Measles Vaccines Designed for Enhanced CD8+ T Cell Activation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Cloning, Rescue and Propagation of Recombinant Measles Vaccine Viruses
- ggOVA MluI for (5′→3′) tttacgcgtgccaccatgggctccatcggcg
- ggOVA AscI rev (5′→3′) tttggcgcgcctattaaggggaaacacatctgcca
- mTRP-2 MluI for (5′→3′) tttacgcgtgccaccatgggccttgtggga
- mTRP-2 AscI rev (5′→3′) tttggcgcgcctaggcttcctccgtgt
- pN1 NUb fw (5′→3′) tttacgcgtgccaccatgcagatttttgtgaag
- pN1 NUb TRP-2 rev (5′→3′) ttttttgcgcgctcattagtcgacataggctgccaa
- pN1 NUb OVA rev (5′→3′) ttttttgcgcgctcattagtcgacataggctgccaa
2.3. Growth Curves
2.4. Western Blot
2.5. Synthetic Peptides
2.6. Enzyme-Linked Immunospot (ELISpot) Assay
2.7. Flow Cytometry
2.8. Data Analyses
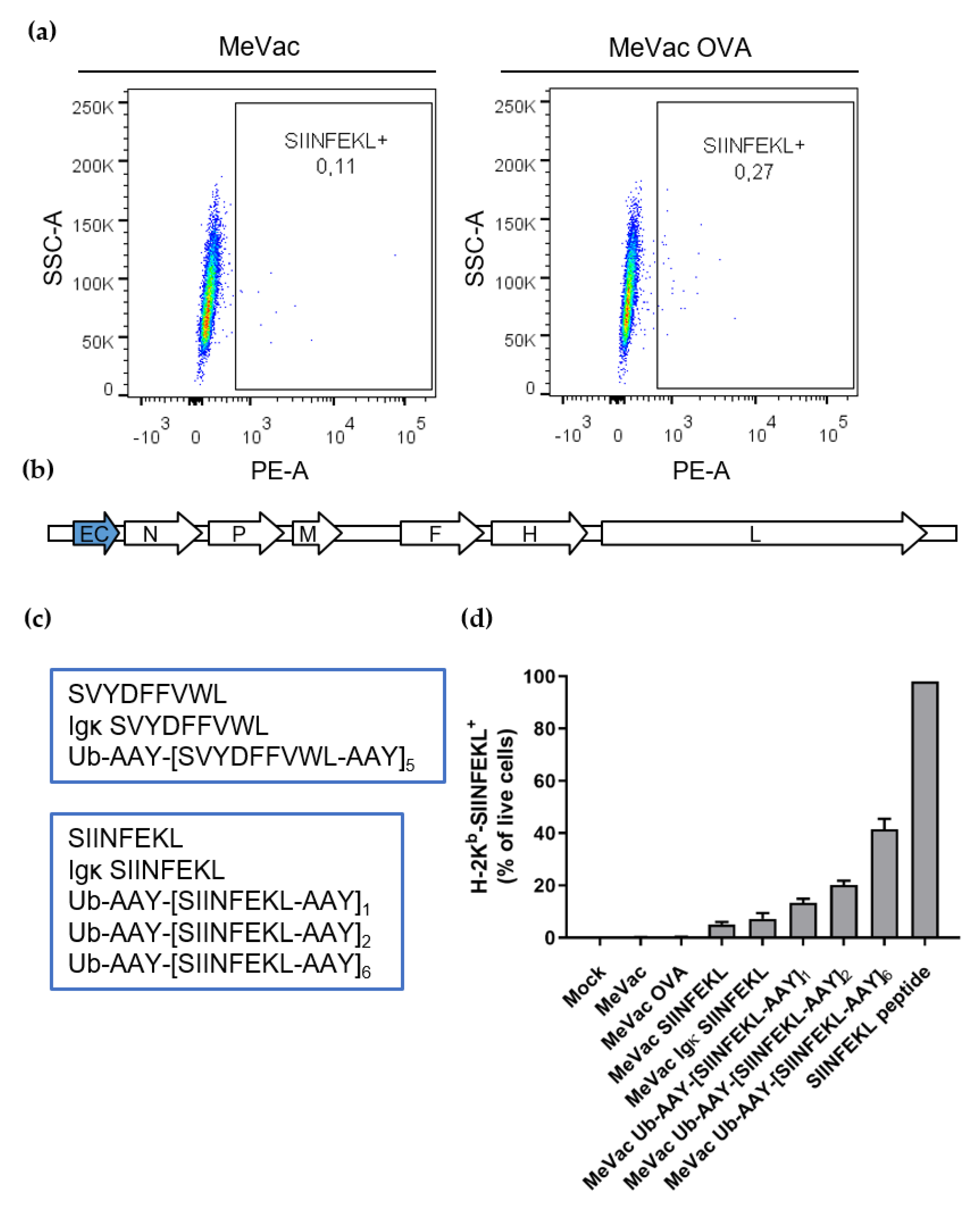
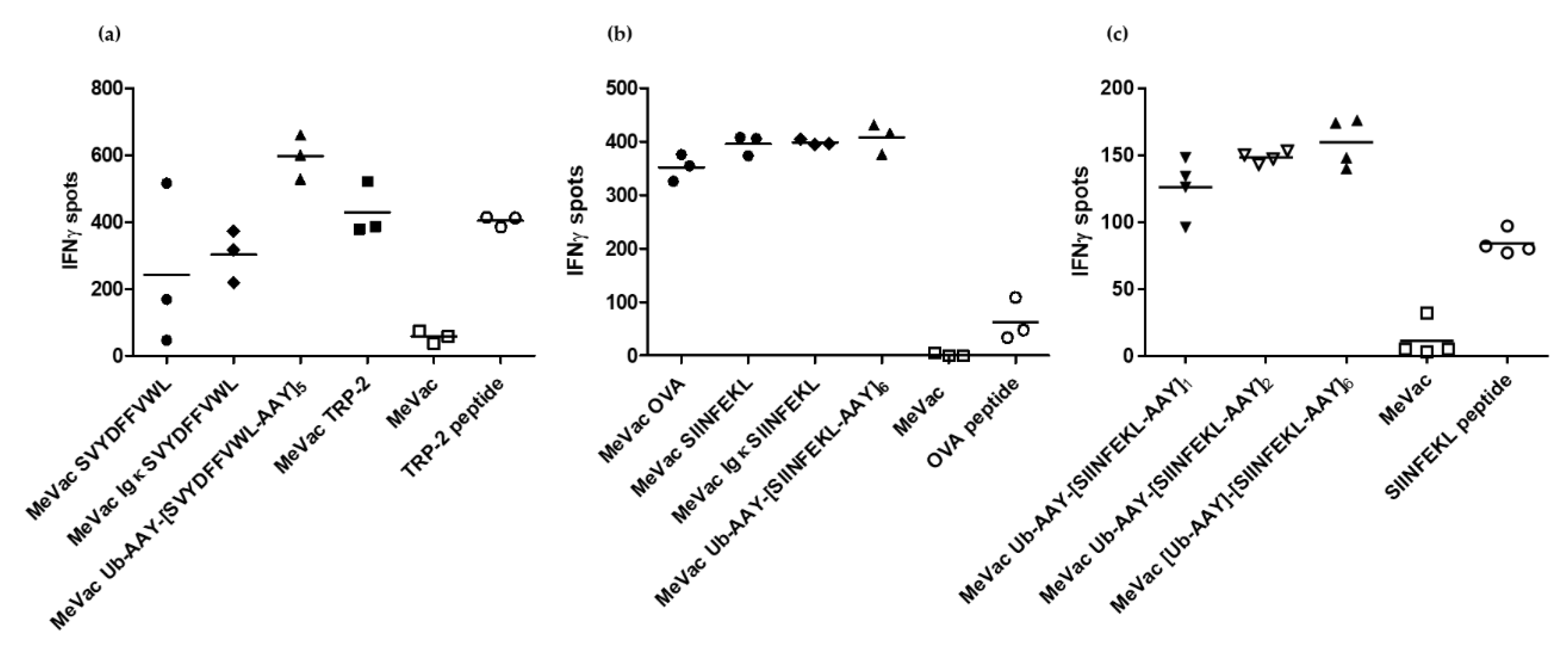
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Demicheli, V.; Rivetti, A.; Debalini, M.G.; Di Pietrantonj, C. Vaccines for measles, mumps and rubella in children. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef]
- Griffin, D.E. Measles Vaccine. Viral Immunol. 2018, 31, 86–95. [Google Scholar] [CrossRef]
- Muhlebach, M.D. Vaccine platform recombinant measles virus. Virus Genes 2017, 53, 733–740. [Google Scholar] [CrossRef]
- Hu, H.M.; Chen, H.W.; Hsiao, Y.J.; Wu, S.H.; Chung, H.H.; Hsieh, C.H.; Chong, P.; Leng, C.H.; Pan, C.H. The successful induction of T-cell and antibody responses by a recombinant measles virus-vectored tetravalent dengue vaccine provides partial protection against dengue-2 infection. Hum. Vaccines Immunother. 2016, 12, 1678–1689. [Google Scholar] [CrossRef]
- Lorin, C.; Mollet, L.; Delebecque, F.; Combredet, C.; Hurtrel, B.; Charneau, P.; Brahic, M.; Tangy, F. A single injection of recombinant measles virus vaccines expressing human immunodeficiency virus (HIV) type 1 clade B envelope glycoproteins induces neutralizing antibodies and cellular immune responses to HIV. J. Virol. 2004, 78, 146–157. [Google Scholar] [CrossRef]
- Singh, M.; Cattaneo, R.; Billeter, M.A. A recombinant measles virus expressing hepatitis B virus surface antigen induces humoral immune responses in genetically modified mice. J. Virol. 1999, 73, 4823–4828. [Google Scholar] [CrossRef]
- Liniger, M.; Zuniga, A.; Tamin, A.; Azzouz-Morin, T.N.; Knuchel, M.; Marty, R.R.; Wiegand, M.; Weibel, S.; Kelvin, D.; Rota, P.A.; et al. Induction of neutralising antibodies and cellular immune responses against SARS coronavirus by recombinant measles viruses. Vaccine 2008, 26, 2164–2174. [Google Scholar] [CrossRef] [PubMed]
