
Immunization with Virus-Like Particle Vaccine Protects Rabbits against Hepatitis E-3 Virus Infection

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. VLP Vaccine and Virus
2.2. Animals, Immunization, and Challenge Schedule
2.3. HEV RNA PCR and Anti-HEV Total Antibody Analysis
2.4. Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) Levels Assessment
2.5. Cytokine Level Assessment Using ELISA
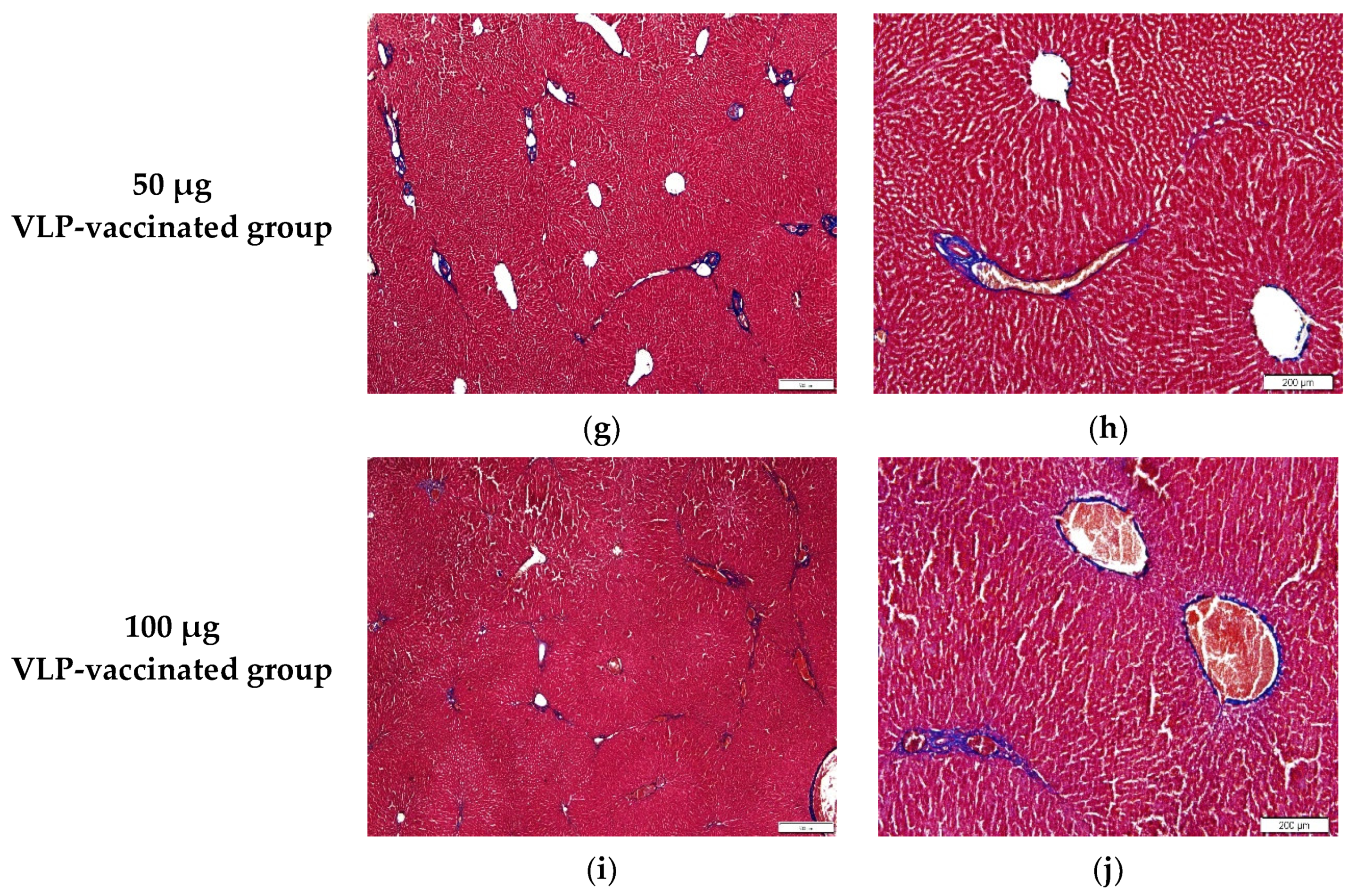
2.6. Histopathological Examination
2.7. Statistical Analyses
3. Results
3.1. HEV RNA Was Not Detected in the Liver, Serum, and Feces of Rabbits Vaccinated with 100 μg of VLP
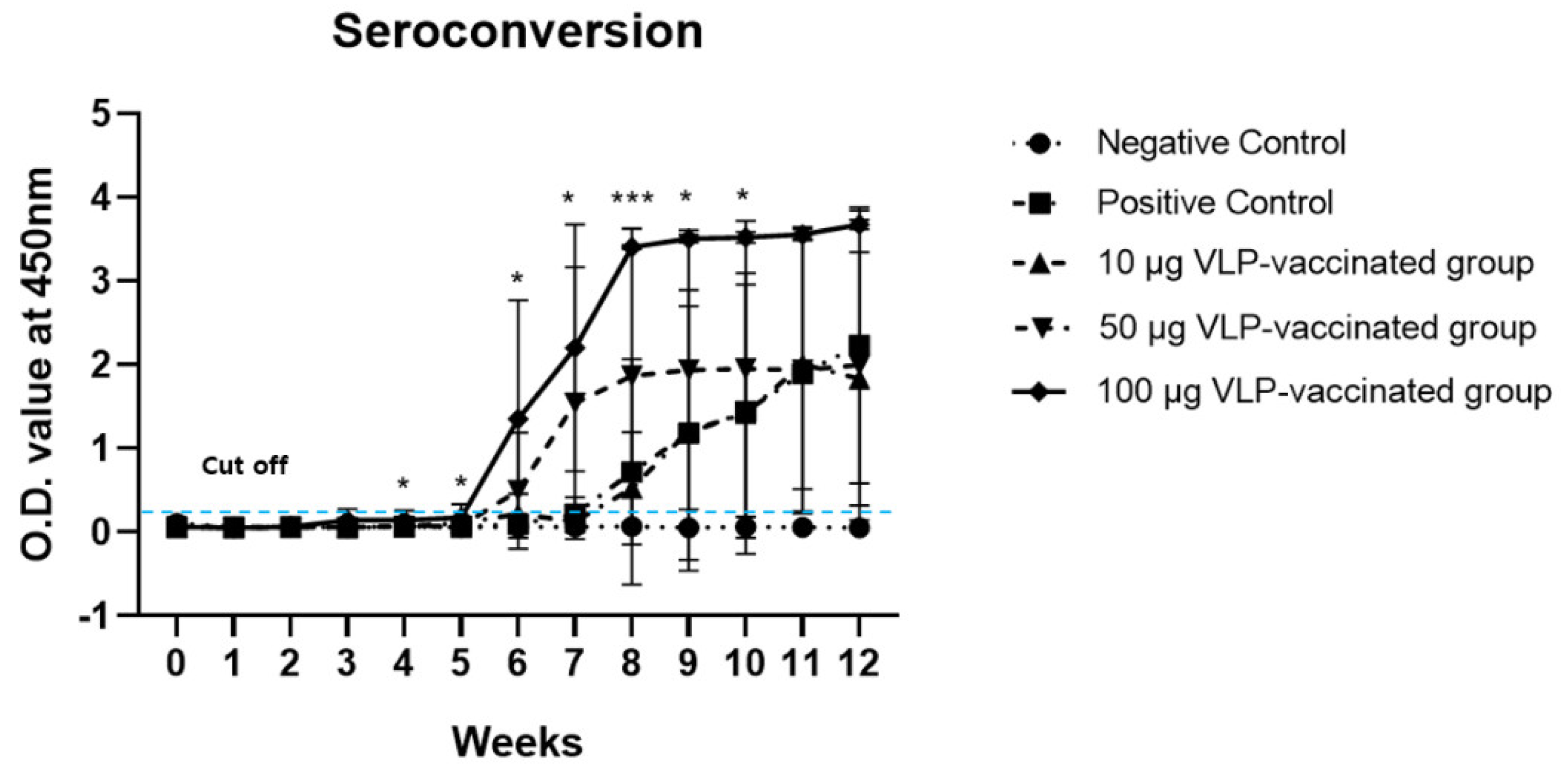
3.2. Anti-HEV Antibodies Were Produced after Vaccination with HEV-3-239-VLP in a Dose-Dependent Manner
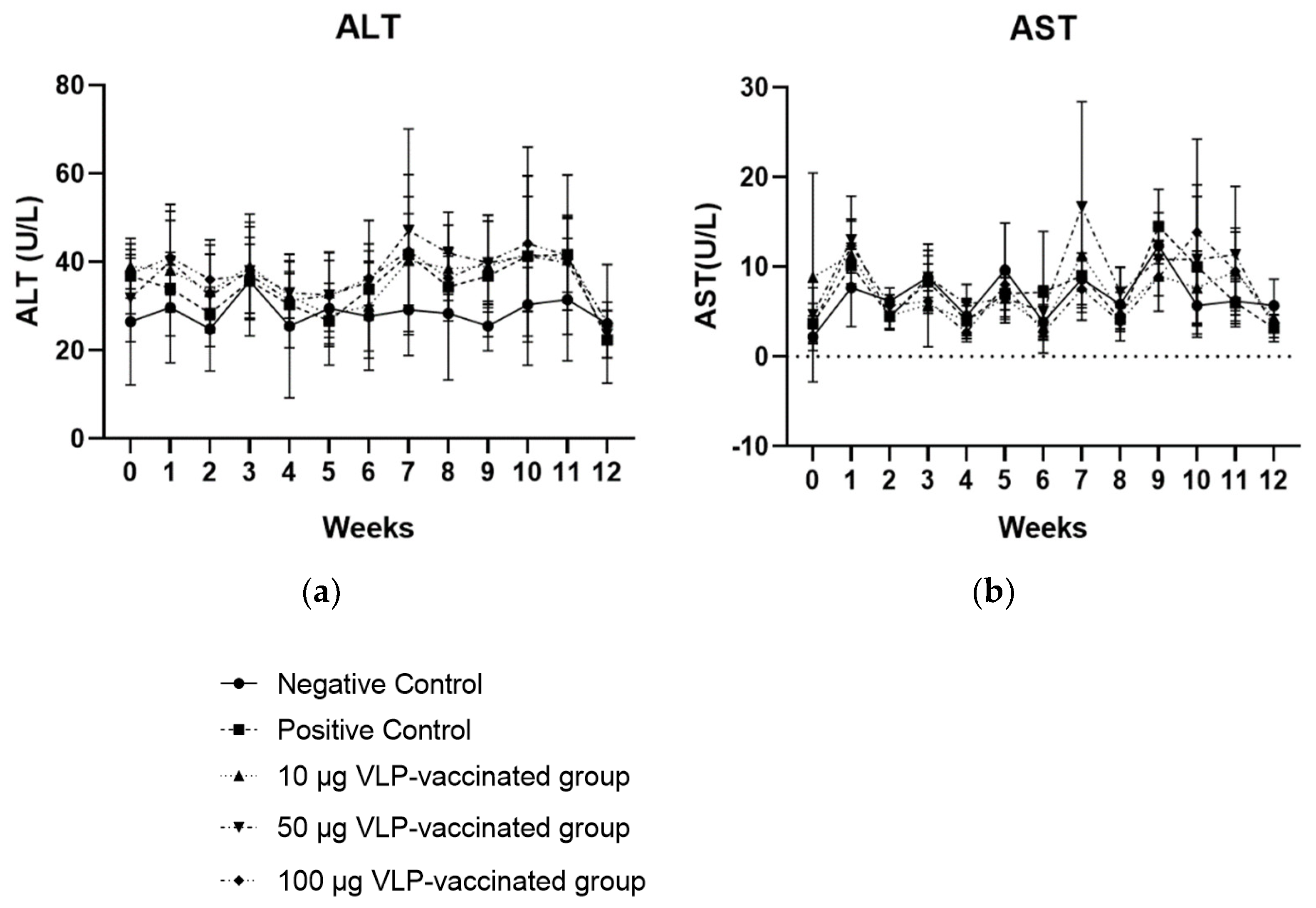
3.3. ALT and AST Levels of the Positive Control and Three Vaccinated Groups Were Not Significantly Higher than Those of the Negative Control Group
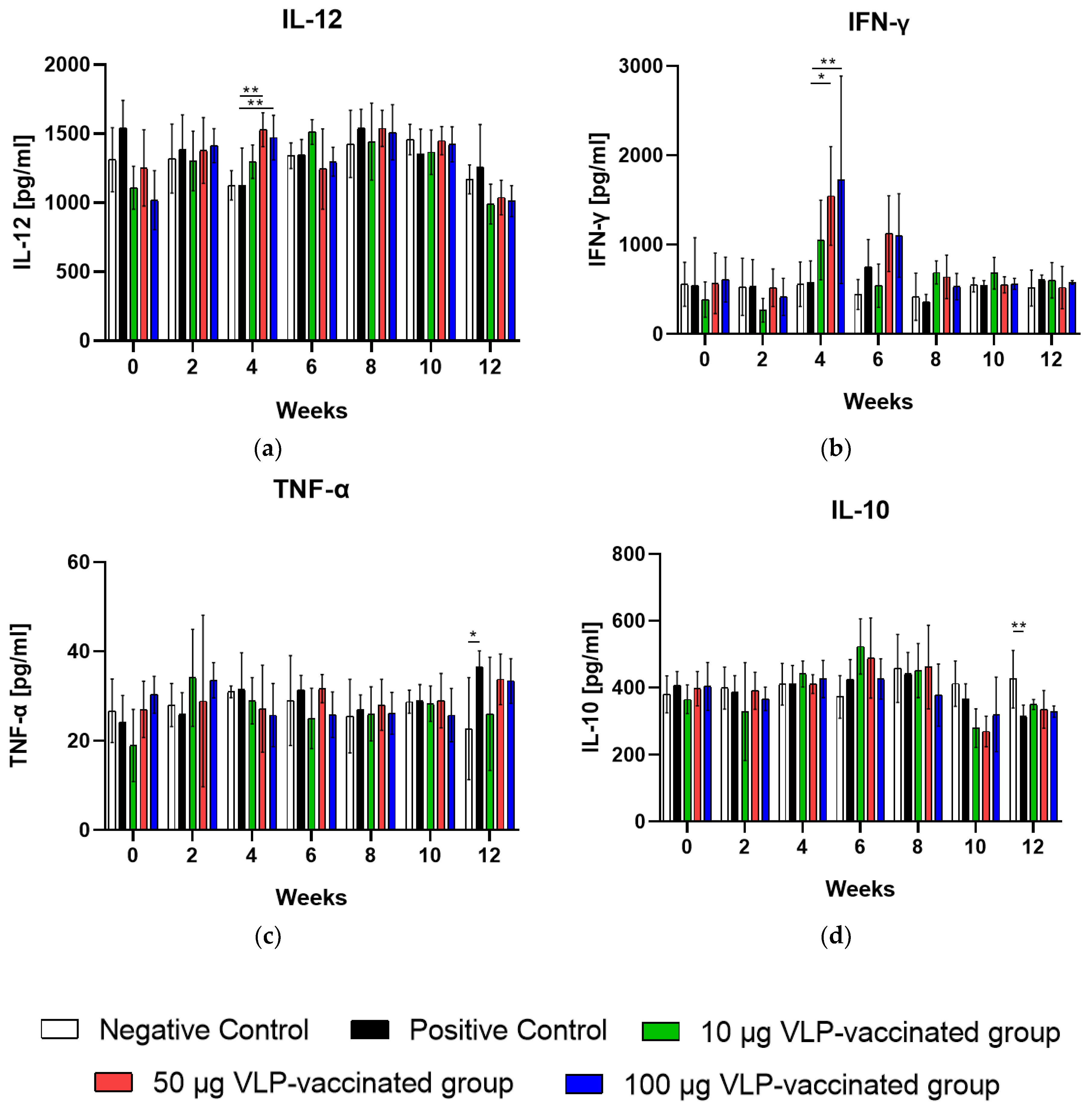
3.4. Serum IL-12 and IFN-γ Levels Increased after 50 and 100 μg VLP Administration
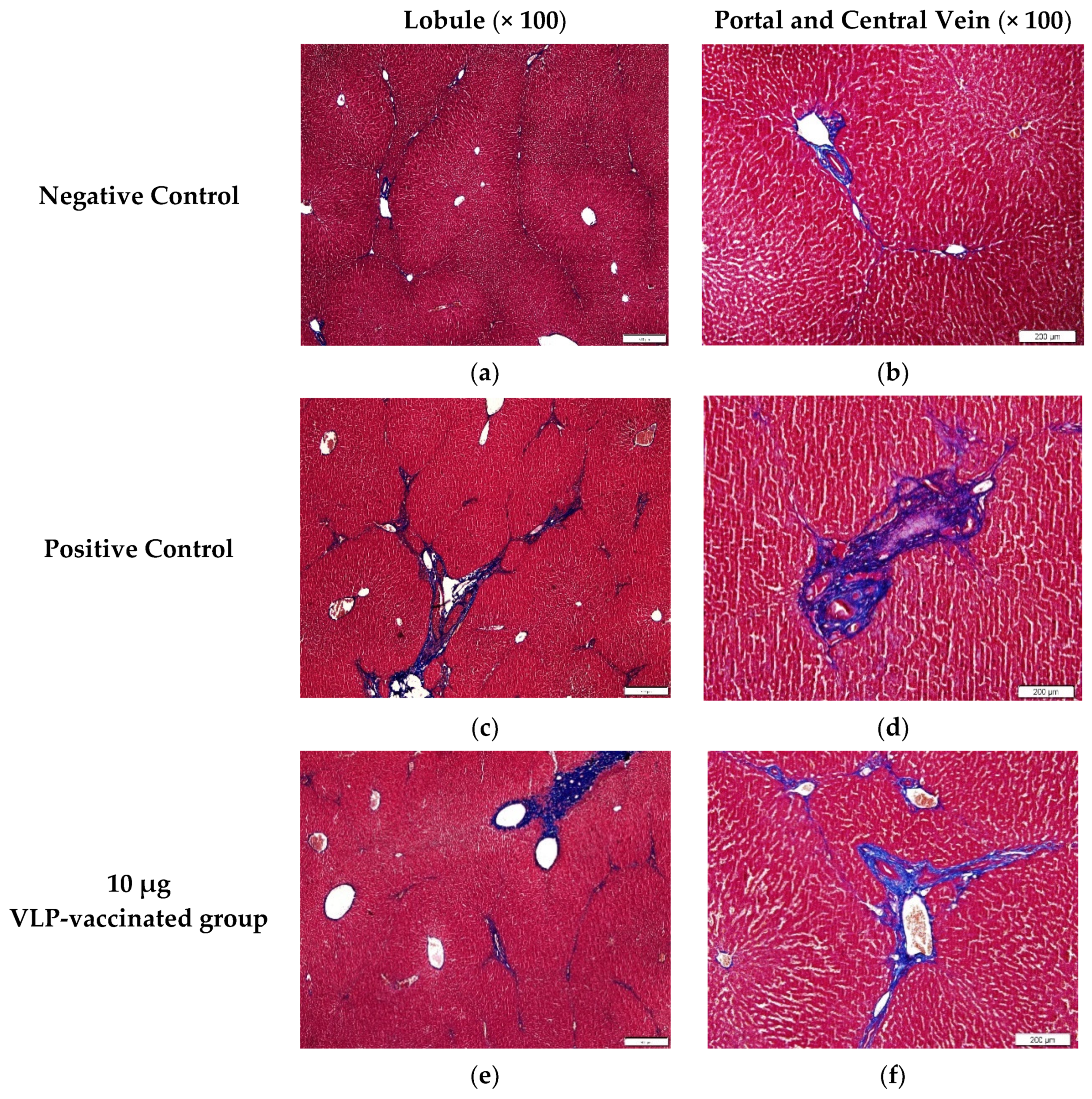
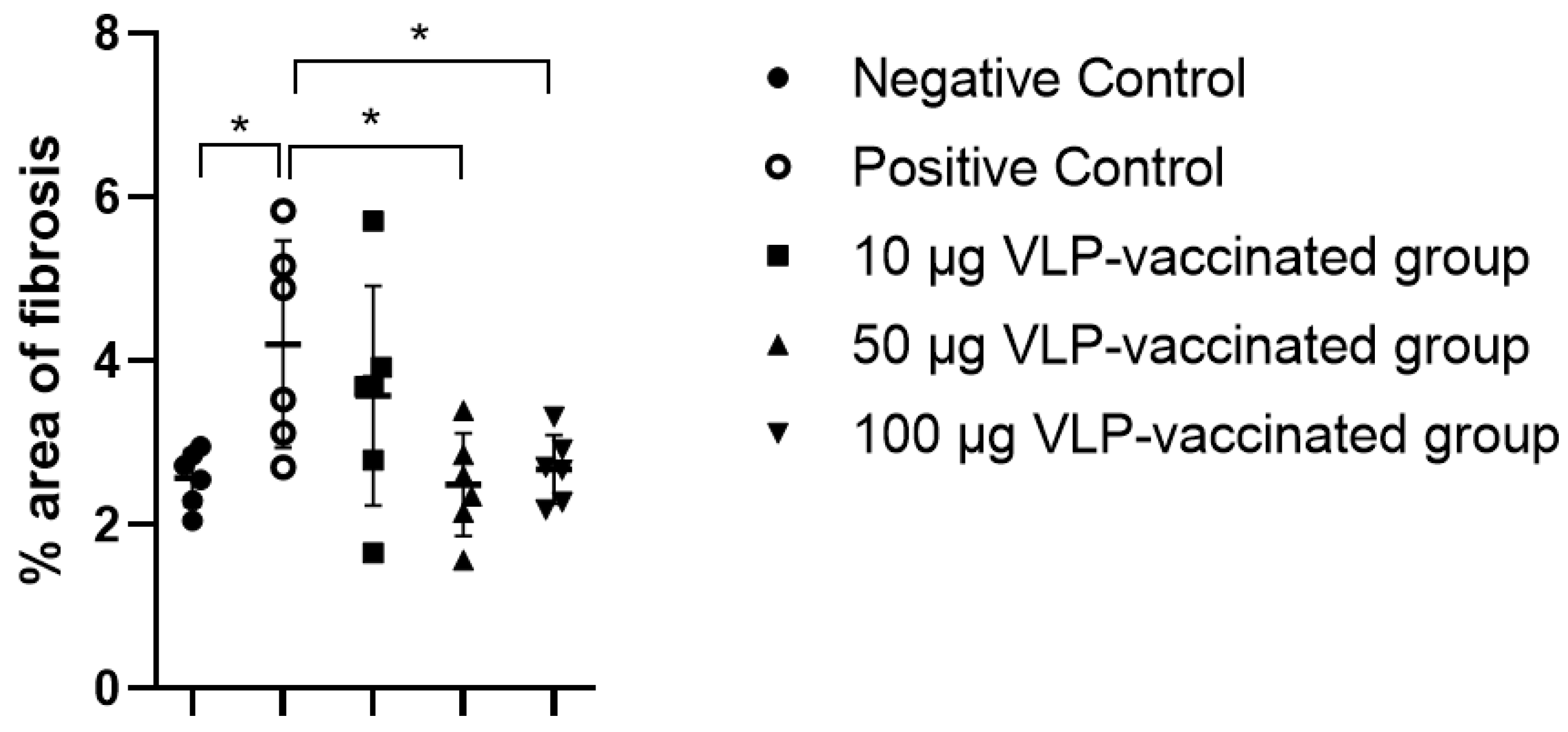
