Predicting Understory Species Richness from Stand and Management Characteristics Using Regression Trees

 ,
,
Abstract
1. Introduction
2. Experimental Section
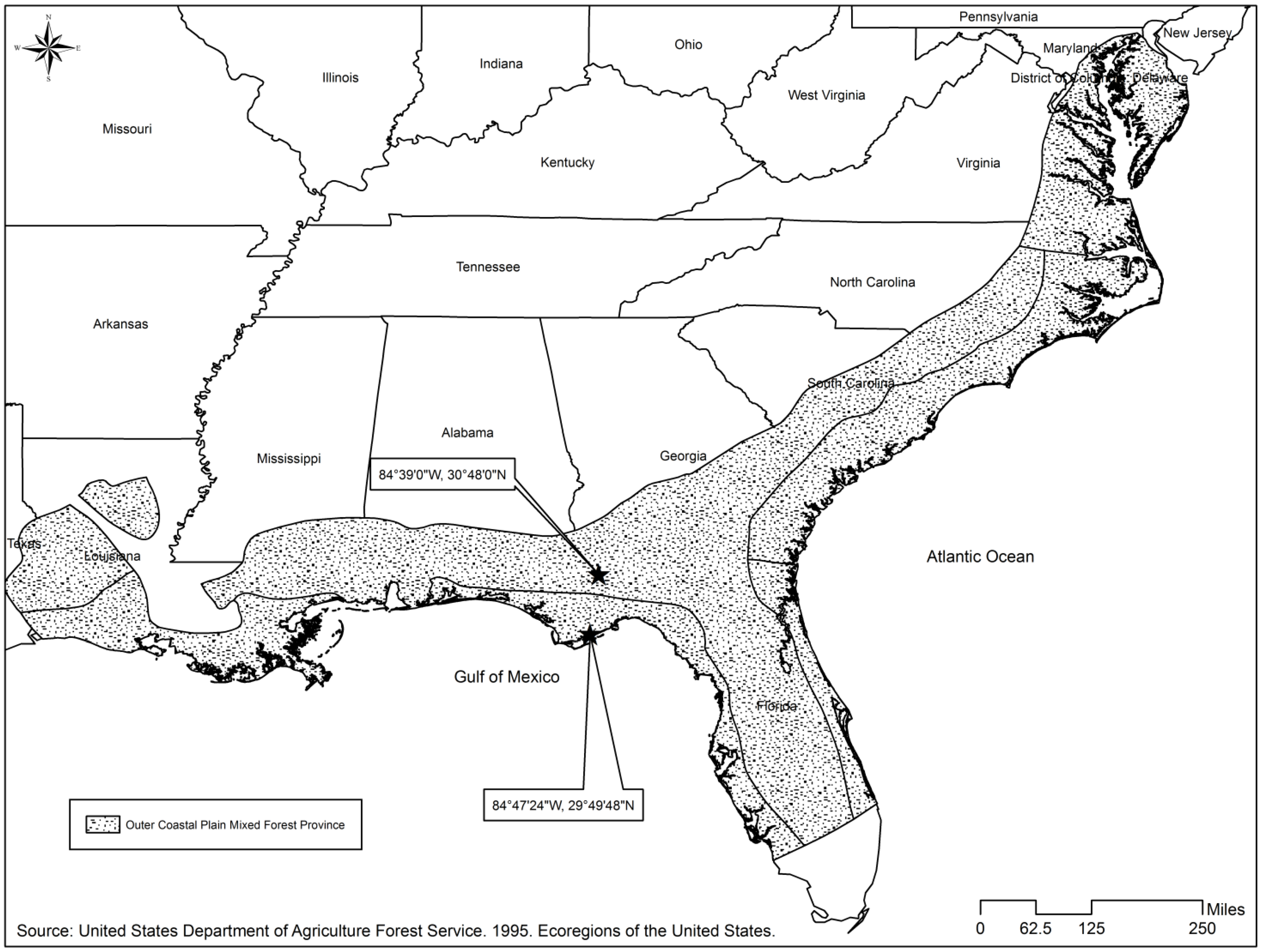

{kind=link}
{kind=link}
{kind=link}
| Physiographic zones | Selected Studies |
|---|---|
| Upper coastal plain (UCP) | [12,30,31,32,33,34,35,36] |
| Middle coastal plain(MCP) | [21,37,38,39,40,41] |
| Lower coastal plain (LCP) | [16,37,42,43,44,45,46,47,48,49,50,51,52] |

| Variables used in the model | Categories | Range/Frequency |
|---|---|---|
| Species richness (number/4 m2) | NA | 0–53 |
| Stand age (years) | NA | 1–85 |
| Physiographic zone | Xeric site (Sandhills type), Mesic (Flatwood type), | (52, 117, 6) * |
| and Hydric (Swamp) | ||
| Stand Origin | Plantation and Natural | (100, 75) * |
| Fire | Yes (1) and No (0) | (146, 29) * |
| Time since the last fire (years) | NA | 0.5 to >50 |
| Site preparation | Yes (1) and No (0) | (79, 96) * |
| Time since site preparation | NA | 1 to >50 |
| (years) | ||
| Herbicide and Fertilizer | Yes (1) and No (0) | (56, 119) * |
| Time since herbicide and | NA | 1 to >50 |
| fertilizer (years) | ||