- Malczyk, A.H.; Kupke, A.; Prufer, S.; Scheuplein, V.A.; Hutzler, S.; Kreuz, D.; Beissert, T.; Bauer, S.; Hubich-Rau, S.; Tondera, C.; et al. A Highly Immunogenic and Protective Middle East Respiratory Syndrome Coronavirus Vaccine Based on a Recombinant Measles Virus Vaccine Platform. J. Virol. 2015, 89, 11654–11667. [Google Scholar] [CrossRef]
- Mura, M.; Ruffie, C.; Combredet, C.; Aliprandini, E.; Formaglio, P.; Chitnis, C.E.; Amino, R.; Tangy, F. Recombinant measles vaccine expressing malaria antigens induces long-term memory and protection in mice. Npj Vaccines 2019, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, E.C.; Tschismarov, R.; Beubler, E.; Wiedermann, U.; Firbas, C.; Loebermann, M.; Pfeiffer, A.; Muellner, M.; Tauber, E.; Ramsauer, K. Immunogenicity, safety, and tolerability of the measles-vectored chikungunya virus vaccine MV-CHIK: A double-blind, randomised, placebo-controlled and active-controlled phase 2 trial. Lancet (Lond. Engl.) 2019, 392, 2718–2727. [Google Scholar] [CrossRef]
- Mateo, M.; Reynard, S.; Carnec, X.; Journeaux, A.; Baillet, N.; Schaeffer, J.; Picard, C.; Legras-Lachuer, C.; Allan, R.; Perthame, E.; et al. Vaccines inducing immunity to Lassa virus glycoprotein and nucleoprotein protect macaques after a single shot. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.J.; Barber, G.N. Oncolytic Viruses as Antigen-Agnostic Cancer Vaccines. Cancer Cell 2018, 33, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Pol, J.G.; Levesque, S.; Workenhe, S.T.; Gujar, S.; Le Boeuf, F.; Clements, D.R.; Fahrner, J.E.; Fend, L.; Bell, J.C.; Mossman, K.L.; et al. Trial Watch: Oncolytic viro-immunotherapy of hematologic and solid tumors. Oncoimmunology 2018, 7, e1503032. [Google Scholar] [CrossRef] [PubMed]
- Leber, M.F.; Hoyler, B.; Prien, S.; Neault, S.; Engeland, C.E.; Foerster, J.M.; Bossow, S.; Springfeld, C.; Von Kalle, C.; Jaeger, D.; et al. Sequencing of Serially Passaged Measles Virus Affirms its Genomic Stability and Reveals a Nonrandom Distribution of Consensus Mutations. J. Gen. Virol. 2020. [Google Scholar] [CrossRef]
- Robinson, S.; Galanis, E. Potential and clinical translation of oncolytic measles viruses. Expert Opin. Biol. Ther. 2017, 17, 353–363. [Google Scholar] [CrossRef]
- Grote, D.; Cattaneo, R.; Fielding, A.K. Neutrophils contribute to the measles virus-induced antitumor effect: Enhancement by granulocyte macrophage colony-stimulating factor expression. Cancer Res. 2003, 63, 6463–6468. [Google Scholar]
- Grossardt, C.; Engeland, C.E.; Bossow, S.; Halama, N.; Zaoui, K.; Leber, M.F.; Springfeld, C.; Jaeger, D.; von Kalle, C.; Ungerechts, G. Granulocyte-macrophage colony-stimulating factor-armed oncolytic measles virus is an effective therapeutic cancer vaccine. Hum. Gene Ther. 2013, 24, 644–654. [Google Scholar] [CrossRef]