3.5. HEV-3-Induced Liver Fibrosis Was Prevented by 50 and 100 μg VLP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emerson, S.U.; Purcell, R.H. Hepatitis E virus. Rev. Med. Virol. 2003, 13, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Ticehurst, J.; Popkin, T.J.; Bryan, J.P.; Innis, B.L.; Duncan, J.F.; Ahmed, A.; Iqbal, M.; Malik, I.; Kapikian, A.Z.; Legters, L.J.; et al. Association of hepatitis E virus with an outbreak of hepatitis in Pakistan: Serologic responses and pattern of virus excretion. J. Med. Virol. 1992, 36, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692. [Google Scholar] [CrossRef]
- Nelson, K.E.; Labrique, A.B.; Kmush, B.L. Epidemiology of Genotype 1 and 2 Hepatitis E Virus Infections. Cold Spring Harb. Perspect. Med. 2019, 9, a031732. [Google Scholar] [CrossRef] [PubMed]
- Renou, C.; Afonso, A.M.R.; Pavio, N. Foodborne Transmission of Hepatitis E Virus from Raw Pork Liver Sausage, France. Emerg. Infect. Dis. 2014, 20, 1945. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Okada, K.; Takahashi, K.; Mishiro, S. Severe Hepatitis E Virus Infection after Ingestion of Uncooked Liver from a Wild Boar. J. Infect. Dis. 2003, 188, 944. [Google Scholar] [CrossRef]
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): Molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120–131. [Google Scholar] [CrossRef]
