
Divergent Responses of Temperature Sensitivity to Rising Incubation Temperature in Warmed and Un-Warmed Soil: A Mesocosm Experiment from a Subtropical Plantation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Soil Sampling
2.2. Laboratory Incubation Experiment
2.3. Soil Chemical Analyses
2.4. Microbial Biomass C and N
2.5. Enzyme Analysis
2.6. Phospholipid Fatty Acids (PLFAs) Analysis
2.7. Calculation of Organic Carbon Mineralization Rate, Cumulative Mineralization and Temperature Sensitivity
2.8. Statistical Analyses
3. Results
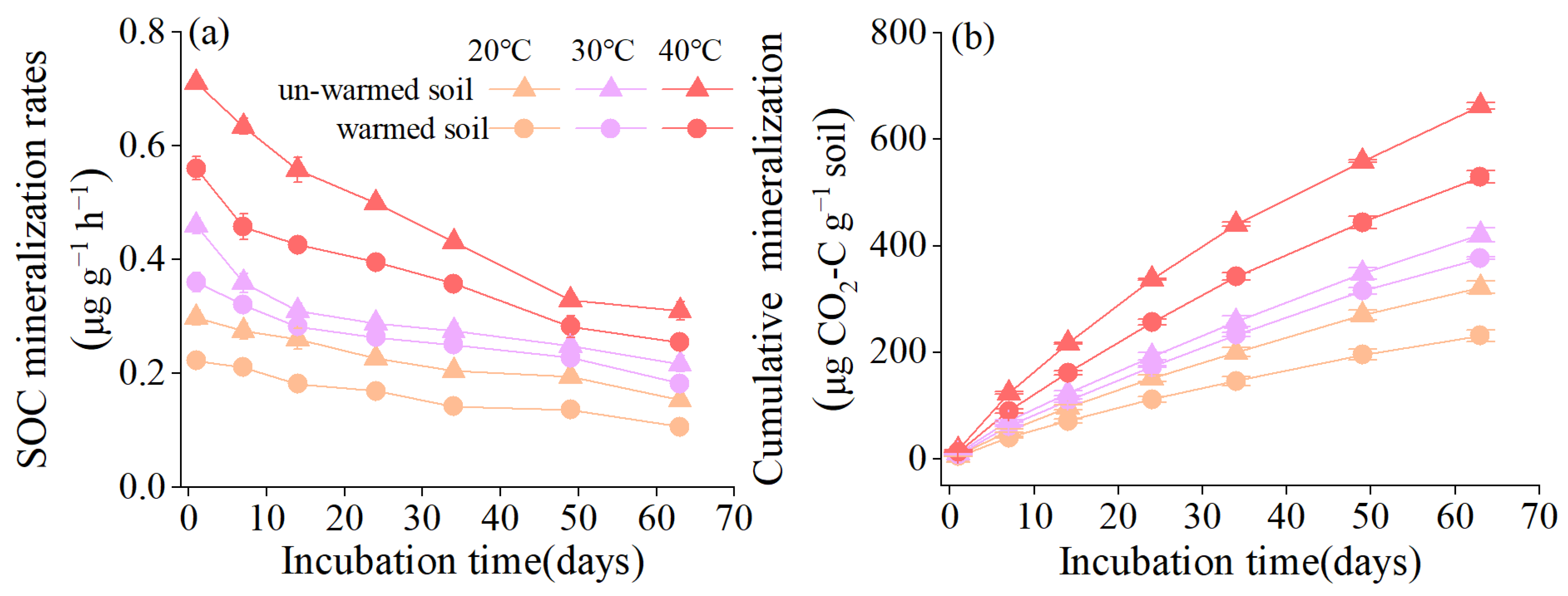
3.1. Soil Organic Carbon Mineralization and Its Temperature Sensitivity
3.2. Soil Nutrients, MBC, Microbial Metabolic Quotients
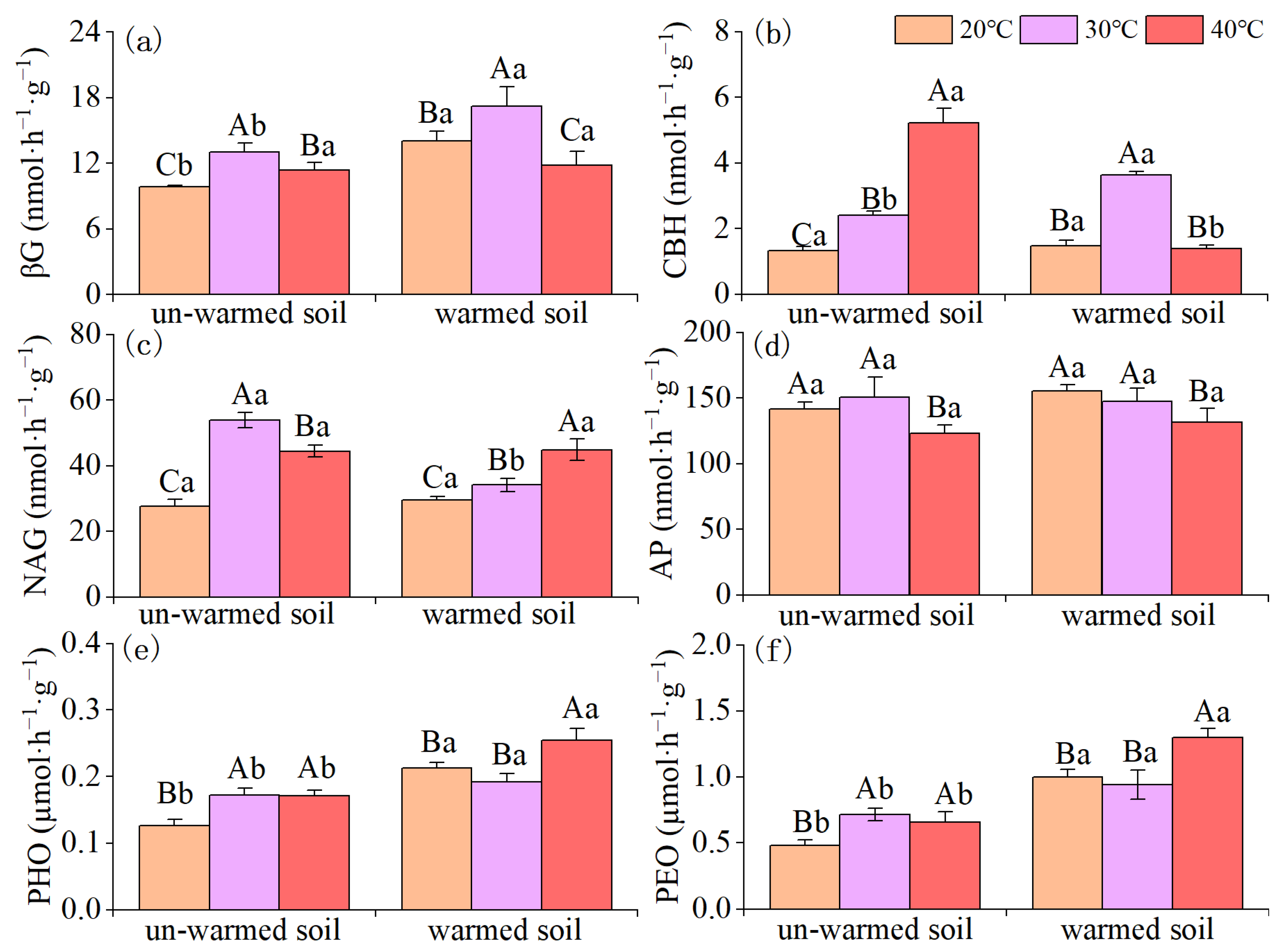
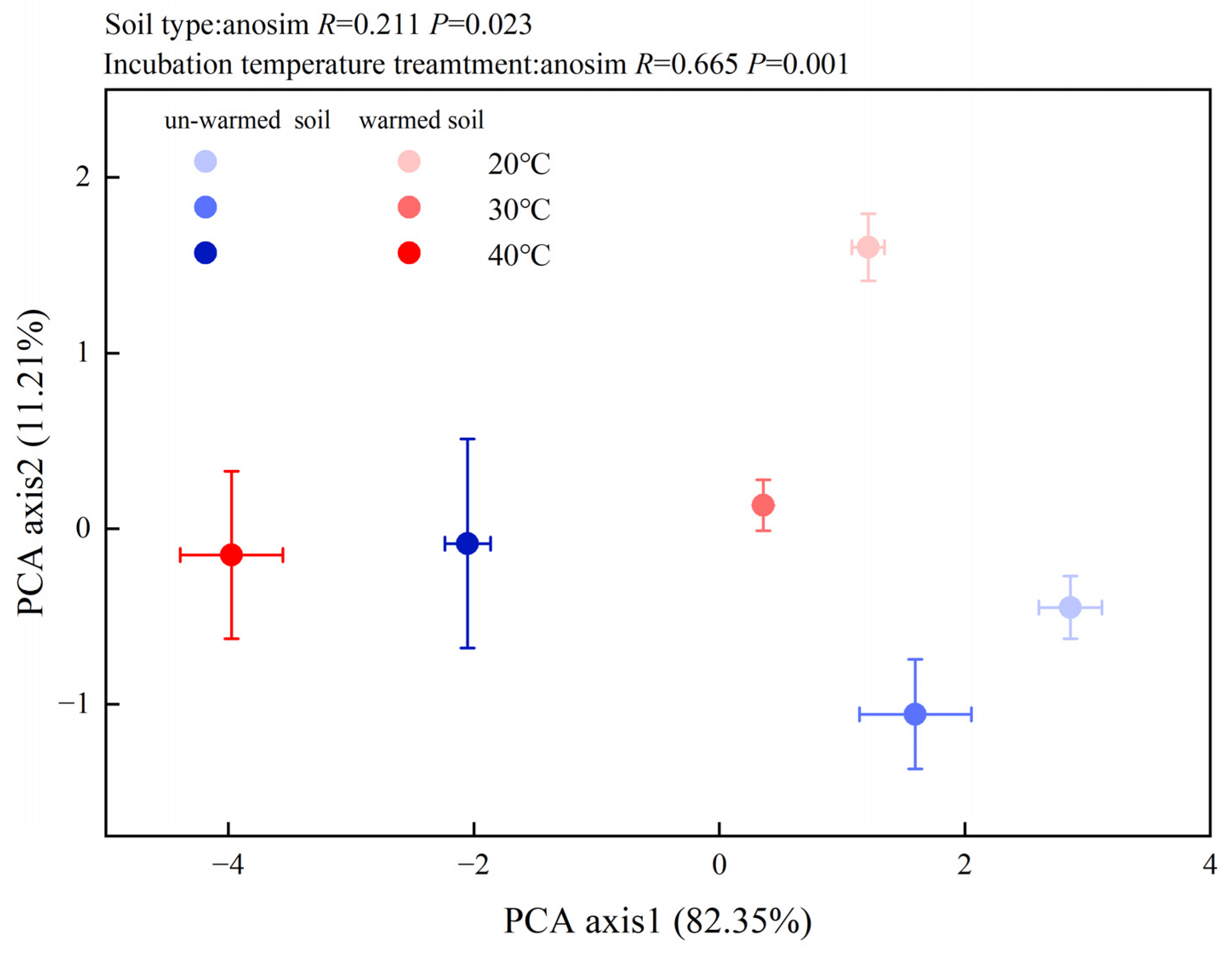
3.3. Soil Enzyme Activities and Microbial PLFAs
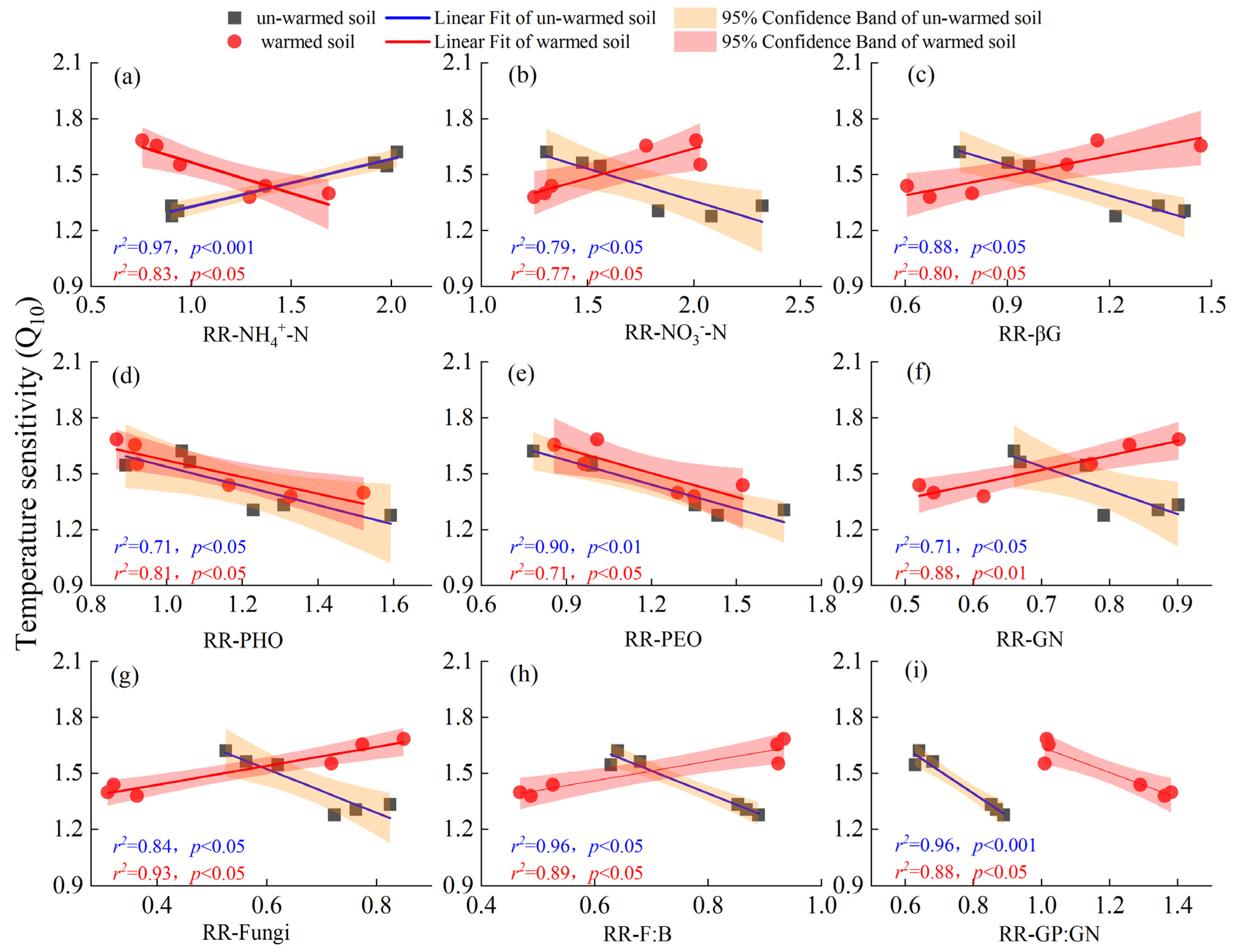
3.4. Factors Affecting the Temperature Sensitivity of SOC Mineralization
4. Discussion
4.1. Response of SOC Mineralization of Un-Warmed and Warmed Soil to Warming
4.2. Response of the Q10 of SOC Mineralization of Un-Warmed and Warmed Soil to Rising Incubation Temperature
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R. (Eds.) Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Bond-Lamberty, B.; Thomson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Lugo, A.E. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 1982, 14, 161–187. [Google Scholar] [CrossRef]
- Litton, C.M.; Giardina, C.P.; Albano, J.K.; Long, M.S.; Asner, G.P. The magnitude and variability of soil-surface CO2 efflux increase with mean annual temperature in Hawaiian tropical montane wet forests. Biochemistry 2011, 43, 2315–2323. [Google Scholar] [CrossRef]
- Giardina, C.P.; Litton, C.M.; Crow, S.E.; Asner, G.P. Warming-related increases in soil CO2 efflux are explained by increased below-ground carbon flux. Nat. Clim. Chang. 2014, 4, 822–827. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Semenov, M.V.; Yao, F.; Ye, J.I.; Bu, R.; Ma, R.; Lin, J.; Kurganova, I.; Wang, X.; et al. Temperature sensitivity of SOM decomposition is linked with a K-selected microbial community. Glob. Chang. Biol. 2021, 27, 2763–2779. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Kou, D.; Mao, C.; Chen, Y.; Chen, L.; Yang, Y. Temperature sensitivity of permafrost carbon release mediated by mineral and microbial properties. Sci. Adv. 2021, 7, eabe3596. [Google Scholar] [CrossRef]