- Iankov, I.D.; Allen, C.; Federspiel, M.J.; Myers, R.M.; Peng, K.W.; Ingle, J.N.; Russell, S.J.; Galanis, E. Expression of immunomodulatory neutrophil-activating protein of Helicobacter pylori enhances the antitumor activity of oncolytic measles virus. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 1139–1147. [Google Scholar] [CrossRef]
- Engeland, C.E.; Grossardt, C.; Veinalde, R.; Bossow, S.; Lutz, D.; Kaufmann, J.K.; Shevchenko, I.; Umansky, V.; Nettelbeck, D.M.; Weichert, W.; et al. CTLA-4 and PD-L1 checkpoint blockade enhances oncolytic measles virus therapy. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 1949–1959. [Google Scholar] [CrossRef]
- Veinalde, R.; Grossardt, C.; Hartmann, L.; Bourgeois-Daigneault, M.C.; Bell, J.C.; Jager, D.; von Kalle, C.; Ungerechts, G.; Engeland, C.E. Oncolytic measles virus encoding interleukin-12 mediates potent antitumor effects through T cell activation. Oncoimmunology 2017, 6, e1285992. [Google Scholar] [CrossRef]
- Speck, T.; Heidbuechel, J.P.W.; Veinalde, R.; Jaeger, D.; von Kalle, C.; Ball, C.R.; Ungerechts, G.; Engeland, C.E. Targeted BiTE Expression by an Oncolytic Vector Augments Therapeutic Efficacy Against Solid Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 2128–2137. [Google Scholar] [CrossRef] [PubMed]
- Backhaus, P.S.; Veinalde, R.; Hartmann, L.; Dunder, J.E.; Jeworowski, L.M.; Albert, J.; Hoyler, B.; Poth, T.; Jager, D.; Ungerechts, G.; et al. Immunological Effects and Viral Gene Expression Determine the Efficacy of Oncolytic Measles Vaccines Encoding IL-12 or IL-15 Agonists. Viruses 2019, 11, 914. [Google Scholar] [CrossRef] [PubMed]
- Dietz, L.; Engeland, C.E. Immunomodulation in Oncolytic Measles Virotherapy. Methods Mol. Biol. (Clifton N.J.) 2020, 2058, 111–126. [Google Scholar] [CrossRef]
- Kottke, T.; Errington, F.; Pulido, J.; Galivo, F.; Thompson, J.; Wongthida, P.; Diaz, R.M.; Chong, H.; Ilett, E.; Chester, J.; et al. Broad antigenic coverage induced by vaccination with virus-based cDNA libraries cures established tumors. Nat. Med. 2011, 17, 854–859. [Google Scholar] [CrossRef]
- Pulido, J.; Kottke, T.; Thompson, J.; Galivo, F.; Wongthida, P.; Diaz, R.M.; Rommelfanger, D.; Ilett, E.; Pease, L.; Pandha, H.; et al. Using virally expressed melanoma cDNA libraries to identify tumor-associated antigens that cure melanoma. Nat. Biotechnol. 2012, 30, 337–343. [Google Scholar] [CrossRef]