- Nan, Y.; Zhang, Y.J. Molecular biology and infection of hepatitis E virus. Front. Microbiol. 2016, 7, 1419. [Google Scholar] [CrossRef]
- Robinson, R.A.; Burgess, W.H.; Emerson, S.U.; Leibowitz, R.S.; Sosnovtseva, S.A.; Tsarev, S.; Purcell, R.H. Structural Characterization of Recombinant Hepatitis E Virus ORF2 Proteins in Baculovirus-Infected Insect Cells. Protein Expr. Purif. 1998, 12, 75–84. [Google Scholar] [CrossRef]
- Huang, Y.W.; Opriessnig, T.; Halbur, P.G.; Meng, X.J. Initiation at the Third In-Frame AUG Codon of Open Reading Frame 3 of the Hepatitis E Virus Is Essential for Viral Infectivity In Vivo. J. Virol. 2007, 81, 3018–3026. [Google Scholar] [CrossRef]
- Yamada, K.; Takahashi, M.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Nagashima, S.; Tanaka, T.; Okamato, H. ORF3 protein of hepatitis E virus is essential for virion release from infected cells. J. Gen. Virol. 2009, 90, 1880–1891. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Zhang, J.; Li, Y.M.; Ou, S.H.; Huang, G.Y.; He, Z.Q.; Ge, S.X.; Xian, Y.L.; Pang, S.Q.; Ng, M.H.; et al. A bacterially expressed particulate hepatitis E vaccine: Antigenicity, immunogenicity and protectivity