Morphological Characteristics and Allometric Relationships of Shoot in Two Undergrowth Plants: Polygonatum odoratum and Polygonatum multiflorum

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Morpho-Anatomical Analysis of Polygonatum Shoots
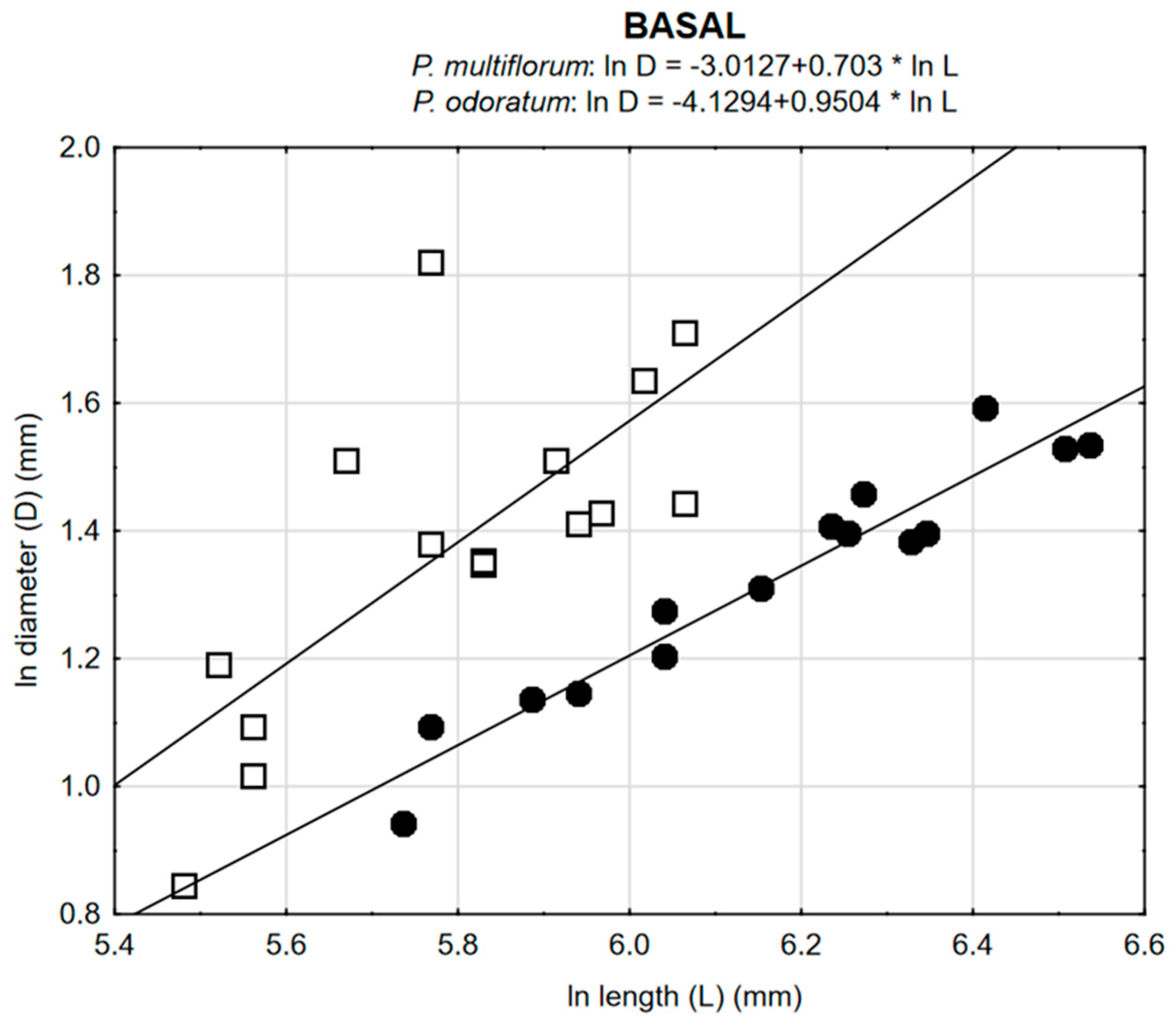
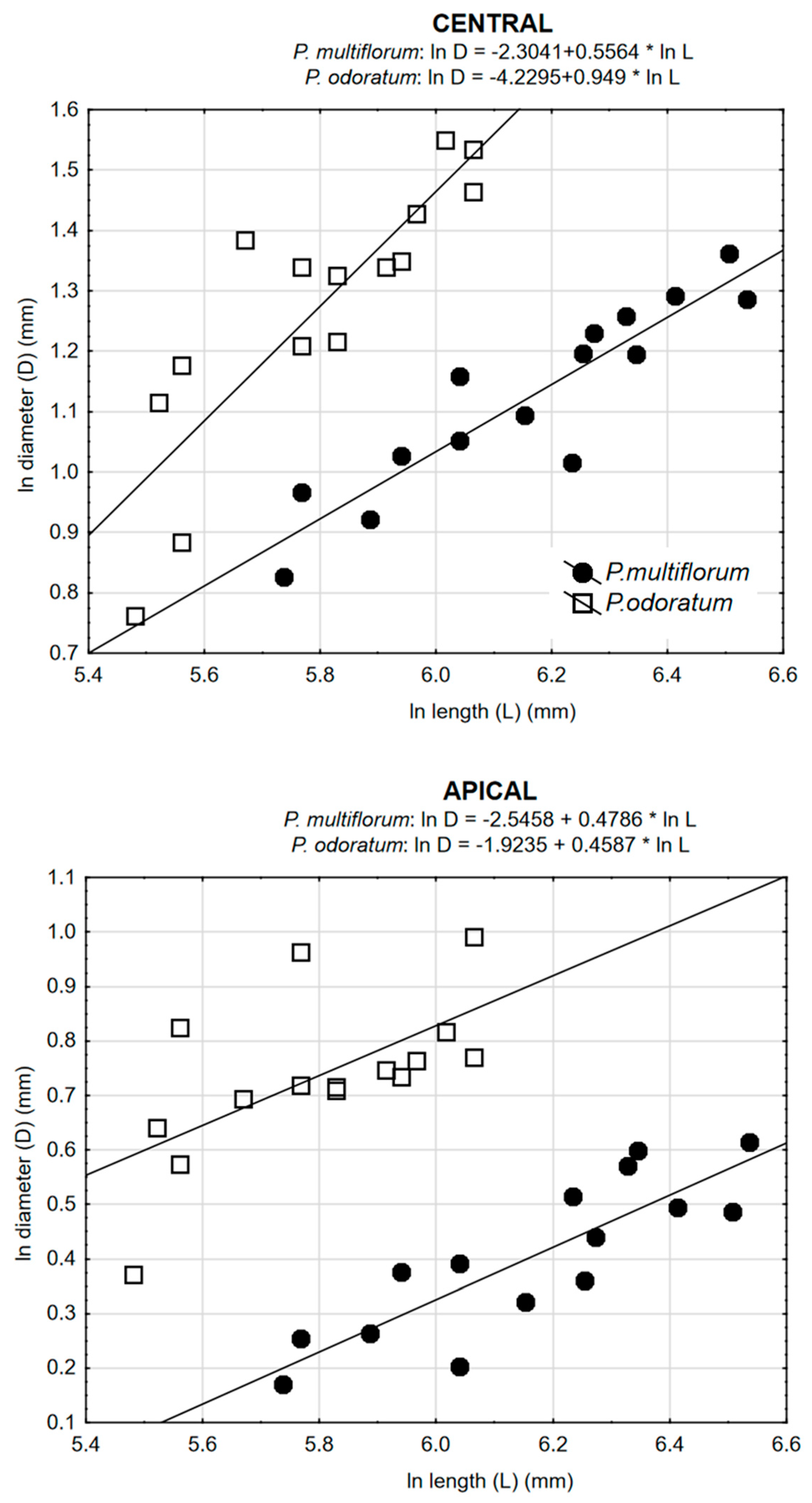
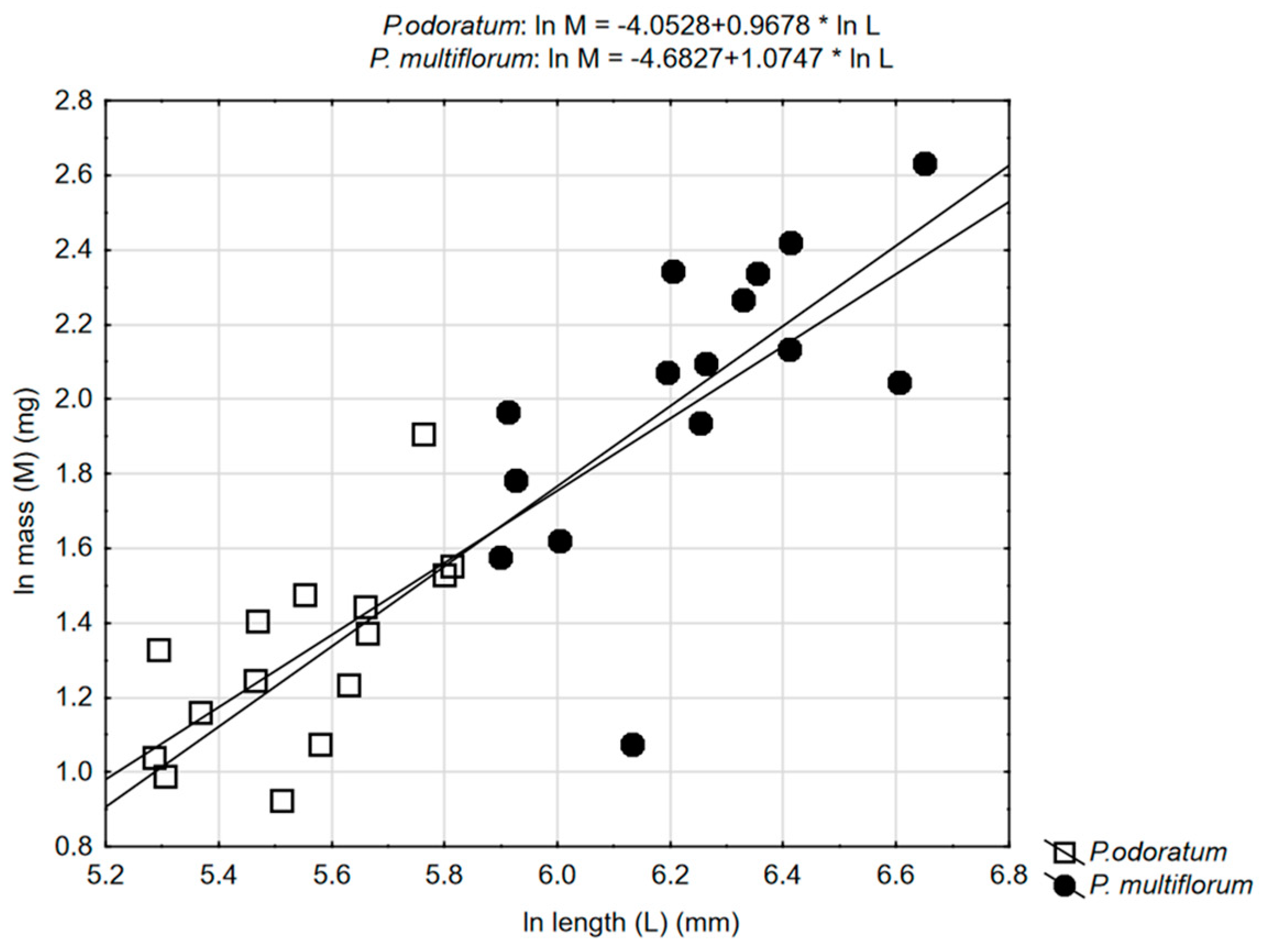
3.2. Allometric Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reinhard, D.; Kuhlemeier, C. Plant architecture. EMBO Rep. 2002, 3, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Crook, M.J.; Ennos, A.R. Mechanical differences between free-standing and supported wheat plants, Triticum aestivum L. Ann. Bot. 1996, 77, 197–202. [Google Scholar] [CrossRef]
- Gartner, B.L. Root biomechanics and whole plant allocation patterns: Responses of tomato to stem flexure. J. Exp. Bot. 1994, 45, 1647–1654. [Google Scholar] [CrossRef]
- Crook, M.J.; Ennos, A.R.; Sellers, E.K. Structural development of the shoot and root systems of two winter wheat cultivars. J. Exp. Bot. 1994, 45, 857–863. [Google Scholar] [CrossRef]
