Long-Term Soil Productivity in Christmas Tree Farms of Oregon and Washington: A Comparative Analysis between First- and Multi-Rotation Plantations

Abstract
:1. Introduction
2. Experimental Section
2.1. Site Description
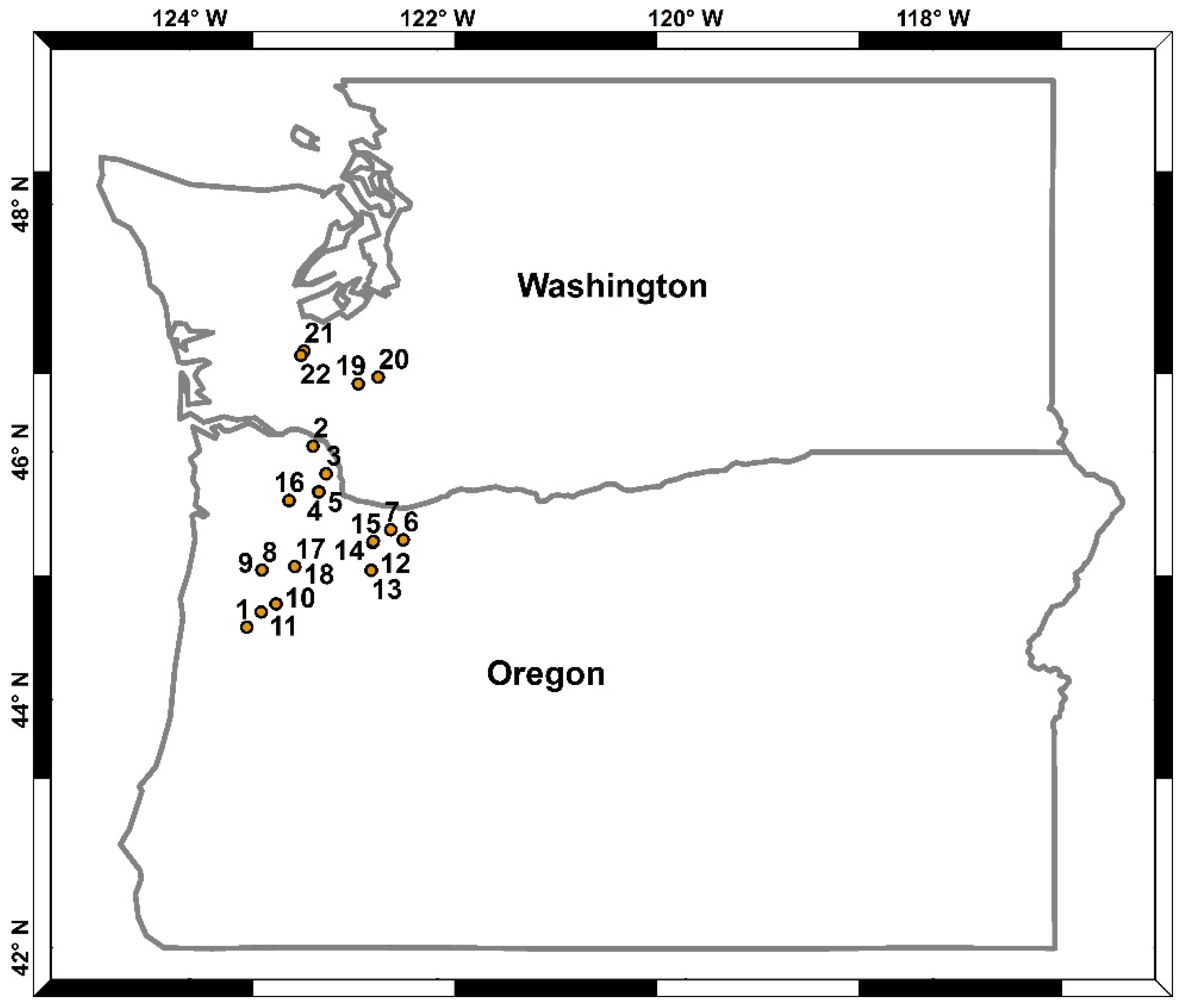

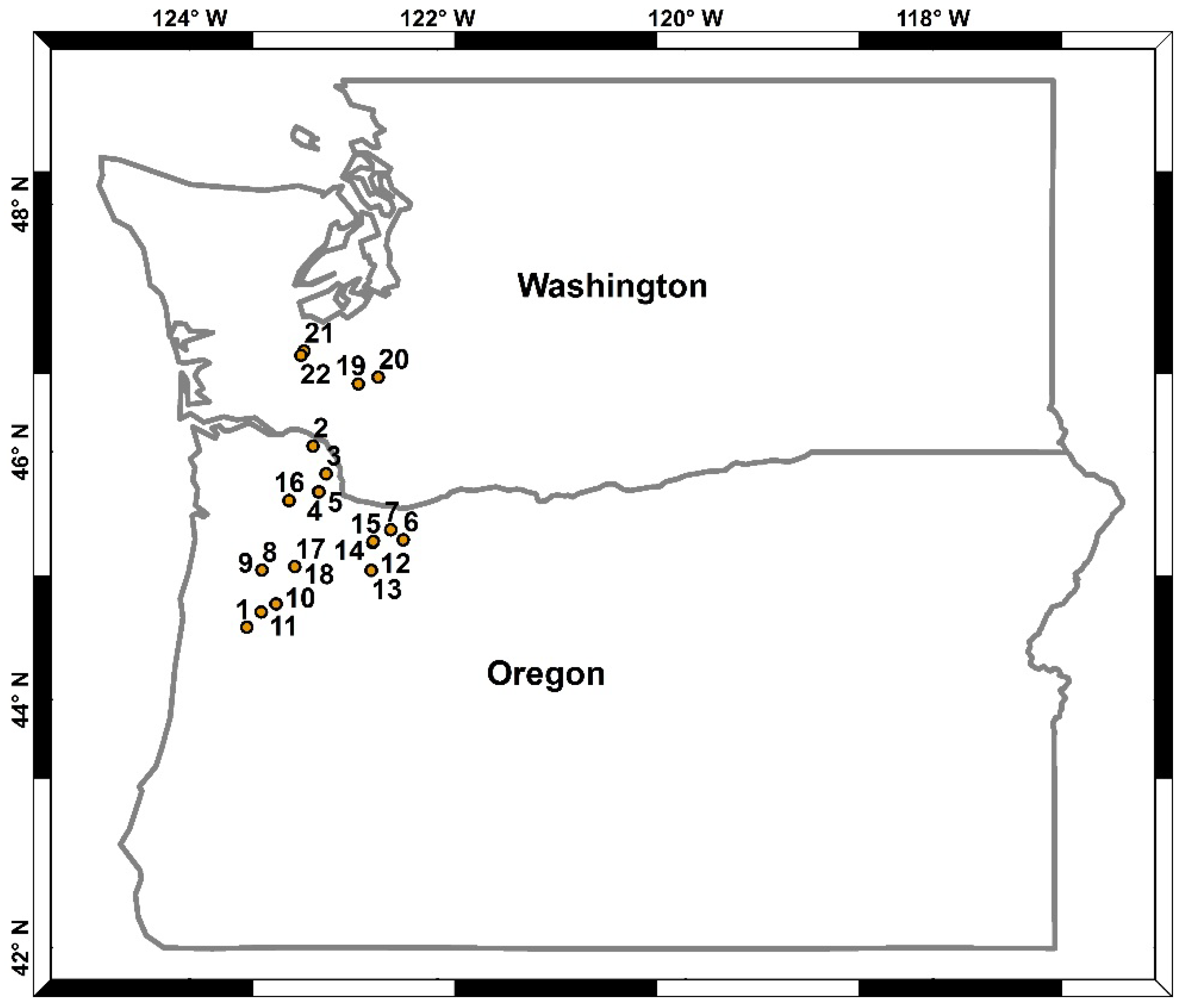{kind=link}
{kind=link}
| Pair | Elev. (m) | MAT (°C) | MAP (mm) | USDA Subgroup Soil Classification | USDA Soil Texture | # of Rot. 1 | Years in Prod. 2 | Years Since Liming | Years Since Tillage |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 201 | 11 | 1853 | Typic Humult | clay loam | 1 | 3 | na | 3 |
| silty clay loam | 4 | 28 | 2 | 2 | |||||
| 2 | 251 | 10 | 1506 | Andic Fragiudepts | gravelly silt loam | 1 | 9 | 1 | na |
| silt loam | 4 | 31 | na | na | |||||
| 3 | 97 | 11 | 1251 | Aquic Haploxerepts | silt loam | 1 | 0 | na | na |
| silt loam | 3 | 15 | na | 16 | |||||
| 4 | 342 | 10 | 1590 | Andic Fragiudepts | silt loam | 1 | 9 | na | 36 |
| silt loam | 6 | 35 | 1.5 | 39 | |||||
| 5 | 382 | 10 | 1599 | Andic Fragiudepts | silt loam | 1 | 3 | 1.5 | 26 |
| silt loam | 4 | 25 | 1.5 | 3 | |||||
| 6 | 348 | 11 | 1545 | Typic Paleudults | silt loam | 1 | 3 | na | 3 |
| loam | 3 | 20 | na | 2 | |||||
| 7 | 94 | 12 | 1213 | Ultic Argixerolls | silty clay loam | 1 | 4 | na | na |
| loam | 4 | 20 | na | 22 | |||||
| 8 | 175 | 11 | 1227 | Xeric Palehumults | silty clay loam | 1 | 2 | na | 2 |
| silty clay loam | 3 | 27 | na | 9 | |||||
| 9 | 175 | 11 | 1227 | Xeric Haplohumults | silty clay loam | 1 | 2 | na | 2 |