on primates. Vaccine 2005, 23, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Zhang, J.; Zhang, X.F.; Zhou, C.; Wang, Z.Z.; Huang, S.J.; Wang, H.; Yang, C.L.; Jiang, H.M.; Cai, J.P.; et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: A large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895–902. [Google Scholar] [CrossRef]
- Zhang, M.; Emerson, S.U.; Nguyen, H.; Engle, R.; Govindarajan, S.; Blackwelder, W.C.; Gerin, J.; Purcell, R.H. Recombinant vaccine against hepatitis E: Duration of protective immunity in rhesus macaques. Vaccine 2002, 20, 3285–3291. [Google Scholar] [CrossRef]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M.; Mammen, M.P.; Thapa, G.B.; Thapa, N.; Myint, K.S.A.; Fourneau, M.; Kuschner, R.A.; Shrestha, S.K.; et al. Safety and Efficacy of a Recombinant Hepatitis E Vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.-b.; Li, R.-c.; Li, Y.-m.; Zheng, Y.-j.; Li, Y.-p.; Luo, D.; Pan, B.-b.; Nong, Y.; Ge, S.X.; et al. Randomized-controlled phase II clinical trial of a bacterially expressed recombinant hepatitis E vaccine. Vaccine 2009, 27, 1869–1874. [Google Scholar] [CrossRef]