- Niklas, K.J. Plant allometry: The Scaling of Form and Process; University of Chicago Press: Chicago, IL, USA, 1994; ISBN 9780226580814. [Google Scholar]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Amer. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Schmidt, W.; Weitemeier, M.; Holzapfel, C. Vegetation dynamics in canopy gaps of beech forest on limestone—The influence of the light gradient on species richness. Verh. Ges. Ökol. 1996, 25, 253–260. [Google Scholar]
- Elemans, M. Light, nutirents and the growth of herbaceous forest species. Acta Oecol. 2004, 26, 197–202. [Google Scholar] [CrossRef]
- Tinya, F.; Márialigeti, S.; Király, B.; Németh, B.; Ódor, P. The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant Ecol. 2009, 204, 69–81. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Pearcy, R.W. The importance of sunflecks for forest understory plants. BioScience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Kitajima, K.; Mulkey, S.S.; Wright, J. Variation in crown light utilization characteristics among tropical canopy trees. Ann. Bot. 2005, 95, 535–547. [Google Scholar] [CrossRef]
- Nűnements, Ű. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Obmiński, Z. Ekologia Lasu; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1977; pp. 260–268. [Google Scholar]
- Gärtner, H.; Schweingruber, F.H. Microscopic Preparation Techniques for Plant Stem Analysis; Kessel Publishing House: Remagen, Germany, 2013; ISBN 978-3-941300-76-7. [Google Scholar]
- Broda, B. Metody Histochemii Roślinnej; Państwowy Zakład Wydawnictw Lekarskich: Warszawa, Poland, 1971. [Google Scholar]
- Weiner, J.; Thomas, S.C. Competition and allometry in three species of annual plants. Ecology 1992, 73, 648–656. [Google Scholar] [CrossRef]
- Poorter, H.; Jagodziński, A.M.; Ruiz-Peinado, R.P.; Kuyah, S.; Luo, Y.; Oleksyn, J.; Usoltsev, V.A.; Buckley, T.N.; Reich, T.N.; Reich, P.B.; et al. Howe does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five country. New Phytol. 2015, 208, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.F.V. The mechanical design of grass. J. Mater. Sci. 1982, 17, 856–860. [Google Scholar] [CrossRef]
