
Consecutive Pruning Enhances Leaf Flavonoids, Leaf Yield, and Cutting Rooting in Ginkgo biloba


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Pruning
2.2. Morphological Observation
2.3. Physiological Measurement
2.4. Determination of Flavonoid Content
2.5. Total RNA Extraction and qRT-PCR Experiment
2.6. Rooting of Stem Cutting
2.7. Statistical Analysis
3. Results
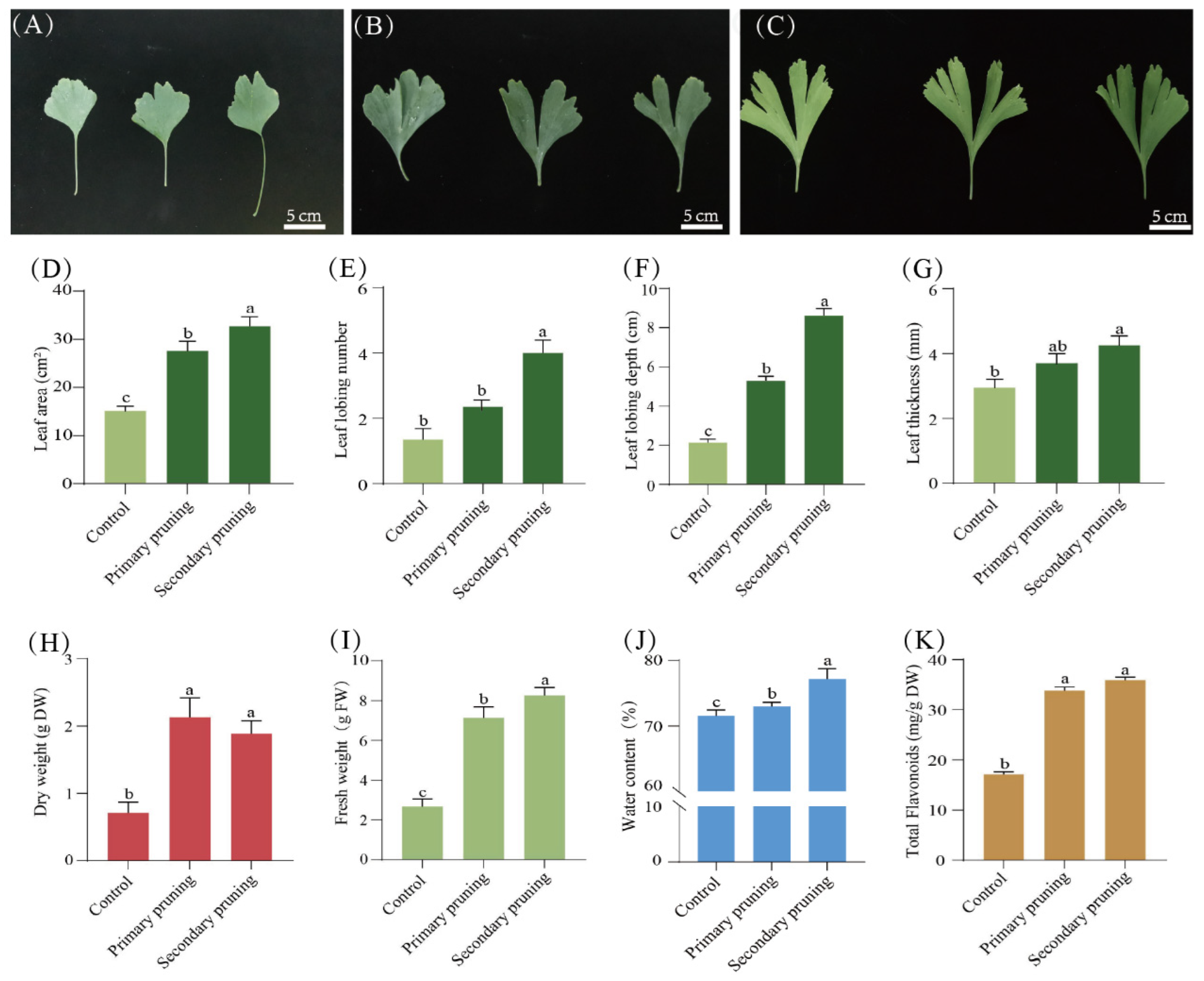
3.1. Effects of Various Pruning Methods on Plant Growth and Leaf Development
3.2. Effects of Pruning Times on Plant Growth and Leaf Development
3.3. Expression Changes of Genes Related to Bud Sprouting and Flavonoid Biosynthesis in Pruned Seedlings
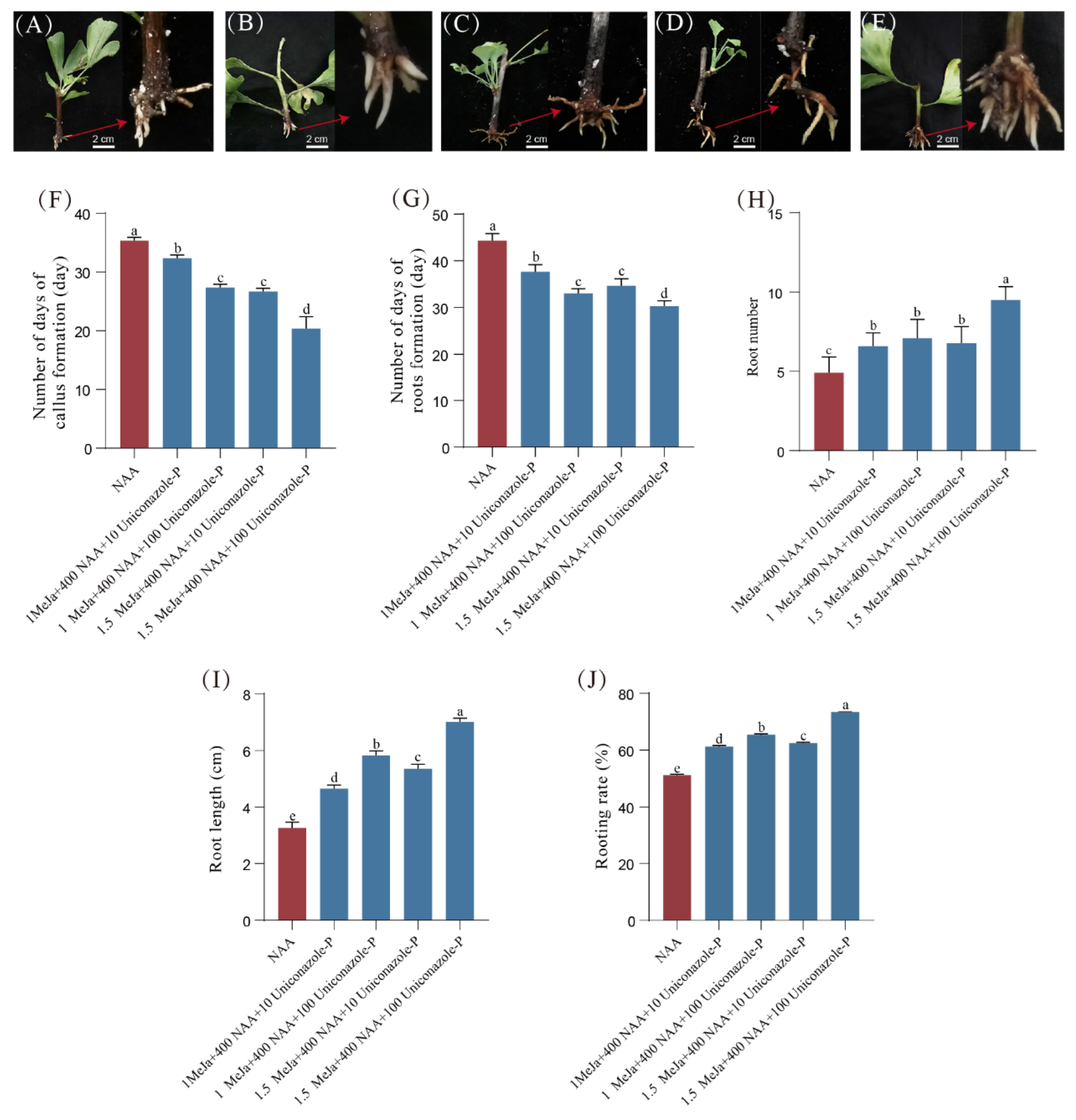
3.4. Repetitive Pruning Promotes the Rooting of Cuttings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.F.; Huang, L.B.; Zhong, Y.B.; Zhou, Q.H.; Wang, H.L.; Zheng, G.Q.; Lin, Y. An overview of systematic reviews of Ginkgo biloba extracts for mild cognitive impairment and dementia. Front. Aging Neurosci. 2016, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Akaberi, M.; Baharara, H.; Amiri, M.S.; Moghadam, A.T.; Sahebkar, A.; Emami, S.A. Ginkgo biloba: An update review on pharmacological, ethnobotanical, and phytochemical studies. Pharmacol. Res.-Mod. Chin. Med. 2023, 9, 100331. [Google Scholar] [CrossRef]
- Mao, D.Y.; Zhong, L.; Zhao, X.Y.; Wang, L. Function, biosynthesis, and regulation mechanisms of flavonoids in Ginkgo biloba. Fruit Res. 2023, 3, 0018. [Google Scholar] [CrossRef]
- Guo, J.; Zhou, X.; Wang, T.L.; Wang, G.B.; Cao, F.L. Regulation of flavonoid metabolism in ginkgo leaves in response to different day-night temperature combinations. Plant. Physiol. Bioch. 2020, 147, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Rethinking Ginkgo biloba L.: Medicinal uses and conservation. Pharmacogn. Rev. 2015, 9, 140–148. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.F.; Gan, Q.; Liu, S.A.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Wang, Q.J.; Jiang, Y.; Mao, X.Y.; Yu, W.W.; Lu, J.K.; Wang, L. Integration of morphological, physiological, cytological, metabolome and transcriptome analyses reveal age inhibited accumulation of flavonoid biosynthesis in Ginkgo biloba leaves. Ind. Crop. Prod. 2022, 187, 115405. [Google Scholar] [CrossRef]