- Liu, X.F.; Yang, Z.J.; Lin, C.F.; Giardina, C.P.; Xiong, D.C.; Lin, W.S.; Chen, S.D.; Xu, C.; Chen, G.S.; Xie, J.S.; et al. Will nitrogen deposition mitigate warming-increased soil respiration in a young subtropical plantation? Agric. For. Meteorol. 2017, 246, 78–85. [Google Scholar] [CrossRef]
- Wang, Q.K.; Liu, S.; Tian, P. Carbon quality and soil microbial property control the latitudinal pattern in temperature sensitivity of soil microbial respiration across Chinese forest ecosystems. Glob. Chang. Biol. 2018, 24, 2841–2849. [Google Scholar] [CrossRef]
- Wang, X.W.; Li, X.Z.; Hu, Y.M.; Lv, J.J.; Sun, J.; Li, Z.M.; Wu, Z.F. Effect of temperature and moisture on soil organic carbon mineralization of predominantly permafrost peatland in the Great Hing’an Mountains, Northeastern China. J. Environ. Sci. 2010, 22, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Liu, C.; Song, C.C.; Wang, X.W.; Ma, X.Y.; Gao, J.L.; Gao, S.Q.; Wang, L.L. Linking soil organic carbon mineralization with soil microbial and substrate properties under warming in permafrost peatlands of Northeastern China. Catena 2021, 203, 105348. [Google Scholar] [CrossRef]
- Jiang, L.; Ma, X.; Song, Y.Y.; Gao, S.Q.; Ren, J.S.; Zhang, H.; Wang, X.W. Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland. Microorganisms 2022, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Pold, G.; Grandy, A.S.; Melillo, J.M.; Deangelis, K.M. Changes in substrate availability drive carbon cycle response to chronic warming. Soil Biol. Biochem. 2017, 110, 68–78. [Google Scholar] [CrossRef]
- Li, X.J.; Xie, J.S.; Zhang, Q.F.; Lyu, M.K.; Xiong, X.L.; Liu, X.F.; Lin, T.C.; Yang, Y.S. Substrate availability and soil microbes drive temperature sensitivity of soil organic carbon mineralization to warming along an elevation gradient in subtropical Asia. Geoderma 2020, 364, 114198. [Google Scholar] [CrossRef]
- Hassan, W.; Bano, R.; Khatak, B.U.; Hussain, I.; Yousaf, M.; David, J. Temperature sensitivity and soil organiccarbon pools decomposition under different moisture regimes: Effect on total microbial and enzymatic activity. CLEAN-Soil Air Water 2015, 43, 391–398. [Google Scholar] [CrossRef]