| silty clay loam | 3 | 28 | na | 5 | |||||
| 10 | 118 | 11 | 1406 | Ultic Haploxerolls | loam | 1 | 12 | 1.5 | 6 |
| loam | 4 | 22 | na | na | |||||
| 11 | 156 | 11 | 1675 | Xeric Haplohumults | silty clay loam | 1 | 9 | 0 | 10 |
| silty clay loam | 5 | 38 | 2 | na | |||||
| 12 | 475 | 10 | 1755 | Xeric Palehumults | clay loam | 1 | 1 | na | 1 |
| clay loam & silt loam | 4 | 43 | 6 | 6 | |||||
| 13 | 475 | 10 | 1755 | Xeric Palehumults | clay loam | 1 | 1 | na | |
| clay loam & silt loam | 3 | 23 | na | 6 | |||||
| 14 | 245 | 11 | 1343 | Xeric Palehumults | silt loam & clay loam | 1 | 1 | na | 1 |
| silt loam | 3 | na | na | na | |||||
| 15 | 232 | 11 | 1340 | Xeric Palehumults | silt loam | 1 | 2 | na | 3 |
| clay loam & loam | 4 | na | na | na | |||||
| 16 | 150 | 11 | 1249 | Ultic Haploxeralfs | silt loam | 1 | 6 | na | 7 |
| silt loam | 5 | 36 | 1 | 1 | |||||
| 17 | 326 | 10 | 1320 | Xeric Haplohumults | silty clay loam | 1 | 6 | 0 | 1 |
| silty clay loam | 4 | 31 | na | na | |||||
| 18 | 326 | 10 | 1320 | Xeric Haplohumults | silt loam | 1 | 5 | na | na |
| silt loam | 3 | 30 | na | 19 | |||||
| 19 | 231 | 10 | 1381 | Humic Haploxerands | silt loam | 1 | 1 | na | 1 |
| loam & sandy loam | 3 | 20 | na | 5 | |||||
| 20 | 327 | 10 | 1713 | Humic Haploxerands | silt loam | 1 | 6 | na | 8 |
| silt loam | 4 | na | na | na | |||||
| 21 | 71 | 10 | 1342 | Xeric Palehumults | loam | 1 | 3 | na | 4 |
| silt & silt loam | 3 | na | na | 4 | |||||
| 22 | 147 | 10 | 1359 | Xeric Palehumults | silt loam | 1 | 1 | na | 1 |