| Thinning | Yes (1) and No (0) | (38, 136) * |
| Grazing | Yes (1) and No (0) | (23, 152) * |
| Forest types | Longleaf pine, Longleaf/Oak, Slash pine, Slash pine/Oak, Loblolly pine, Loblolly pine/Oak | (67, 25, 21, 5, 47, 10) * |




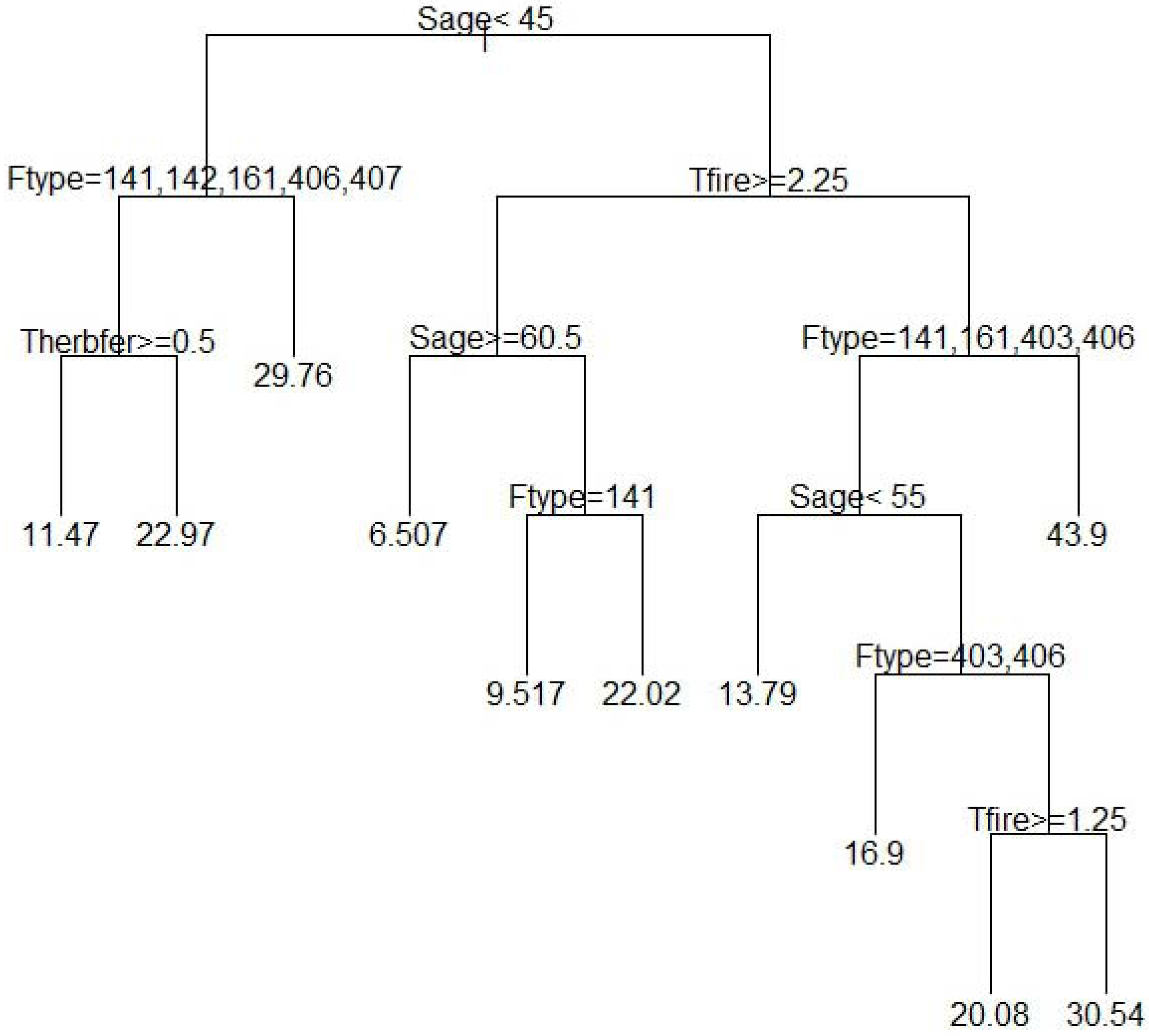
3. Results and Discussion
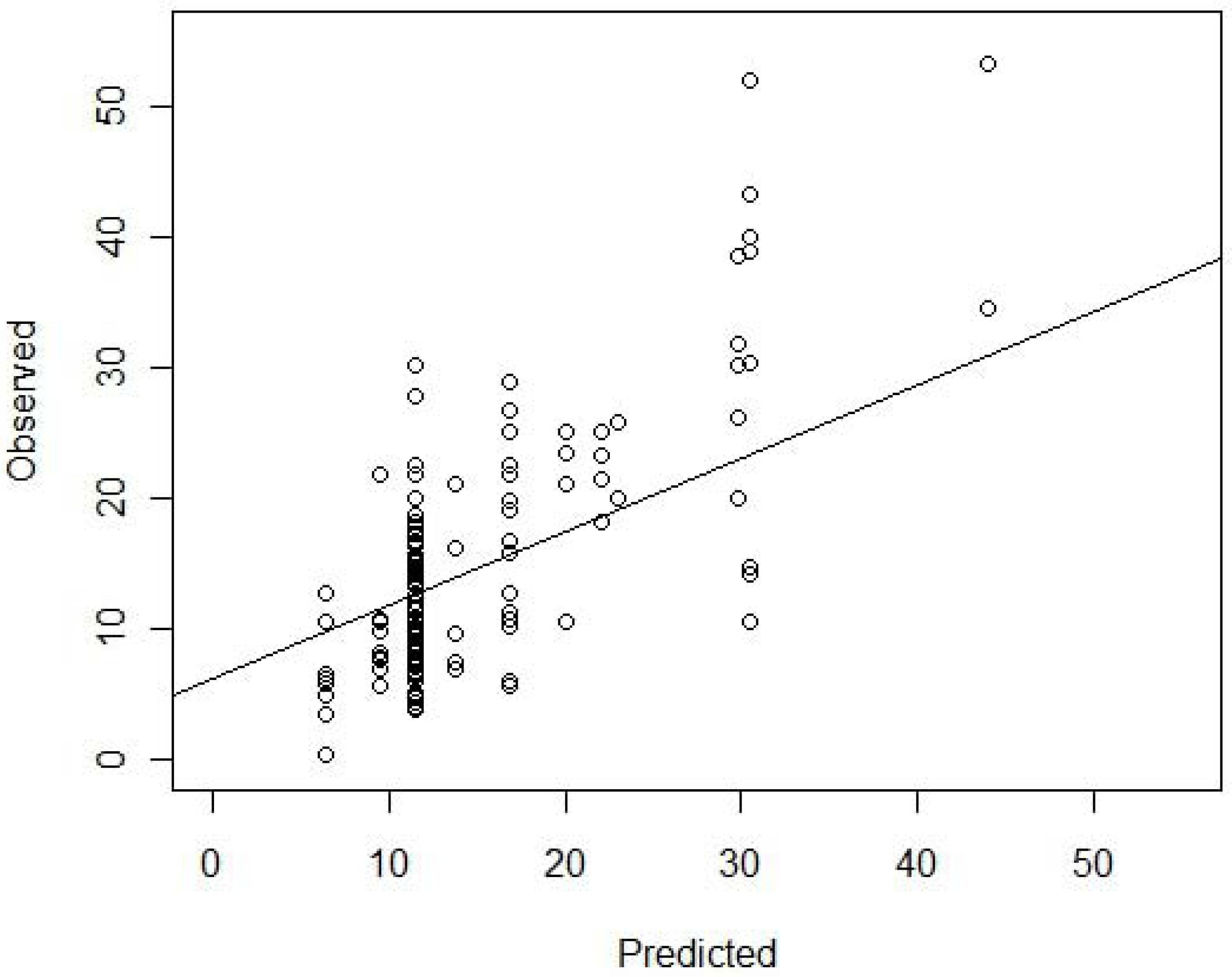
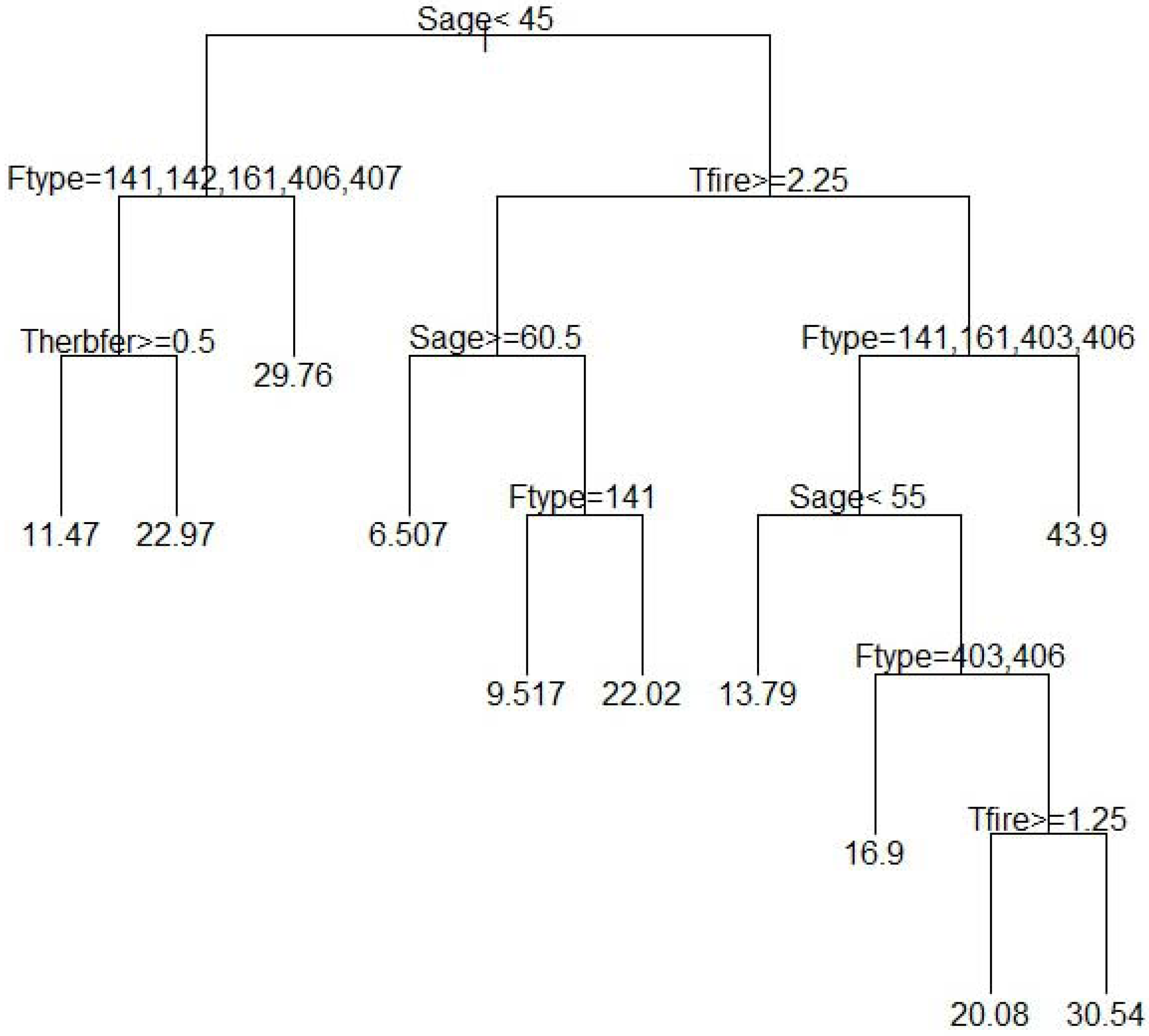
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Sader, S.; Hoppus, M.; Metzler, J.; Jin, S. Perspectives of Maine forest cover change from Landsat imagery and Forest Inventory Analysis (FIA). J. Forest. 2005, 103, 299–303. [Google Scholar]
- Ciais, P.; Schelhaas, M.J.; Zaehle, S.; Piao, S.L.; Cescatti, A.; Liski, J.; Luyssaert, S.; Le-Marie, G.; Schulze, E.D.; Bouriaud, O.; Freibauer, A.; Valentini, R.; Nabuurs, G.J. Carbon accumulation in European forests. Nat. Geosci. 2008, 1, 425–429. [Google Scholar] [CrossRef]
- Yemshanov, D.; McKenney, D.W.; de Groot, P.; Haugen, D.; Sidders, D.; Joss, B. A bioeconomic approach to assess the impact of an alien invasive insect on timber supply and harvesting: A case study with Sirex noctilio in eastern Canada. Can. J. For. Res. 2009, 39, 154–168. [Google Scholar] [CrossRef]