- Hannig, G.; Makrides, S.C. Strategies for Optimizing Heterologous protein expression in Escherichia coli. Trends Biotechnol. 1998, 16, 54–60. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.; Li, L.; Liu, Z.; Wang, Z. Use of baculovirus expression system for generation of virus-like particles: Successes and challenges. Protein Expr. Purif. 2013, 90, 104–116. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.-F.; Huang, S.-J.; Wu, T.; Hu, Y.-M.; Wang, Z.-Z.; Wang, H.; Jiang, H.-M.; Wang, Y.-J.; Yan, Q.; et al. Long-Term Efficacy of a Hepatitis E Vaccine. N. Engl. J. Med. 2015, 372, 914–922. [Google Scholar] [CrossRef]
- Zhang, M.; Emerson, S.U.; Nguyen, H.; Engle, R.E.; Govindarajan, S.; Gerin, J.L.; Purcell, R.H. Immunogenicity and protective efficacy of a vaccine prepared from 53 kDa truncated hepatitis E virus capsid protein expressed in insect cells. Vaccine 2001, 20, 853–857. [Google Scholar] [CrossRef]
- Go, H.J.; Park, B.J.; Ahn, H.S.; Kim, D.H.; Kim, D.Y.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; et al. Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus. Vaccines 2021, 9, 1265. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, E.L.; Park, B.J.; Ahn, H.S.; Han, S.H.; Go, H.J.; Kim, D.H.