (inhibitor miR-6937-5p) | 1.11× 1012 vp/mL |
| Gene Symbol | Fold-Change | Pathways | ||
|---|---|---|---|---|
| 6937 vs. SC | 6937 vs. C | SC vs. C | ||
| * Angptl1 | 1.5 | 1.67 | −1.12 | Akt signaling/Transmembrane receptor protein tyrosine kinase signaling pathway |
| * Bcl2a1d | 1.6 | 1.5 | 1.22 | Negative regulation of apoptotic process/T cell receptor signaling pathway |
| * Exosc6 | 1.62 | 1.58 | 1.02 | Negative regulation of neuron apoptotic process/rRNA processing |
| Ntng2 | 2.31 | 1.69 | 1.36 | Nervous system development/axonogenesis/cell differentiation |
| Cpne8 | 2.25 | 1.78 | 1.26 | Calcium-dependent membrane-binding |
| Cryba1 | 2.16 | 2.14 | 1.01 | Visual perception/camera-type eye development/lens development in camera-type eye |
| 1600002K03Rik | 2.1 | 1.73 | 1.22 | --- |
| Atxn2l | 2.06 | 1.78 | 1.16 | Regulation of cytoplasmic mRNA processing body assembly/stress granule assembly |
| Cldn5 | 2.03 | 2.39 | −1.18 | Single organismal cell-cell adhesion/myelination/calcium-independent cell-cell adhesion |
| Cog2 | 2.01 | 1.78 | 1.13 | Protein transport // intra-Golgi vesicle-mediated transport |
| Mapk1 | 2 | 1.65 | 1.21 | Apoptotic process/cellular response to DNA damage stimulus |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anasagasti, A.; Lara-López, A.; Milla-Navarro, S.; Escudero-Arrarás, L.; Rodríguez-Hidalgo, M.; Zabaleta, N.; González Aseguinolaza, G.; de la Villa, P.; Ruiz-Ederra, J. Inhibition of MicroRNA 6937 Delays Photoreceptor and Vision Loss in a Mouse Model of Retinitis Pigmentosa. Pharmaceutics 2020, 12, 913. https://doi.org/10.3390/pharmaceutics12100913
Anasagasti A, Lara-López A, Milla-Navarro S, Escudero-Arrarás L, Rodríguez-Hidalgo M, Zabaleta N, González Aseguinolaza G, de la Villa P, Ruiz-Ederra J. Inhibition of MicroRNA 6937 Delays Photoreceptor and Vision Loss in a Mouse Model of Retinitis Pigmentosa. Pharmaceutics. 2020; 12(10):913. https://doi.org/10.3390/pharmaceutics12100913
Chicago/Turabian StyleAnasagasti, Ander, Araceli Lara-López, Santiago Milla-Navarro, Leire Escudero-Arrarás, María Rodríguez-Hidalgo, Nerea Zabaleta, Gloria González Aseguinolaza, Pedro de la Villa, and Javier Ruiz-Ederra. 2020. "Inhibition of MicroRNA 6937 Delays Photoreceptor and Vision Loss in a Mouse Model of Retinitis Pigmentosa" Pharmaceutics 12, no. 10: 913. https://doi.org/10.3390/pharmaceutics12100913
APA StyleAnasagasti, A., Lara-López, A., Milla-Navarro, S., Escudero-Arrarás, L., Rodríguez-Hidalgo, M., Zabaleta, N., González Aseguinolaza, G., de la Villa, P., & Ruiz-Ederra, J. (2020). Inhibition of MicroRNA 6937 Delays Photoreceptor and Vision Loss in a Mouse Model of Retinitis Pigmentosa. Pharmaceutics, 12(10), 913. https://doi.org/10.3390/pharmaceutics12100913