
Mycophagy of White-Tailed Deer (Odocoileus virginianus Zimmermann) in the Boreal Forest



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. DNA Extraction and Sequencing
2.4. Bioinformatic Analyses
2.5. Statistical Analyses
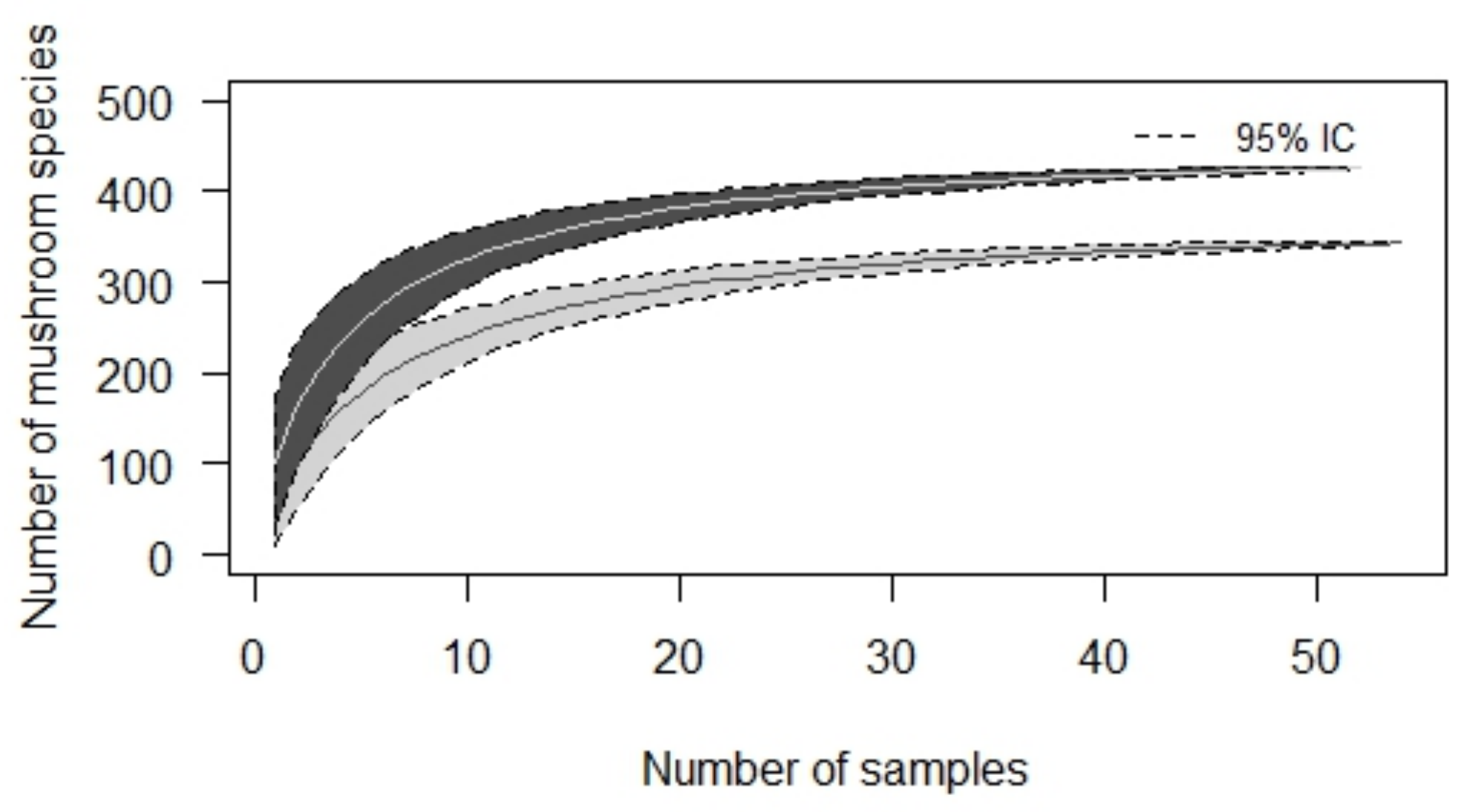
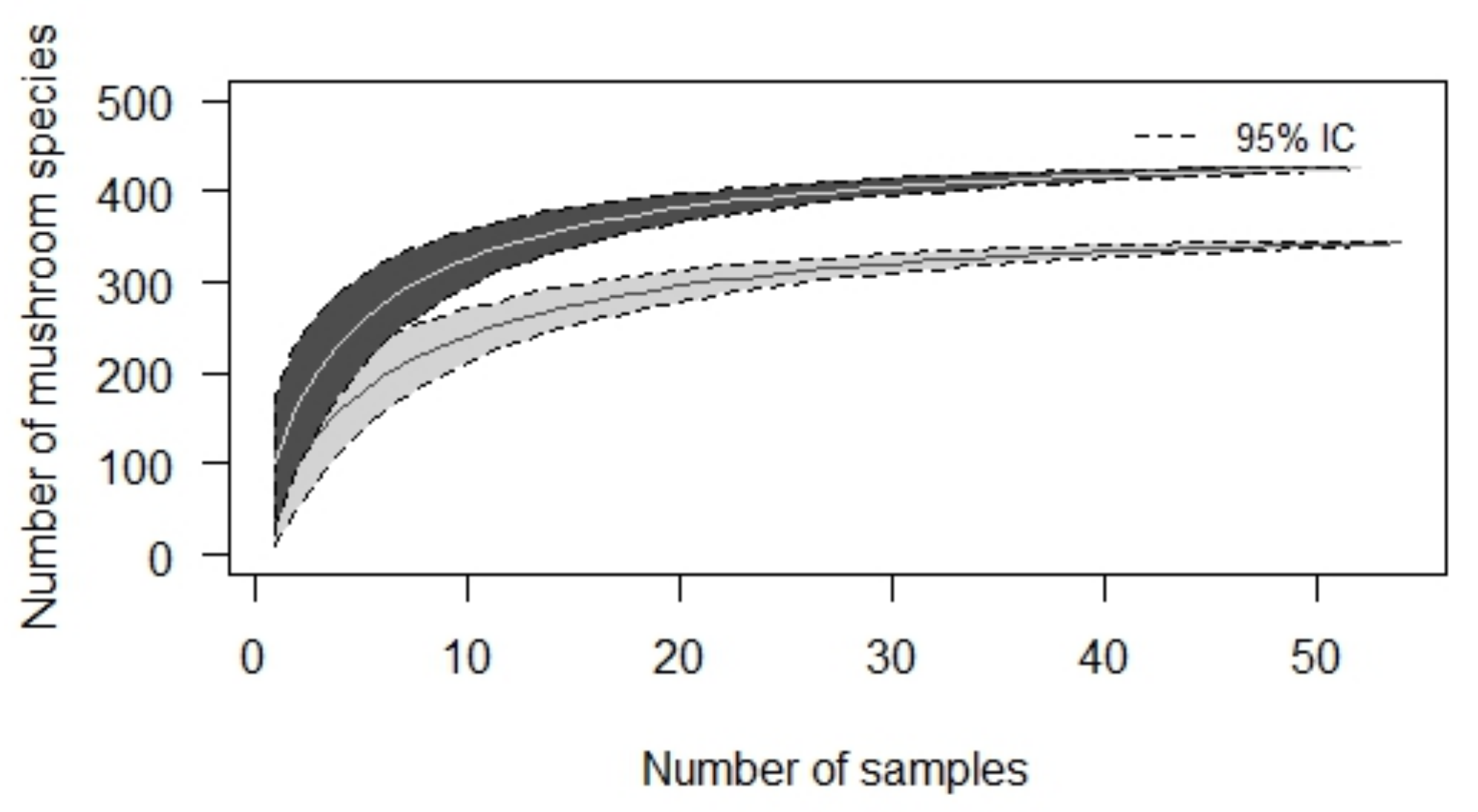
3. Results
3.1. Consumed Mushroom Species
3.2. Effects of Intrinsic and Extrinsic Variables on the Diversity of Mushrooms in Deer Diet
4. Discussion
4.1. Consumed Mushroom Species
4.2. Effects of Intrinsic and Extrinsic Variables on the Diversity of Mushrooms in Deer Diet
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Species | Groups | Crude Protein (% of Dry Weight) | References |
|---|---|---|---|
| Tuber Hafizi | Hypogeous | 19.2 | [79] |
| Tuber nivea | Hypogeous | 16.0 | [79] |
| Tuber Boudieri | Hypogeous | 15.8 | [79] |
| Tuber claveriy | Hypogeous | 15.0 | [79] |
| Ophiocordyceps sinensis | Hypogeous | 38.6 | [80] |
| Cordyceps militaris | Hypogeous | 29.7 | [80] |
| Armillaria mellea | Armillarioids | 21.1 | [81] |
| Lactarius volemus | Lactarius | 25.2 | [81] |
| Agaricus arvensis | Agaricus | 56.3 | [71] |
| Armillariella mellea | Armillarioids | 22.3 | [71] |
| Boletus edulis | Boletoids | 33.1 | [71] |
| Craterellus cornucopioides | Chanterelloids | 22.3 | [71] |
| Cantharellus cibarius | Chanterelloids | 18.7 | [71] |
| Lactarius deliciosus | Lactarius | 29.8 | [71] |
| Agaricus campestris | Agaricus | 18.6 | [72] |
| Armillaria mellea | Armillarioids | 16.4 | [72] |
| Suillus mediterraneensis | Boletoids | 24.3 | [72] |
| Boletus reticulatus | Boletoids | 22.6 | [72] |
| Boletus edulis | Boletoids | 21.1 | [72] |
| Boletus erythropus | Boletoids | 20.9 | [72] |
| Boletus armeniacus | Boletoids | 18.3 | [72] |
| Boletus aereus | Boletoids | 17.9 | [72] |
| Suillus variegatus | Boletoids | 17.6 | [72] |
| Suillus granulatus | Boletoids | 16.5 | [72] |
| Cantharellus cibarius | Chanterelloids | 35.8 | [72] |
| Lactarius salmonicolor | Lactarius | 37.3 | [72] |
| Lactarius deliciosus | Lactarius | 20.2 | [72] |
| Russula delica | Russula | 50.6 | [72] |
| Russula olivacea | Russula | 16.8 | [72] |
| Russula cyanoxantha | Russula | 16.8 | [72] |
| Boletus armeniacus | Boletoids | 18.3 | [73] |
| Suillus variegatus | Boletoids | 17.6 | [73] |
| Boletus impolitus | Boletoids | 16.0 | [73] |
| Cortinarius praestans | Cortinarius | 14.6 | [73] |
| Tuber Zubaidi | Hypogeous | 27.2 | [74] |
| Tuber Gibaah | Hypogeous | 25.0 | [74] |
| TuberKolehissi | Hypogeous | 19.6 | [74] |
| Genus and Species | Deer Species | Methods | References |
|---|---|---|---|
| Daedalia confragosa | Odocoileus virginianus | Behavioral observation | [14] |
| Panus stipticus | Odocoileus virginianus | Behavioral observation | [14] |
