
Vertical Stratification and Seasonal Dynamics of Coleoptera in Open Biotopes of Forest Ecosystems (Centre of European Russia)



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Procedures
2.2. Identification and Taxonomic Position of Samples
2.3. Data Analyses
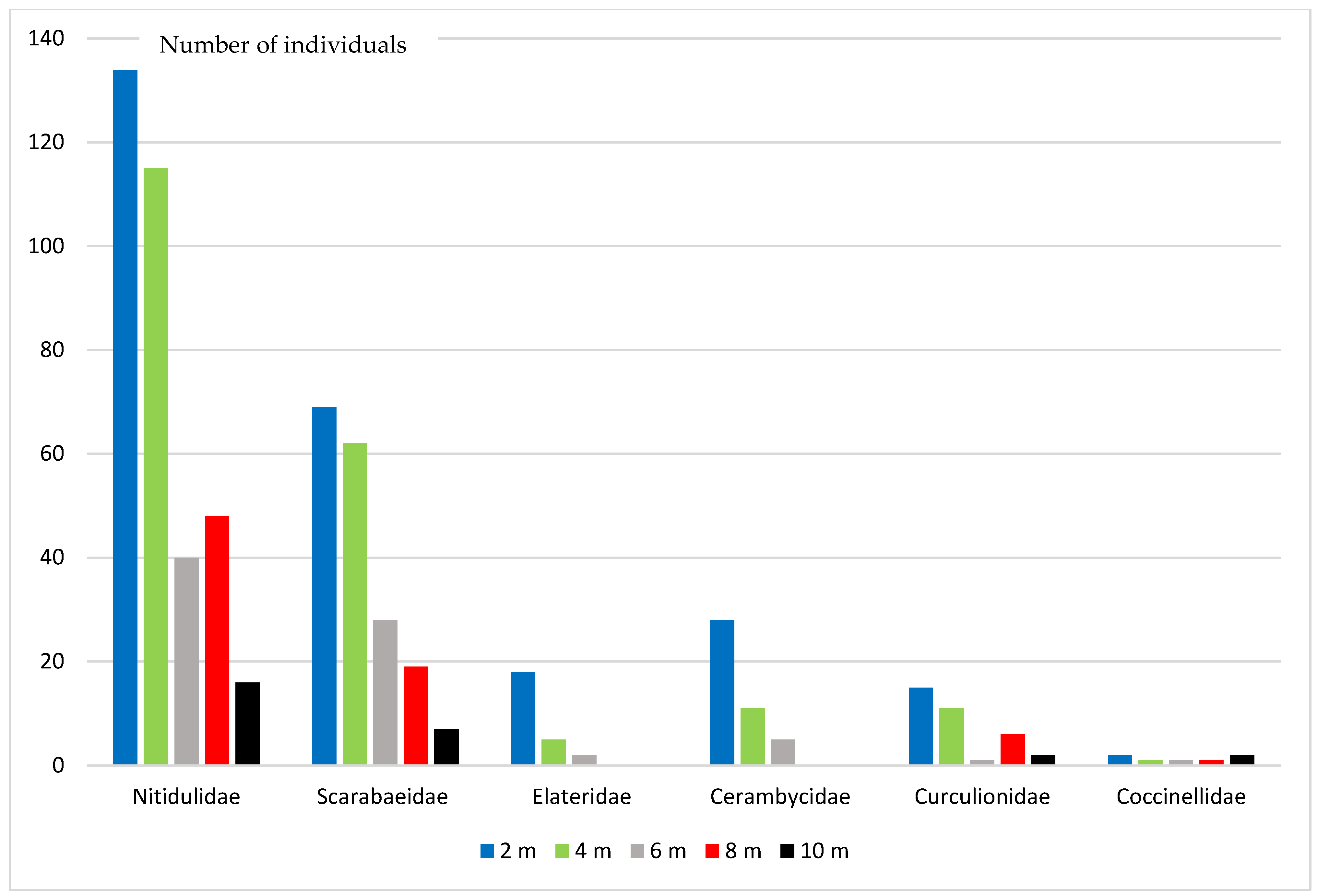
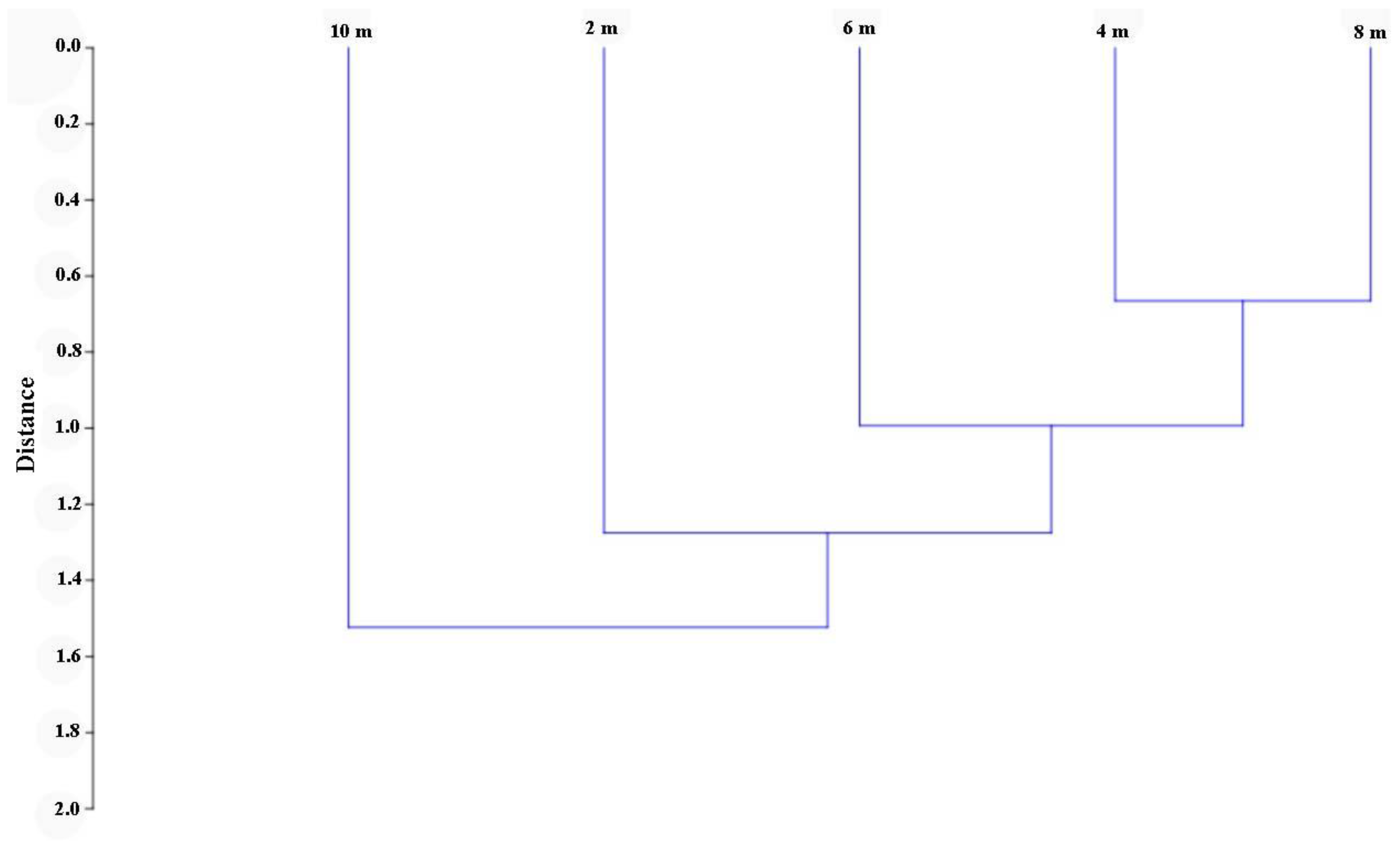
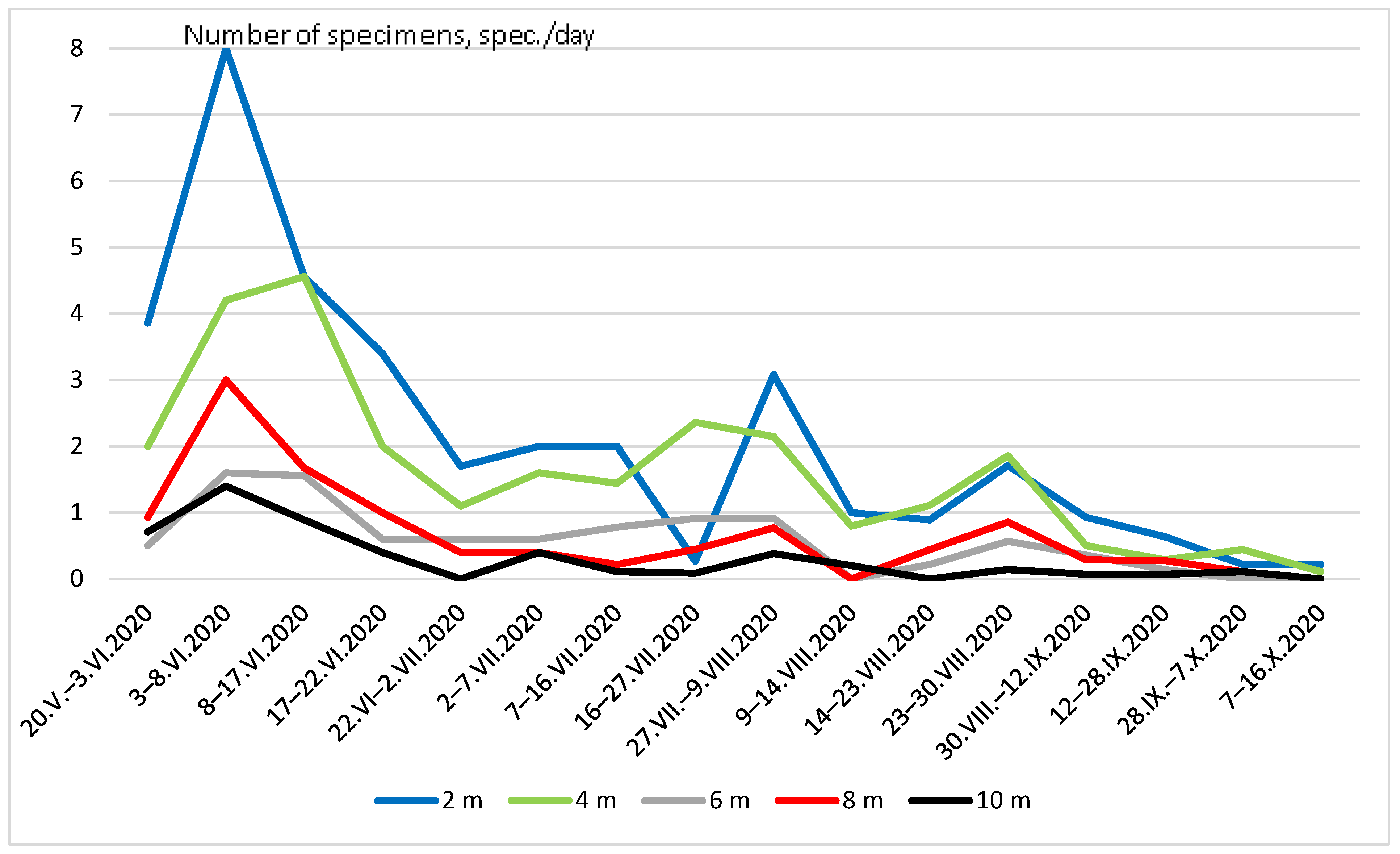
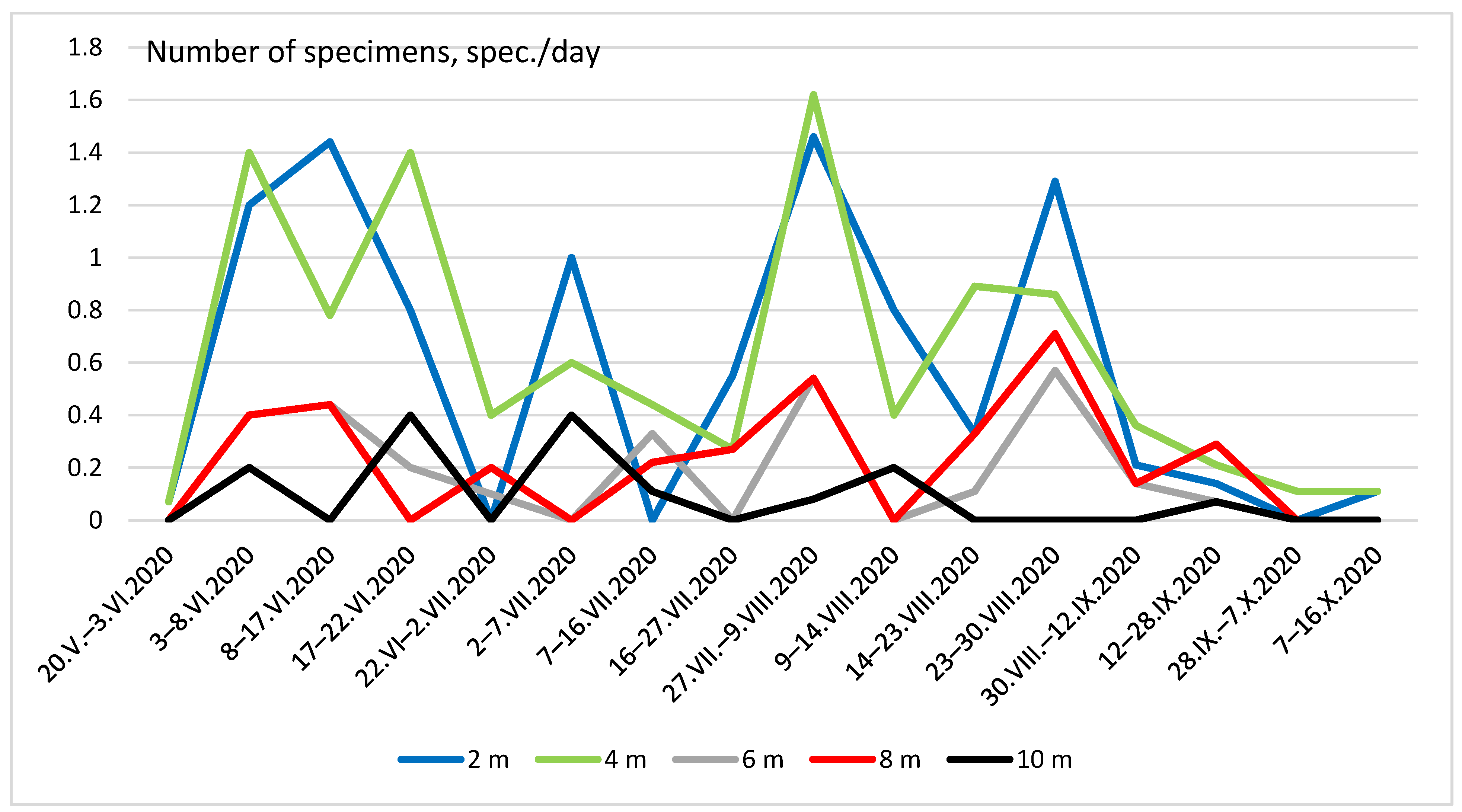
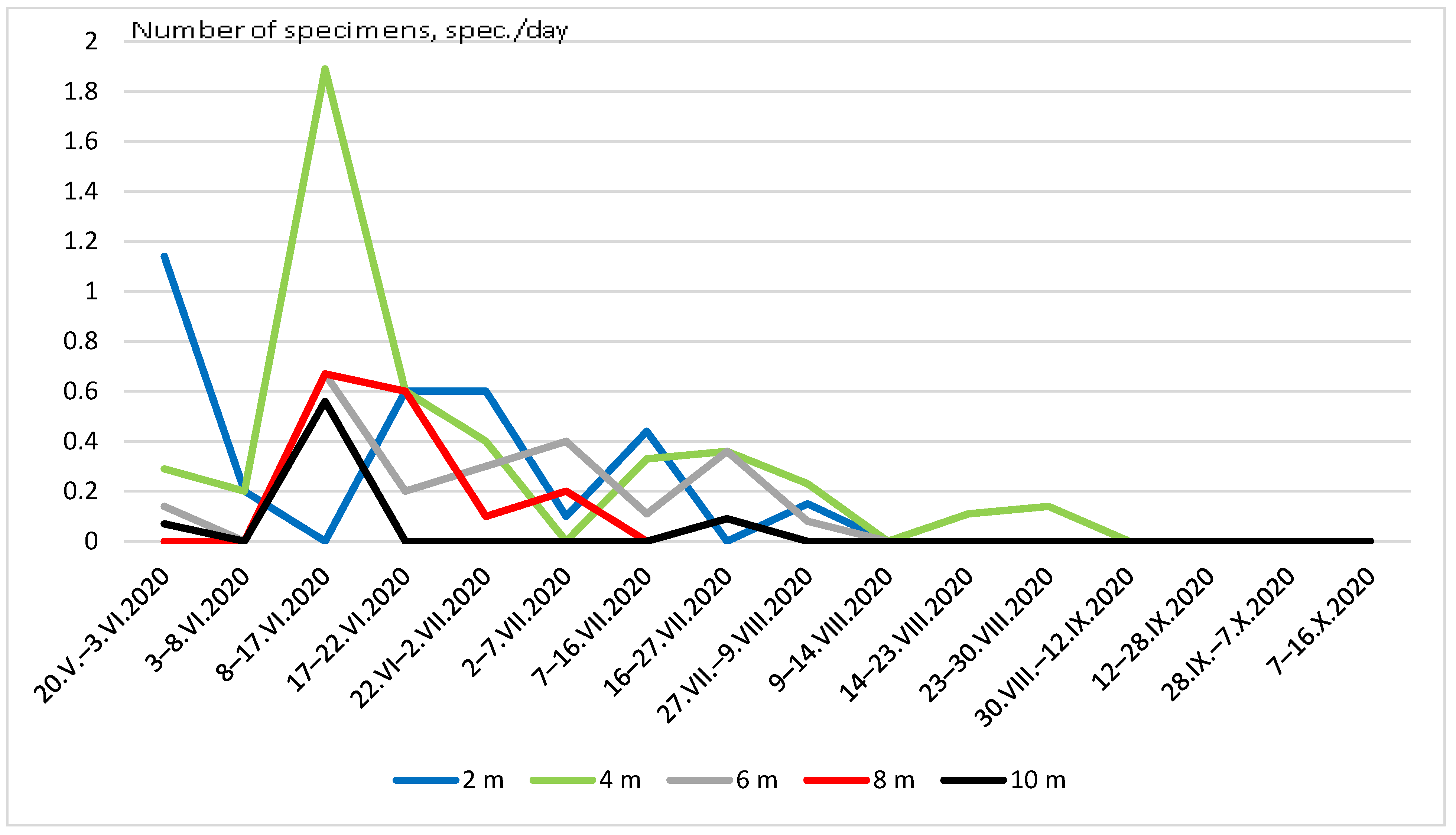
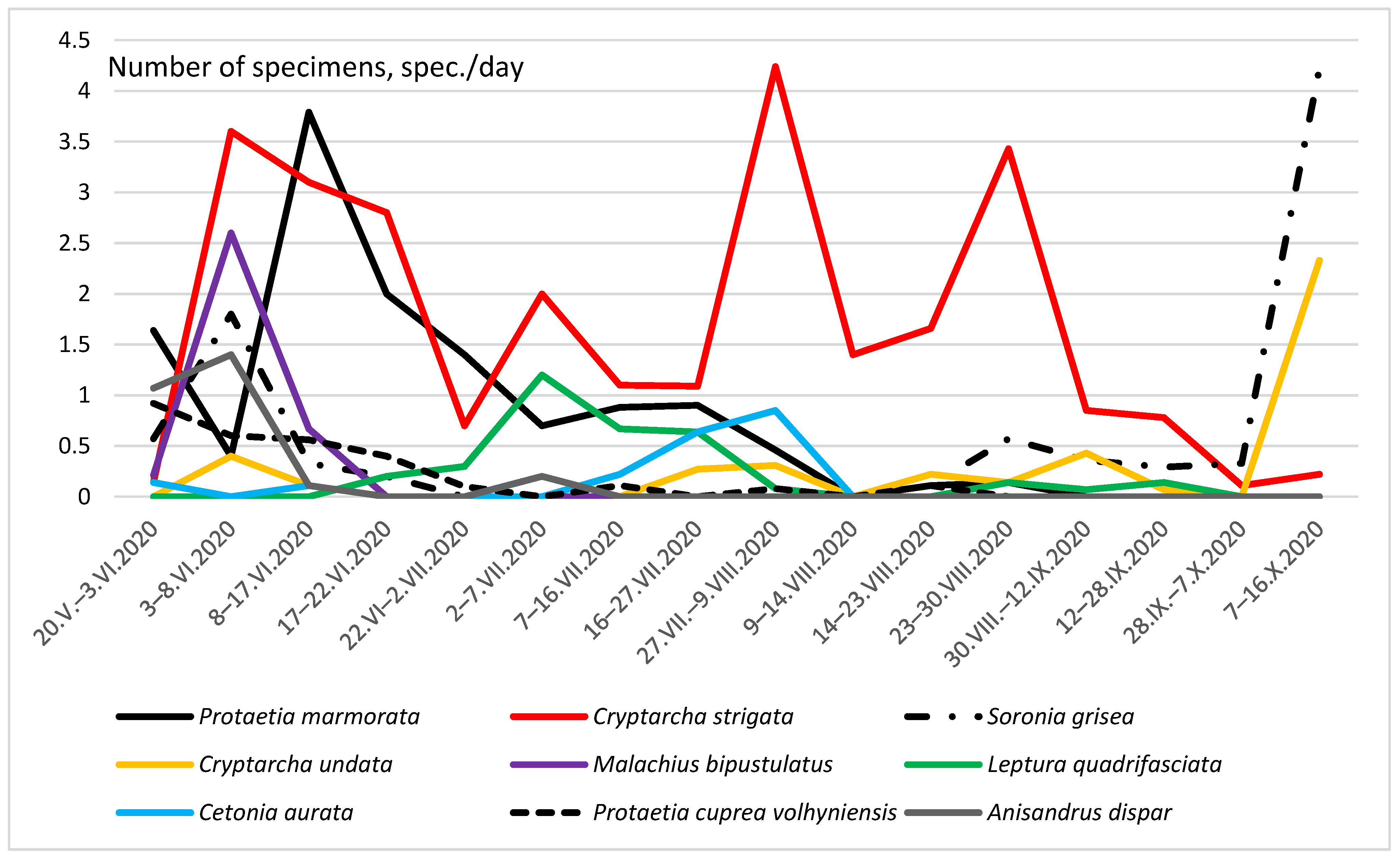
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Ried, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [PubMed]
- Olsson, J.; Johansson, T.; Jonsson, B.G.; Hjältén, J.; Edman, M.; Ericson, L. Landscape and substrate properties affect species richness and community composition of saproxylic beetles. For. Ecol. Manag. 2012, 286, 108–120. [Google Scholar] [CrossRef]
- Dedyukhin, S.V. Phytophagous beetles (Coleoptera: Chrysomelidae and Curculionoidea), protected and recommended for protection in the regions of the Middle Volga and the Urals. Nat. Conserv. Res. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Glotov, S.V.; Hushtan, K.V. Rove beetles of the subfamily Aleocharinae (Coleoptera: Staphylinidae) from the Hutsulshchyna National Nature Park. Biosyst. Divers. 2020, 28, 364–369. [Google Scholar] [CrossRef]
- Sundukov, Y.N.; Makarov, K.V. The ground beetles of the tribus Trechini (Carabidae) on the Southern Kuril Islands. Nat. Conserv. Res. 2021, 6, 15–51. [Google Scholar] [CrossRef]
- Prokin, A.A.; Sazhnev, A.S.; Philippov, D.A. Water beetles (Insecta: Coleoptera) of some peatlands in the North Caucasus. Nat. Conserv. Res. 2019, 4, 57–66. [Google Scholar] [CrossRef]
- Bondarenko, A.S.; Zamotajlov, A.S.; Belyi, A.I.; Khomitskiy, E.E. Fauna and ecological characteristics of ground beetles (Coleoptera, Carabidae) of the Nature Sanctuaries «Prichernomorskiy» and «Tuapsinskiy» (Russia). Nat. Conserv. Res. 2020, 5, 66–85. [Google Scholar] [CrossRef]
- Khalimov, F. The ground beetles (Coleoptera, Carabidae) of the Karatepa and Chakilkalyan mountains (west part of Zarafshan Mountains Range, Uzbekistan). Biosyst. Divers. 2020, 28, 265–271. [Google Scholar] [CrossRef]
- Avtaeva, T.A.; Sukhodolskaya, R.A.; Brygadyrenko, V.V. Modeling the bioclimatic range of Pterostichus melanarius (Coleoptera, Carabidae) in conditions of global climate change. Biosyst. Divers. 2021, 29, 140–150. [Google Scholar] [CrossRef]
- Zamotajlov, A.S.; Serdyuk, V.Y.; Khomitskiy, E.E.; Belyi, A.I. New data on distribution and biology of some rare ground beetles (Coleoptera, Carabidae) in South Russia. Nat. Conserv. Res. 2019, 4, 81–90. [Google Scholar] [CrossRef]
- Kabak, I.I.; Liang, H.-B. An annotated list of the genus Chlaenius Bonelli, 1810 (Coleoptera: Carabidae) of Xinjiang Uygur Autonomous Region of China. Far East. Entomol. 2021, 429, 12–28. [Google Scholar] [CrossRef]
- Tomaszewska, W.; Egorov, L.V.; Ruchin, A.B.; Vlasov, D.V. First record of Clemmus troglodytes (Coleoptera: Coccinelloidea, Anamorphidae) for the fauna of Russia. Nat. Conserv. Res. 2018, 3, 103–105. [Google Scholar] [CrossRef]