| silt loam | 3 | 21 | na | na |
2.2. Soil Sample Collection and Analysis
2.3. Soil Resistance
2.4 Statistical Analysis
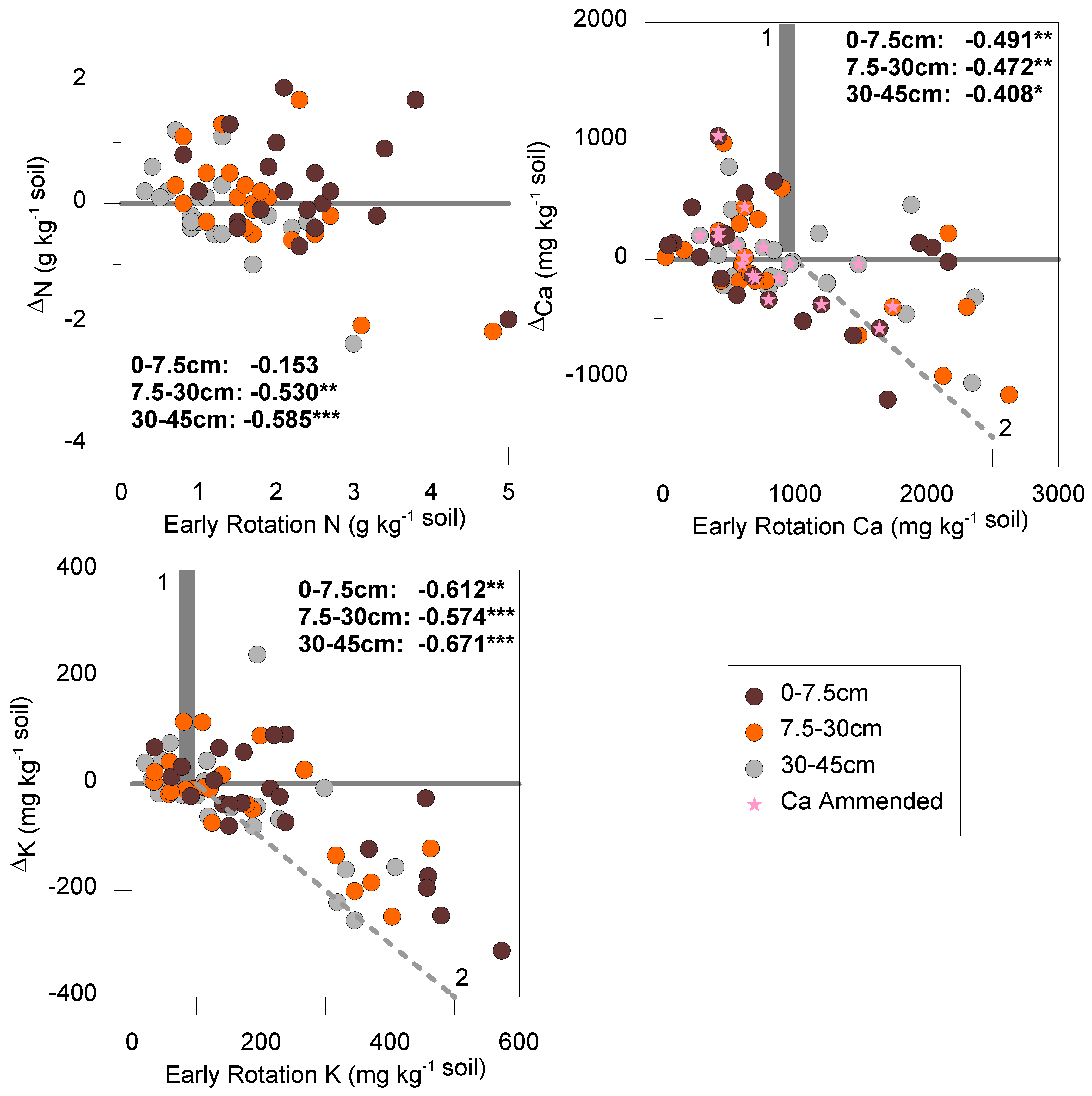
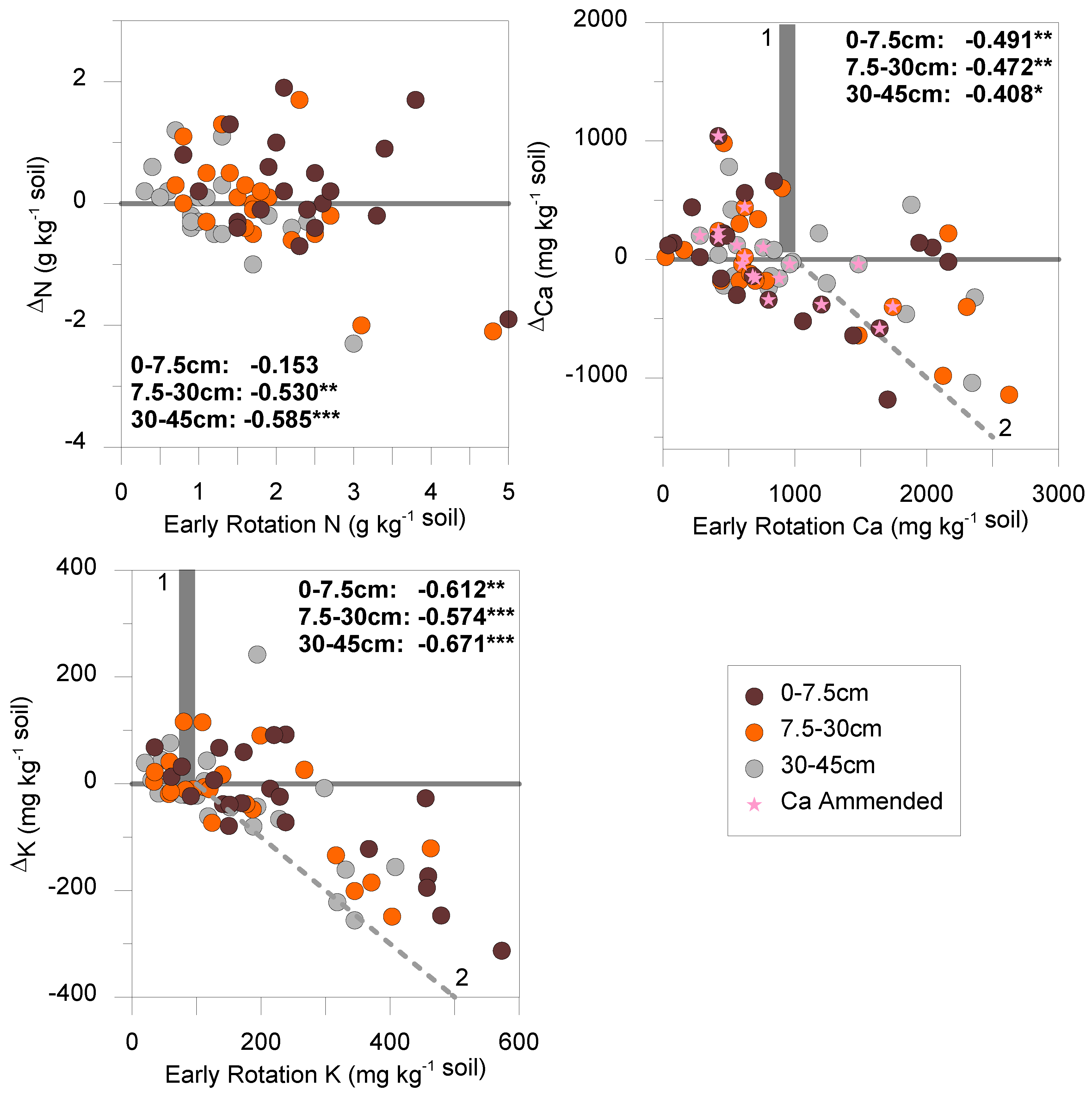
3. Results and Discussion
| Parameter | Depth (cm) | First Rotation | Late Rotation | ||
|---|---|---|---|---|---|
| Average | Range | Average | Range | ||
| pH | 0 to 7.5 | 5.4 | 4.5 to 6.3 | 5.5 | 4.8 to 7.0 |
| 7.5 to 30 | 5.6 | 4.8 to 6.2 | 5.5 | 4.8 to 6.2 | |
| 30 to 45 | 5.6 | 5.0 to 6.1 | 5.7 | 5.1 to 6.0 | |
| P (mg kg−1) | 0 to 7.5 | 28 | 10 to 76 | 28 | 6 to 103 |
| 7.5 to 30 | 21 | 9 to 68 | 21 | 5 to 81 | |
| 30 to 45 | 14 | 6 to 34 | 14 | 4 to 43 | |
| K (mg kg−1) | 0 to 7.5 | 235 | 35 to 573 | 196 | 74 to 428 |
| 7.5 to 30 | 177 | 34 to 463 | 139 | 38 to 342 | |
| 30 to 45 | 154 | 20 to 408 | 130 | 37 to 436 | |
| Ca (mg kg−1) | 0 to 7.5 | 932 | 40 to 2160 | 822 | 80 to 2140 |
| 7.5 to 30 | 1080 | 20 to 2620 | 800 | 40 to 2380 | |
| 30 to 45 | 1030 | 20 to 2360 | 914 | 40 to 2340 | |
| Mg (mg kg−1) | 0 to 7.5 | 143 | 12 to 411 | 150 | 12 to 496 |
| 7.5 to 30 | 156 | 12 to 593 | 148 | 12 to 557 | |
| 30 to 45 | 184 | 12 to 629 | 200 | 12 to 750 | |
| C (g kg−1) | 0 to 7.5 | 37.9 | 11.9 to 76.9 | 36.4 | 13.9 to 107 |
| 7.5 to 30 | 30 | 11.6 to 72.0 | 27.5 | 8.9 to 75 | |
| 30 to 45 | 27.5 | 5.0 to 43.8 | 17.4 | 5.5 to 45.8 | |
| N (g kg−1) | 0 to 7.5 | 2.6 | 0.8 to 5.0 | 2.3 | 1.0 to 5.5 |
| 7.5 to 30 | 1.9 | 1.0 to 4.8 | 1.8 | 0.7 to 4.0 | |
| 30 to 45 | 1.2 | 0.3 to 3.0 | 1.1 | 0.4 to 2.4 | |