- Heath, L.S.; Smith, J.E.; Woodall, C.W.; Azuma, D.L.; Waddell, K.L. Carbon stocks on forestland of the United States, with emphasis on USDA Forest Service Ownership. Ecosphere 2011, 2, 1–21. [Google Scholar]
- Managing Forest Ecosystems, National Forest Inventories: Contributions to Forest Biodiversity Assessments; Chirici, G.; Winter, G.; McRoberts, R.E. (Eds.) Springer: Heidelberg, Germany, 2011; p. 206.
- Allen, A.W.; Bernal, Y.K.; Moulton, R.J. Pine plantations and Wildlife in the Southeastern United States: An Assessment of Impacts and Opportunities; US Department of the Interior National Biological Service Information and Technology Report 3; US Department of Interior: Washington, DC, USA, 1996; p. 32. [Google Scholar]
- Baker, J.C.; Hunter, W.C. Effects of forest management on terrestrial ecosystems. In Southern Forest Resource Assessment; General Technical Report SRS-53; Wear, D.N., Greis, J.G., Eds.; USDA Forest Service Southern Research Station: Ashville, NC, USA, 2002; pp. 91–112. [Google Scholar]
- Moore, S.E.; Allen, H.L. Plantation forestry. In Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 1999; pp. 400–433. [Google Scholar]