- Domeignoz-Horta, L.A.; Pold, G.; Erb, H.; Sebag, D.; Verrecchia, E.; Northen, T.; Louie, K.; Eloe-Fadrosh, E.; Pennacchio, C.; Knorr, M.A.; et al. Substrate availability and not thermal acclimation controls microbial temperature sensitivity response to long-term warming. Glob. Chang. Biol. 2023, 29, 1574–1590. [Google Scholar] [CrossRef]
- Zelikova, T.J.; Housman, D.C.; Grote, E.E.; Neher, D.A.; Belnap, J. Warming and increased precipitation frequency on the Colorado plateau: Implications for biological soil crusts and soil processes. Plant Soil 2012, 355, 265–282. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Xue, K.; Xie, J.P.; Deng, Y.; Wu, L.Y.; Cheng, X.H.; Fei, S.F.; Deng, S.P.; He, Z.L.; Van Nostrand, J.D.; et al. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Chang. 2012, 2, 106–110. [Google Scholar] [CrossRef]
- Karhu, K.; Auffret, M.D.; Dungait, J.A.J.; Hopkins, D.W.; Prosser, J.I.; Singh, B.K.; Subke, J.-A.; Wookey, P.A.; Ågren, G.I.; Sebastià, M.-T.; et al. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 2014, 513, 81–84. [Google Scholar] [CrossRef]
- Wang, C.; Morrissey, E.M.; Mau, R.L.; Hayer, M.; Piñeiro, J.; Mack, M.C.; Marks, J.C.; Bell, S.L.; Miller, S.N.; Schwartz, E.; et al. The temperature sensitivity of soil: Microbial biodiversity, growth, and carbon mineralization. ISME J. 2021, 15, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.; Chen, H.; Liu, H.; Nie, M. Enzymic moderations of bacterial and fungal communities on short-and long-term warming impacts on soil organic carbon. Sci. Total Environ. 2022, 804, 150197. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Liu, X.R.; Li, Y.; Ran, Y.; Liu, Y.P.; Zhang, Q.C.; Li, Z.; He, Y.; Xu, J.H.; Di, H.J. High temperatures inhibited the growth of soil bacteria and archaea but not that of fungi and altered nitrous oxide production mechanisms from different nitrogen sources in an acidic soil. Soil Biol. Biochem. 2017, 107, 168–179. [Google Scholar] [CrossRef]
- Li, Y.Q.; Qing, Y.X.; Lyu, M.K.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Yang, Y.S. Effects of artificial warming on different soil organic carbon and nitrogen pools in a subtropical plantation. Soil Biol. Biochem. 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Meir, P.; Velasquez, E.; Turner, B.L. Soil carbon loss by experimental warming in a tropical forest. Nature 2020, 584, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.L.; Wheeler, C.E.; Mitchard, E.T.; Koch, A. Regenerate natural forests to store carbon. Nature 2019, 568, 25–28. [Google Scholar] [CrossRef]