- Goodman, A.M. Mechanical adaptations of Cleavers (Gallium aparine). Ann. Bot. 2005, 95, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Koehl, M.A.R.; Wainwright, S.A. Mechanical adaptation of a giant kelp. Limnol. Oceanogr. 1997, 22, 1067–1071. [Google Scholar] [CrossRef]
- Rüggeberg, M.; Bugert, I.; Speck, T. Structural and mechanical design of tissue interfaces in the giant reed Arundo donax. J. R. Soc. Interface 2010, 7, 499–506. [Google Scholar] [CrossRef]
- Rich, P.M. Mechanical structure of the stem of arborescent palms. Bot. Gaz. 1987, 148, 42–50. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The Structural Biology of Palms; Clarendon Press: Oxford, UK, 1990; ISBN 10: 019854572X. [Google Scholar]
- Spatz, H.C.; Beismann, H.; Brüchert, F.; Emanns, A.; Speck, T. Biomechanics of the giant reed Arundo donax. Philos. Trans. R. Soc. Lond. 1997, 352, 1–10. [Google Scholar] [CrossRef]
- Frey, L. Trawy niezwyciężone. Łąkarstwo W Pol. 2000, 3, 9–20. [Google Scholar]
- Winer, J.; Bernston, G.M.; Thomas, S.C. Competition and growth form in a woodland annual. J. Ecol. 1990, 78, 459–469. [Google Scholar] [CrossRef]
- Watari, R.; Nagashima, H.; Hirose, T. Stem extension and mechanical stability of Xantium canadense grown in an open or in dense stand. Ann. Bot. 2014, 114, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Zobel, B.J.; Sprague, J.R. Juvenille Wood in Forest Trees; Springer-Verlag: Berlin, Germany, 1998; ISBN 978-3-642-72126-7. [Google Scholar]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1517. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. Plant height and the properties of some herbaceous stems. Ann. Bot. 1995, 75, 133–142. [Google Scholar] [CrossRef]
- Castelan-Estrada, M.; Vivin, P.; Gaudillére, J.P. Allometric relationships estimate seasonal above-ground vegetative and reproductive biomass of Vitis vinifera L. Ann. Bot. 2002, 89, 401–408. [Google Scholar] [CrossRef]