- Atherton, M.J.; Stephenson, K.B.; Tzelepis, F.; Bakhshinyan, D.; Nikota, J.K.; Son, H.H.; Jirovec, A.; Lefebvre, C.; Dvorkin-Gheva, A.; Ashkar, A.A.; et al. Transforming the prostatic tumor microenvironment with oncolytic virotherapy. Oncoimmunology 2018, 7, e1445459. [Google Scholar] [CrossRef]
- McGray, A.J.R.; Huang, R.Y.; Battaglia, S.; Eppolito, C.; Miliotto, A.; Stephenson, K.B.; Lugade, A.A.; Webster, G.; Lichty, B.D.; Seshadri, M.; et al. Oncolytic Maraba virus armed with tumor antigen boosts vaccine priming and reveals diverse therapeutic response patterns when combined with checkpoint blockade in ovarian cancer. J. Immunother. Cancer 2019, 7, 189. [Google Scholar] [CrossRef]
- Pol, J.G.; Acuna, S.A.; Yadollahi, B.; Tang, N.; Stephenson, K.B.; Atherton, M.J.; Hanwell, D.; El-Warrak, A.; Goldstein, A.; Moloo, B.; et al. Preclinical evaluation of a MAGE-A3 vaccination utilizing the oncolytic Maraba virus currently in first-in-human trials. Oncoimmunology 2019, 8, e1512329. [Google Scholar] [CrossRef]
- Pol, J.G.; Zhang, L.; Bridle, B.W.; Stephenson, K.B.; Resseguier, J.; Hanson, S.; Chen, L.; Kazdhan, N.; Bramson, J.L.; Stojdl, D.F.; et al. Maraba virus as a potent oncolytic vaccine vector. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 420–429. [Google Scholar] [CrossRef]
- Atherton, M.J.; Stephenson, K.B.; Pol, J.; Wang, F.; Lefebvre, C.; Stojdl, D.F.; Nikota, J.K.; Dvorkin-Gheva, A.; Nguyen, A.; Chen, L.; et al. Customized Viral Immunotherapy for HPV-Associated Cancer. Cancer Immunol. Res. 2017, 5, 847–859. [Google Scholar] [CrossRef]
- Hutzler, S.; Erbar, S.; Jabulowsky, R.A.; Hanauer, J.R.H.; Schnotz, J.H.; Beissert, T.; Bodmer, B.S.; Eberle, R.; Boller, K.; Klamp, T.; et al. Antigen-specific oncolytic MV-based tumor vaccines through presentation of selected tumor-associated antigens on infected cells or virus-like particles. Sci. Rep. 2017, 7, 16892. [Google Scholar] [CrossRef]
- Lei, J.; Osen, W.; Gardyan, A.; Hotz-Wagenblatt, A.; Wei, G.; Gissmann, L.; Eichmuller, S.; Lochelt, M. Replication-Competent Foamy Virus Vaccine Vectors as Novel Epitope Scaffolds for Immunotherapy. PLoS ONE 2015, 10, e0138458. [Google Scholar] [CrossRef]
- Falk, K.; Rotzschke, O.; Stevanovic, S.; Jung, G.; Rammensee, H.G. Allele-specific motifs revealed by sequencing of self-peptides eluted from MHC molecules. Nature 1991, 351, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Bloom, M.B.; Perry-Lalley, D.; Robbins, P.F.; Li, Y.; el-Gamil, M.; Rosenberg, S.A.; Yang, J.C. Identification of tyrosinase-related protein 2 as a tumor rejection antigen for the B16 melanoma. J. Exp. Med. 1997, 185, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Samorski, R.; Gissmann, L.; Osen, W. Codon optimized expression of HPV 16 E6 renders target cells susceptible to E6-specific CTL recognition. Immunol. Lett. 2006, 107, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Radecke, F.; Spielhofer, P.; Schneider, H.; Kaelin, K.; Huber, M.; Dotsch, C.; Christiansen, G.; Billeter, M.A. Rescue of measles viruses from cloned DNA. EMBO J. 1995, 14, 5773–5784. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Staeheli, P.; Schneider, U. RNA polymerase II-controlled expression of antigenomic RNA enhances the rescue efficacies of two different members of the Mononegavirales independently of the site of viral genome replication. J. Virol. 2006, 80, 5708–5715. [Google Scholar] [CrossRef]
- Calain, P.; Roux, L. The rule of six, a basic feature for efficient replication of Sendai virus defective interfering RNA. J. Virol. 1993, 67, 4822–4830. [Google Scholar] [CrossRef]
- Heidbuechel, J.P.W.; Engeland, C.E. Paramyxoviruses for Tumor-targeted Immunomodulation: Design and Evaluation Ex Vivo. J. Vis. Exp. Jove 2019. [Google Scholar] [CrossRef]
- Merrifield, R.B. Solid phase peptide synthesis. I. The synthesis of a tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- Carpino, L.A.; Han, G. 9-Fluorenylmethoxycarbonyl amino-protecting group. J. Org. Chem. 1972, 37, 3404–3409. [Google Scholar] [CrossRef]
- Grigorov, B.; Rabilloud, J.; Lawrence, P.; Gerlier, D. Rapid titration of measles and other viruses: Optimization with determination of replication cycle length. PLoS ONE 2011, 6, e24135. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, R.; Rebmann, G.; Baczko, K.; ter Meulen, V.; Billeter, M.A. Altered ratios of measles virus transcripts in diseased human brains. Virology 1987, 160, 523–526. [Google Scholar] [CrossRef]
- Wu, Y.; Kipps, T.J. Deoxyribonucleic acid vaccines encoding antigens with rapid proteasome-dependent degradation are highly efficient inducers of cytolytic T lymphocytes. J. Immunol. (Baltimore, Md.: 1950) 1997, 159, 6037–6043. [Google Scholar]
- Rodriguez, F.; Zhang, J.; Whitton, J.L. DNA immunization: Ubiquitination of a viral protein enhances cytotoxic T-lymphocyte induction and antiviral protection but abrogates antibody induction. J. Virol. 1997, 71, 8497–8503. [Google Scholar] [CrossRef]
- Bergmann, C.C.; Yao, Q.; Ho, C.K.; Buckwold, S.L. Flanking residues alter antigenicity and immunogenicity of multi-unit CTL epitopes. J. Immunol. (Baltimore, Md.: 1950) 1996, 157, 3242–3249. [Google Scholar]
- Nussbaum, A.K.; Dick, T.P.; Keilholz, W.; Schirle, M.; Stevanovic, S.; Dietz, K.; Heinemeyer, W.; Groll, M.; Wolf, D.H.; Huber, R.; et al. Cleavage motifs of the yeast 20S proteasome beta subunits deduced from digests of enolase 1. Proc. Natl. Acad. Sci. USA 1998, 95, 12504–12509. [Google Scholar] [CrossRef]