- Yang, Z.J.; Chen, S.D.; Liu, X.F.; Xiong, D.C.; Xu, C.; Arthur, M.A.; McCulley, R.L.; Shi, S.H.; Yang, Y.S. Loss of soil organic carbon following natural forest conversion to Chinese fir plantation. For. Ecol. Manag. 2019, 449, 117476. [Google Scholar] [CrossRef]
- Liu, X.F.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Xiong, D.C.; Lin, W.S.; Xu, C.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; et al. Will heterotrophic soil respiration be more sensitive to warming than autotrophic respiration in subtropical forests? Warming effect on soil respiration. Eur. J. Soil Sci. 2019, 70, 655–663. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014. In International Soil Classification System for Naming soils and Creating Legends for Soil Maps; World Soil Resources Reports No.106; FAO: Rome, Italy, 2014. [Google Scholar]
- Hamdi, S.; Chevallier, T.; Aïssa, N.B.; Hammouda, M.B.; Gallali, T.; Chotte, J.; Bernoux, M. Short-term temperature dependence of heterotrophic soil respiration after one-month of pre-incubation at different temperatures. Soil Biol. Biochem. 2011, 43, 1752–1758. [Google Scholar] [CrossRef]
- Huang, J.X.; Lin, T.C.; Xiong, D.C.; Yang, Z.J.; Liu, X.F.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; Yang, Y.S. Organic carbon mineralization in soils of a natural forest and a forest plantation of southeastern China. Geoderma 2019, 344, 119–126. [Google Scholar] [CrossRef]
- Jones, D.; Willett, V. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil sampling and methods of analysis. In Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, FL, USA, 2007; Chapter 19; pp. 207–214. [Google Scholar]
- Vance, E.; Brookes, P.; Jenkinson, D. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Xu, G.; Chen, J.; Berninger, F.; Pumpanen, J.; Bai, J.; Yu, L.; Duan, B. Labile, recalcitrant, microbial carbon and nitrogen and the microbial community composition at two Abies faxoniana forest elevations under elevated temperatures. Soil Biol. Biochem. 