- Gobbi, M.; Fontaneto, D. Biodiversity of ground beetles (Coleoptera: Carabidae) in different habitats of the Italian Po lowland. Agric. Ecosyst. Environ. 2008, 127, 273–276. [Google Scholar] [CrossRef]
- Sergeev, M.E. Species composition and biotopic distribution of leaf beetles (Coleoptera: Megalopodidae, Chrysomelidae) in the Sikhote-Alin State Nature Reserve (Russia). Nat. Conserv. Res. 2020, 5, 80–88. [Google Scholar] [CrossRef]
- Asbeck, T.; Großmann, J.; Paillet, Y.; Winiger, N.; Bauhus, J. The use of tree-related microhabitats as forest biodiversity indicators and to guide integrated forest management. Curr. For. Rep. 2021, 7, 59–68. [Google Scholar] [CrossRef]
- Allison, J.; Strom, B.; Sweeney, J.; Mayo, P. Trap deployment along linear transects perpendicular to forest edges: Impact on capture of longhorned beetles (Coleoptera: Cerambycidae). J. Pest Sci. 2019, 92, 299–308. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L.; Horn, S.; Kilgo, J.C.; Moorman, C.E. Spatial and temporal patterns of beetles associated with coarse woody debris in managed bottomland hardwood forests. For. Ecol. Manag. 2004, 199, 259–272. [Google Scholar] [CrossRef]
- Dunn, E.; Hough-Goldstein, J.; Hanks, L.M.; Millar, J.G.; D’Amico, V. Range of attraction of pheromone lures and dispersal behavior of cerambycid beetles. Ann. Entomol. Soc. Am. 2016, 109, 872–880. [Google Scholar] [CrossRef]
- Kozel, P.; Sebek, P.; Platek, M.; Benes, J.; Zapletal, M.; Dvorsky, M.; Lanta, V.; Dolezal, J.; Bace, R.; Zbuzek, B.; et al. Connectivity and succession of open structures as a key to sustaining light-demanding biodiversity in deciduous forests. J. Appl. Ecol. 2021, 58, 2951–2961. [Google Scholar] [CrossRef]
- Dodds, K. Effects of habitat type and trap placement on captures of bark (Coleoptera: Scolytidae) and longhorned (Coleoptera: Cerambycidae) beetles in semiochemical-baited traps. J. Econ. Entomol. 2011, 104, 879–888. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Dung beetles in continuous forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodivers. Conserv. 2002, 11, 1903–1918. [Google Scholar] [CrossRef]
- Schroeder, B.; Buddle, C.; Saint-Germain, M. Activity of flying beetles (Coleoptera) at two heights in canopy gaps and intact forests in a hardwood forest in Quebec. Can. Entomol. 2009, 141, 515–520. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Arthropod vertical stratification in temperate deciduous forests: Implications for conservation-oriented management. For. Ecol. Manag. 2011, 261, 1479–1489. [Google Scholar] [CrossRef]
- Kirstová, M.; Pyszko, P.; Šipoš, J.; Drozd, P.; Kočárek, P. Vertical distribution of earwigs (Dermaptera: Forficulidae) in a temperate lowland forest, based on sampling with a mobile aerial lift platform. Entomol. Sci. 2017, 20, 57–64. [Google Scholar] [CrossRef]
- Sheehan, T.N.; Ulyshen, M.D.; Horn, S.; Hoebeke, E.R. Vertical and horizontal distribution of bark and woodboring beetles by feeding guild: Is there an optimal trap location for detection? J. Pest Sci. 2019, 92, 327–341. [Google Scholar] [CrossRef]
- Weiss, M.; Didham, R.K.; Procházka, J.; Schlaghamerský, J.; Basset, Y.; Odegaard, F.; Tichechkin, A.; Schmidl, J.; Floren, A.; Curletti, G.; et al. Saproxylic beetles in tropical and temperate forests—A standardized comparison of vertical stratification patterns. For. Ecol. Manag. 2019, 444, 50–58. [Google Scholar] [CrossRef]