- Lu, Z.G.; Zhu, L.K.; Lu, J.K.; Shen, N.; Wang, L.; Liu, S.A.; Wang, Q.J.; Yu, W.W.; Noguchi, H.K.; Li, W.X.; et al. Rejuvenation increases leaf biomass and flavonoid accumulation in Ginkgo biloba. Hortic Res. 2022, 9, uhab018. [Google Scholar] [CrossRef]
- Vellas, B.; Coley, N.; Ousset, P.J.; Berrut, G.; Dartigues, J.F.; Dubois, B.; Grandjean, H.; Pasquier, F.; Piette, F.; Robert, P.; et al. Long-term use of stand-ardised Ginkgo biloba extract for the prevention of Alzheimer’s disease (GuidAge): A randomised placebo-controlled trial. Lancet Neurol. 2012, 11, 851–859. [Google Scholar] [CrossRef]
- Herrschaft, H.; Nacu, A.; Likhachev, S.; Sholomov, I.; Hoerr, R.; Schlaefke, S. Ginkgo biloba extract EGb 761(R) in dementia with neuropsychiatric features: A randomised, placebo-controlled trial to confirm the efficacy and safety of a daily dose of 240 mg. Psychiatr. Res. 2012, 46, 716–723. [Google Scholar] [CrossRef]
- Bitencourt, J.; Zuffellato-Ribas, K.C.; Koehler, H.S. Ginkgo biloba L. cutting using three substrates. Rev. Bras. Plantas Med. 2010, 12, 135–140. [Google Scholar] [CrossRef]
- Kosenko, I.S.; Tsybrovska, N.V.; Balabak, O.A.; Hrabovyi, V.M.; Muzyka, H.I.; Shvets, T.A.; Oksantiuk, V.M. Introduction of Ginkgo biloba and its cultivars by vegetative propagation. Ukr. J. Ecol. 2021, 11, 65–76. [Google Scholar]
- Wang, L.; Lu, Z.; Li, W.; Xu, J.; Luo, K.; Lu, W.; Zhang, L.; Jin, B. Global comparative analysis of expressed genes in ovules and leaves of Ginkgo biloba L. Tree Genet. Genomes. 2016, 12, 29. [Google Scholar] [CrossRef]
- Zhang, G.F.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants. 2019, 5, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.R.; Matheny, N. The research foundation to tree pruning: A review of the literature. Arboric. Urban For. 2010, 36, 110. [Google Scholar] [CrossRef]
- Long, H.; James, S. Sensing and automation in pruning of apple trees: A Review. Agronomy 2018, 8, 211. [Google Scholar] [CrossRef]
- Forrester, D.I. Growth responses to thinning, pruning and fertiliser application in eucalyptus plantations: A review of their production ecology and interactions. Forest Ecol. Manag. 2013, 310, 336–347. [Google Scholar] [CrossRef]
- De la Paz, A.V.C.; Espinosa, B.G.G.; Brizuela, E.I.L.; Bravo, D.E.L.; Cueto, D.O.G. Fructification pruning in guava Crop (Psidium guajava L.) and its influence on yield. Rev. Cienc. Técnicas Agropecu. 2019, 28, 81–88. [Google Scholar]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—Part I: Concepts, regulation and consequences of phase change. New For. 2014, 45, 449–471. [Google Scholar] [CrossRef]