| Polypores elegans | Odocoileus virginianus | Behavioral observation | [14] |
| Scleroderma vulgare | Odocoileus virginianus | Behavioral observation | [14] |
| Stereum rameale | Odocoileus virginianus | Behavioral observation | [14] |
| Polypores arcularius | Odocoileus virginianus | Rumen samples | [18] |
| Usnea sp. | Odocoileus virginianus | Rumen samples | [18] |
| Boletoids subaureus | Odocoileus hermianus | Stomach contents | [10] |
| Clitocybe gigantea | Odocoileus hermianus | Stomach contents | [10] |
| Morchella esculenta | Odocoileus hermianus | Stomach contents | [10] |
| Russula atropurpurea | Odocoileus hermianus | Stomach contents | [10] |
| Usnea barbata | Odocoileus hermianus | Stomach contents | [10] |
| Amanita muscaria | Odocoileus hermianus | Behavioral observation | [17] |
| Boletoids oriantiachus | Odocoileus hermianus | Behavioral observation | [17] |
| Cortinarius sp. | Odocoileus hermianus | Behavioral observation | [17] |
| Russula emetica | Odocoileus hermianus | Behavioral observation | [17] |
| Agaricus sp. | Odocoileus hermianus | Rumen samples | [9] |
| Rhizopogon evadens | Deer in North America | Fecal inoculant | [20] |
| Rhizopogon fuscorubens | Deer in North America | Fecal inoculant | [20] |
| Rhizopogon occidentalis | Deer in North America | Fecal inoculant | [20] |
| Rhizopogon salebrosus | Deer in North America | Fecal inoculant | [20] |
| Suillus brevipes | Deer in North America | Fecal inoculant | [20] |
| Suillus tomentosus | Deer in North America | Fecal inoculant | [20] |
| Suillus umbonatus | Deer in North America | Fecal inoculant | [20] |
| Thelephora americana | Deer in North America | Fecal inoculant | [20] |
| Thelephoraceae sp. | Deer in North America | Fecal inoculant | [20] |
| Tomentella sublilicina | Deer in North America | Fecal inoculant | [20] |
| Elaphomyces anthracinus | Deer in North America | Behavioral observation | [15] |
| Elaphomyces granulatus | Deer in North America | Behavioral observation | [15] |
| Rhizopogon roseolus | Deer in North America | Behavioral observation | [15] |
| Armillarias ventricosa | Deer in North America | - | [16] |
| Clavaria sp. | Deer in North America | - | [16] |
| Lactarius sp. | Deer in North America | - | [16] |
| Suillus granulatus | Deer in North America | - | [16] |
| Amanita sp. | Capreolus capreolus | Organs and feces | [19] |
| Boletuss sp. | Capreolus capreolus | Organs and feces | [19] |
| Calvatia sp. | Capreolus capreolus | Organs and feces | [19] |
| Coprinus sp. | Capreolus capreolus | Organs and feces | [19] |
| Cortinarius sp. | Capreolus capreolus | Organs and feces | [19] |
| Elaphomyces sp. | Capreolus capreolus | Organs and feces | [19] |