- Palik, B.; Engstrom, R.T. Species composition. In Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 1999; pp. 65–94. [Google Scholar]
- Twedt, D.J.; Tirpak, J.M.; Jones-Farrand, D.T.; Thompson, F.R., III; Uihlein, W.B., III; Fitzgerald, J.A. Change in avian abundance predicted from regional forest inventory data. For. Ecol. Manage. 2010, 260, 241–1250. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manage. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Hedman, C.W.; Grace, S.L.; King, S.E. Vegetation composition and structure of southern coastal plain pine forests: an ecological comparison. For. Ecol. Manage. 2000, 134, 233–247. [Google Scholar] [CrossRef]
- Moore, W.H.; Swindel, B.F.; Terry, W.S. Vegetative response to prescribed fire in a north Florida flatwoods forest. J. Range Manage. 1982, 35, 386–389. [Google Scholar] [CrossRef]
- Iglay, R.B.; Leopold, B.D.; Miller, D.A.; Burger, L.W., Jr. Effect of plant community composition on plant response to fire and herbicide treatments. For. Ecol. Manage. 2010, 260, 543–548. [Google Scholar] [CrossRef]
- Swindel, B.F.; Conde, L.F.; Smith, J.E. Effects of forest regeneration practices on plant diversity and succession in Florida ecosystems. In Annual Symposium Proceedings on Impacts of Intensive Management Practices; School of Forest Resources and Conservation, University of Florida: Gainesville, FL, USA, 1982; pp. 5–15. [Google Scholar]
- Swindel, B.F.; Conde, L.F.; Smith, J.E. Successional changes in Pinus elliottii plantation following two regeneration treatments. Can. J. For. Res. 1986, 16, 630–636. [Google Scholar] [CrossRef]
- Conde, L.F.; Swindel, B.F.; Smith, J.E. Plant species cover, frequency, and biomass: Early responses to clearcutting, chopping, and bedding in Pinus elliottii flatwoods. For. Ecol. Manage. 1983, 6, 307–318. [Google Scholar] [CrossRef]
- Conde, L.F.; Swindel, B.F.; Smith, J.E. Plant species cover, frequency, and biomass: Early responses to clearcutting, burning, windrowing, discing, and bedding in Pinus elliottii flatwoods. For. Ecol. Manage. 1983, 6, 319–331. [Google Scholar] [CrossRef]
- Miller, K.V.; Miller, J.H. Forestry herbicide influences on biodiversity and wildlife habitat in southern forests. Wildlife Soc. B 2004, 32, 1049–1060. [Google Scholar] [CrossRef]