- Enquist, B.J. Universal scaling in tree and vascular plant allometry toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiol. 2002, 22, 1045–1064. [Google Scholar] [CrossRef]
- Prior, S.A.; Rogers, H.H. Soybean growth response to water supply. J. Plant Nutr. 1995, 18, 617–636. [Google Scholar] [CrossRef]
- Wang, P.; Weiner, J.; Cahill, J.F.; Zhou, D.W.; Bian, H.F.; Song, Y.T.; Sheng, L.X. Shoot competition, root competition and reproductive allocation in Chenopodium accuminatum. J. Ecol. 2014, 102, 1688–1696. [Google Scholar] [CrossRef]
- Jarzyna, I. Plant morphology and allometric relationships in competing and non-competing plants of Tagetes patula L. Acta Soc. Bot. Pol. 2002, 71, 149–154. [Google Scholar] [CrossRef]








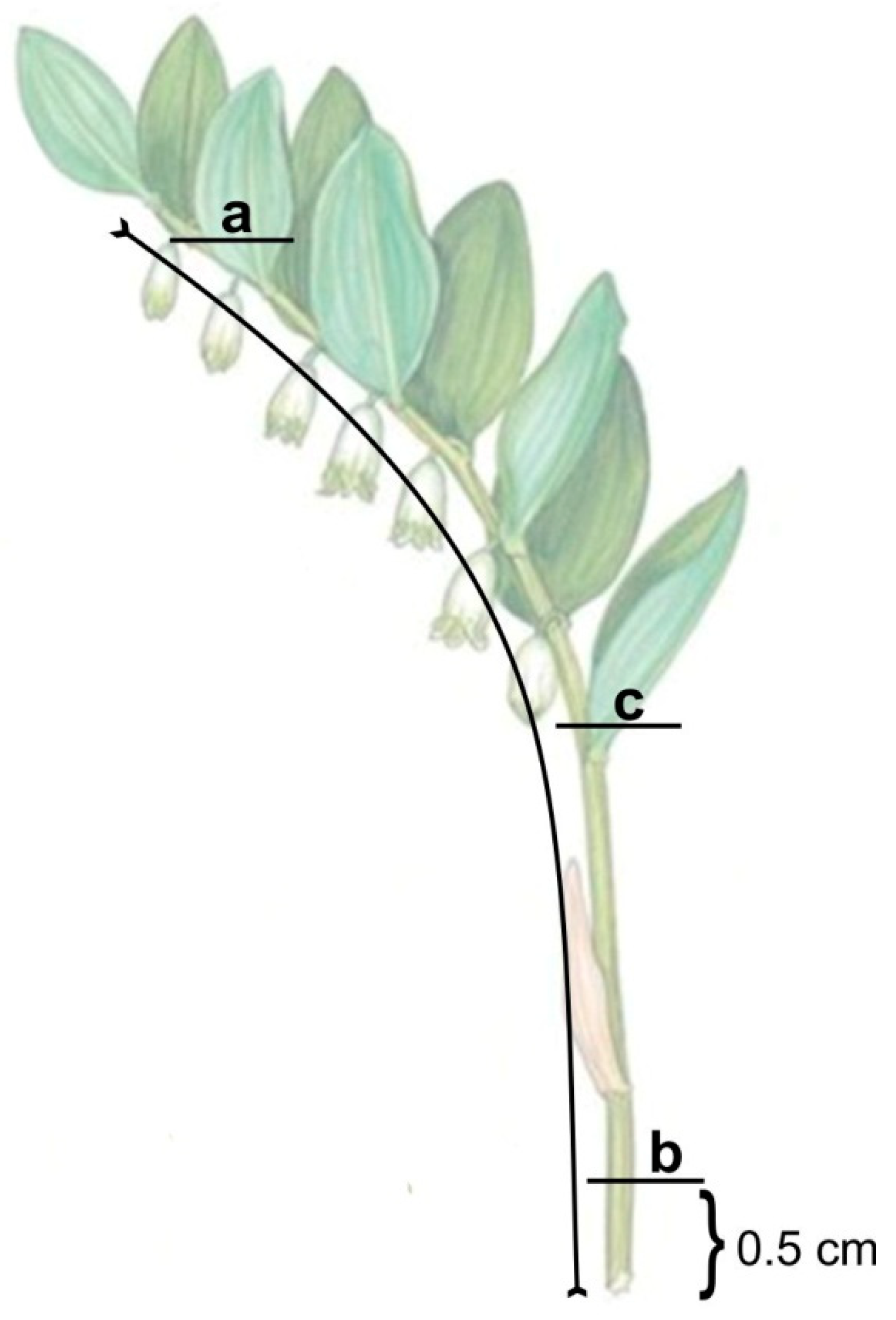{kind=link}
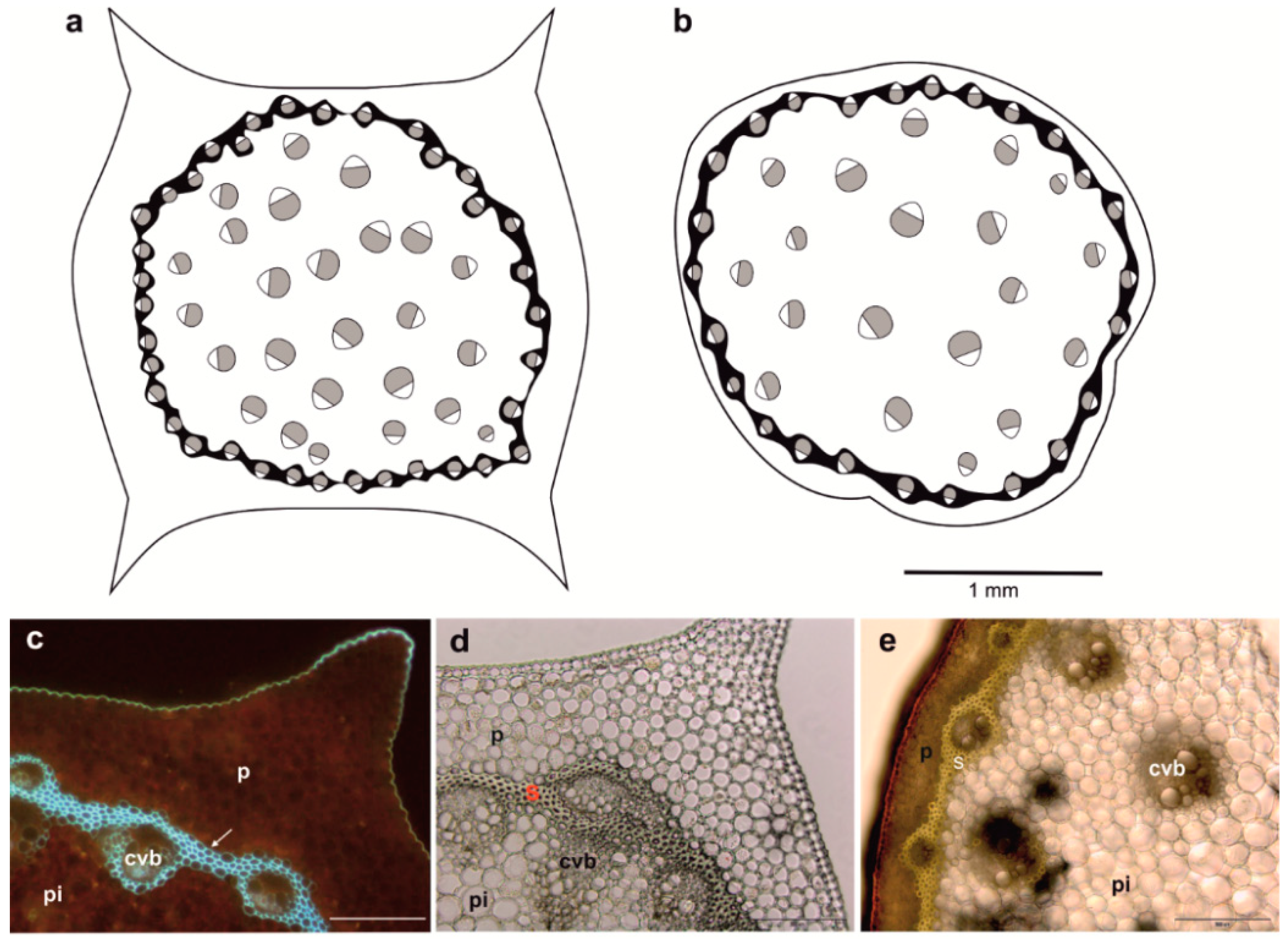{kind=link}
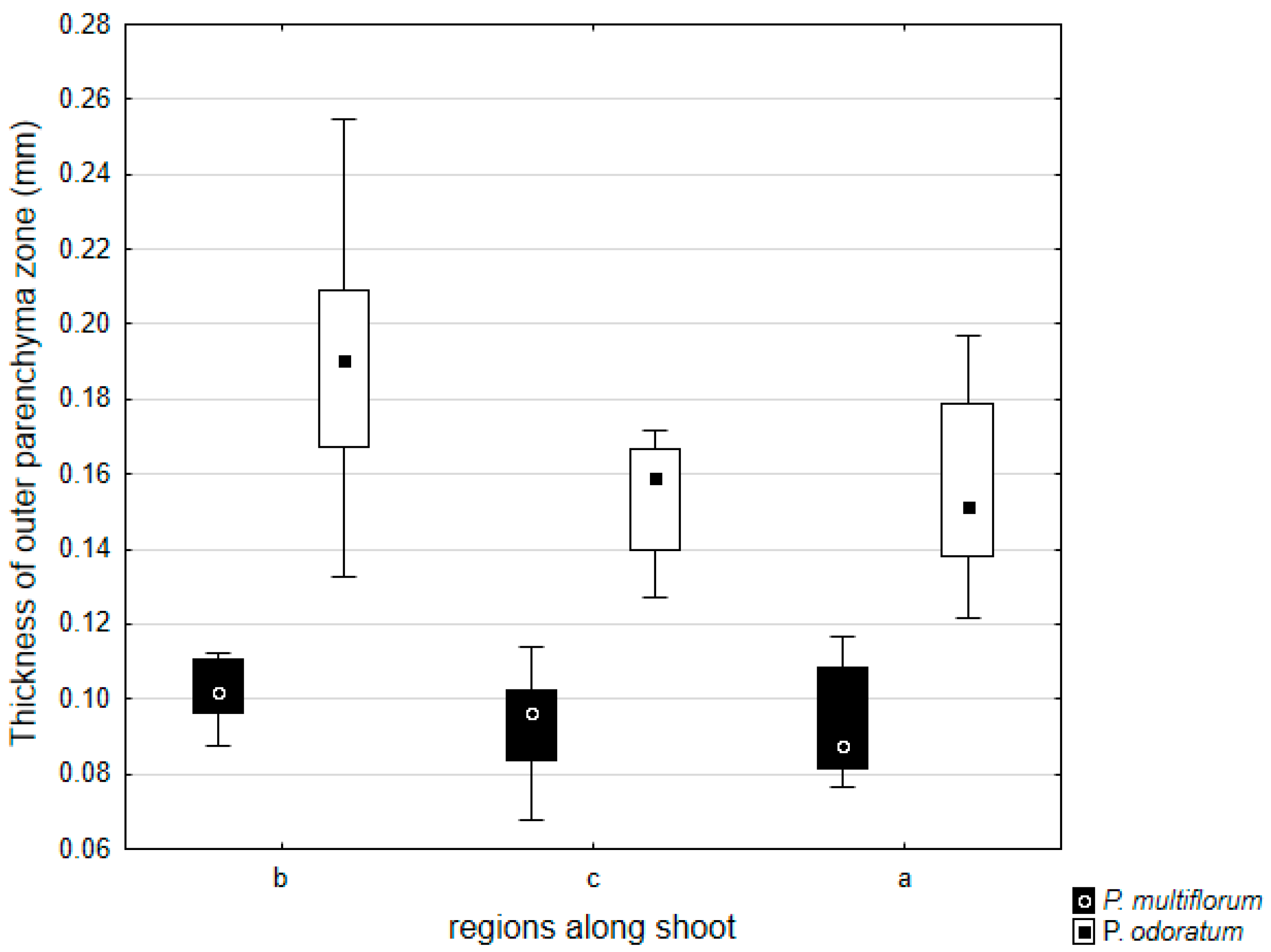{kind=link}
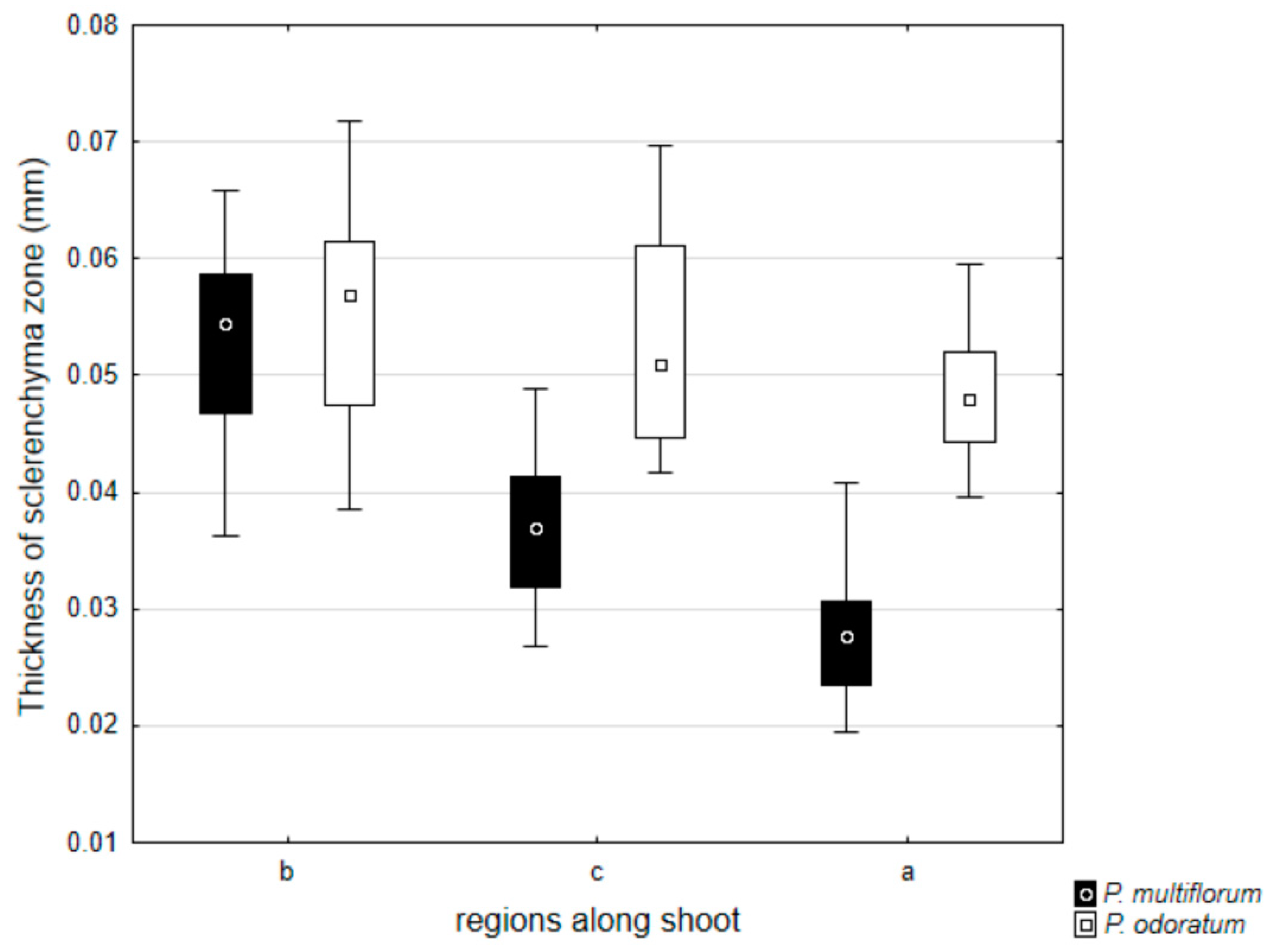{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable at Categorized Region | Mean | t-Value | p-Value | |
|---|---|---|---|---|
| P. o | P. m | |||
| Basal | 0.1876 | 0.1047 | 8.6023 | 0.0000 |