- Pereira, A.C.; Scaloppi Junior, E.J.; Costa, E.; Maitins, G.L.M.; Souza, N.C. Efeito da poda apical nos atributos morfofisiológicos do porta-enxerto clonal de seringueira GT 1. Ciência Florest. 2019, 29, 900–912. [Google Scholar] [CrossRef]
- Su, M.L.; Wu, S.; Ma, L.Y.; Duan, J.; Rong, G.C.; Su, S.C.; Ao, Y. Effects of three pruning methods on endogenous hormone and shoot growth of Xanthoceras sorbifolia Bunge buds. J. Northwest A F Univ. 2017, 45, 101–108. [Google Scholar]
- Duan, Y.J.; Meng, F.X.; Yang, S.Y.; Sun, M.Y.; Yang, Y.J.; Guo, S.P.; Liu, H.G.; Fang, H.D. Effects of different pruning methods on the growth of new shoots of Annona atemoya. Yunnan Acad. Agric. Sci. 2021, 7, 63–65. [Google Scholar]
- Csihon, Á.; Dremák, P.; Gonda, I. Partial and total rejuvenation pruning of sour cherry trees. Int. J. Hortic. Sci. 2015, 21, 11–15. [Google Scholar] [CrossRef]
- Zhu, L.G.; Sun, J.; Zhang, W.J.; Chen, Z.Z.; Wu, Z.D.; Jiang, F.Y. Water retention and biomass of litter from varied pruning practices on tea bushes. Fujian J. Agric. Sci. 2016, 31, 1210–1215. [Google Scholar]
- Zhang, C.; Liu, D.H.; Yang, T.F.; Zhang, Q.; Li, M.J.; Zhang, J.K. Different winter pruning amounts affect the growth and fruiting of ‘Fuji’ apples. Acta Agric. Boreali-Occident. Sin. 2016, 25, 1650–1655. [Google Scholar] [CrossRef]
- Atmaca, S.; Ulger, S. The effects of different planting densities and pruning methods on changes of endogenous hormone levels in shoot tips and flowering in ‘Gemlik’ Olive cultivar. Erwerbs-Obstbau 2021, 63, 201–207. [Google Scholar] [CrossRef]
- Chen, M.; Cao, M.; Lin, L.X. Research advances in regeneration of woody plants by sprouting. J. Plant Ecol. 2007, 26, 1114–1118. [Google Scholar]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: How buds, protection and resources drive persistence after fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef]
- Cao, M.; Gan, Q.; Xu, Y.; Lu, J.K.; Zhong, L.; Wang, M.X.; Liu, S.A.; Wang, L. Pruning improves seedling development and bioactive secondary metabolite accumulation in the leaves of Ginkgo biloba. Trees 2022, 36, 953–966. [Google Scholar] [CrossRef]
- Wang, Q.J.; Xu, S.Y.; Zhong, L.; Zhao, X.Y.; Wang, L. Effects of Zinc Oxide Nanoparticles on Growth, Development, and Flavonoid Synthesis in Ginkgo biloba. Int. J. Mol. Sci. 2023, 24, 15775. [Google Scholar] [CrossRef]