| C:N | 0 to 7.5 | 18.4 | 13.3 to 25.5 | 17.3 | 8.1 to 23.0 |
| 7.5 to 30 | 18.6 | 13.2 to 24.9 | 18.2 | 12.4 to 23.6 | |
| 30 to 45 | 18.4 | 11.7 to 24.5 | 18.6 | 11.7 to 24.6 | |
| AS (%) | 0 to 7.5 | 94.3 | 64.6 to 99.7 | 91.2 | 48.7 to 99.2 |
| Depth | ∆pH | ∆P | ∆K | ∆Ca | ∆Mg | ∆C | ∆N | ∆C:N | ∆AS | |
|---|---|---|---|---|---|---|---|---|---|---|
| mg kg−1 | g kg−1 | % | ||||||||
| 0 to 7.5 cm | median | 0.1 | −5.5 | −25.5 | 60.12 | 36.46 | −1.750 | 0.200 | −0.7 | −1.8 |
| stdev | 0.4 | 17.1 | 108.5 | 489.14 | 296.63 | 11.295 | 0.834 | 3.3 | 10.2 | |
| pdiff | 0.861 | 0.648 | 0.110 | 0.062 | 0.483 | 0.247 | 0.056 | 0.421 | 0.011 | |
| pnorm | 0.935 | 0.615 | 0.560 | 0.273 | 0.273 | 0.334 | 0.124 | 0.367 | 0.011 | |
| 7.5 to 30 cm | median | 0.1 | −2.0 | −10.5 | −80.16 | −48.61 | −1.250 | 0.050 | −1.0 | nm |
| stdev | 0.3 | 9.8 | 96.5 | 484.46 | 293.80 | 11.721 | 0.876 | 2.7 | nm | |
| pdiff | 0.178 | 0.753 | 0.252 | 0.005 | 0.475 | 0.187 | 0.329 | 0.626 | nm | |
| pnorm | 0.140 | 0.790 | 0.695 | 0.025 | 0.025 | 0.317 | 0.475 | 0.649 | nm | |
| 30 to 45 cm | median | 0.0 | −0.5 | −20.5 | −30.06 | −18.23 | −3.100 | −0.200 | −0.6 | nm |
| stdev | 0.3 | 4.7 | 104.7 | 360.09 | 218.37 | 11.032 | 0.705 | 3.2 | nm | |
| pdiff | 0.373 | 0.742 | 0.318 | 0.092 | 0.331 | 0.963 | 0.987 | 0.725 | nm | |
| pnorm | 0.242 | 0.647 | 0.814 | 0.367 | 0.367 | 0.458 | 0.448 | 0.700 | nm | |
| Depth | Rotation | 22 | Drip Line | Tire Tracks | Mid-Row | |||
|---|---|---|---|---|---|---|---|---|
| Mean | Max | Mean | Max | Mean | Max | |||