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; et al. Detection and genetic analysis of zoonotic hepatitis E virus, rotavirus, and sapovirus in pigs. Korean J. Vet. Res. 2020, 60, 61–68. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, S.; Dai, X.; Shi, C.; Wen, Y.; Zhu, M.; Zhan, S.; Meng, J. Rabbit as a novel animal model for hepatitis E virus infection and vaccine evaluation. PLoS ONE 2012, 7, e51616. [Google Scholar] [CrossRef] [PubMed]
- Sanford, B.; Opriessnig, T.; Kenney, S.; Dryman, B.A.; Córdoba, L.; Meng, X.J. Assessment of the cross-protective capability of recombinant capsid proteins derived from pig, rat, and avian hepatitis E viruses (HEV) against challenge with a genotype 3 HEV in pigs. Vaccine 2012, 30, 6249–6255. [Google Scholar] [CrossRef]
- Mizuo, H.; Suzuki, K.; Takikawa, Y.; Sugai, Y.; Tokita, H.; Akahane, Y.; Itoh, K.; Gotanda, Y.; Takahashi, M.; Nishizawa, T.; et al. Polyphyletic strains of hepatitis E virus are responsible for sporadic cases of acute hepatitis in Japan. J. Clin. Microbiol. 2002, 40, 3209–3218. [Google Scholar] [CrossRef]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- The Global Prevalence of Hepatitis E Virus Infection and Susceptibility: A Systematic Review. Available online: https://apps.who.int/iris/handle/10665/70513 (accessed on 9 May 2022).