| Elaphomyces virgatosporus | Capreolus capreolus | Organs and feces | [19] |
| Entoloma sp. | Capreolus capreolus | Organs and feces | [19] |
| Hypholoma sp. | Capreolus capreolus | Organs and feces | [19] |
| Inocybe sp. | Capreolus capreolus | Organs and feces | [19] |
| Lycoperdon sp. | Capreolus capreolus | Organs and feces | [19] |
| Macrolepiota sp. | Capreolus capreolus | Organs and feces | [19] |
| Pluteus sp. | Capreolus capreolus | Organs and feces | [19] |
| Russula sp. | Capreolus capreolus | Organs and feces | [19] |
| Suillus sp. | Capreolus capreolus | Organs and feces | [19] |
| Xerocomus sp. | Capreolus capreolus | Organs and feces | [19] |
| Models | K | AICc | Delta_AICc | AICcWt | LL |
|---|---|---|---|---|---|
| Age + Condition + Year | 5 | 257.84 | 0 | 0.44 | −122.56 |
| Age + Year | 4 | 258.82 | 0.98 | 0.27 | −124.54 |
| Null | 2 | 260.19 | 2.34 | 0.14 | −127.85 |
| Year | 3 | 261.62 | 3.78 | 0.07 | −127.31 |
| Age + Month + Year | 6 | 262.84 | 5 | 0.04 | −123.42 |
| Condition + Year | 4 | 263.94 | 6.1 | 0.02 | −127.1 |
| Stand_Age + Year | 4 | 264.16 | 6.32 | 0.02 | −127.21 |
| Month + Year | 5 | 265.56 | 7.72 | 0.01 | −126.42 |
| Stand_type + Year | 6 | 266.24 | 8.4 | 0.01 | −125.12 |
| Condition + Month + Year | 6 | 268.32 | 10.47 | 0 | −126.16 |
| Stand_type + Stand_Age + Year | 7 | 269.77 | 11.93 | 0 | −125.09 |
| Condition + Stand_type + Stand_Age + Year | 8 | 273.30 | 15.46 | 0 | −124.86 |
| Month + Stand_type + Stand_Age + Year | 9 | 277.62 | 19.78 | 0 | −124.81 |
| Condition + Stand_type + Stand_Age + Month + Year | 10 | 282.02 | 24.18 | 0 | −124.54 |
| Models | K | AICc | Delta_AICc | AICcWt | LL |
|---|---|---|---|---|---|
| Age + Year | 4 | 643.59 | 0 | 0.49 | −317.47 |
| Age + Condition + Year | 5 | 645.41 | 1.82 | 0.20 | −317.22 |
| Year | 3 | 646.37 | 2.77 | 0.12 | −319.99 |
| Age + Month + Year | 6 | 647.75 | 4.15 | 0.06 | −317.17 |
| Condition + Year | 4 | 647.94 | 4.35 | 0.06 | −319.65 |
| Stand_Age + Year | 4 | 648.50 | 4.91 | 0.04 | −319.93 |
| Month + Year | 5 | 650.73 | 7.13 | 0.01 | −319.87 |
| Condition + Month + Year | 6 | 652.60 | 9.01 | 0.01 | −319.6 |
| Stand_type + Year | 8 | 652.93 | 9.34 | 0 | −317.23 |
| Stand_type + Stand_Age + Year | 9 | 654.51 | 10.92 | 0 | −316.68 |
| Condition + Stand_type + Stand_Age + Year | 10 | 656.97 | 13.38 | 0 | −316.52 |
| Month + Stand_type + Stand_Age + Year | 11 | 659.74 | 16.15 | 0 | −316.47 |
| Condition + Stand_type + Stand_Age + Month + Year | 12 | 662.56 | 18.96 | 0 | −316.39 |
| Null | 2 | 663.25 | 19.65 | 0 | −329.53 |
| Models | K | AICc | Delta_AICc | AICcWt | LL |
|---|---|---|---|---|---|
| Year + Lactation | 4 | 454.95 | 0 | 0.52 | −223 |
| Stand_Age + Year + Lactation | 5 | 456.51 | 1.56 | 0.24 | −222.52 |