- Holzhutter, H.G.; Frommel, C.; Kloetzel, P.M. A theoretical approach towards the identification of cleavage-determining amino acid motifs of the 20 S proteasome. J. Mol. Biol. 1999, 286, 1251–1265. [Google Scholar] [CrossRef]
- Velders, M.P.; Weijzen, S.; Eiben, G.L.; Elmishad, A.G.; Kloetzel, P.M.; Higgins, T.; Ciccarelli, R.B.; Evans, M.; Man, S.; Smith, L.; et al. Defined flanking spacers and enhanced proteolysis is essential for eradication of established tumors by an epitope string DNA vaccine. J. Immunol. (Baltimore, Md.: 1950) 2001, 166, 5366–5373. [Google Scholar] [CrossRef]
- Koup, R.A.; Douek, D.C. Vaccine design for CD8 T lymphocyte responses. Cold Spring Harb. Perspect. Med. 2011, 1, a007252. [Google Scholar] [CrossRef]
- Reguzova, A.; Antonets, D.; Karpenko, L.; Ilyichev, A.; Maksyutov, R.; Bazhan, S. Design and evaluation of optimized artificial HIV-1 poly-T cell-epitope immunogens. PLoS ONE 2015, 10, e0116412. [Google Scholar] [CrossRef] [PubMed]
- Rennick, L.J.; de Vries, R.D.; Carsillo, T.J.; Lemon, K.; van Amerongen, G.; Ludlow, M.; Nguyen, D.T.; Yuksel, S.; Verburgh, R.J.; Haddock, P.; et al. Live-attenuated measles virus vaccine targets dendritic cells and macrophages in muscle of nonhuman primates. J. Virol. 2015, 89, 2192–2200. [Google Scholar] [CrossRef] [PubMed]
- Guillerme, J.B.; Boisgerault, N.; Roulois, D.; Menager, J.; Combredet, C.; Tangy, F.; Fonteneau, J.F.; Gregoire, M. Measles virus vaccine-infected tumor cells induce tumor antigen cross-presentation by human plasmacytoid dendritic cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Gauvrit, A.; Brandler, S.; Sapede-Peroz, C.; Boisgerault, N.; Tangy, F.; Gregoire, M. Measles virus induces oncolysis of mesothelioma cells and allows dendritic cells to cross-prime tumor-specific CD8 response. Cancer Res. 2008, 68, 4882–4892. [Google Scholar] [CrossRef]
- Bolton, D.L.; Santra, S.; Swett-Tapia, C.; Custers, J.; Song, K.; Balachandran, H.; Mach, L.; Naim, H.; Kozlowski, P.A.; Lifton, M.; et al. Priming T-cell responses with recombinant measles vaccine vector in a heterologous prime-boost setting in non-human primates. Vaccine 2012, 30, 5991–5998. [Google Scholar] [CrossRef]
- Borst, J.; Ahrends, T.; Babala, N.; Melief, C.J.M.; Kastenmuller, W. CD4(+) T cell help in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2018, 18, 635–647. [Google Scholar] [CrossRef]