- Makarkin, V.N.; Ruchin, A.B. Materials on the Neuroptera and Raphidioptera fauna in Mordovia and adjacent regions of European Russia. Proc. Mordovia State Nat. Reserve 2020, 24, 161–181. [Google Scholar]
- Perry, K.I.; Sivakoff, F.S.; Wallin, K.F.; Wenzel, J.W.; Herms, D.A. Forest disturbance and arthropods: Small-scale canopy and understory disturbances alter movement of mobile arthropods. Ecosphere 2021, 12, e03771. [Google Scholar] [CrossRef]
- Urban-Mead, K.R.; Muñiz, P.; Gillung, J.; Espinoza, A.; Fordyce, R.; van Dyke, M.; McArt, S.H.; Danforth, B.N. Bees in the trees: Diverse spring fauna in temperate forest edge canopies. For. Ecol. Manag. 2021, 482, 118903. [Google Scholar] [CrossRef]
- Hanski, I. Habitat fragmentation and species richness. J. Biogeogr. 2015, 42, 989–993. [Google Scholar] [CrossRef]
- Ekström, A.L.; Bergmark, P.; Hekkala, A.M. Can multifunctional forest landscapes sustain a high diversity of saproxylic beetles? For. Ecol. Manag. 2021, 490, 119107. [Google Scholar] [CrossRef]
- Kärvemo, S.; Jönsson, M.; Hekkala, A.M.; Sjögren, J.; Strengbom, J. Multi-taxon conservation in northern forest hot-spots: The role of forest characteristics and spatial scales. Landsc. Ecol. 2021, 36, 989–1002. [Google Scholar] [CrossRef]
- Cicort-Lucaciu, A.Ș. Road-killed ground beetles prove the presence of Carabus hungaricus (Coleoptera: Carabidae) in North-Western Romania. Nat. Conserv. Res. 2020, 5, 134–138. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Makarkin, N.V. Neuroptera and Raphidioptera in the Mordovia State Nature Reserve. Nat. Conserv. Res. 2017, 2, 38–46. [Google Scholar] [CrossRef][Green Version]
- Ruchin, A.B.; Khapugin, A.A. Red Data Book Invertebrates in a Protected Area of European Russia. Acta Zool. Acad. Sci. Hung. 2019, 65, 349–370. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Alekseev, S.K.; Khapugin, A.A. Post-fire fauna of carabid beetles (Coleoptera, Carabidae) in forests of the Mordovia State Nature Reserve (Russia). Nat. Conserv. Res. 2019, 4 (Suppl. S1), 11–20. [Google Scholar] [CrossRef]
- Chugunov, G.G.; Vargot, E.V.; Khapugin, A.A. Materials for the flora of the Mordovia State Nature Reserve named after P.G. Smidovich (Report 3). Proc. Mordovia State Nat. Reserve 2017, 19, 220–228. [Google Scholar]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A.; Vikhrev, N.E.; Esin, M.N. The use of simple crown traps for the insects collection. Nat. Conserv. Res. 2020, 5, 87–108. [Google Scholar] [CrossRef]
- Bouchard, P.; Bousquet, Y. Additions and corrections to “Family-group names in Coleoptera (Insecta)”. ZooKeys 2020, 922, 65–139. [Google Scholar] [CrossRef]