| Condition + Year + Lactation | 5 | 457.43 | 2.48 | 0.15 | −222.98 |
| Month + Year + Lactation, | 6 | 459.34 | 4.39 | 0.06 | −222.62 |
| Condition + Month + Year + Lactation | 7 | 461.88 | 6.93 | 0.02 | −222.5 |
| Stand_type + Year + Lactation | 8 | 463.26 | 8.31 | 0.01 | −221.74 |
| Stand_type + Stand_Age + Year + Lactation | 9 | 465.62 | 10.67 | 0 | −221.38 |
| Condition + Stand_type + Stand_Age + Year + Lactation | 10 | 468.67 | 13.72 | 0 | −221.28 |
| Month + Stand_type + Stand_Age + Year + Lactation | 11 | 471.81 | 16.86 | 0 | −221.13 |
| Condition + Stand_type + Stand_Age + Month + Year + Lactation | 12 | 474.97 | 20.02 | 0 | −220.89 |
| Year | 3 | 551.51 | 96.56 | 0 | −272.53 |
| Condition + Year | 4 | 552.06 | 97.11 | 0 | −271.64 |
| Stand_Age + Year | 4 | 552.97 | 98.02 | 0 | −272.1 |
| Condition + Month + Year | 6 | 555.54 | 100.59 | 0 | −270.93 |
| Month + Year | 5 | 555.93 | 100.98 | 0 | −272.37 |
| Stand_type + Year | 7 | 559.61 | 104.66 | 0 | −271.66 |
| Stand_type + Stand_Age + Year | 8 | 560.63 | 105.68 | 0 | −270.82 |
| Condition + Stand_type + Stand_Age + Year | 9 | 561.22 | 106.27 | 0 | −269.7 |
| Condition + Stand_type + Stand_Age + Month + Year | 11 | 565.81 | 110.86 | 0 | −268.97 |
| Month + Stand_type + Stand_Age + Year | 10 | 566.1 | 111.15 | 0 | −270.66 |
| Null | 2 | 568.04 | 113.09 | 0 | −281.91 |
| Species and Genus | Reference |
|---|---|
| Agaricus arvensis | [40] |
| Albatrellus confluens | [40] |
| Aleuria aurantia | [40] |
| Amanita muscaria | [40] |
| Auricularia auricula-judae | [40] |
| Cantharellus cibarius | [40] |
| Clavariadelphus sp. | [40] |
| Clavulinopsis fusiformis | [40] |
| Coprinus atramentarius | [40] |
| Coprinus comatus | [40] |
| Cortinarius alboviolaceus | [40] |
| Cortinarius armillatus | [40] |
| Cortinarius violaceus | [40] |
| Craterelle tubaeformis | [40] |
| Geastrum sp. | [40] |
| Gyromitra sp. | [40] |
| Helvella crispa | [40] |
| Hydnum repandum | [40] |
| Hygrocybe coccinea | [40] |
| Hygrophoropsis aurantiaca | [40] |
| Hypomyces lactifluorum | [40] |
| Hypsizygus ulmarius | [40] |
| Inocybe sp. | [40] |
| Laccaria laccata | [40] |
| Lactarius deterrimus | [40] |
| Lepiota sp. | [40] |
| Lycoperdon perlatum | [40] |
| Lycoperdon pyriforme | [40] |
| Marasmius scorodonius | [40] |
| Mutinus caninus | [40] |
| Neolecta irregularis | [40] |
| Paxillus involutus | [40] |
| Paxillus sp. | [40] |
| Pleurocybella porrigens | [40] |
| Pleurotus ostreatus | [40] |
| Russula decolorans | [40] |
| Russula emetica | [40] |
| Sarcodon imbricatus | [40] |
| Suillus cavipes | [40] |
| Suillus grevillei | [40] |
| Tremella foliacea | [40] |
| Tremellodon gelatinosum | [40] |
| Tremiscus helvelloides | [40] |
| Tricholoma sp. | [40] |