- Billeter, M.A.; Naim, H.Y.; Udem, S.A. Reverse genetics of measles virus and resulting multivalent recombinant vaccines: Applications of recombinant measles viruses. Curr. Top. Microbiol. Immunol. 2009, 329, 129–162. [Google Scholar]
- Yewdell, J.W. Confronting complexity: Real-world immunodominance in antiviral CD8+ T cell responses. Immunity 2006, 25, 533–543. [Google Scholar] [CrossRef]
- Kotturi, M.F.; Scott, I.; Wolfe, T.; Peters, B.; Sidney, J.; Cheroutre, H.; von Herrath, M.G.; Buchmeier, M.J.; Grey, H.; Sette, A. Naive precursor frequencies and MHC binding rather than the degree of epitope diversity shape CD8+ T cell immunodominance. J. Immunol. (Baltimore, Md.: 1950) 2008, 181, 2124–2133. [Google Scholar] [CrossRef]
- La Gruta, N.L.; Kedzierska, K.; Pang, K.; Webby, R.; Davenport, M.; Chen, W.; Turner, S.J.; Doherty, P.C. A virus-specific CD8+ T cell immunodominance hierarchy determined by antigen dose and precursor frequencies. Proc. Natl. Acad. Sci. USA 2006, 103, 994–999. [Google Scholar] [CrossRef]
- Ovsyannikova, I.G.; Reid, K.C.; Jacobson, R.M.; Oberg, A.L.; Klee, G.G.; Poland, G.A. Cytokine production patterns and antibody response to measles vaccine. Vaccine 2003, 21, 3946–3953. [Google Scholar] [CrossRef]
- Butler, N.S.; Nolz, J.C.; Harty, J.T. Immunologic considerations for generating memory CD8 T cells through vaccination. Cell. Microbiol. 2011, 13, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Peng, K.W.; Dingli, D.; Kratzke, R.A.; Russell, S.J. Oncolytic measles viruses encoding interferon beta and the thyroidal sodium iodide symporter gene for mesothelioma virotherapy. Cancer Gene Ther. 2010, 17, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Pol, J.G.; Atherton, M.J.; Bridle, B.W.; Stephenson, K.B.; Le Boeuf, F.; Hummel, J.L.; Martin, C.G.; Pomoransky, J.; Breitbach, C.J.; Diallo, J.S.; et al. Development and applications of oncolytic Maraba virus vaccines. Oncolytic Virotherapy 2018, 7, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Bridle, B.W.; Nguyen, A.; Salem, O.; Zhang, L.; Koshy, S.; Clouthier, D.; Chen, L.; Pol, J.; Swift, S.L.; Bowdish, D.M.; et al. Privileged Antigen Presentation in Splenic B Cell Follicles Maximizes T Cell Responses in Prime-Boost Vaccination. J. Immunol. (Baltimore, Md.: 1950) 2016, 196, 4587–4595. [Google Scholar] [CrossRef]
- Zitvogel, L.; Galluzzi, L.; Kepp, O.; Smyth, M.J.; Kroemer, G. Type I interferons in anticancer immunity. Nat. Rev. Immunol. 2015, 15, 405–414. [Google Scholar] [CrossRef]
- Cai, L.; Michelakos, T.; Yamada, T.; Fan, S.; Wang, X.; Schwab, J.H.; Ferrone, C.R.; Ferrone, S. Defective HLA class I antigen processing machinery in cancer. Cancer Immunol. Immunother. Cii 2018, 67, 999–1009. [Google Scholar] [CrossRef]