| Central | 0.1589 | 0.0930 | 7.3910 | 0.0000 |
| Apical | 0.1555 | 0.0931 | 8.8375 | 0.0000 |
| Variable | Mean | t-Value | p-Value | |
|---|---|---|---|---|
| P. o | P. m | |||
| Radius | 1.6416 | 1.4050 | 2.0269 | 0.0456 |
| Thickness of outer parenchyma zone | 0.1674 | 0.0969 | 13.0812 | 0.0000 |
| Thickness of sclerenchyma zone | 0.05217 | 0.0395 | 5.28161 | 0.0000 |
| r D–L | |||
|---|---|---|---|
| Basal | Central | Apical | |
| P. odoratum | 0.7305 ** | 0.8555 *** | 0.6224 * |
| P. multiflorum | 0.9626 *** | 0.9152 *** | 0.8511 *** |
| P. odoratum | ||||
| Ws-D | Ws-L | Wp-D | Wp-L | |
| basal | 0.529657 * | 0.664209 ** | NS | 0.599483 * |
| central | NS | |||
| apical | NS | NS | NS | |
| P. multiflorum | ||||
| Ws-D | Ws-L | Wp-D | Wp-L | |
| basal | 0.816187 *** | 0.799026 *** | NS | |
| central | NS | NS | NS | NS |
| apical | 0.522822 * | 0.741862 ** | NS | NS |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tulik, M.; Karczewski, J.; Szeliga, N.; Jura-Morawiec, J.; Jarzyna, I. Morphological Characteristics