Abstract
Non-structural carbohydrates’ (NSCs) allocation pattern is closely related to environmental factors, plant metabolism, and xylem function. At the same time, we know little about whole-tree NSC allocation patterns in different seasons, especially the high-elevation species which suffer from environmental stress. We examined the concentration of NSCs in various parts of Faxon fir trees (needles, branches, trunks, and roots) at five elevations (2800 m, 3000 m, 3200 m, 3400 m, and 3600 m) over four months (July 2019, October 2019, January 2020, and April 2020). The goal was to understand how NSC allocation patterns vary by location and time in high-elevation species and what factors contribute to these variations. The results showed that the needles had the highest concentration, followed by roots, branches, and trunks. The NSC concentration was highest in January 2020 and lowest in July 2019. The total non-structural carbohydrates (TNSCs) and soluble sugar concentrations of roots and needles were substantially higher in the cold (non-growing season) than in the warm (growing season) season. At different elevations, the soluble sugar concentrations in the needles and trunks remained the highest and lowest, respectively. Branches and roots’ soluble sugar concentrations alternated and varied with the seasons at all elevations. Many factors, such as climate, morphological traits, and carbon content, affected the spatial and temporal patterns of non-structural carbohydrates, with temperature, plant moisture conditions, and carbon content being the main driving factors. Various factors’ interaction mainly influenced NSCs’ spatial and temporal patterns. Non-structural carbohydrates significantly improve the resistance of Faxon fir trees’ terminal organs in adverse environments.
1. Introduction
Photosynthesis produces and stores non-structural carbohydrates (NSCs) in plants, which include readily usable soluble sugar and stored starch. These carbohydrates can support future growth and metabolism by the plant [1,2]. Studies have shown that plant NSCs’ temporal and spatial dynamics occur with phenological changes. Several years ago, researchers discovered that the non-structural carbohydrates (NSCs) of evergreen broad-leaved leaves increased during the late summer and early fall seasons. However, broad-leaved deciduous leaves did not exhibit the same increase in NSCs during this time. Additionally, the concentrations of NSCs in different organs changed significantly with the seasons [3,4]. Researchers have recently found that several deciduous trees growing in temperate forests showed that starch on twigs was heavily consumed during spring germination, indicating C storage’s importance for spreading spring leaves [5]. However, the characteristics of species and their habitat may also cause their NSCs to exhibit different seasonal dynamics [6,7], and in some cases, the seasonal effects on the concentrations of NSCs were more significant than species effects [8].
In addition to the seasonal dynamic changes, there are also some studies on the internal spatial changes of individuals. From the perspective of the whole tree, the differences among different organs were relatively significant, especially in the branches [9]. Previous studies of Chinese fir under diverse environmental conditions found remarkable differences in NSCs concentration between above- and below-ground organs [10]. Organs of different species have different NSCs responses under the same environmental conditions because each plant organ has distinct functions [11]. Therefore, using a single organ to characterize the concentration of the whole tree is unreasonable because the concentrations of the individual organs vary during the season [1]. Current research on NSCs mainly focuses on two or more organs in a season or the seasonal dynamics of two organs [12,13], while little research has been conducted about the seasonal dynamics of all organs (leaf, branch, trunk, and root) of big trees in the wild environment, especially the subalpine treeline.
It was found to be critical for woody plants to store NSCs because it allowed sessile, long-living organisms to grow despite biotic and abiotic stresses [14,15]. The seasonal dynamics of plant organs’ NSCs are influenced by many factors such as carbon starvation, hydraulic dysfunction, pests and diseases, and the combined effects of all these factors [13,16]. Hydraulic dysfunction is a problem that happens when plants do not receive enough carbon, and it can affect how plants store carbohydrates. To avoid this problem, plants need to consume non-structural carbohydrates, especially soluble sugars, which help them maintain their hydraulic system and keep their structure intact [17].
Soluble sugar is released into the embolism xylem and creates a concentration gradient between the embolism and the normal xylem [18]. These factors often appear together and can deplete the non-structural carbohydrates (NSCs) stored by plants for future use [1,15,19]. If not managed, NSCs depletion can cause trees to die. It can even lead to ecosystem degradation and affect plant functional processes directly or indirectly related to NSCs. These functions include growth activity, osmoregulation, osmoprotection, pest and disease resistance, water and nutrient transport, storage, and drought and cold resistance [9,20,21,22,23]. The higher ratio of soluble sugars and starches in plant organs explains the ability of some species to grow in low-temperature stressed environments at high elevations [24].
Global climate change results in an increasing frequency and intensity of extreme weather, which affects tree growth and forest ecosystem function [25,26,27,28] and also affects trees’ morphological traits (such as leaf size and leaf mass per unit area) [29,30]. Trees often store NSCs in the stem and roots and use them to support plant growth or resist environmental stress in the future [31]. The growth of trees and NSCs reserves are strongly correlated, and NSCs have a crucial role in trees’ resilience to extreme weather events in the Mediterranean area [32].
Researchers used NSCs to improve predictions of a forest’s gross primary productivity and promote further understanding of the complex relationship between the environment and photosynthesis [33], showing that trees’ NSCs concentration was essential for the stability of the forest ecosystem functions. Especially in an extreme climate, tree stems’ concentration of non-structural carbohydrates was a good indicator of tree mortality [34]. Global climate change induces carbon starvation, one of the causes of tree mortality, and is expected to exacerbate forest vulnerability [35].
Global climate change has increased the frequency of extreme weather events, which has affected the structure and composition of forests and the quality and structure of habitats [36]. According to predictions, chronic temperature increases are responsible for massive conifer mortality [37]. In stressful situations, trees’ carbon reserves play an important role in resilience, and the different components of NSCs play different roles [32]. A recent study confirmed that Faxon fir radial growth is constrained by thermal and hydraulic factors, while hydrothermal conditions are highly affected by global climate change [38]. Conversely, southwest China is one of the regions most vulnerable to climate change, and the conifer forest vulnerability of this area showed an increasing trend with global climate change [39,40]. Hence, there is a rising need to be expanded in both breadth and depth to comprehensively assess typical conifer species NSCs allocation, storage, and seasonal dynamics in southwest China better to understand the function of NSCs in subalpine forest ecosystems.
In this paper, we study the NSCs of Faxon fir for four months (July 2019, October 2019, January 2020, and April 2020). We set five elevation gradients at intervals of 200 m (2800–3600 m) in Wolong Nature Reserve in Sichuan province. We aim to reveal the effects of seasonal variations on NSCs storage and consumption at different elevations and the NSCs spatial–temporal distribution patterns. We also want to know which carbon strategies Faxon fir adopts to cope with climate change and the main driving factors for these distribution patterns. Specifically, we will test the following hypotheses:
- The whole-tree allocation pattern will be as follows: the terminal organs contribute more due to Faxon fir trees needing to store more energy to cope with environmental stress during the cold season, and NSCs are significantly lower in the warmer season than the cold season.
- Faxon fir organs (especially the terminal organs) accumulate a higher concentration of NSCs in winter to enhance tissue resistance to high-altitude low temperature so as to successfully overwinter.
- Due to its unique geographical environment, the distribution pattern of NSCs in Faxon fir at high altitudes may be influenced by various factors.