- Löbl, I.; Smetana, A. (Eds.) Curculionoidea I. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2011; Volume 7, p. 373. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Curculionoidea II. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2013; Volume 8, p. 707. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Revised and Updated Version. Hy-Drophiloidea–Staphylinoidea. In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2015; Volume 2/1, p. 1702. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Revised and Updated Version. Scara-Baeoidea–Scirtoidea–Dascilloidea–Buprestoidea–Byrrhoidea. In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2016; Volume 3, p. 983. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Revised and Updated Version. Archostemata–Adephaga–Myxophaga. In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2017; Volume 1, p. 1443. [Google Scholar]
- Iwan, D.; Löbl, I. (Eds.) Revised and Updated Second Edition. Tenebrionoidea. In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 5, p. 945. [Google Scholar]
- Danilevsky, M. (Ed.) Updated and Revised Second Edition. Chrysomeloidea I (Vesperidae, Disteniidae, Cerambycidae). In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 6/1, p. 712. [Google Scholar]
- Robertson, J.; Ślipiński, A.; Moulton, M.; Shockley, F.W.; Giorgi, A.; Lord, N.P.; McKenna, D.D.; Tomaszewska, W.; Forrester, J.; Miller, K.B.; et al. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst. Entomol. 2015, 40, 745–778. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.H.C.; et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea. Monogr. Electrón. SEA 2017, 8, 1–729. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Elateroidea–Derodontoidea–Bostrichoidea–Lymexyloidea–Cleroidea–Cucujoidea. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2007; Volume 4, p. 935. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Chrysomeloidae. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2010; Volume 6, p. 924. [Google Scholar]
- Bousquet, Y. Litteratura Coleopterologica (1758–1900): A guide to selected books related to the taxonomy of Coleoptera with publication dates and notes. ZooKeys 2016, 583, 1–776. [Google Scholar] [CrossRef]
- Lachat, T.; Wermelinger, B.; Gossner, M.M.; Bussler, H.; Isacsson, G.; Müller, J. Saproxylic beetles as indicator species for dead-wood amount and temperature in European beech forests. Ecol. Indic. 2012, 23, 323–331. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. A red list of Italian saproxylic beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Entomol. 2015, 47, 53–126. [Google Scholar] [CrossRef]
- Gutowski, J.M.; Sućko, K.; Borowski, J.; Kubisz, D.; Mazur, M.A.; Melke, A.; Mokrzycki, T.; Plewa, R.; Żmihorski, M. Post-fire beetle succession in a biodiversity hotspot: Białowieża Primeval Forest. Forest Ecol. Manag. 2020, 461, 117893. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423; 623–659. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Chapman & Hall: London, UK, 1996; 179p. [Google Scholar]
- Allemand, R.; Aberlenc, H.P. Une methode efficace d’echantillonnage de l’entomofaune des frondaisons: Le piege attractif aerien. Bull. De La Soc. Entomol. Suisse 1991, 64, 293–305. [Google Scholar]
- Brustel, H. Coleopteres Saproxyliques et Valeur Biologique des Forets Françaises. Perspectives Pour la Conservation du Patrimoine Naturel; Les Dossiers Forestiers 13, Office National des Forets: Paris, France, 2004; 297p. [Google Scholar]