2015, 91, 1–13. [Google Scholar] [CrossRef]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: Lessons learned about soil quality and indications for improving sustainability. Soil Biol. Biochem. 2010, 42, 1–13. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. Effects of long-term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E.; McClaugherty, C.A.; Rayburn, L.; Repert, D.A.; Weiland, T. Wood decomposition over a first-order watershed: Mass loss as a function of lignocellulase activity. Soil Biol. Biochem. 1992, 24, 743–749. [Google Scholar] [CrossRef]
- Wan, X.H.; Huang, Z.Q.; He, Z.M.; Yu, Z.P.; Wang, M.H.; Davis, M.R.; Yang, Y.S. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- McKinley, V.L.; Peacock, A.D.; White, D.C. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biol. Biochem. 2005, 37, 1946–1958. [Google Scholar] [CrossRef]
- Hu, R.G.; Hatano, R.; Kusa, K.; Sawamoto, T. Soil respiration and net ecosystem production in an onion field in Central Hokkaido, Japan. Soil Sci. Plant Nutr. 2004, 50, 27–33. [Google Scholar] [CrossRef]
- Creamer, C.A.; De Menezes, A.B.; Krull, E.S.; Sanderman, J.; Newton-Walters, R.; Farrell, M. Microbial community structure mediates response of soil C decomposition to litter addition and warming. Soil Biol. Biochem. 2015, 80, 175–188. [Google Scholar] [CrossRef]
- Tian, Q.X.; Wang, X.G.; Wang, D.Y.; Wang, M.; Liao, C.; Yang, X.L.; Liu, F. Decoupled linkage between soil carbon and nitrogen mineralization among soil depths in a subtropical mixed forest. Soil Biol. Biochem. 2017, 109, 135–144. [Google Scholar] [CrossRef]
- Lefcheck, J.S. Piecewise SEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Gudasz, C.; Bastviken, D.; Steger, K.; Premke, K.; Sobek, S.; Tranvik, L.J. Temperature-controlled organic carbon mineralization in lake sediments. Nature 2010, 466, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.K.; Wang, S.L.; He, T.X.; Liu, L.; Wu, J.W. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Wang, Q.K.; Zeng, Z.Q.; Zhong, M.C. Soil moisture alters the response of soil organic carbon mineralization to litter addition. Ecosystems 2016, 19, 450–460. [Google Scholar] [CrossRef]