| 0 to 7.5 cm | Early | median | 670 | 1021 | 882 | 1317 | 757 | 1284 |
| stdev | 389 | 620 | 385 | 585 | 393 | 579 | ||
| Late | median | 537 | 860 | 754 | 1073 | 733 | 1112 | |
| stdev | 382 | 655 | 327 | 573 | 345 | 649 | ||
| 7.5 to 30 cm | Early | median | 1926 | 2339 | 2072 | 2420 | 2122 | 3746 |
| stdev | 382 | 368 | 342 | 354 | 331 | 640 | ||
| Late | median | 1918 | 2325 | 2067 | 2388 | 2116 | 3878 | |
| stdev | 466 | 473 | 429 | 445 | 426 | 744 | ||
| 30 to 45 cm | Early | median | 2308 | 2455 | 2309 | 2525 | 2387 | 2616 |
| stdev | 383 | 423 | 410 | 395 | 405 | 435 | ||
| Late | median | 2213 | 2412 | 2260 | 2461 | 2320 | 2442 | |
| stdev | 406 | 473 | 395 | 443 | 411 | 439 | ||
| Depth | Rotation | Drip Line | Tire Tracks | Mid-Row | |||
|---|---|---|---|---|---|---|---|
| Mean | Max | Mean | Max | Mean | Max | ||
| 0 to 7.5 cm | median | −91 | −120 | −80 | −63 | −159 | −174 |
| stdev | 490 | 811 | 477 | 786 | 490 | 855 | |
| pdiff | 0.478 | 0.518 | 0.458 | 0.498 | 0.384 | 0.582 | |
| 7.5 to 30 cm | median | −21 | −39 | −59 | −80 | 15 | −186 |
| stdev | 408 | 377 | 370 | 343 | 370 | 811 | |
| pdiff | 0.718 | 0.350 | 0.539 | 0.245 | 0.478 | 0.439 | |
| 30 to 45 cm | median | −76 | −36 | −83 | −66 | −59 | −122 |
| stdev | 335 | 339 | 389 | 410 | 379 | 388 | |
| pdiff | 0.070 | 0.245 | 0.478 | 0.814 | 0.439 | 0.334 | |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- USDA. Agriculture Census; United States Department of Agriculture: Washington, DC, USA, 2012. [Google Scholar]