- Zhang, R.; Ding, J.; Liu, C.; Cai, C.; Zhou, B.; Zhang, T.; Guo, W. Molecular evolution and phylogenetic analysis of eight COL superfamily genes in group I related to photoperiodic regulation of fowering time in wild and domesticated cotton (Gossypium) species. PLoS ONE 2015, 10, e0118669. [Google Scholar]
- Xu, Y.; Lu, J.K.; Liu, S.A.; Wang, L. Research progress on the mechanism of resprouting in trees. J. Plant Physiol. 2023, 59, 803–816. [Google Scholar]
- Yang, B.; Hu, L.; Shen, H.; Sun, X.; Zhang, P. Effect of mounding and top-pruning on survival and growth of manchurian ash seedlings planted under the secondary forest of the species. Sci. Silvae Sin. 2018, 51, 104–113. [Google Scholar]
- Guo, C.J. Impact of pruning frequency and retention of branches on raspberry yield and benefits. North. Fruits 2016, 3, 15–17. [Google Scholar]
- Yu, Y.J.; Zhang, D.B.; Yuan, Z. The updated functional study of WOX protein family in regulating stem cell development. Chin. Bull. Bot. 2016, 51, 565–574. [Google Scholar]
- Lou, H.; Huang, Y.T.; Wang, W.Z.; Cai, Z.Y.; Cai, H.Y.; Liu, Z.Q.; Sun, L.; Xu, Q.J. Overexpression of the AtWUSCHEL gene promotes somatic embryogenesis and lateral branch formation in birch (Betula platyphylla Suk.). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 150, 371–383. [Google Scholar] [CrossRef]
- Gao, R.; Gruber, M.Y.; Amyot, L.; Hannoufa, A. SPL13 regulates shoot branching and flowering time in Medicago sativa. Plant. Mol. Biol. 2018, 96, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.L.; Yang, H.Y.; Yang, S.H.; Wang, J.H. Overexpression of short-root2 transcription factor enhances the outgrowth of mature axillary buds in poplar trees. J. Exp. Bot. 2022, 73, 2469–2486. [Google Scholar] [CrossRef]
- Tian, Y.T.; Wang, J.N.; Guo, H.P.; Qu, K.; Xu, D.; Hou, L.L.; Li, J.H. Transcriptome analysis of active axillary buds from narrow-crown and broad-crown poplars provides insight into the phytohormone regulatory network for branching angle. Plant Mol. Biol. Rep. 2021, 39, 595–606. [Google Scholar] [CrossRef]
- Kareem, A.; Durgaprasad, K.; Sugimoto, K.; Du, Y.J.; Pulianmackal, A.J.; Trivedi, Z.B.; Abhayadev, P.V.; Pinon, V.; Meyerowitz, E.M.; Scheres, B.P. PLETHORA genes control regeneration by a two-step mechanism. Curr. Biol. 2015, 25, 1017–1030. [Google Scholar] [CrossRef]