- Moore, W.F. Effects of Chemical Site Preparation on Vegetative, Small Mammal, and Songbird Communities in the Georgia Sandhills. Master’s Thesis, University of Georgia, Athens, GA, USA, 1996; p. 208. [Google Scholar]
- Miller, J.H.; Boyd, R.S.; Edwards, M.B. Floristic diversity, stand structure, and composition 11 years after herbicide site preparation. Can. J. For. Res. 1999, 29, 1073–1083. [Google Scholar] [CrossRef]
- Timilsina, N.; Escobedo, F.; Cropper, W.P., Jr.; Abd-Elrahman, A.; Brandeis, T.; Delphin, S.; Lambert, S. A framework for identifying carbon hotspots and forest management drivers. J. Environ. Manage. 2012. [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. Plant Sci. 2006, 25, 381–397. [Google Scholar]
- Hagar, J.C. Wildlife species associated with non-coniferous vegetation in Pacific Northwest conifer forests: A review. For. Ecol. Manage. 2007, 246, 108–122. [Google Scholar] [CrossRef]
- Marsland, S. Machine Learning: An Algorithmic Perspective; CRC Press: Boca Raton, FL, USA, 2009; p. 406. [Google Scholar]
- McNab, W.H.; Avers, P.E. USDA Forest Service WO-WSA-5, 1994. Ecological Subregions of the United States. Available online: http://www.fs.fed.us/land/pubs/ecoregions/ (accessed on 4 September 2012).