- Bai, E.; Li, S.L.; Xu, W.H.; Li, W.; Dai, W.W.; Jiang, P. A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics. New Phytol. 2013, 199, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Dawes, M.A.; Schleppi, P.; Hättenschwiler, S.; Rixen, C.; Hagedorn, F. Soil warming opens the nitrogen cycle at the alpine treeline. Glob. Chang. Biol. 2017, 23, 421–434. [Google Scholar] [CrossRef]
- Vogel, J.G.; Valentine, D.W.; Ruess, R.W. Soil and root respiration in mature Alaskan black spruce forests that vary in soil organic matter decomposition rates. Can. J. For. Res. 2005, 35, 161–174. [Google Scholar] [CrossRef]
- Zhou, P.; Li, Y.; Ren, X.E.; Xiao, H.A.; Tong, C.L.; Ge, T.; Brookes, P.; Shen, J.L.; Wu, J.S. Organic carbon mineralization responses to temperature increases in subtropical paddy soils. J. Soils Sediments 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Niu, B.; Zhang, X.Z.; Piao, S.L.; Janssens, I.A.; Fu, G.; He, Y.; He, Y.T.; Zhang, Y.J.; Shi, P.L.; Dai, E.; et al. Warming homogenizes apparent temperature sensitivity of ecosystem respiration. Sci. Adv. 2021, 7, eabc7358. [Google Scholar] [CrossRef]
- Bradford, M.A.; Davies, C.A.; Frey, S.D.; Maddox, T.R.; Mohan, J.E.; Reynolds, J.F.; Treseder, K.K.; Wallenstein, M.D. Thermal adaptation of soil microbial respiration to elevated temperature. Ecol. Lett. 2008, 11, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, S.; Moyano, F.; Sall, S.; Bernoux, M.; Chevallier, T. Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions. Soil Biol. Biochem. 2013, 58, 115–126. [Google Scholar] [CrossRef]
- Gershenson, A.; Bader, N.E.; Cheng, W.C. Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition. Glob. Chang. Biol. 2009, 15, 176–183. [Google Scholar] [CrossRef]
- Su, J.; Zhang, H.Y.; Han, X.G.; Penuelas, J.; Filimonenko, E.; Jiang, Y.; Kuzyakov, Y.; Wei, C.Z. Low carbon availability in paleosols nonlinearly attenuates temperature sensitivity of SOM decomposition. Glob. Chang. Biol. 2022, 28, 4180–4193. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.H.; Domsch, K.H. Soil microbial biomass: The eco-physiological approach. Soil Biol. Biochem. 2010, 42, 2039–2043. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kuzyakov, Y.; Sanaullah, M.; Heitkamp, F.; Zelenev, V.; Kumar, A.; Blagodatskaya, E. Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues: Mechanisms and thresholds. Biol. Fertil. Soils 2017, 53, 287–301. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.M.; Jehmlich, N.; von Bergen, M.; Griffiths, R.I.; Gleixner, G. Soil fungal:bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Xing, B.; Dukes, J.S. Warming alters potential enzyme activity but precipitation regulates chemical transformations in grass litter exposed to simulated climatic changes. Soil Biol. Biochem. 2014, 75, 102–112. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Follstad Shah, J.J. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef]
- Ding, F.; Sun, W.J.; Huang, Y.; Hu, X.Y. Larger Q10 of carbon decomposition in finer soil particles does not bring long-lasting dependence of Q10 on soil texture. Eur. J. Soil Sci. 2018, 69, 336–347. [Google Scholar] [CrossRef]
- Xu, X.; Luo, Y.Q.; Zhou, J.Z. Carbon quality and the temperature sensitivity of soil organic carbon decomposition in a tallgrass prairie. Soil Biol. Biochem. 2012, 50, 142–148. [Google Scholar] [CrossRef]
- Bai, Z.; Ma, Q.; Wu, X.; Zhang, Y.L.; Yu, W.T. Temperature sensitivity of a PLFA-distinguishable microbial community differs between varying and constant temperature regimes. Geoderma 2017, 308, 54–59. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | SOC | TN | NH4+-N | NO3−-N | DOC | MBC | MBN |
|---|---|---|---|---|---|---|---|---|
| (mg·g−1) | (mg·g−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | ||
| un-warmed soil | 4.31 ± 0.08 A | 13.02 ± 1.14 A | 1.12 ± 0.08 A | 4.48 ± 0.60 A | 2.04 ± 0.54 A | 13.58 ± 1.46 A | 285.50 ± 20.74 A | 24.60 ± 2.20 A |
| warmed soil | 4.27 ± 0.09 A | 11.60 ± 1.38 A | 0.98 ± 0.09 A | 4.08 ± 1.38 A | 2.32 ± 0.45 A | 9.86 ± 2.24 B | 203.55 ± 28.75 B | 19.07 ± 2.43 B |
| Treatment | Cumulative SOC Mineralization | ||||||
|---|---|---|---|---|---|---|---|
| 1 d | 7 d | 14 d | 24 d | 34 d | 49 d | 63 d | |
| Temperature | 1010.31 *** | 807.56 *** | 1130.97 *** | 1718.71 *** | 1449.05 *** | 1380.53 *** | 1515.73 *** |
| Soil type | 249.58 *** | 215.57 *** | 228.79 *** | 386.28 *** | 307.45 *** | 308.32 *** | 349.39 *** |
| Temperature × Soil type | 10.36 *** | 37.93 *** | 45.45 *** | 58.25 *** | 39.60 *** | 31.91 *** | 29.24 *** |
| Treatment | NH4+-N | NO3−-N | DON | DOC | MBC | MBN | Microbial Quotient | Metabolic Quotient | |
|---|---|---|---|---|---|---|---|---|---|
| (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (%) | (mg CO2-C g−1 MBC h−1) | ||