| Xerocomus badius | [40] |
| Alectoria sarmentosa | [66] |
| Amanitopsis sp. | [66] |
| Arthonia swartziana | [66] |
| Biatora campestris | [66] |
| Biatora decipiens | [66] |
| Biatora sanguineo-atra | [66] |
| Biatora uliginosa | [66] |
| Boletinus sp. | [66] |
| Boletus sp. | [66] |
| Bryopogon jubata | [66] |
| Buellia parasema | [66] |
| Cetraria islandica | [66] |
| Chone unfundibulformis | [66] |
| Cladonia gracilis | [66] |
| Cladonia turgida | [66] |
| Clavaria aurea | [66] |
| Clitocybe laccata | [66] |
| Collema flaccidum | [66] |
| Collybia sp. | [66] |
| Exoascus sp. | [66] |
| Fistulina sp. | [66] |
| Fomes fomentarius | [66] |
| Giromitra sp. | [66] |
| Graphis scripta | [66] |
| Heterothecium grossum | [66] |
| Heterothecium pezizoideum | [66] |
| Heterothecium sanguinarium | [66] |
| Hirneola sp. | [66] |
| Lactarius piperatus | [66] |
| Lecanora prinigna | [66] |
| Lecanora surfusca | [66] |
| Lecidea enteroleuca | [66] |
| Lenzites betulina | [66] |
| Lycoperdon sp. | [66] |
| Morchella deliciosa | [66] |
| Nephroma levigatum | [66] |
| Pannaria brunnea | [66] |
| Pannaria lepidiota | [66] |
| Pannaria macounii | [66] |
| Pannaria mycrophylla | [66] |
| Parmelia saxatilis | [66] |
| Peltigera aphtosa | [66] |
| Peltigera canina | [66] |
| Pertusaria multipunctata | [66] |
| Peziza odorata | [66] |
| Placodium aurantiacum | [66] |
| Placodium elegans | [66] |
| Pleurotus sp. | [66] |
| Pluteus cervinus | [66] |
| Polyporus versicolor | [66] |
| Ramalina calicaris | [66] |
| Ramalina polymorpha | [66] |
| Sphaerophorus globiferous | [66] |
| Stereocaulon pileatum | [66] |
| Stereocaulon tomentosum | [66] |
| Sticta pulmonaria | [66] |
| Sticta scorbiculata | [66] |
| Theloshistes polycarpus | [66] |
| Umbilicaria hyperborea | [66] |
| Usnea barbata | [66] |
References
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Berteaux, D.; Crête, M.; Huot, J.; Maltais, J.; Ouellet, J.-P. Food choice by white-tailed deer in relation to protein and energy content of the diet: A field experiment. Oecologia 1998, 115, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Miles, P.G. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Launchbaugh, K.L.; Urness, P.J. Mushroom consumption (mycophagy) by north american cervids. Great Basin Nat. 1992, 52, 321–327. [Google Scholar]
- Hanley, T.A.; Robbins, C.T.; Hagerman, A.E.; McArthur, C. Predicting digestible dry matter in tannin-containing forages consumed by ruminants. Ecology 1992, 73, 537–541. [Google Scholar] [CrossRef]
- Beck, J.L.; Peek, J.M. Great basin summer range forage quality: Do plant nutrient meet elk requirements? West. N. Am. Nat. 2005, 65, 516–527. [Google Scholar]