- Seale, M.; Bennett, T.; Leyser, O. BRC1 expression regulates bud activation potential but is not necessary or sufficient for bud growth inhibition in Arabidopsis. Development 2017, 144, 1661–1673. [Google Scholar] [PubMed]
- Zou, K.; Liu, X.; Zhang, D.; Yang, Q.; Fu, S.; Meng, D.; Chang, W.; Li, R.; Yin, H.; Liang, Y. Flavonoid biosynthesis is likely more susceptible to elevation and tree age than other branch pathways involved in phenylpropanoid biosynthesis in Ginkgo leaves. Front. Plant. Sci. 2019, 10, 983. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.A.; Meng, Z.L.; Zhang, H.Y.; Chu, Y.X.; Qiu, Y.Y.; Jin, B.; Wang, L. Identification and characterization of thirteen gene families involved in flavonoid biosynthesis in Ginkgo biloba. Ind. Crop. Prod. 2022, 188, 115576. [Google Scholar] [CrossRef]
- Zhao, B.B.; Wang, L.; Pang, S.Y.; Jia, Z.C.; Wang, L.; Li, W.X.; Jin, B. UV-B promotes flavonoid synthesis in Ginkgo biloba leaves. Ind. Crop. Prod. 2020, 151, 112483. [Google Scholar] [CrossRef]
- Jia, Z.; Zhao, B.; Liu, S.; Lu, Z.; Chang, B.; Jiang, H.; Cui, H.; He, Q.; Li, W.; Jin, B.; et al. Embryo transcriptome and miRNA analyses reveal the regulatory network of seed dormancy in Ginkgo biloba. Tree Physiol. 2020, 41, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gao, M.; Kang, S.C.; Yang, C.M.; Meng, H.; Yang, Y.; Zhao, X.S.; Gao, Z.H.; Xu, Y.H.; Jin, Y.; et al. Molecular mechanism underlying mechanical wounding-induced flavonoid accumulation in Dalbergia odorifera T. Chen, an endangered tree that produces Chinese rosewood. Genes 2020, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, Y.; Li, Y. Plant rejuvenation: From phenotypes to mechanisms. Plant Cell Rep. 2020, 39, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Hechmi, M.; Khaled, M.; Abed, S.; EL-Hassen, A.; Faiez, R.; M’hamed, A. Performance of olive cuttings (Olea europaea L.) of different cultivars growing in the agro-climatic conditions of Al-Jouf (Saudi Arabia). Am. J. Plant Physiol. 2013, 8, 41–49. [Google Scholar] [CrossRef][Green Version]
- Fan, H.Y.; Wang, Y.; Tang, F.; Lu, C. Determination of the mimic epitope of the M-like protein adhesin in swine Streptococcus equi subsp zooepidemicus. BMC Microbiol. 2008, 8, 170. [Google Scholar] [CrossRef]
- Xiao, Z.F.; Ji, N.; Zhang, X.Z.; Zhang, Y.Z.; Wang, Y.; Wu, T.; Xu, X.F.; Han, Z.H. The lose of juvenility elicits adventitious rooting recalcitrance in apple rootstocks. Plant Cell 2014, 119, 51–63. [Google Scholar] [CrossRef]