2. Materials and Methods
Study Area
We carried out this experiment at Wolong Nature Reserve (WoNR, latitude 30°53′ N, longitude 102°58′ E) in Sichuan province, southwest China. WoNR is located in the western Sichuan Plateau, with a subtropical semi-humid climate characterized by dry, cold winters and wet, cool summers [41,42]. The mean annual temperature is 4.06 °C, and the mean annual precipitation is about 1026 mm. In WoNR, the dominant tree species are Faxon fir, accompanied by Rhododendron faberi Hemsl, Rhododendron asterochnoum Diels, Picea purpurea Mast, Betula albosinensis Burkill, and Rosa moyesii [41,43]. The Faxon fir grows in different altitude ranges. It co-exists with understory bamboo (Fargesia spp.) at 2800–3100 m above sea level and with Rhododendron spp. at 3200–3600 m above sea level [41]. According to the Chinese Soil Taxonomy classification, the soil type is dark brown [44]. Faxon fir (Abies fargesii var. faxoniana) survives in the cool and wet subalpine region of Sichuan province in southwest China, and its distribution area is also the main habitat of Chinese giant pandas (Ailuropoda melanoleuca David) [43]. It is the dominant tree species of the subalpine coniferous forest [45]. It is naturally distributed from 2700 to 3900 m a.s.l in the subalpine area of Sichuan province [41]. In the genus Abies, Faxon fir is an ancient species that has survived through glacial and interglacial periods [46].
3. Field Data Collection
3.1. Climate Data
Using data from the observations of over 2400 stations in China, the average seasonal temperature for the study sites was calculated from 1997 to 2016 using an interpolation method [41,47]. We used topographic correction to calculate the air temperature at each elevation, applying a lapse rate of 0.65 °C per 100 m [48]. Soil temperature was measured using soil button thermometers (iButton, DS1922L) set in WoNR from 2018.5 to 2019.4 (Figure 1). The soil depth was 10 cm. Two soil button thermometers were set at each altitude and recorded one reading each hour, and the average value was used to express soil temperature [41]. The growing season shortens with increasing altitude. The growing season lasts five months (May–September) at 2800 m above sea level (a.s.l). However, at 3600 m a.s.l, only two months, July and August, are included in the growing season, as shown in Figure 1.
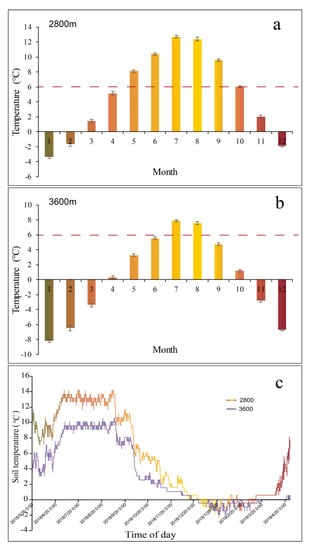
Figure 1.
Mean monthly temperature for several years at Wolong Nature Reserve at (a) 2800 m a.s.l and (b) 3600 m a.s.l; (c) soil temperature dynamic at 2800 and 3600 m a.s.l from May 2018 to April 2019. The red dotted line means the 6 °C datum line, presenting the data as the mean value ± SE.
3.2. Growing Season and Non-Growing Season Determination
During the growing season, tree line areas naturally have a mean temperature of around 6 °C. However, as temperatures decrease below this threshold, tree species’ root growth is limited [49]. The temperature controls the germination of species, and the threshold for the air temperature to trigger germination is approximately 6 °C [50]. Thus, we considered the growing season as the period when the average monthly temperature was above 6 °C. The average monthly air temperature at the 2800 m a.s.l. research sites was over 6 °C from May to September and below 6 °C between October and April. Meanwhile, at the highest sites at 3600 m a.s.l, the mean air temperatures were higher than 6 °C only in July and August. The months selected for this study were April, July, October, and January, and we used July to indicate the growing season and April, October, and January to indicate the non-growing season.
3.3. Sample Collection and Pre-Dawn Leaf Water Potential Measurement
We selected healthy adult individuals with similar diameters from each sample site and selected branch samples with mature spreading leaves on the sunny side. In this experiment, we measured pre-dawn leaf water potential (Ψ) with a pressure chamber (SKPM1400-50, SKYE, UK). We took measurements on shoots from the four selected months and elevations and collected samples for Ψ before sunrise (06:00–07:00 h). After cutting them from the trees, we wrapped 1.5 m long branch samples in small plastic bags containing moist paper towels. All the samples were sealed immediately in big black plastic bags containing moist paper towels and sent to the laboratory for hydraulic traits measurement. We selected three current-year shoots from the branches of the samples and measured all samples within an hour of the samples leaving the tree.
3.4. Laboratory Measurement
3.4.1. Morphological Traits Measurement and Calculation
Morphological traits include LA/SA (the ratio of leaf area and sapwood area), LMA (leaf mass per area), and tracheid lumen diameter (TLD); a detailed description is provided in Pan’s study [51].
3.4.2. Non-Structural Carbohydrate Measurements
We simultaneously measured the non-structural carbohydrates of Faxon fir trees’ roots, trunks, branches, and needles across five different altitudes in January, April, July, and October. We also measured the hydraulic properties of the xylem. To collect data, we took fine roots (with a diameter of less than 5 mm) from the topsoil at a depth of approximately 20 cm. At 1.35 m, we drilled two 5-mm diameter increment borer holes on opposing sides of the trunk, and at each of the six heights, we obtained two tree cores from each of the trees. We used a microwave oven at 600 W for 40 s to kill the samples in time for non-structural carbohydrate analysis. Then, we dried them to a constant mass at 70 °C for more than 48 h.
We first weighed 0.1 g of the sample and placed it in a 10-mL centrifuge tube for chemical analysis. Next, we added 5 mL of an 80% ethanol solution to the tube. The mixture was then incubated in a boiling water bath at 80 °C for 30 min before being centrifuged at 5000 r for 10 min. We collected the supernatant and extracted the pellet twice with an 80% ethanol solution. Finally, we combined the supernatant and stored it at −20 °C before determining the soluble sugar content using the Anthrone method [17,52].
To prepare the extract, we evaporated the alcohol in the precipitate to dryness. Then, we added 2 mL of distilled water and shook it well. Finally, we gelatinized the mixture in a boiling water bath for 15 min. Once the mixture was cooled to room temperature, we added 2 mL of 9.2 mol/L perchloric acid (HClO4) and stirred it for 15 min. After hydrolyzing the starch, we added 4 mL of distilled water to the mixture and centrifuged it at 4000 r for 10 min. Then, we transferred the supernatant to a 50 mL volumetric flask. Next, we added 2 mL of 4.6 mol/L perchloric acid (HClO4) to the precipitate and stirred it. After 15 min, we added 5 mL of distilled water and mixed it well. Then, we centrifuged the mixture for 10 min and transferred the supernatant to a 50 mL volumetric flask. We washed the precipitate twice with distilled water and combined the supernatants before making the volume constant. The resulting extract was used to determine the starch content [17].
3.5. Chemical Analyses
To analyze the carbon (C) concentration in our samples, we dried the needles, branches, trunks, and roots in an oven and used an elemental analyzer (Vario EL III, CHNOS Elemental Analyzer; Elementar Analysensysteme GmbH, Frankfurt, Germany).