- Windels, S.K.; Hewitt, D.G. Effects of plant secondary compounds on nutritional carrying capacity estimates of a browsing ungulate. Rangel Ecol. Manag. 2011, 64, 264–275. [Google Scholar] [CrossRef]
- Golley, F.B. Energy values of ecological materials. Ecology 1961, 42, 581–584. [Google Scholar] [CrossRef]
- Lovaas, A.L. Mule deer food habits and range use, little belt mountains, Montana. J. Wildl. Manage. 1958, 22, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Cowan, I.M. The ecological relationships of the food of the columbian black-tailed deer, Odocoileus hemionus columbianus (Richardson), in the coast forest region of southern Vancouver Island, British Columbia. Ecol. Monogr. 1945, 15, 110–139. [Google Scholar] [CrossRef]
- Cook, J.G.; Cook, R.C.; Davis, R.W.; Irwin, L.L. Nutritional ecology of elk during summer and autumn un the Pacific northwest. Wildl. Monogr. 2016, 195, 1–81. [Google Scholar] [CrossRef]
- Denryter, K.A.; Cook, R.C.; Cook, J.G.; Parker, K.L. Straight from the caribou’s (Rangifer tarandus) mouth: Detailes observations of tame caribou reveal insights into summer-autumn diets. Can. J. Zool. 2017, 95, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Maser, C.; Trappe, J.M.; Claridge, A.W. Trees, Truffles, and Beasts: How Forests Function; Rutgers University Press: New Brunswick, NJ, USA, 2008. [Google Scholar]
- Atwood, E.L. White-tailed deer foods of the United States. J. Wildl. Manag. 1941, 5, 314–332. [Google Scholar] [CrossRef]
- Gillis, W.T. Subterranean elaphomyces and rhizopogon in the Michigan jack-pine region. Mycologia 1959, 51, 364–367. [Google Scholar] [CrossRef]
- Miller, H.A.; Halls, L.K. Fleshy Fungi Commonly Eaten by Southern Wildlife; Forest Service Research Paper SO-49; U.S. Department of Agriculture: New Orleans, LA, USA, 1969.
- Hungerford, C.R. Response of kaibab mule deer to management of summer range. J. Wildl. Manag. 1970, 34, 852–862. [Google Scholar] [CrossRef]
- Skinner, W.R.; Telfer, E.S. Spring, summer, and fall foods of deer in New Brunswick. J. Wildl. Manag. 1974, 38, 210–214. [Google Scholar] [CrossRef]
- Pokorny, B.; Al Sayegh-Petkovsek, S.; Ribaric-Lasnik, C.; Vrtacnik, J.; Doganoc, D.Z.; Adamic, M. Fungi ingestion as an important factor influencing heavy metal intake in roe deer: Evidence from faeces. Sci. Total. Environ. 2004, 324, 223–234. [Google Scholar] [CrossRef]
- Ashkannejhad, S.; Horton, T.R. Ectomycorrhizal ecology under primary succession on coastal sand dunes: Interactions involving Pinus contorta, suilloid fungi and deer. New Phytol. 2006, 169, 345–354. [Google Scholar] [CrossRef]