- Amri, E.; Lyaruu, H.V.M.; Nyomora, A.S.; Kanyeka, Z.L. Vegetative propagation of African Blackwood (Dalbergia melanoxylon Guill. & Perr.): Effects of age of donor plant, IBA treatment and cutting position on rooting ability of stem cuttings. New For. 2010, 39, 183–194. [Google Scholar] [CrossRef]
- Hong, H.H. The effect of cutting age on the rooting process, biochemical characteristics, and seedling growth of white poplar. Beijing For. Univ. 2018, 2, 274–281. [Google Scholar]
- Tong, P.X.; Ma, K.P.; Hu, Y.; Wang, L. Relationship between ginkgo cutting types and rooting rate. Mod. Hortic. 2019, 11, 10–11. [Google Scholar]
- Wang, R.M.; Zhu, L.G.; Chen, Y.; Yao, X.W.; Bai, J.W.; Chen, X.; Zhang, Q.Q.; Cao, F.L. Study on the rooting process and mechanism of hard branch cuttings of Ginkgo biloba. J. Cent. South Univ. For. Technol. 2020, 40, 28–37. [Google Scholar]
- He, Q.Q.; Zhang, Q.; Mu, X.P.; Zhang, J.C.; Du, J.J.; Wang, P.F. Effects of foliar spraying plant growth regulator on rooting and physiological changes of Cerasus humilis softwood cuttings. J. Plant Physiol. 2024, 60, 108–116. [Google Scholar] [CrossRef]
- Zhou, W.; Lozano-Torres, J.L.; Blilou, I.; Zhang, X.; Zhai, Q.; Smant, G.; Li, C.; Scheres, B. A jasmonate signaling network activates root stem cells and promotes regeneration. Cell 2019, 177, 942–956.e14. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, L.; Xu, S.; Xu, S.; Zhou, W.; Lu, Z.; Jin, B.; Wang, L. Consecutive Pruning Enhances Leaf Flavonoids, Leaf Yield, and Cutting Rooting in Ginkgo biloba. Forests 2024, 15, 761. https://doi.org/10.3390/f15050761
Zhong L, Xu S, Xu S, Zhou W, Lu Z, Jin B, Wang L. Consecutive Pruning Enhances Leaf Flavonoids, Leaf Yield, and Cutting Rooting in Ginkgo biloba. Forests. 2024; 15(5):761. https://doi.org/10.3390/f15050761
Chicago/Turabian StyleZhong, Lei, Shiyuan Xu, Shuwen Xu, Wanxiang Zhou, Zhaogeng Lu, Biao Jin, and Li Wang. 2024. "Consecutive Pruning Enhances Leaf Flavonoids, Leaf Yield, and Cutting Rooting in Ginkgo biloba" Forests 15, no. 5: 761. https://doi.org/10.3390/f15050761
APA StyleZhong, L., Xu, S., Xu, S., Zhou, W., Lu, Z., Jin, B., & Wang, L. (2024). Consecutive Pruning Enhances Leaf Flavonoids, Leaf Yield, and Cutting Rooting in Ginkgo biloba. Forests, 15(5), 761. https://doi.org/10.3390/f15050761