- Noss, R.F. Longleaf pine and wiregrass: Keystone components of an endangered ecosystem. Nat. Areas J. 1989, 9, 211–213. [Google Scholar]
- Peet, R.K.; Allard, D.J. Longleaf Pine Vegetation of the Southern Atlantic and Eastern Gulf Coast Regions: A Preliminary Classification. P. In Proceedings of the 18th Tall Timbers Fire Ecology Conference on the Longleaf Pine Ecosystem: Ecology, Restoration, and Management, Tallahassee, FL, USA, 30 May–2 June, 1991; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; pp. 45–82. [Google Scholar]
- Myers, R. Fire and the dynamic relationship between Florida sandhill and sand pine scrub vegetation. B. Torrey Bot. Club. 1985, 112, 241–252. [Google Scholar] [CrossRef]
- Archer, J.K.; Miller, D.L.; Tanner, G.W. Changes in understory vegetation and soil characteristics following silvicultural activities in a southeastern mixed forest. J. Torrey Bot. Soc. 2007, 134, 489–504. [Google Scholar] [CrossRef]
- Hebb, E.A.; Clewell, A.F. A remnant stand of old-growth slash pine in the Florida panhandle. B. Torrey Bot. Club 1976, 103, 1–9. [Google Scholar] [CrossRef]
- Brewer, J.S. Patterns of plant species richness in a wet slash-pine (Pinus elliottii) savanna. J. Torrey Bot. Soc. 1998, 125, 216–224. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Damschen, E.I. Land-use history, historical connectivity, and land management interact to determine longleaf pine woodland understory richness and composition. Ecography 2011, 34, 257–266. [Google Scholar] [CrossRef]
- Harrington, T.B.; Edwards, M.B. Understory vegetation, resource availability, and litterfall responses to pine thinning and woody vegetation control in longleaf pine plantations. Can. J. For. Res. 1999, 29, 1055–1064. [Google Scholar] [CrossRef]
- Harrington, T.B. Overstory and understory relationships in longleaf pine plantations 14 years after thinning and woody control. Can. J. For. Res. 2011, 41, 2301–2314. [Google Scholar] [CrossRef]
- Provencher, L.; Litt, A.R.; Gordon, D.R. Predictors of species richness in northwest Florida longleaf pine sandhills. Conserv. Biol. 2003, 17, 1660–1671. [Google Scholar] [CrossRef]
- Miller, J.H.; Zutter, B.R.; Newbold, R.A.; Edwards, M.B.; Zedaker, S.M. Stand dynamics and plant associates of loblolly pine plantations to midrotation after early intensive vegetation management—A southeastern United States regional study. South. J. Appl. For. 2003, 27, 1–16. [Google Scholar]
- Blake, P.M.; Hurst, G.A.; Terry, T.A. Response of vegetation and deer forage following application of hexazinone. South. J. Appl. For. 1987, 11, 176–180. [Google Scholar]
- Boyd, R.S.; Freeman, J.D.; Miller, J.H.; Edwards, M.B. Forest herbicide influences on floristic diversity seven years after broadcast pine release treatments in central Georgia, USA. New For. 1995, 10, 17–37. [Google Scholar]
- Brockway, D.G.; Lewis, C.E. Influence of deer, cattle grazing and timber harvest on plant species diversity in a longleaf pine bluestem ecosystem. For. Ecol. Manage. 2003, 175, 49–69. [Google Scholar] [CrossRef]
- Kush, J.S.; Meldahl, R.S.; Boyer, W.D. Understory plant community response after 23 years of hardwood control treatments in natural longleaf pine (Pinus palustris) forests. Can J. For. Res. 1999, 29, 1047–1054. [Google Scholar] [CrossRef]
- Maliakal, S.K.; Menges, E.S.; Denslow, J.S. Community composition and regeneration of Lake Wales Ridge wiregrass flatwoods in relation to time-since-fire. J. Torrey Bot. Soc. 2000, 127, 125–138. [Google Scholar] [CrossRef]
- Lane, V.R.; Miller, K.V.; Castleberry, S.B.; Miller, D.A.; Wigley, T.B.; Marsh, G.M.; Mihalco, R.L. Plant community responses to a gradient of site preparation intensities in pine plantations in the Coastal Plain of North Carolina. For. Ecol. Manage. 2011, 262, 370–378. [Google Scholar] [CrossRef]