| un-warmed soil | 20 °C | 15.50 ± 0.76 Ba | 7.73 ± 0.67 Ca | 1.42 ± 0.27 Ba | 26.03 ± 1.99 Aa | 233.18 ± 14.38 Aa | 23.31 ± 3.11 Aa | 1.80 ± 0.13 Aa | 0.66 ± 0.06 Ca |
| 30 °C | 14.14 ± 0.55 Ba | 11.08 ± 0.76 Bb | 2.57 ± 0.8 Ba | 18.67 ± 1.68 Ba | 167.65 ± 9.02 Ba | 25.82 ± 4.54 Aa | 1.34 ± 0.22 Ba | 1.30 ± 0.09 Ba | |
| 40 °C | 27.88 ± 0.72 Aa | 15.96 ± 0.51 Ab | 5.68 ± 0.18 Aa | 15.88 ± 0.43 Ca | 94.26 ± 6.12 Ca | 17.54 ± 2.28 Ba | 0.78 ± 0.07 Ca | 3.30 ± 0.38 Aa | |
| warmed soil | 20 °C | 15.95 ± 1.47 Ba | 6.88 ± 0.28 Ca | 1.48 ± 0.37 Ba | 15.71 ± 1.05 Ab | 178.58 ± 12.37 Ab | 18.03 ± 3.31 Aa | 1.44 ± 0.15 Ab | 0.60 ± 0.08 Ca |
| 30 °C | 13.46 ± 2.09 Ba | 13.32 ± 0.66 Ba | 1.71 ± 0.15 Ba | 13.45 ± 0.74 Bb | 136.89 ± 9.21 Bb | 20.69 ± 2.27 Aa | 1.15 ± 0.08 Ba | 1.34 ± 0.10 Ba | |
| 40 °C | 19.22 ± 0.63 Ab | 17.20 ± 0.30 Aa | 4.54 ± 0.41 Ab | 10.96 ± 1.08 Cb | 79.23 ± 7.18 Cb | 13.32 ± 1.72 Bb | 0.67 ± 0.06 Ca | 3.23 ± 0.25 Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Yang, Z.; Zhou, J.; Zheng, W.; Chen, S.; Lin, W.; Xiong, D.; Xu, C.; Liu, X.; Yang, Y. Divergent Responses of Temperature Sensitivity to Rising Incubation Temperature in Warmed and Un-Warmed Soil: A Mesocosm Experiment from a Subtropical Plantation. Forests 2023, 14, 2164. https://doi.org/10.3390/f14112164
Zheng Y, Yang Z, Zhou J, Zheng W, Chen S, Lin W, Xiong D, Xu C, Liu X, Yang Y. Divergent Responses of Temperature Sensitivity to Rising Incubation Temperature in Warmed and Un-Warmed Soil: A Mesocosm Experiment from a Subtropical Plantation. Forests. 2023; 14(11):2164. https://doi.org/10.3390/f14112164
Chicago/Turabian StyleZheng, Yong, Zhijie Yang, Jiacong Zhou, Wei Zheng, Shidong Chen, Weisheng Lin, Decheng Xiong, Chao Xu, Xiaofei Liu, and Yusheng Yang. 2023. "Divergent Responses of Temperature Sensitivity to Rising Incubation Temperature in Warmed and Un-Warmed Soil: A Mesocosm Experiment from a Subtropical Plantation" Forests 14, no. 11: 2164. https://doi.org/10.3390/f14112164
APA StyleZheng, Y., Yang, Z., Zhou, J., Zheng, W., Chen, S., Lin, W., Xiong, D., Xu, C., Liu, X., & Yang, Y. (2023). Divergent Responses of Temperature Sensitivity to Rising Incubation Temperature in Warmed and Un-Warmed Soil: A Mesocosm Experiment from a Subtropical Plantation. Forests, 14(11), 2164. https://doi.org/10.3390/f14112164