3.6. Data Manipulation and Statistical Analysis
This study analyzed the seasonal variations of non-structural carbohydrates among different sites and the seasonal variations in each organ at each site. We used SPSS 18.0 to analyze the changes in air and soil temperatures. To check if our data followed a normal distribution, we employed the Lilliefors test and the Shapiro–Wilk test. Additionally, we used the F test and Levene’s test to assess whether the variances of our data were homogeneous.
We used a function called “Varpart” from the “Vegan” package to partition and analyze the variation in the concentrations of soluble sugar and starch across different organs (needles, branches, trunks, and roots) into components under four categories of predictors (i.e., climate factors, the carbon content of organs, physiological trait factors, and morphological trait factors) in R Studio with version 3.5.2.
We used a redundancy analysis (RDA) to determine the relationship between the climate variables, carbon content of organs, and physiological and morphological factors with non-structural carbohydrates (soluble sugar and starch) across all sites. We subjected the results of the RDA to a Monte Carlo permutation test to determine their significance level (999 permutations). We transformed the explain and response factors using either Hellinger or standardized transformations in the RDA.
We used linear regression models to examine the relationship between soluble sugar and starch concentrations, MAT, Ψ, LMA, and root C content across the study site. We used a one-way analysis of variance (ANOVA) to examine the variations in soluble sugar and starch concentration among needles, branches, trunks, and roots. We conducted multiple comparisons using the LSD test.
4. Results
4.1. Spatio-Temporal Non-Structural Carbohydrate Pattern of Faxon Fir
The soluble sugar concentration of the whole trees reached the highest value in January 2019 across all the sampling sites. According to Figure 2a, soluble sugar followed a similar pattern to TNSCs, while starch remained stable with seasonal variations. Figure 2b shows that the order of TNSCs concentrations in organs from highest to lowest was needle > root > branch > trunk during the four selected months. In Figure 2c, the concentration of starch in organs exhibits a “V”-shape variation with the seasons, with root and needle concentrations being similar, as were branch and trunk concentrations. Finally, Figure 2d demonstrates that the order of soluble sugar concentrations in organs from highest to lowest was needle > root > branch > trunk during the four selected months. The soluble sugar concentrations in the needles and roots showed similar seasonal trends. In contrast, the maximum concentration in the branches and trunks occurred between October 2019 and January 2020, with the highest in January 2020.
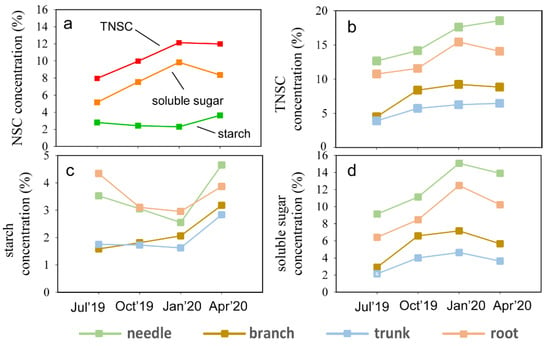
Figure 2.
Spatio-temporal distribution pattern of non-structural carbohydrates in Faxon fir. (a) Total non-structural carbohydrates (TNSCs), soluble sugar, and starch variations with seasons; (b) each organ’s total non-structural carbohydrates (TNSCs) variations with season; (c) each organ’s starch concentration variations with season; (d) each organ’s soluble sugar concentration variations with season.
The seasonal pattern of soluble sugar concentration in Faxon fir organs was similar at different elevations (Figure 3 and Figure 4). The concentration of soluble sugar in needles, branches, and roots increased with the elevation each season and stayed higher from October to April, but the concentrations in the trunk were stable (Figure 3a,c,e,g,i and Figure 4a,c,e,g,i). The spatial pattern of soluble sugar concentration was also consistent at different elevations, from high to low as follows: needles > roots > branches > trunks. The starch concentration of Faxon fir showed different spatial and temporal patterns at different elevations (Figure 3b,d,f,h,j and Figure 4b,d,f,h,j). The starch concentration in needles was higher than that in other organs at 2800 m altitude, increasing with elevation. The starch concentration in roots gradually increased as compared to needles, and the starch concentration in needles and roots showed a “V” shape from July to April at all elevations. The value was lowest in January.
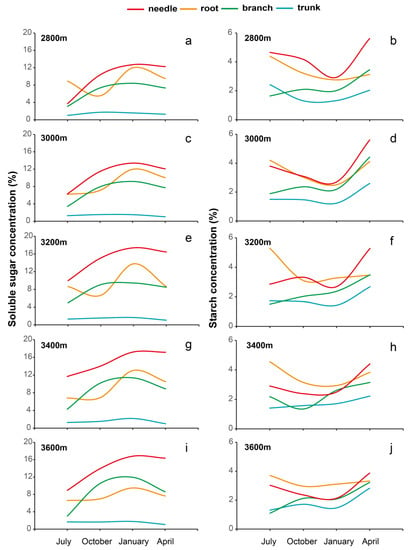
Figure 3.
Changes in soluble sugar concentration (a,c,e,g,i) and starch concentration (b,d,f,h,j) of Faxon fir tree organs (roots, trunks, branches, and needles) with seasonal variations across different altitudes in WoNR. Lines in different colors represent different non-structural carbohydrate types and organs.
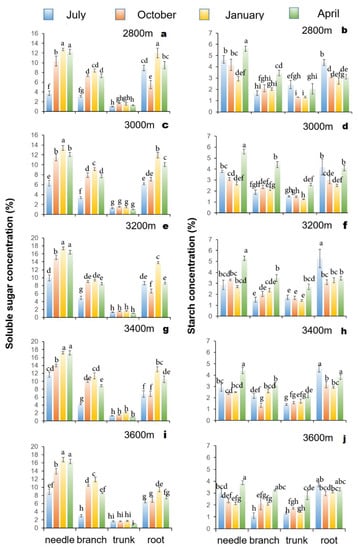
Figure 4.
Different altitude Faxon fir soluble sugar (a,c,e,g,i) and starch concentrations (b,d,f,h,j) of organs (needles, branches, trunks, and roots) in different seasons (July, October, January, and April). Data are presented as means ± SE. Different letters indicate significant differences among organs and seasons based on an LSD test (n = 6, p < 0.05).
The starch concentration in branches was lowest in July and highest in April at 2800 m, 3000 m, 3200 m, and 3600 m a.s.l, while it was lowest in October and highest in April at 3400 m a.s.l. (Figure 3b,d,f,h,j and Figure 4b,d,f,h,j). The starch concentration in trunks showed a “V” shape at 2800 m and increased with elevation, being significantly higher in April than in other seasons (Figure 3b,d,f,h,j and Figure 4b,d,f,h,j). Soluble sugar and starch showed substantial organ variations across altitudes in the four selected months, and the combined effect among seasons, organs, and altitude showed an influence (Table 1).

Table 1.
The effects of altitude, season, organ, and the interaction between altitude, season, and organ, as analyzed using a three-way ANOVA.