- Morris, L.A.; Miller, R.E. Evidence for long-term productivity change as provided by field trials. In Impacts of Forest Harvesting on Long-Term Site Productivity; Dyck, W.J., Cole, D.W., Comerford, N.B., Eds.; Chapman and Hall: London, UK, 1994; pp. 41–80. [Google Scholar]
- Richardson, B.; Skinner, M.F.; West, G. The role of forest productivity in defining the sustainability of plantation forests in New Zealand. For. Ecol. Manag. 1999, 122, 125–137. [Google Scholar]
- Burger, J.A.; Kelting, D.L. Using soil quality indicators to assess forest stand management. For. Ecol. Manag. 1999, 122, 155–166. [Google Scholar]
- Greacen, E.; Barley, K.; Farrell, D. The mechanics of root growth in soils with particular reference to the implications for root distribution. In Root Growth; Whittington, W.J., Ed.; Butterworths: London, UK, 1969; pp. 256–269. [Google Scholar]
- Greacen, E.L.; Sands, R. Compaction of Forest Soils. A Review. Aust. J. Soil Res. 1980, 18, 163–189. [Google Scholar]
- Sands, R.; Greacen, E.; Gerard, C. Compaction of sandy soils in radiata pine forests. I. A penetrometer study. Soil Res. 1979, 17, 101–113. [Google Scholar]
- Amaranthus, M.P.; Page-Dumroese, D.; Harvey, A.; Cazares, E.; Bednar, L.F. Soil Compaction and Organic Matter Affect Conifer Seedling Nonmychorizal and Ectomychorrrhizal Root Tip Abundance and Diversity; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1996. [Google Scholar]
- Frank, R.; Sirons, G.; Anderson, G. Atrazine: The impact of persistent residues in soil on susceptible crop species. Can. J. Soil Sci. 1983, 63, 315–325. [Google Scholar]