- Li, Y.; Meng, Q.; Silk, P.; Gao, W.; Mayo, P.; Sweeney, J. Effect of semiochemicals and trap height on catch of Neocerambyx raddei in Jilin province, China. Entomol. Exp. Appl. 2017, 164, 94–101. [Google Scholar] [CrossRef]
- Vance, C.C.; Kirby, K.R.; Malcolm, J.R.; Smith, S.M. Community composition of longhorned beetles (Coleoptera: Cerambycidae) in the canopy and understorey of sugar maple and white pine stands in south-central Ontario. Environ. Entomol. 2003, 32, 1066–1074. [Google Scholar] [CrossRef]
- Leksono, A.S.; Takada, K.; Koji, S.; Nakagoshi, N.; Anggraeni, T.; Nakamura, K. Vertical and seasonal distribution of flying beetles in a suburban temperate deciduous forest collected by water pan trap. Insect Sci. 2005, 12, 199–206. [Google Scholar] [CrossRef]
- Hirao, T.; Murakami, M.; Kashizaki, A. Importance of the understory stratum to entomofaunal diversity in a temperate deciduous forest. Ecol. Res. 2009, 24, 263–272. [Google Scholar] [CrossRef]
- Bouget, C.; Brin, A.; Brustel, H. Exploring the “last biotic frontier”: Are temperate forest canopies special for saproxylic beetles? For. Ecol. Manag. 2011, 261, 211–220. [Google Scholar] [CrossRef]
- Graham, E.E.; Poland, T.M.; McCullough, D.G.; Millar, J.G. A comparison of trap type and height for capturing cerambycid beetles (Coleoptera). J. Econ. Entomol. 2012, 105, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Vodka, Š.; Cizek, L. The effects of edge-interior and understorey-canopy gradients on the distribution of saproxylic beetles in a temperate lowland forest. For. Ecol. Manag. 2013, 304, 33–41. [Google Scholar] [CrossRef]
- Procházka, J.; Cizek, L.; Schlaghamerský, J. Vertical stratification of scolytine beetles in temperate forests. Insect Conserv. Divers. 2018, 11, 534–544. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. Vertical stratification of beetles in deciduous forest communities in the Centre of European Russia. Diversity 2021, 13, 508. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L. A comparison of the beetle (Coleoptera) fauna captured at two heights above the ground in a North American temperate deciduous forest. Am. Midl. Nat. 2007, 158, 260–278. [Google Scholar] [CrossRef]
- Kurochkin, A.S. Fauna and bionomy of sap beetles (Coleoptera, Nitidulidae) and kateretid beetles (Coleoptera, Kateretidae) of Krasnosamarskoe forestry farm (Samara Region, Russia). Vestn. Samara Univ. Nat. Sci. Ser. 2007, 8, 120–128. [Google Scholar]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Seasonal activity of Coleoptera attracted by fermental crown traps in forest ecosystems of Central Russia. Ecol. Quest. 2021, 32, 37–53. [Google Scholar] [CrossRef]
- Oleksa, A.; Chybicki, I.J.; Gawronski, R.; Svensson, G.P.; Burczyk, J. Isolation by distance in saproxylic beetles may increase with niche specialization. J. Insects Conserv. 2013, 17, 219–233. [Google Scholar] [CrossRef]
- Urban, P.; Schulze, W. Ein aktueller Nachweis des Marmorierten Rosenkäfers Pro-taetia marmorata (Fabricius, 1792) in der Senne (Nordrhein-Westfalen) (Mitteilungen zur Insektenfauna Westfalens XXII). Mitt. Der Arb. Westfälischer Entomol. 2017, 33, 15–19. [Google Scholar]