4.2. Various Factors Influence the Temporal and Spatial Distribution Pattern of Non-Structural Carbohydrates in Faxon Fir Organs
Values in the figure represent the detailed variations in each category of elements and various interactions (Figure 5). Climate factors include soil temperature (Soil.T), mean annual temperature (MAT), and mean monthly temperature (MMT) (Figure 1); Ψ represents the pre-dawn leaf water potential, which showed a significant difference among study sites at different measurement times and decreased with the increasing elevation in general (Table 2). Carbon content includes the root, needle, trunk, and branch carbon content (CNeedle, CBranch, CTrunk, and CRoot). Morphological traits include leaf area per unit sapwood area (LA/SA), leaf mass per area (LMA), and tracheid lumen diameter (TLD); LMA showed a trend of gradual increase with increasing elevation, LA/SA showed a significant difference among study sites, and TLD exhibited a decreasing trend with increasing elevation (Table 3).
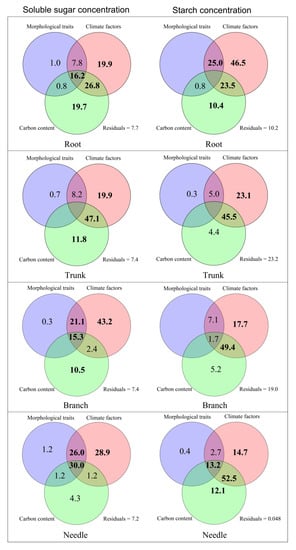
Figure 5.
Schematic diagram of variation partitioning to determine the effects of climate factors, carbon content, and morphological traits on the amount of sugar and starch in different parts of a Faxon fir tree.
Table 2.
Pre-dawn leaf water potential of Faxon fir in four experimental periods at different altitudes (letters indicate significance with different traits at different altitudes).
Table 3.
Morphological traits (LMA, LA/SA, and TLD) of Faxon fir at different altitudes (letters indicate significance with different traits at different altitudes).
We found that climate factors, carbon content, and morphological traits primarily influenced needles’ soluble sugar concentration, showing 30% variance through the variance partitioning analysis. Climate factors alone accounted for 28.9% of the variation, while the combined effect of climate factors and morphological traits accounted for 26% (Figure 5). The needle starch concentration was affected by the combined effect of climate factors and carbon content with 52.5% variation; was jointly influenced by climate factors, carbon content, and morphological traits, showing 13.2% variation; and was further affected by the two factors with 14.7% and 12.1% variation (Figure 5).
Climate factors accounting for 43.2% variation primarily described the branch soluble sugar concentration variance. The combined effect of morphological traits and climate factors accounted for 21.1% of the variation. In contrast, the combined effect of morphological traits, climate factors, and carbon content accounted for 15.3% of the variation. The carbon content effect was 10.5% of the variation. The combined effect of climate factors and carbon content on starch concentration showed 49.4% variation, and climate factors further independently explained 17.7% of the variation (Figure 5).
We interpreted the soluble sugar concentration variation of the trunk by the combined effect of climate factors and carbon content, which accounted for 47.1%. Climate factors and carbon content individually accounted for 19.9% and 11.8%, respectively (Figure 5). The trunk starch concentration also showed 45.5% variance by the combined effect of climate factors and carbon content, with climate factors further independently explaining 23.1% of the variation (Figure 5).
We explained the variance of the root soluble sugar concentration by the combined effect of climate factors and carbon content, with a 26.8% variance. Climate factors and carbon content individually accounted for 19.9% and 19.7%, respectively, and the combined effect of morphological traits, climate factors, and carbon content showed a 16.2% variance (Figure 5). Variation in root starch concentration due to climate factors was 46.5%, that from the combined effect of morphological traits and climate factors was 25.0%, that due to climate factors and carbon content was 23.5%, and that further independently explained by carbon content was 10.4% (Figure 5).
The results of the RDA explain the variations of Faxon fir organs’ soluble sugar and starch concentrations across the sampling sites in four months, with 47% at axis 1 and 41% at axis 2 (Figure 6). Based on the Monte Carlo test in the RDA, MMT (F = 759.5, p < 0.001), Soil.T (F = 674.9, p < 0.001), Ψ (F = 264.5, p < 0.001), and CBranch (F = 114.2, p < 0.001) were the dominant driving factors of Faxon fir organs’ soluble sugar and starch concentration variations. The RDA ordination biplot (Figure 6) shows that carbon content (CNeedle, CBranch, CTrunk, and CRoot) was an important factor that caused changes in the concentrations of soluble sugar and starch in the Faxon fir organs. These changes occurred along RDA axis 1 and affected the starch concentration in needles, trunks, and branches and the sugar concentration in roots and trunks.
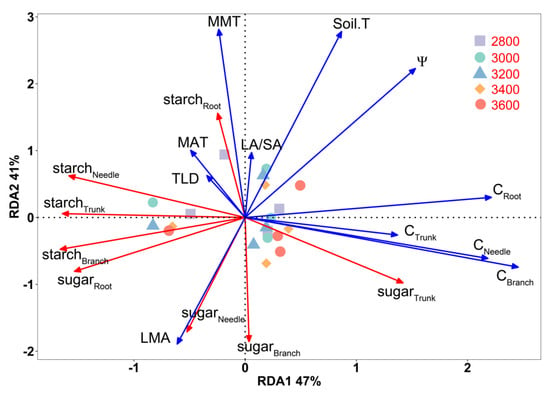
Figure 6.
Redundancy analysis (RDA) of environmental factors and different organs’ non-structural carbohydrate concentrations across sampling sites in different measurement months.
Figure 6 indicates that environmental factors (MAT, MMT, and Soil.T) and morphological traits (LA/SA and LMA) significantly influenced soluble sugar and starch concentration variations in the Faxon fir organs along RDA axis 2. This affected starch levels in roots and sugar levels in branches and needles. The variables Ψ, Soil.T, CNeedle, CBranch, CTrunk, and CRoot all displayed negative associations with starchNeedle, starchTrunk, starchBranch, sugarRoot, and sugarTrunk and were positively associated with sugarTrunk (Figure 6, Figures S2 and S3). The variables Ψ, TLD, MAT, MMT, Soil.T, and LA/SA showed positive associations with starchRoot and were negatively associated with sugarBranch and sugarNeedle. LMA was positively associated with sugarBranch and sugarNeedle but negatively associated with starchRoot (Figure 6 and Figures S1–S3). TLD and MAT were positively associated with starchNeedle, starchTrunk, starchBranch, and sugarRoot but negatively associated with sugarTrunk (Figure 6 and Figure S1).
Variables displayed in Figure 6 include the following: Ψ, pre-dawn leaf water potential; Soil.T, soil temperature; MAT, mean annual temperature; MMT, mean monthly temperature; starchNeedle, needle starch concentration; starchBranch, branch starch concentration; starchTrunk, trunk starch concentration; starchRoot, root starch concentration; sugarNeedle, needle soluble sugar concentration; sugarBranch, branch soluble sugar concentration; sugarTrunk, trunk soluble sugar concentration; sugarRoot, root soluble sugar concentration; CRoot, carbon content of root; CTrunk, carbon content of trunk; CNeedle, carbon content of needle; CBranch, carbon content of branch; LA/SA, leaf area per unit sapwood area; LMA, leaf mass per area; TLD, tracheid lumen diameter. The points with different colors and shapes represent different elevations.