and Allometric Relationships of Shoot in Two Undergrowth Plants: Polygonatum odoratum and Polygonatum multiflorum. Forests 2018, 9, 783. https://doi.org/10.3390/f9120783
Tulik M, Karczewski J, Szeliga N, Jura-Morawiec J, Jarzyna I. Morphological Characteristics and Allometric Relationships of Shoot in Two Undergrowth Plants: Polygonatum odoratum and Polygonatum multiflorum. Forests. 2018; 9(12):783. https://doi.org/10.3390/f9120783
Chicago/Turabian StyleTulik, Mirela, Jerzy Karczewski, Natalia Szeliga, Joanna Jura-Morawiec, and Ingeborga Jarzyna. 2018. "Morphological Characteristics and Allometric Relationships of Shoot in Two Undergrowth Plants: Polygonatum odoratum and Polygonatum multiflorum" Forests 9, no. 12: 783. https://doi.org/10.3390/f9120783
APA StyleTulik, M., Karczewski, J., Szeliga, N., Jura-Morawiec, J., & Jarzyna, I. (2018). Morphological Characteristics and Allometric Relationships of Shoot in Two Undergrowth Plants: Polygonatum odoratum and Polygonatum multiflorum. Forests, 9(12), 783. https://doi.org/10.3390/f9120783