- Carroll, G. Fungal endophytes in stems and leaves: From latent pathogen to mutualistic symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Provenza, F.; Villalba, J.; Dziba, L.; Atwood, S.B.; Banner, R. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rumin. Res. 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Demment, M.W.; Van Soest, P.J. A nutritional explanation for body-size patterns of ruminant and nonruminant herbivores. Am. Nat. 1985, 125, 641–672. [Google Scholar] [CrossRef]
- Holter, J.B.; Urban, W.E.; Hayes, H.H. Nutrition of northern white-tailed deer throughout the year. J. Anim. Sci. 1977, 45, 365. [Google Scholar] [CrossRef] [PubMed]
- Simard, A.; Huot, J.; De Bellefeuille, S.; Côté, S.D. Influences of habitat composition, plant phenology, and population density on autumn indices of body condition in a northern white-tailed deer population. Wildl. Monogr. 2014, 187, 1–28. [Google Scholar] [CrossRef]
- Hudson, R.J.; White, R.G. Bioenergetics of Wild Herbivores; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
- Twieg, B.D.; Durall, D.M.; Simard, S.W. Ectomycorrhizal fungal succession in mixed temperate forests. New Phytol. 2007, 176, 437–447. [Google Scholar] [CrossRef]
- McNeil, R. Le Grand Livre Des Champignons du Québec et de l’Est du Canada; Éditions Michel Quintin: Waterloo, ON, Canada, 2006. [Google Scholar]
- Kelty, M.J.; Larson, B.C.; Oliver, C.D.; Smith, D.M. The Ecology and Silviculture of Mixed-Species Forests: A Festschrift for David M. Smith; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992. [Google Scholar]
- Wang, S.; Chen, H.Y.H. Diversity of northern plantations peaks at intermediate management intensity. For. Ecol. Manag. 2010, 259, 360–366. [Google Scholar] [CrossRef]
- Nash, T.H. Lichen Biology; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Parker, K.L.; Barboza, P.S.; Stephenson, T.R. Protein conservation in female caribou (Rangifer tarandus): Effects of decreasing diet quality during winter. J. Mammal. 2005, 86, 610–622. [Google Scholar] [CrossRef] [Green Version]
- Lefort, S.; Tremblay, J.-P.; Fournier, F.; Potvin, F.; Huot, J. Importance of balsam fir as winter forage for white-tailed deer at the northeastern limit of their distribution range. Ecoscience 2007, 14, 109–116. [Google Scholar] [CrossRef]
- Phillips, R.; Foy, N.; Kibby, G. Les Champignons du Québec: Guide D’identification; Broquet: Saint-Constant, QC, Canada, 2002. [Google Scholar]
- McNeil, R. Champignons Communs du Québec et de l’Est du Canada; Éditions Michel Quintin: Waterloo, ON, Canada, 2007. [Google Scholar]
- Thomas, A.C.; Jarman, S.N.; Haman, K.H.; Trites, A.W.; Deagle, B.E. Improving accuracy of DNA diet estimates using food tissue control materials and an evaluation of proxies for digestion bias. Mol. Ecol. 2014, 23, 3706–3718. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T. Its primer with enhanced specificity for basidiomycetes-Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Gales, N.J.; Cheal, A.J. Estimating diet composition of the Australian sea lion (Neophoa cinerea) from scat analysis: An unreliable technique. Wildl. Res. 1992, 19, 447–455. [Google Scholar] [CrossRef]