5. Discussion
5.1. Terminal Organs Needles and Roots’ NSC Concentration Is Higher than That of Branches and Trunk
Our results show that the order of Faxon fir organs from high to low NSCs concentration is needle > root > branch > trunk (Figure 2, Figure 3 and Figure 4). This difference probably resulted from differences in the physiology of each organ [10] and also due to the dry density and activity in the different tissues. Needles are photosynthesis organs that produce sugars, which constitute a significant NSCs component (Figure 2, Figure 3 and Figure 4). Previous research has confirmed that photosynthesis rates positively correlate with leaf mass per unit area (LMA) [53], which is consistent with this paper’s result of LMA showing a significant relationship with needle sugar concentration (Figure 6). Soluble sugar’s contribution to LMA may differ in different plant species. For example, in tomato, the contribution of soluble sugar to LMA was not significant in a day’s growth, but the contribution of starch was very significant [54]. However, in P-limited soil environments in tropical forests, it was found that higher concentrations of NSCs may lead to a lower LMA; this may be due to the close correlation between leaf area and NSCs [55]. In this study, the soluble sugar concentrations in leaves and branches were closely correlated with LMA (Figure 6) due to the plant’s self-protection mechanism; in an advanced environment, plants want to reserve more NSCs for enhanced resistance [56].
Kozlowski’s research showed that high NSCs concentrations in conifer leaves can be essential in storage and cold tolerance [57]. On the other hand, a high NSCs concentration in needles reflects their high metabolic rates, and needles need high NSCs concentrations to maintain the turgor pressure of many living cells [58,59]. Roots had the second highest NSCs concentrations across the sample sites in the four months (Figure 2, Figure 3 and Figure 4). In contrast to other organs, roots are believed to specialize as storage organs [57], so roots would contain the most starch [9]; this agrees with our finding that roots stored the most starch with seasonal variations, while storing less than the needles in April 2020 (Figure 2, Figure 3 and Figure 4). The osmotic and metabolic demands are intermediate in roots, which results in a medium NSCs concentration [58]. In contrast, roots have a relatively high NSCs concentration and form a concentration gradient between root and soil water, which plays a crucial part in roots absorbing water from the soil [60].
Below-ground carbon allocation is an essential process of the terrestrial ecosystem carbon cycle and couples the activity of sink organs [60]. Furthermore, in unfavorable conditions, plants invest C into storage pools (roots) to prepare for environmental condition improvement. It enables plants to regrow and increase their resilience under adverse environmental conditions [60]. The branch NSCs concentration was slightly lower than that of the roots; the Faxon fir needle has a short petiole, and the resistance of energy transport between needle and petiole is reduced. Leaf size affects the leaf photosynthesis capacity and its nutrient acquisition and indirectly affects nutrient distribution in other organs. Non-structural carbohydrates depletion in the branches and trunk, in addition to affecting xylem growth and sap chemistry, could indirectly increase xylem hydraulic vulnerability [61].
The stems had a lower NSCs concentration, which may affect their ability to refill embolized xylem conduits [61,62]. For low-temperature environment species, NSCs, especially sugars, were positively related to cold tolerance in perennial organs and used to adjust intracellular osmotic concentration [22,63]. This paper’s results on organ NSCs distribution patterns were consistent with Martínez-Vilalta’s study showing that leaf NSCs concentrations change according to biome and plant functional type. In contrast, root NSCs concentrations were intermediate [58]. The NSCs storage and distribution patterns of various plant organs resulted from a long-term adaptation of the species to the environment and climate change. It is a true reflection of plant survival strategies [64].
5.2. Plant Organs Have Higher Concentrations of NSCs in the Low-Temperature Season than in the Warmer Season
Faxon fir is mainly distributed at 2700 m to 3900 m a.s.l. The subalpine region has a significant temperature difference between the growing season (May–September) and the non-growing season (October–April). The length of the growing season decreased with the increasing elevation (Figure 1). Lower soluble sugar and TNSCs concentrations of needles and branches were found in July 2019 (growing season), while being higher in October 2019, January 2020, and April 2020 (non-growing season); furthermore, a higher starch concentration was found in July 2019 and April 2020. The NSCs seasonal changes in our study were similar to those in Zhang’s previous study [65], which found a significant reduction in NSCs concentration at the end of the growing season in branches and trunks. Phenological changes were one of the trigger factors of NSCs allocations during the year [66].
During the growing season, plants need newly produced sugars to support crucial processes such as bud break and sprouting, as stated by previous studies [67,68]. As a result, they have limited reserves for the rest of the growing season. However, evergreen species begin photosynthesizing early in the spring, allowing them to accumulate some sugars. The high leaf grafting and radial growth consumption impede NSCs accumulation in the growing season, such as in L. decidua and P. cembra [20,66]. Our findings aligned with Hoch’s study, which reported a high proportion of free sugars and TNSCs in Faxon fir during the non-growing season. Hoch et al. suggested that the elevated concentration of NSCs throughout the non-growing season prepared the trees for winter freezing tolerance [20].
Additionally, our results demonstrated that the NSCs concentration of Faxon fir organs had a significant negative correlation with MAT and soil temperature, as shown in Figures S1 and S2. Research has confirmed that when exposed to low temperatures, starch converts into soluble sugars, which then act as osmoprotectants to improve the freezing tolerance of overwinter organs [69]. Figure 2, Figure 3 and Figure 4 and Table 1 indicate that the NSCs concentration in the trunks varied with the seasons at partial elevation, which aligns with Martinez-Vilalta’s findings. They discovered that seasonal changes in stems of all functional types are statistically significant due to the large stem biomass of trees. Even though there may be minor variations in concentration, stems play a crucial role in plants’ overall seasonal carbohydrate dynamics [58].
Figure 2, Figure 3 and Figure 4 and Table 1 indicate that the roots’ soluble sugar and NSCs concentrations fluctuated with seasonal variations across all the sample sites. Roots are often considered a long-term storage pool, only utilized during catastrophic events such as loss of aboveground biomass [9]. Our results showed that the starch concentration in needles and roots exhibited a “V” shape, which means that the stored starch in needles and roots decreased in the non-growing season but decreased with soluble sugar increase. This may be due to the conversion of starch into soluble sugar to improve organs’ cold resistance [13,17]. For high-elevation trees, temperature was an important driving factor of carbon storage states, explaining slow growth and treeline formation [70] (Figures S1 and S2).
5.3. NSCs Improve the Resistance of Faxon Fir to Environmental Stress
Bush et al. confirmed that environmental factors affect plants’ composition and the distribution of non-structural carbohydrates [71]. Furthermore, Swanson et al. found that environmental disturbances are the main drivers shaping forest structure and succession [72]. Turco et al. predicted that the severity and frequency of abiotic disturbances would increase due to climate change [73]. Piper and Paula suggested that this may lead to an increased reliance on the allocation and storage of carbon by plants [74]. Similarly, Figure 5 and Figure 6 present related information. Besides moisture, temperature, and light conditions, carbon content is essential in NSCs storage (Figure 5, Figure 6 and Figure S4) when carbon gains are higher than demand, leading to NSCs storage [75]. Our results showed that root carbon content negatively correlated with soluble sugar and starch concentrations. This confirmed that a dynamic trade-off of photosynthesis products exists between NSCs and C content [75]. As a result of organs not being resupplied with C quickly enough, plants could starve and die [76].