- Berglund, H.L.; Milberg, P. Sampling of flower-visiting insects: Poor correspondence between the catches of colour pan-trap and sweep netting. Europ. J. Entomol. 2019, 116, 425–431. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; MacGowan, I.; Makarkin, V.N.; Antropov, A.V.; Gornostaev, N.G.; Khapugin, A.A.; Dvořák, L.; Esin, M.N. Post-fire insect fauna explored by crown fermental traps in forests of the European Russia. Sci. Rep. 2021, 11, 21334. [Google Scholar] [CrossRef]
- Nasir, S.; Akram, W.; Ahmed, F. The population dynamics, ecological and seasonal activity of Paederus fuscipes Curtis (Staphylinidae; Coleoptera) in the Punjab, Pakistan. APCBEE Procedia 2012, 4, 36–41. [Google Scholar] [CrossRef][Green Version]
- Ruchin, A.B.; Egorov, L.V.; Semishin, G.B. Fauna of click beetles (Coleoptera: Elateridae) in the interfluve of Rivers Moksha and Sura, Republic of Mordovia, Russia. Biodiversitas 2018, 19, 1352–1365. [Google Scholar] [CrossRef]
- Batschynskaja, J.A.; Komisova, T.E.; Lykova, I.O. Peculiarities of the development and seasonal dynamics of the activity of epigean beetles of the Polyphaga suborder (Coleoptera) in fields of winter wheat in the conditions of Southern Ukraine. Biosyst. Divers. 2020, 28, 243–249. [Google Scholar] [CrossRef]
- Zemoglyadchuk, A.V.; Ruchin, A.B.; Egorov, L.V. An annotated checklist of the tumbling flower beetles (Coleoptera, Mordellidae) of the Republic of Mordovia, with a short review of the family in European Russia. Entomol. Rev. 2020, 100, 771–787. [Google Scholar] [CrossRef]
- Evangelista, J.; Rocha, M.V.C.; Monné, M.L.; Monné, M.A.; Frizzas, M.R. Diversity of Cerambycidae (Insecta: Coleoptera) in the Cerrado of Central Brazil using a new type of bait. Biota Neotrop. 2021, 21, e20201103. [Google Scholar] [CrossRef]
- Karolyi, F.; Gorb, S.N.; Krenn, H.W. Trapping pollen by the moist mouth: Structure and function of the mouthparts in the flower visiting Cetonia aurata (Scarabeidae, Coleoptera). Arthropod Plant Interact. 2009, 3, 1–8. [Google Scholar] [CrossRef]
- Fremlin, M. The rose chafer Cetonia aurata L. (Coleoptera: Scarabaeidae: Cetoniinae) in Essex: Distribution and some aspects of its ecology. Essex Nat. 2018, 35, 167–178. [Google Scholar]
- Egorov, L.V.; Ruchin, A.B.; Semishin, G.B. Some data concerning the Coleoptera fauna of the Mordovia State Nature Reserve. Information 9. Proc. Mordovia State Nat. Reserve 2020, 24, 61–150. [Google Scholar]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Usage of fermental traps for studying the species diversity of Coleoptera. Insects 2021, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Maican, S.; Munteanu, C. The diversity of urban coleopterans. In Species Monitoring in the Central Parks of Bucharest/Coord.: Marilena Onete; Ars Docendi: Bucureti, Romania, 2009; pp. 106–112. [Google Scholar]
- Ruchin, A.B.; Egorov, L.V. Fauna of longicorn beetles (Coleoptera: Cerambycidae) of Mordovia. Rus. Entomol. J. 2018, 27, 161–177. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family, Taxon | 2 m | 4 m | 6 m | 8 m | 10 m |
|---|---|---|---|---|---|
| Carabidae | |||||
| Harpalus signaticornis (Duftschmid, 1812) | 1 | ||||
| Stenolophus mixtus (Herbst, 1784) | 1 | ||||
| Hydrophilidae | |||||
| Hydrobius fuscipes (Linnaeus, 1758) | 1 | ||||
| Histeridae | |||||