- Hart, J.; Landgren, C.; Fletcher, R.; Bondi, M.; Withrow-Robinson, B.; Chastagner, G. Christmas Tree Nutrient Management Guide: Western Oregon and Washington; Oregon State Univeristy Extension Service, EM 8856-E: Corvallis, OR, USA, 2009. [Google Scholar]
- Daly, C.; Gibson, W.P.; Taylor, G.H.; Johnson, G.L.; Pasteris, P. A knowledge-based approach to the statistical mapping of climate. Clim. Res. 2002, 22, 99–113. [Google Scholar]
- Thomas, G.W. Soil pH and Soil Acidity. In Methods of Soil Analysis; Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America and America Society of Agronomy: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Sumner, M.; Miller, W.; Sparks, D.; Page, A.; Helmke, P.; Loeppert, R.; Soltanpour, P.; Tabatabai, M.; Johnston, C. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis; Part 3 Chemical Methods; Soil Science Society of America and America Society of Agronomy: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar]
- Olsen, S.; Sommers, L. Phosphorus. In Methods of Soils Analysis; Part II, 2nd ed.; Page, A.L., Ed.; Agronomy Monograph 9; ASA/SSA: Madison, WI, USA, 1982. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle Size Analysis. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 383–412. [Google Scholar]
- Worrell, R.; Hampson, A. The influence of some forest operations on the sustainable management of forest soils—a review. Forestry 1997, 70, 61–85. [Google Scholar]