- Freeman, J.E.; Jose, S. The role of herbicide in savanna restoration: Effects of shrub reduction treatments on the understory and overstory of a longleaf pine flatwoods. For. Ecol. Manage. 2009, 257, 978–986. [Google Scholar] [CrossRef]
- Platt, W.J.; Carr, S.M.; Reilly, M.; Fahr, J. Pine savanna overstorey influences on ground-cover biodiversity. Appl. Veg. Sci. 2006, 9, 37–50. [Google Scholar] [CrossRef]
- Keddy, P.A.; Smith, L.; Campbell, D.R.; Clark, M.; Montz, G. Patterns of herbaceous plant diversity in southeastern Louisiana pine savannas. Appl. Veg. Sci. 2006, 9, 17–26. [Google Scholar] [CrossRef]
- Brockway, D.G.; Lewis, C.E. Long-term effects of dormant-season prescribed fire on plant community diversity, structure and productivity in a longleaf pine wiregrass ecosystem. For. Ecol. Manage. 1997, 96, 167–183. [Google Scholar] [CrossRef]
- Kirkman, K.; Mitchell, R.J.; Helton, R.C.; Drew, M.B. Productivity and species richness across an environmental gradient in a fire-dependent ecosystem. Am. J. Bot. 2001, 88, 2119–2128. [Google Scholar]
- Glitzenstein, J.S.; Streng, D.R.; Wade, D.D. Fire frequency effects on Longleaf Pine (Pinus palustris P. Miller) vegetation in South Carolina and Northeast Florida USA. Nat. Area J. 2003, 23, 22–37. [Google Scholar]
- Ruth, A.D.; Miller, D.L.; Jose, S.; Long, A. Effects of reintroduction of fire into fire suppressed coastal scrub and longleaf pine communities along the lower gulf coastal plain. Nat. Areas J. 2007, 271, 332–344. [Google Scholar]
- Beckage, B.; Stout, I.J. Effects of repeated burning on species richness in a Florida pine savanna: A test of the intermediate disturbance hypothesis. J. Veg. Sci. 2000, 11, 113–122. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Christensen, N.L. Herb-layer response to burning in pine flatwoods of the lower coastal plain of South Carolina. B. Torrey Bot. Club. 1986, 113, 42–45. [Google Scholar] [CrossRef]
- Fridley, J.D.; Peet, P.K.; Wentworth, T.R.; White, P.S. Connecting fine- and broad-scale species-area relationships of southeastern U.S. flora. Ecology 2005, 86, 1172–1177. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.G. Classification and Regression Trees; Wadsworth International Group: Belmont, CA, USA, 1984; p. 358. [Google Scholar]
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: Cambridge, UK, 1996; p. 415. [Google Scholar]
- De’ath, G.; Fabricus, K.E. Classification and regression trees: A powerful yet simple technique for ecological data analysis. Ecology 2000, 81, 3178–3192. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; McGraw-Hill: New York, NY, USA, 1990; p. 544. [Google Scholar]
- Holland, D.N.; Lilieholm, R.J.; Roberts, D.W. Economic trade-offs of managing forests for timber production and vegetative diversity. Can. J. For. Res. 1994, 24, 1260–1265. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Timilsina, N.; Cropper, W.P., Jr.; Escobedo, F.J.; Lima, J.M.T. Predicting Understory Species Richness from Stand and Management Characteristics Using Regression Trees. Forests 2013, 4, 122-136. https://doi.org/10.3390/f4010122
Timilsina N, Cropper WP Jr., Escobedo FJ, Lima JMT. Predicting Understory Species Richness from Stand and Management Characteristics Using Regression Trees. Forests. 2013; 4(1):122-136. https://doi.org/10.3390/f4010122
Chicago/Turabian StyleTimilsina, Nilesh, Wendell P. Cropper, Jr., Francisco J. Escobedo, and Joanna M. Tucker Lima. 2013. "Predicting Understory Species Richness from Stand and Management Characteristics Using Regression Trees" Forests 4, no. 1: 122-136. https://doi.org/10.3390/f4010122
APA StyleTimilsina, N., Cropper, W. P., Jr., Escobedo, F. J., & Lima, J. M. T. (2013). Predicting Understory Species Richness from Stand and Management Characteristics Using Regression Trees. Forests, 4(1), 122-136. https://doi.org/10.3390/f4010122