| Carcinops pumilio (Erichson, 1834) | 1 | ||||
| Silphidae | |||||
| Necrodes littoralis (Linnaeus, 1758) | 1 | 1 | 1 | ||
| Oiceoptoma thoracicum (Linnaeus, 1758) | 1 | ||||
| Staphylinidae | |||||
| Staphylinidae sp. | 4 | 6 | 2 | ||
| Quedius dilatatus (Fabricius, 1787) | 1 | ||||
| Scarabaeidae | |||||
| Cetonia aurata (Linnaeus, 1758) | 11 | 8 | 3 | 1 | |
| Gnorimus variabilis (Linnaeus, 1758) | 2 | 1 | |||
| Phyllopertha horticola (Linnaeus, 1758) | 1 | 1 | 1 | ||
| Protaetia fieberi (Kraatz, 1880) | 6 | 6 | 2 | ||
| Protaetia marmorata (Fabricus, 1792) | 33 | 41 | 20 | 12 | 7 |
| Protaetia speciosissima (Scopoli, 1786) | 1 | 1 | |||
| Protaetia cuprea volhyniensis (Gory & Percheron, 1833) | 18 | 5 | 1 | 3 | |
| Scirtidae | |||||
| Contacyphon variabilis (Thunberg, 1787) | 1 | ||||
| Elateridae | |||||
| Agrypnus murinus (Linnaeus, 1758) | 3 | ||||
| Ampedus pomonae (Stephens, 1830) | 6 | ||||
| Dalopius marginatus (Linnaeus, 1758) | 1 | ||||
| Denticollis borealis (Paykull, 1800) | 1 | ||||
| Limonius minutus (Linnaeus, 1758) | 9 | 5 | |||
| Buprestidae | |||||
| Agrilus sp. | 1 | 2 | 1 | ||
| Anthaxia quadripunctata (Linnaeus, 1758) | 1 | ||||
| Coraebus elatus (Fabricius, 1787) | 1 | ||||
| Byrrhidae | |||||
| Lamprobyrrhulus nitidus (Schaller, 1783) | 1 | ||||
| Lycidae | |||||
| Lygistopterus sanguineus (Linnaeus, 1758) | 1 | ||||
| Cantharidae | |||||
| Cantharis fusca Linnaeus, 1758 | 1 | 1 | |||
| Cantharis rustica Fallén, 1807 | 1 | 2 | 2 | ||
| Rhagonycha fugax Mannerheim, 1843 | 1 | ||||
| Rhagonycha fulva (Scopoli, 1763) | 1 | 1 | |||
| Dermestidae | |||||
| Attagenus schaefferi (Herbst, 1792) | 1 | ||||
| Trogoderma glabrum (Herbst, 1783) | 3 | ||||
| Ptinidae | |||||
| Hadrobregmus pertinax (Linnaeus, 1758) | 1 | ||||
| Melyridae | |||||
| Dasytes niger (Linnaeus, 1761) | 1 | ||||
| Malachius bipustulatus (Linnaeus, 1758) | 7 | 4 | 2 | 5 | 4 |
| Erotylidae | |||||
| Dacne bipustulata (Thunberg, 1781) | 1 | ||||
| Nitidulidae | |||||
| Carpophilus dimidiatus (Fabricius, 1792) | 2 | ||||
| Carpophilus hemipterus (Linnaeus, 1758) | 2 | 1 | |||
| Cryptarcha strigata (Fabricius, 1787) | 76 | 83 | 26 | 34 | 9 |
| Cryptarcha undata (G.-A. Olivier, 1790) | 2 | 6 | 6 | 5 | 2 |
| Cychramus luteus (Fabricius, 1787) | 5 | ||||
| Cyllodes ater (Herbst, 1792) | 1 | ||||
| Epuraea sp. | 19 | 11 | 2 | 4 | 4 |
| Glischrochilus grandis (Tournier, 1872) | 5 | 1 | 1 | ||
| Glischrochilus hortensis (Geoffroy, 1785) | 2 | ||||
| Glischrochilus quadriguttatus (Fabricius, 1777) | 1 | 1 | |||
| Glischrochilus quadripunctatus (Linnaeus, 1758) | 1 | 1 | |||
| Meligethes sp. | 1 | 1 | |||
| Soronia grisea (Linnaeus, 1758) | 18 | 10 | 5 | 4 | 1 |
| Coccinellidae | |||||
| Calvia quatuordecimguttata (Linnaeus, 1758) | 2 | ||||
| Coccinella magnifica L. Redtenbacher, 1843 | 1 | 1 | |||
| Coccinella septempunctata Linnaeus, 1758 | 1 | ||||
| Hyperaspis campestris (Herbst, 1783) | 1 | ||||
| Propylea quatuordecimpunctata (Linnaeus, 1758) | 1 | ||||
| Melandryidae | |||||
| Osphya bipunctata (Fabricius, 1775) | 1 | ||||
| Wanachia triguttata (Gyllenhal, 1810) | 1 | ||||
| Zopheridae | |||||
| Bitoma crenata (Fabricius, 1775) | 1 | ||||
| Mordellidae | |||||
| Mordella sp. | 1 | 1 | |||