- Wedemeyer, H.; Pischke, S.; Manns, M.P. Pathogenesis and Treatment of Hepatitis E Virus Infection. Gastroenterology 2012, 142, 1388–1397.e1. [Google Scholar] [CrossRef]
- Todt, D.; Gisa, A.; Radonic, A.; Nitsche, A.; Behrendt, P.; Suneetha, P.V.; Pischke, S.; Bremer, B.; Brown, R.J.P.; Manns, M.P.; et al. In vivo evidence for ribavirin-induced mutagenesis of the hepatitis E virus genome. Gut 2016, 65, 1733–1743. [Google Scholar] [CrossRef]
- Lhomme, S.; Kamar, N.; Nicot, F.; Ducos, J.; Bismuth, M.; Garrigue, V.; Petitjean-Lecherbonnier, J.; Ollivier, I.; Alessandri-Gradt, E.; Goria, O.; et al. Mutation in the hepatitis E virus polymerase and outcome of ribavirin therapy. Antimicrob. Agents Chemother. 2016, 60, 1608–1614. [Google Scholar] [CrossRef]
- Zaman, K.; Dudman, S.; Stene-Johansen, K.; Qadri, F.; Yunus, M.; Sandbu, S.; Gurley, E.S.; Overbo, J.; Julin, C.H.; Dembinski, J.L.; et al. HEV study protocol: Design of a cluster-randomised, blinded trial to assess the safety, immunogenicity and effectiveness of the hepatitis E vaccine HEV 239 (Hecolin) in women of childbearing age in rural Bangladesh. BMJ Open 2020, 10, e033702. [Google Scholar] [CrossRef]
- Yousefi, Z.; Aria, H.; Ghaedrahmati, F.; Bakhtiari, T.; Azizi, M.; Bastan, R.; Hosseini, R.; Eskandari, N. An Update on Human Papilloma Virus Vaccines: History, Types, Protection, and Efficacy. Front. Immunol. 2022, 12, 805695. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Ahn, H.S.; Han, S.H.; Go, H.J.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Paik, H.J.; Choi, Y.K.; et al. Evaluation of the protective effects of a nanogel-based vaccine against rabbit hepatitis E virus. Vaccine 2019, 37, 5972–5978. [Google Scholar] [CrossRef] [PubMed]
- Han, S.H.; Park, B.J.; Ahn, H.S.; Kim, Y.H.; Go, H.J.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Choi, Y.K.; et al. Cross-Species Transmission of Swine Hepatitis E Virus Genotype 3 to Rabbits. Viruses 2020, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zheng, L.; Liu, Y.; Zhao, C.; Harrison, T.J.; Ma, Y.; Sun, S.; Zhang, J.; Wang, Y. Experimental Infection of Rabbits with Rabbit and Genotypes 1 and 4 Hepatitis E Viruses. PLoS ONE 2010, 5, e9160. [Google Scholar] [CrossRef]
- Wu, Q.; An, J.; She, R.; Shi, R.; Hao, W.; Soomro, M.H.; Yuan, X.; Yang, J.; Wang, J. Detection of Genotype 4 Swine Hepatitis E Virus in Systemic Tissues in Cross-Species Infected Rabbits. PLoS ONE 2017, 12, e0171277. [Google Scholar] [CrossRef] [PubMed]
- Taherkhani, R.; Farshadpour, F.; Makvandi, M.; Rajabi Memari, H.; Samarbafzadeh, A.R.; Sharifi, N.; Naeimi, B.; Tajbakhsh, S.; Akbarzadeh, S. Cytokine Profiles and Cell Proliferation Responses to Truncated ORF2 Protein in Iranian Patients Recovered from Hepatitis e Infection. J. Trop. Med. 2015, 2015, 523560. [Google Scholar] [CrossRef] [PubMed]
- Pape, G.R.; Gerlach, T.J.; Diepolder, H.M.; Grüner, N.; Jung, M.C.; Santantonio, T. Role of the specific T-cell response for clearance and control of hepatitis C virus. J. Viral Hepat. 1999, 6, 36–40. [Google Scholar] [CrossRef]
- Rossol, S.; Marinos, G.; Carucci, P.; Singer, M.V.; Williams, R.; Naoumov, N.V. Interleukin-12 induction of Th1 cytokines is important for viral clearance in chronic hepatitis B. J. Clin. Investig. 1997, 99, 3025–3033. [Google Scholar] [CrossRef]
- Stubblefield Park, S.R.; Widness, M.; Levine, A.D.; Patterson, C.E. T Cell-, Interleukin-12-, and Gamma Interferon-Driven Viral Clearance in Measles Virus-Infected Brain Tissue. J. Virol. 2011, 85, 3664–3676. [Google Scholar] [CrossRef][Green Version]
- Rubtsova, K.; Rubtsov, A.V.; Halemano, K.; Li, S.X.; Kappler, J.W.; Santiago, M.L.; Marrack, P. T Cell Production of IFNγ in Response to TLR7/IL-12 Stimulates Optimal B Cell Responses to Viruses. PLoS ONE 2016, 11, e0166322. [Google Scholar] [CrossRef]