Scientists predict that the temperature will rise to 4 °C by 2100, and these alterations in precipitation patterns will make global climate change seem overwhelming [77]. The impact of climate change is often slow and far-reaching, including increases in drought stress, shifts in species’ geographical locations and modifications to their physiology and seasonal behavior [77], and morphological adaptation to the environment, directly or indirectly altering NSCs distribution patterns (Figure 5 and Figure 6). High-elevation species affected by global climate change will lose their habitats at a lower elevation of distribution and need to migrate upward to recouple populations with the climates to which they are adapted [78]. To get used to the new environmental conditions, species at high elevations need to adjust or change their survival strategies.
6. Conclusions
Our results have important implications for understanding the high-elevation Faxon fir NSCs allocation in different organs (spatial pattern) and according to seasonal variation (temporal dynamics). Needles, branches, and roots had a high concentration of NSCs in the non-growing seasons across all the sample sites. In contrast, the starch concentration in each organ fluctuated with the season and accumulated in overwintering organs to improve their cold resistance and prepare for growth in the non-growing season. Plants stored more starch during early spring, whereas soluble sugar consumption was more significant.
The concentration of NSCs in roots followed a wave-like pattern over time, and it was affected by various factors besides moisture and temperature. Many factors influenced when and where the Faxon fir trees stored their NSCs. Biotic–abiotic factors such as temperature, water status, and root carbon content influence the distribution of non-structural carbohydrates (NSCs) in different organs of Faxon fir. These factors also prepare the plant to use NSCs differently in different seasons. For instance, during the growing season, the plant depletes a lot of soluble sugar but stores some starch. The combined effect of these biotic–abiotic factors explains the allocation strategy of NSCs. This allocation strategy resulted from Faxon fir’s adaptation to environmental changes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14071438/s1, Figure S1: The linear relationships of soluble sugar and starch concentration of Faxon fir organs with mean monthly temperature (MMT). Solid lines indicate a significant relationship. Each point is a mean value (n = 6); Figure S2: Correlations between soluble sugar and starch concentration of Faxon fir organs with Soil temperature. Each point is a mean value. Figure S3: Correlations between soluble sugar and starch concentration of Faxon fir organs with Pre-dawn leaf water potential. Each point is a mean value; Figure S4: The linear relationship of soluble sugar and starch concentration of Faxon fir with needle carbon content. CBranch: needle carbon content. Each point is a mean value; Figure S5: One-way analysis of variance of Faxon Fir organ’s (Needle, a b; Branch, c d; Trunk, e f; Root, g h) soluble sugar and starch concentration among five study sites. “⁎” means significant difference; “ns” means no difference.
Author Contributions
Conceptualization, S.P.; Methodology, S.P., X.Y. and X.D.; Formal analysis, S.P. and X.L.; Investigation, S.P.; Data curation, X.Y. and X.D.; Writing—original draft, S.P.; Writing—review & editing, S.A.A. and Z.L.; Supervision, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| NSC | Non-structural carbohydrates |
| Ψ | Pre-dawn leaf water potential |
| Soil.T | Soil temperature |
| MAT | Mean annual temperature |
| MMT | Mean monthly temperature |
| starchNeedle | Needle starch concentration |
| starchBranch | Branch starch concentration |
| starchTrunk | Trunk starch concentration |
| starchRoot | Root starch concentration |
| sugarNeedle | Needle soluble sugar concentration |
| sugarBranch | Branch soluble sugar concentration |
| sugarTrunk | Trunk soluble sugar concentration |
| sugarRoot | Root soluble sugar concentration |
| CRoot | Carbon content of root |
| CTrunk | Carbon content of trunk |
| CNeedle | Carbon content of needle |
| CBranch | Carbon content of branch |
| LA/SA | Leaf area per unit sapwood area |
| LMA | Leaf mass per area |
| TLD | Tracheid lumen diameter |
References
- Richardson, A.D.; Carbone, M.S.; Keenan, T.F.; Czimczik, C.I.; Hollinger, D.Y.; Murakami, P.; Schaberg, P.G.; Xu, X. Seasonal dynamics and age of stemwood non-structural carbohydrates in temperate forest trees. New Phytol. 2013, 197, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Quentin, A.G.; Pinkard, E.A.; Ryan, M.G.; Tissue, D.T.; Baggett, L.S.; Adams, H.D.; Maillard, P.; Marchand, J.; Landhäusser, S.M.; Lacointe, A.; et al. Non-structural carbohydrates in woody plants compared among laboratories. Tree Physiol. 2015, 35, 1146–1165. [Google Scholar] [CrossRef] [PubMed]
- Parker, J. Seasonal variations in sugars of conifers with some observations on cold resistance. For. Sci. 1959, 5, 56–63. [Google Scholar]
- Parker, J. Seasonal changes in cold resistance and free sugars of some hardwood tree barks. For. Sci. 1962, 8, 255–262. [Google Scholar]
- Klein, T.; Vitasse, Y.; Hoch, G. Coordination between growth, phenology and carbon storage in three coexisting deciduous tree species in a temperate forest. Tree Physiol. 2016, 36, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Oleksyn, J.; Zytkowiak, R.; Karolewski, P.; Reich, P.B.; Tjoelker, M.G. Genetic and environmental control of seasonal carbohydrate dynamics in trees of diverse Pinus sylvestris populations. Tree Physiol. 2000, 20, 837–847. [Google Scholar] [CrossRef]
- Smith, M.G.; Miller, R.; Arndt, S.; Kasel, S.; Bennett, L.T. Whole-tree distribution and temporal variation of non-structural carbohydrates in broadleaf evergreen trees. Tree Physiol. 2018, 38, 570–581. [Google Scholar] [CrossRef]
- Liu, W.; Su, J.; Li, S.; Lang, X.; Huang, X. Non-structural carbohydrates regulated by season and species in the subtropical monsoon broad-leaved evergreen forest of Yunnan Province, China. Sci. Rep. 2018, 8, 1083. [Google Scholar] [CrossRef]
- Furze, M.E.; Huggett, B.A.; Aubrecht, D.M.; Stolz, C.D.; Carbone, M.S.; Richardson, A.D. Whole-tree nonstructural carbohydrate storage and seasonal dynamics in five temperate species. New Phytol. 2019, 221, 1466–1477. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, W.; Li, R.; Xu, M.; Wang, S. Different responses of non-structural carbohydrates in above-ground tissues/organs and root to extreme drought and re-watering in Chinese fir (Cunninghamia lanceolata) saplings. Trees 2016, 30, 1863–1871. [Google Scholar] [CrossRef]