| Tenebrionidae | |||||
| Bolitophagus reticulatus (Linnaeus, 1767) | 1 | ||||
| Neomida haemorrhoidalis (Fabricius, 1787) | 1 | ||||
| Lagria hirta (Linnaeus, 1758) | 1 | ||||
| Oedemeridae | |||||
| Oedemera femorata (Scopoli, 1763) | 4 | ||||
| Anthicidae | |||||
| Omonadus floralis (Linnaeus, 1758) | 1 | ||||
| Scraptiidae | |||||
| Anaspis frontalis (Linnaeus, 1758) | 1 | ||||
| Cerambycidae | |||||
| Anastrangalia reyi (Heyden, 1889) | 1 | ||||
| Leptura quadrifasciata Linnaeus, 1758 | 20 | 8 | |||
| Leptura thoracica Creutzer, 1799 | 2 | 2 | |||
| Lepturalia nigripes (DeGeer, 1775) | 3 | 1 | |||
| Pachyta quadrimaculata (Linnaeus, 1758) | 1 | ||||
| Plagionotus arcuatus (Linnaeus, 1758) | 1 | ||||
| Purpuricenus kaehleri (Linnaeus, 1758) | 1 | ||||
| Stenocorus meridianus (Linnaeus, 1758) | 1 | ||||
| Strangalia attenuata (Linnaeus, 1758) | 2 | ||||
| Tetrops praeustus (Linnaeus, 1758) | 1 | ||||
| Orsodacnidae | |||||
| Orsodacne cerasi (Linnaeus, 1758) | 1 | 1 | 1 | ||
| Chrysomelidae | |||||
| Altica sp. | 1 | ||||
| Cassida vittata Villers, 1789 | 1 | ||||
| Chaetocnema hortensis (Geoffroy, 1785) | 1 | ||||
| Galerucella lineola (Fabricius, 1781) | 1 | 1 | 1 | ||
| Hypocassida subferruginea (Schrank, 1776) | 1 | 1 | 1 | ||
| Anthribidae | |||||
| Rhaphitropis marchica (Herbst, 1797) | 1 | ||||
| Curculionidae | |||||
| Anisandrus dispar (Fabricius, 1792) | 12 | 7 | 4 | 1 | |
| Larinus sturnus (Schaller, 1783) | 1 | ||||
| Phyllobius argentatus (Linnaeus, 1758) | 1 | 1 | |||
| Phyllobius pyri (Linnaeus, 1758) | 1 | 3 | 1 | 1 | |
| Xyleborinus saxesenii (Ratzeburg, 1837) | 1 | 1 | |||
| Total of individuals | 302 | 229 | 83 | 90 | 41 |
| Number of taxon (excluding unidentified ones) | 53 | 29 | 22 | 22 | 16 |
| Number of saproxylic taxon (excluding unidentified ones) | 30 | 20 | 13 | 12 | 9 |
| Number of species of saproxylic beetles (% of the total number of taxon) | 56.6 | 69.0 | 59.1 | 54.5 | 56.3 |
| Number of anthophilic taxon (excluding unidentified ones) | 25 | 13 | 13 | 12 | 4 |
| Number of taxon anthophilic beetles (% of the total number of taxon) | 47.2 | 44.8 | 59.1 | 54.5 | 25.0 |
| Shannon index | 2.91 | 2.23 | 2.25 | 2.23 | 2.39 |
| Simpson index | 0.11 | 0.21 | 0.18 | 0.20 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruchin, A.B.; Egorov, L.V. Vertical Stratification and Seasonal Dynamics of Coleoptera in Open Biotopes of Forest Ecosystems (Centre of European Russia). Forests 2022, 13, 1014. https://doi.org/10.3390/f13071014
Ruchin AB, Egorov LV. Vertical Stratification and Seasonal Dynamics of Coleoptera in Open Biotopes of Forest Ecosystems (Centre of European Russia). Forests. 2022; 13(7):1014. https://doi.org/10.3390/f13071014
Chicago/Turabian StyleRuchin, Alexander B., and Leonid V. Egorov. 2022. "Vertical Stratification and Seasonal Dynamics of Coleoptera in Open Biotopes of Forest Ecosystems (Centre of European Russia)" Forests 13, no. 7: 1014. https://doi.org/10.3390/f13071014
APA StyleRuchin, A. B., & Egorov, L. V. (2022). Vertical Stratification and Seasonal Dynamics of Coleoptera in Open Biotopes of Forest Ecosystems (Centre of European Russia). Forests, 13(7), 1014. https://doi.org/10.3390/f13071014