- Ye, J.; Lv, L.; Wu, W.; Li, Y.; Shi, D.; Fang, D.; Guo, F.; Jiang, H.; Yan, R.; Ye, W.; et al. Butyrate Protects Mice Against Methionine–Choline-Deficient Diet-Induced Non-alcoholic Steatohepatitis by Improving Gut Barrier Function, Attenuating Inflammation and Reducing Endotoxin Levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.N.; Schreiner, P.; Ng, B.Y.Y.; Lo, B.; Hughes, M.R.; Scott, R.W.; Gusti, V.; Lecour, S.; Simonson, E.; Manisali, I.; et al. Impact of a CXCL12/CXCR4 antagonist in bleomycin (BLM) induced pulmonary fibrosis and carbon tetrachloride (CCl4) induced hepatic fibrosis in mice. PLoS ONE 2016, 11, e0151765. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Huang, S.; Zhu, N.; Liu, W.; Zhang, Y.; He, Y. Recombinant human histidine triad nucleotide-binding protein 1 attenuates liver fibrosis induced by carbon tetrachloride in rats. Mol. Med. Rep. 2013, 8, 1023–1028. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Péron, J.-M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E Virus and Chronic Hepatitis in Organ-Transplant Recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Gérolami, R.; Moal, V.; Colson, P. Chronic Hepatitis E with Cirrhosis in a Kidney-Transplant Recipient. N. Engl. J. Med. 2009, 358, 859–860. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Mansuy, J.M.; Cointault, O.; Selves, J.; Abravanel, F.; Danjoux, M.; Otal, P.; Esposito, L.; Durand, D.; Izopet, J.; et al. Hepatitis E Virus-Related Cirrhosis in Kidney-and Kidney–Pancreas-Transplant Recipients. Am. J. Transpl. 2008, 8, 1744–1748. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Abravanel, F.; Selves, J.; Garrouste, C.; Esposito, L.; Lavayssière, L.; Cointault, O.; Ribes, D.; Cardeau, I.; Nogier, M.B.; et al. Influence of immunosuppressive therapy on the natural history of genotype 3 hepatitis-e virus infection after organ transplantation. Transplantation 2010, 89, 353–360. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence (5′ to 3′) | Position in HEV-3 * (nt) | |
|---|---|---|
| Forward primer | GGKTRGAATGAATAACATGTY | 5122–5142 |
| Reverse primer | GCATAGGCARAARCACGA | 5198–5215 |
| Probe | CATCGCCCATGGGATCRCCATG | 5148–5169 |
| Number of Rabbits | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Weeks Post Inoculation | ||||||||||||||
| Sample | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | % | |
| Negative control | Serum | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 |
| Feces | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 | |
| Positive control | Serum | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 1/6 | 2/6 | 3/6 | 0/6 | 1/6 | 0/6 | 0/6 | 50 |
| Feces | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 1/6 | 3/6 | 3/6 | 0/6 | 0/6 | 0/6 | 82.3 | |
| 10 μg VLP- vaccinated group | Serum | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 |
| Feces | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 1/6 | 1/6 | 0/6 | 0/6 | 0/6 | 16.7 | |
| 50 μg VLP- vaccinated group | Serum | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 |
| Feces | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 | |
| 100 μg VLP- vaccinated group | Serum | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 |
| Feces | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0 | |
| Group | Number of Rabbits with HEV RNA in the Livers |
|---|---|
| Negative control | 0/6 (0%) |
| Positive control | 3/6 (50%) |
| 10 μg VLP-vaccinated group | 1/6 (16.7%) |
| 50 μg VLP-vaccinated group | 1/6 (16.7%) |
| 100 μg VLP-vaccinated group | 0/6 (0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, H.-J.; Park, B.-J.; Ahn, H.-S.; Han, S.-H.; Kim, D.-H.; Lyoo, E.-L.; Kim, D.-Y.; Kim, J.-H.; Lee, J.-B.; Park, S.-Y.; et al. Immunization with Virus-Like Particle Vaccine Protects Rabbits against Hepatitis E-3 Virus Infection. Viruses 2022, 14, 1432. https://doi.org/10.3390/v14071432
Go H-J, Park B-J, Ahn H-S, Han S-H, Kim D-H, Lyoo E-L, Kim D-Y, Kim J-H, Lee J-B, Park S-Y, et al. Immunization with Virus-Like Particle Vaccine Protects Rabbits against Hepatitis E-3 Virus Infection. Viruses. 2022; 14(7):1432. https://doi.org/10.3390/v14071432
Chicago/Turabian StyleGo, Hyeon-Jeong, Byung-Joo Park, Hee-Seop Ahn, Sang-Hoon Han, Dong-Hwi Kim, Eu-Lim Lyoo, Da-Yoon Kim, Jae-Hyeong Kim, Joong-Bok Lee, Seung-Yong Park, and et al. 2022. "Immunization with Virus-Like Particle Vaccine Protects Rabbits against Hepatitis E-3 Virus Infection" Viruses 14, no. 7: 1432. https://doi.org/10.3390/v14071432
APA StyleGo, H.-J., Park, B.-J., Ahn, H.-S., Han, S.-H., Kim, D.-H., Lyoo, E.-L., Kim, D.-Y., Kim, J.-H., Lee, J.-B., Park, S.-Y., Song, C.-S., Lee, S.-W., Choi, Y.-K., & Choi, I.-S. (2022). Immunization with Virus-Like Particle Vaccine Protects Rabbits against Hepatitis E-3 Virus Infection. Viruses, 14(7), 1432. https://doi.org/10.3390/v14071432