- Miest, T.S.; Yaiw, K.C.; Frenzke, M.; Lampe, J.; Hudacek, A.W.; Springfeld, C.; von Messling, V.; Ungerechts, G.; Cattaneo, R. Envelope-chimeric entry-targeted measles virus escapes neutralization and achieves oncolysis. Mol. Ther. J. Am. Soc. Gene Ther. 2011, 19, 1813–1820. [Google Scholar] [CrossRef]
- Munoz-Alia, M.A.; Russell, S.J. Probing Morbillivirus Antisera Neutralization Using Functional Chimerism between Measles Virus and Canine Distemper Virus Envelope Glycoproteins. Viruses 2019, 11, 688. [Google Scholar] [CrossRef]
- Engeland, C.E.; Bossow, S.; Hudacek, A.W.; Hoyler, B.; Forster, J.; Veinalde, R.; Jager, D.; Cattaneo, R.; Ungerechts, G.; Springfeld, C. A Tupaia paramyxovirus vector system for targeting and transgene expression. J. Gen. Virol. 2017, 98, 2248–2257. [Google Scholar] [CrossRef]
- Ricca, J.M.; Oseledchyk, A.; Walther, T.; Liu, C.; Mangarin, L.; Merghoub, T.; Wolchok, J.D.; Zamarin, D. Pre-existing Immunity to Oncolytic Virus Potentiates Its Immunotherapeutic Efficacy. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Berkeley, R.A.; Steele, L.P.; Mulder, A.A.; van den Wollenberg, D.J.M.; Kottke, T.J.; Thompson, J.; Coffey, M.; Hoeben, R.C.; Vile, R.G.; Melcher, A.; et al. Antibody-Neutralized Reovirus Is Effective in Oncolytic Virotherapy. Cancer Immunol. Res. 2018, 6, 1161–1173. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busch, E.; Kubon, K.D.; Mayer, J.K.M.; Pidelaserra-Martí, G.; Albert, J.; Hoyler, B.; Heidbuechel, J.P.W.; Stephenson, K.B.; Lichty, B.D.; Osen, W.; et al. Measles Vaccines Designed for Enhanced CD8+ T Cell Activation. Viruses 2020, 12, 242. https://doi.org/10.3390/v12020242
Busch E, Kubon KD, Mayer JKM, Pidelaserra-Martí G, Albert J, Hoyler B, Heidbuechel JPW, Stephenson KB, Lichty BD, Osen W, et al. Measles Vaccines Designed for Enhanced CD8+ T Cell Activation. Viruses. 2020; 12(2):242. https://doi.org/10.3390/v12020242
Chicago/Turabian StyleBusch, Elena, Kristina D. Kubon, Johanna K. M. Mayer, Gemma Pidelaserra-Martí, Jessica Albert, Birgit Hoyler, Johannes P. W. Heidbuechel, Kyle B. Stephenson, Brian D. Lichty, Wolfram Osen, and et al. 2020. "Measles Vaccines Designed for Enhanced CD8+ T Cell Activation" Viruses 12, no. 2: 242. https://doi.org/10.3390/v12020242
APA StyleBusch, E., Kubon, K. D., Mayer, J. K. M., Pidelaserra-Martí, G., Albert, J., Hoyler, B., Heidbuechel, J. P. W., Stephenson, K. B., Lichty, B. D., Osen, W., Eichmüller, S. B., Jäger, D., Ungerechts, G., & Engeland, C. E. (2020). Measles Vaccines Designed for Enhanced CD8+ T Cell Activation. Viruses, 12(2), 242. https://doi.org/10.3390/v12020242