- Brewer, L.; Fletcher, R.; Hart, J.; Landgren, C.; Webster, S. Changes in Soil Conditions over Multiple-Tree Cropping Cycles- Final Report to Oregon Department of Agriculture; Oregon State University Extension Service: Corvallis, OR, USA, 2012. [Google Scholar]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar]
- Vazquez, L.; Myhre, D.L.; Hanlon, E.A.; Gallagher, R.N. Soil penetrometer resistance and bulk density relationships after long-term no tillage. Commun. Soil Sci. Plant Anal. 1991, 22, 2101–2117. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatten, J.; Landgren, C.; Hart, J. Long-Term Soil Productivity in Christmas Tree Farms of Oregon and Washington: A Comparative Analysis between First- and Multi-Rotation Plantations. Forests 2014, 5, 2581-2593. https://doi.org/10.3390/f5102581
Hatten J, Landgren C, Hart J. Long-Term Soil Productivity in Christmas Tree Farms of Oregon and Washington: A Comparative Analysis between First- and Multi-Rotation Plantations. Forests. 2014; 5(10):2581-2593. https://doi.org/10.3390/f5102581
Chicago/Turabian StyleHatten, Jeff, Chal Landgren, and John Hart. 2014. "Long-Term Soil Productivity in Christmas Tree Farms of Oregon and Washington: A Comparative Analysis between First- and Multi-Rotation Plantations" Forests 5, no. 10: 2581-2593. https://doi.org/10.3390/f5102581
APA StyleHatten, J., Landgren, C., & Hart, J. (2014). Long-Term Soil Productivity in Christmas Tree Farms of Oregon and Washington: A Comparative Analysis between First- and Multi-Rotation Plantations. Forests, 5(10), 2581-2593. https://doi.org/10.3390/f5102581