- Signori-Müller, C.; Oliveira, R.S.; Barros, F.d.V.; Tavares, J.V.; Gilpin, M.; Diniz, F.C.; Zevallos, M.J.M.; Yupayccana, C.A.S.; Acosta, M.; Bacca, J.; et al. Non-structural carbohydrates mediate seasonal water stress across Amazon forests. Nat. Commun. 2021, 12, 2310. [Google Scholar] [CrossRef]
- De Roo, L.; Salomón, R.L.; Oleksyn, J.; Steppe, K. Woody tissue photosynthesis delays drought stress in Populus tremula trees and maintains starch reserves in branch xylem tissues. New Phytol. 2020, 228, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.S.; Gear, L.; Hultine, K.R.; Koch, G.W.; Ogle, K. Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 2020, 43, 1467–1483. [Google Scholar] [CrossRef] [PubMed]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural Carbon in Woody Plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef]
- Richardson, A.D.; Carbone, M.S.; Huggett, B.A.; Furze, M.E.; Czimczik, C.I.; Walker, J.C.; Xu, X.; Schaberg, P.G.; Murakami, P. Distribution and mixing of old and new nonstructural carbon in two temperate trees. New Phytol. 2015, 206, 590–597. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, L.; Dai, Y.; Wan, X.; Liu, S. Phenology-dependent variation in the non-structural carbohydrates of broadleaf evergreen species plays an important role in determining tolerance to defoliation (or herbivory). Sci. Rep. 2017, 7, 10125. [Google Scholar] [CrossRef]
- Wang, A.-Y.; Han, S.-J.; Zhang, J.-H.; Wang, M.; Yin, X.-H.; Fang, L.-D.; Yang, D.; Hao, G.-Y. The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient. Tree Physiol. 2018, 38, 1792–1804. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling. Plant Cell Environ. 2011, 34, 514–524. [Google Scholar] [CrossRef]
- Erbilgin, N.; Zanganeh, L.; Klutsch, J.G.; Chen, S.; Zhao, S.; Ishangulyyeva, G.; Burr, S.J.; Gaylord, M.; Hofstetter, R.; Keefover-Ring, K.; et al. Combined drought and bark beetle attacks deplete non-structural carbohydrates and promote death of mature pine trees. Plant Cell Environ. 2021, 44, 3866–3881. [Google Scholar] [CrossRef] [PubMed]
- Hoch, G.; Popp, M.; Körner, C. Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline. Oikos 2002, 98, 361–374. [Google Scholar] [CrossRef]
- McDowell, N.G. Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef]
- Piper, F.I.; Fajardo, A.; Hoch, G. Single-provenance mature conifers show higher non-structural carbohydrate storage and reduced growth in a drier location. Tree Physiol. 2017, 37, 1001–1010. [Google Scholar] [CrossRef]
- Han, H.; He, H.; Wu, Z.; Cong, Y.; Zong, S.; He, J.; Fu, Y.; Liu, K.; Sun, H.; Li, Y.; et al. Non-Structural Carbohydrate Storage Strategy Explains the Spatial Distribution of Treeline Species. Plants 2020, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Bastos, A.; Das, A.J.; Esquivel-Muelbert, A.; Hammond, W.M.; Martínez-Vilalta, J.; McDowell, N.G.; Powers, J.S.; Pugh, T.A.; Ruthrof, K.X.; et al. Climate Change Risks to Global Forest Health: Emergence of Unexpected Events of Elevated Tree Mortality Worldwide. Annu. Rev. Plant Biol. 2022, 73, 673–702. [Google Scholar] [CrossRef]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel competitors shape species’ responses to climate change. Nature 2015, 525, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Obser-vations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Petrik, P.; Petek-Petrik, A.; Kurjak, D.; Mukarram, M.; Klein, T.; Gömöry, D.; Střelcová, K.; Frýdl, J.; Konôpková, A. Interannual adjustments in stomatal and leaf morphological traits of European beech (Fagus sylvatica L.) demonstrate its climate change acclimation potential. Plant Biol. 2022, 24, 1287–1296. [Google Scholar] [CrossRef]
- Schuldt, B.; Knutzen, F.; Delzon, S.; Jansen, S.; Müller-Haubold, H.; Burlett, R.; Clough, Y.; Leuschner, C. How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction? New Phytol. 2016, 210, 443–458. [Google Scholar] [CrossRef]
- Hao, B.; Hartmann, H.; Li, Y.; Liu, H.; Shi, F.; Yu, K.; Li, X.; Li, Z.; Wang, P.; Allen, C.D.; et al. Precipitation Gradient Drives Divergent Relationship between Non-Structural Carbohydrates and Water Availability in Pinus tabulaeformis of Northern China. Forests 2021, 12, 133. [Google Scholar] [CrossRef]
- D’andrea, E.; Scartazza, A.; Battistelli, A.; Collalti, A.; Proietti, S.; Rezaie, N.; Matteucci, G.; Moscatello, S. Unravelling resilience mechanisms in forests: Role of non-structural carbohydrates in responding to extreme weather events. Tree Physiol. 2021, 41, 1808–1818. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Lutz, J.A.; Wang, L.; Shugart, H.H.; Yan, X. Using climate-driven leaf phenology and growth to improve predictions of gross primary productivity in North American forests. Glob. Chang. Biol. 2020, 26, 6974–6988. [Google Scholar] [CrossRef]
- Kono, Y.; Ishida, A.; Saiki, S.-T.; Yoshimura, K.; Dannoura, M.; Yazaki, K.; Kimura, F.; Yoshimura, J.; Aikawa, S.-I. Initial hydraulic failure followed by late-stage carbon starvation leads to drought-induced death in the tree Trema orientalis. Commun. Biol. 2019, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- McDowell, N.G.; Williams, A.P.; Xu, C.; Pockman, W.T.; Dickman, L.T.; Sevanto, S.; Pangle, R.E.; Limousin, J.M.; Plaut, J.J.; Mackay, D.S.; et al. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nat. Clim. Chang. 2016, 6, 295–300. [Google Scholar] [CrossRef]
- Keyimu, M.; Li, Z.; Wu, X.; Fu, B.; Liu, G.; Shi, S.; Fan, Z.; Wang, X. Recent decline of high altitude coniferous growth due to thermo-hydraulic constrains: Evidence from the Miyaluo Forest Reserve, Western Sichuan Plateau of China. Dendrochronologia 2020, 63, 125751. [Google Scholar] [CrossRef]
- He, M.; Wang, H.S.; Sun, J.X. Characters of ecosystem vulnerability in southwestern China based on vegetation productivity. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2019, 30, 429–438. [Google Scholar]
- Xu, K.; Wang, X.; Jiang, C.; Sun, O.J. Assessing the vulnerability of ecosystems to climate change based on climate exposure, vegetation stability and productivity. For. Ecosyst. 2020, 7, 23. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, C.; Wang, X.; Feng, Q.; Liu, X.; Tang, Z.; Sun, O.J. Nutrient trade-offs mediated by ectomycorrhizal strategies in plants: Evidence from an Abies species in subalpine forests. Ecol. Evol. 2021, 11, 5281–5294. [Google Scholar] [CrossRef]
- Li, Z.; Keyimu, M.; Fan, Z.; Wang, X. Climate sensitivity of conifer growth doesn’t reveal distinct low–high dipole along the elevation gradient in the Wolong National Natural Reserve, SW China. Dendrochronologia 2020, 61, 125702. [Google Scholar] [CrossRef]
- Chen, L.; Wang, M.; Jiang, C.; Wang, X.; Feng, Q.; Liu, X.; Sun, O.J. Choices of ectomycorrhizal foraging strategy as an important mechanism of environmental adaptation in Faxon fir (Abies fargesii var. faxoniana). For. Ecol. Manag. 2021, 495, 119372. [Google Scholar] [CrossRef]
- Zhang, W.R. The forest soils of Wolong Natural Reserve and its vertical zonalties distribution. Sci. Silvae Sin. 1983, 19, 254–268. [Google Scholar]
- Kang, W.; Tian, C.; Kang, D.; Wang, M.; Li, Y.; Wang, X.; Li, J. Effects of gap microsites and bamboo on Abies faxoniana regeneration in a subalpine forest, China. J. Plant Interact. 2015, 10, 59–64. [Google Scholar] [CrossRef]
- Florin, R. The distribution of conifer and taxad genera in time and space. Acta Horti Bergiani 1963, 20, 121–324. [Google Scholar]
- Xu, Y.; Gao, X.; Shen, Y.; Xu, C.; Shi, Y.; Giorgi, F. A daily temperature dataset over China and its application in validating a RCM simulation. Adv. Atmos. Sci. 2009, 26, 763–772. [Google Scholar] [CrossRef]
- Zhao, T.; Guo, W.; Fu, C. Calibrating and Evaluating Reanalysis Surface Temperature Error by Topographic Correction. J. Clim. 2008, 21, 1440–1446. [Google Scholar] [CrossRef]
- Alvarez-Uria, P.; Körner, C. Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct. Ecol. 2007, 21, 211–218. [Google Scholar] [CrossRef]
- Ferrar, P.J.; Cochrane, P.M.; Slatyer, R.O. Factors influencing germination and establishment of Eucalyptus pauciflora near the alpine tree line. Tree Physiol. 1988, 4, 27–43. [Google Scholar] [CrossRef]
- Pan, S.-A.; Hao, G.; Li, X.; Feng, Q.; Liu, X.; Sun, O.J. Altitudinal variations of hydraulic traits in Faxon fir (Abies fargesii var. faxoniana): Mechanistic controls and environmental adaptability. For. Ecosyst. 2022, 9, 100040. [Google Scholar] [CrossRef]
- Seifter, S.; Muntwyler, E.; Harkness, D.M. Some Effects of Continued Protein Deprivation, with and without Methionine Supplementation, on Intracellular Liver Components. Exp. Biol. Med. 1950, 75, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U. Components of leaf dry mass per area—Thickness and density—Alter photosynthetic capacity in reverse directions in woody plants. New Phytol. 1999, 144, 35–47. [Google Scholar] [CrossRef]
- Bertin, N.; Tchamitchian, M.; Baldet, P.; Devaux, C.; Brunel, B.; Gary, C. Contribution of carbohydrate pools to the variations in leaf mass per area within a tomato plant. New Phytol. 1999, 143, 53–61. [Google Scholar] [CrossRef]
- Mo, Q.; Chen, Y.; Yu, S.; Fan, Y.; Peng, Z.; Wang, W.; Li, Z.; Wang, F. Leaf nonstructural carbohydrate concentrations of understory woody species regulated by soil phosphorus availability in a tropical forest. Ecol. Evol. 2020, 10, 8429–8438. [Google Scholar] [CrossRef]
- Imaji, A.; Seiwa, K. Carbon allocation to defense, storage, and growth in seedlings of two temperate broad-leaved tree species. Oecologia 2010, 162, 273–281. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Carbohydrate sources and sinks in woody plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef]
- Sala, A.; Mencuccini, M. Plump trees win under drought. Nat. Clim. Chang. 2014, 4, 666–667. [Google Scholar] [CrossRef]
- Hasibeder, R.; Fuchslueger, L.; Richter, A.; Bahn, M. Summer drought alters carbon allocation to roots and root respiration in mountain grassland. New Phytol. 2015, 205, 1117–1127. [Google Scholar] [CrossRef]
- Tomasella, M.; Casolo, V.; Natale, S.; Petruzzellis, F.; Kofler, W.; Beikircher, B.; Mayr, S.; Nardini, A. Shade-induced reduction of stem nonstructural carbohydrates increases xylem vulnerability to embolism and impedes hydraulic recovery in Populus nigra. New Phytol. 2021, 231, 108–121. [Google Scholar] [CrossRef] [PubMed]
- McCulloh, K.A.; Johnson, D.M.; Meinzer, F.C.; Lachenbruch, B. An annual pattern of native embolism in upper branches of four tall conifer species. Am. J. Bot. 2011, 98, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, Z.; Zhang, K.; Cornelissen, J.H.C. Geographic pattern and effects of climate and taxonomy on nonstructural carbohydrates of Artemisia species and their close relatives across northern China. Biogeochemistry 2015, 125, 337–348. [Google Scholar] [CrossRef]
- Long, R.W.; Dudley, T.L.; D’Antonio, C.M.; Grady, K.C.; Bush, S.E.; Hultine, K.R. Spenders versus savers: Climate-induced carbon allocation trade-offs in a recently introduced woody plant. Funct. Ecol. 2021, 35, 1640–1654. [Google Scholar] [CrossRef]
- Zhang, T.; Cao, Y.; Chen, Y.; Liu, G. Non-structural carbohydrate dynamics in Robinia pseudoacacia saplings under three levels of continuous drought stress. Trees 2015, 29, 1837–1849. [Google Scholar] [CrossRef]
- Hoch, G.; Richter, A.; Körner, C. Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- Wingler, A.; Henriques, R. Sugars and the speed of life—Metabolic signals that determine plant growth, development and death. Physiol. Plant. 2022, 174, e13656. [Google Scholar] [CrossRef]
- Hansen, J.; Beck, E. Seasonal changes in the utilization and turnover of assimilation products in 8-year-old Scots pine (Pinus sylvestris L.) trees. Trees 1994, 8, 172–182. [Google Scholar] [CrossRef]
- Yano, R.; Nakamura, M.; Yoneyama, T.; Nishida, I. Starch-Related α-Glucan/Water Dikinase Is Involved in the Cold-Induced Development of Freezing Tolerance in Arabidopsis. Plant Physiol. 2005, 138, 837–846. [Google Scholar] [CrossRef]
- Dang, H.; Zhang, K.; Zhang, Q.; Xu, Y. Temporal variations of mobile carbohydrates in Abies fargesii at the upper tree limits. Plant Biol. 2015, 17, 106–113. [Google Scholar] [CrossRef]
- Bush, E.; Wilson, P.; Shepard, D.; McCrimmon, J. Freezing Tolerance and Nonstructural Carbohydrate Composition of Carpetgrass (Axonopus affinis Chase). Hortscience 2000, 35, 187–189. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Turco, M.; Rosa-Cánovas, J.J.; Bedia, J.; Jerez, S.; Montávez, J.P.; Llasat, M.C.; Provenzale, A. Exacerbated fires in Mediterranean Europe due to anthropogenic warming projected with non-stationary climate-fire models. Nat. Commun. 2018, 9, 3821. [Google Scholar] [CrossRef]
- Piper, F.I.; Paula, S. The Role of Nonstructural Carbohydrates Storage in Forest Resilience under Climate Change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, Z.; Wang, Z.; Chen, Y.; Wen, Z.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.-N.; Gessler, A.; Saurer, M.; Hagedorn, F.; Gao, D.-C.; Wang, X.-Y.; Schaub, M.; Li, M.-H.; Shen, W.-J.; Schönbeck, L. Root carbon and nutrient homeostasis determines downy oak sapling survival and recovery from drought. Tree Physiol. 2021, 41, 1400–1412. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W. Climate change and the ecologist. Nature 2007, 448, 550–552. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Pineda, E.; Sáenz-Romero, C.; Ortega-Rodríguez, J.M.; Blanco-García, A.; Madrigal-Sánchez, X.; Lindig-Cisneros, R.; Lopez-Toledo, L.; Pedraza-Santos, M.E.; Rehfeldt, G.E. Suitable climatic habitat changes for Mexican conifers along altitudinal gradients under climatic change scenarios. Ecol. Appl. 2020, 30, e02041. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).