- Côté, S.D.; Dussault, C.; Huot, J.; Potvin, F.; Tremblay, J.-P.; Viera, V. High herbivore density and boreal forest ecology: White-tailed deer on Anticosti Island. In Lessons from the Islands: Introduced Species and What They Tell Us about How Ecosystems Work; Gaston, A.J., Golumbia, T.E., Martin, J.-L., Sharpe, S.T., Eds.; Canadian Wildlife Service: Ottawa, ON, Canada, 2008. [Google Scholar]
- MFFP. Zones de Végétation et Domaines Bioclimatiques du Québec; Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2015. [Google Scholar]
- Potvin, F.; Beaupré, P.; Gingras, A.; Pothier, D. Le Cerf et Les Sapinières de L’île D’anticosti; Société de la Faune et des Parcs du Québec: Québec, QC, Canada, 2000. [Google Scholar]
- Hewison, A.J.M.; Angibault, J.M.; Bideau, E.; Vincent, J.P.; Bonin, J.; Sempr, A. Annual variation in body composition of roe deer (Capreolus capreolus) in moderate environmental conditions. Can. J. Zool. 1996, 74, 245–253. [Google Scholar] [CrossRef]
- Taillon, J.; Brodeur, V.; Festa-Bianchet, M.; Côté, S.D. Variation in body condition of migratory caribou at calving and weaning: Which measures should we use? Ecoscience 2011, 18, 295–303. [Google Scholar] [CrossRef]
- Cook, R.C.; Cook, J.G.; Murray, D.L.; Zager, P.; Johnson, B.K.; Gratson, M.W. Nutritional condition models for elk: Which are the most sensitive, accurate, and precise? J. Wildl. Manag. 2001, 65, 988–997. [Google Scholar] [CrossRef]
- Chanmcleod, A.C.A.; White, R.G.; Russel, D. Body-mass and composition indexes for female barren-ground caribou. J. Wildl. Manag. 1995, 59, 278–291. [Google Scholar] [CrossRef]
- Cangelosi, R.; Goriely, A. Component retention in principal component analysis with application to cDNA microarray data. Biol. Direct 2007, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mautz, W.W.; Petrides, G.A. Food passage rate in the white-tailed deer. J. Wild. Manag. 1971, 35, 723–731. [Google Scholar] [CrossRef]
- Massé, A.; Côté, S.D. Spatiotemporal variations in resources affect activity and movement patterns of white-tailed deer (Odocoileus virginianus) at high density. Can. J. Zool. 2013, 91, 252–263. [Google Scholar] [CrossRef] [Green Version]
- MFFP. Couche Géographique des Peuplements Écoforestiers sur L’île D’anticosti; Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2011. [Google Scholar]
- Bérubé, J.A.; Gagné, P.N.; Ponchart, J.P.; Tremblay, É.D.; Bilodeau, G.J. Detection of Diplodia corticola spores in Ontario and Québec based on High Throughput Sequencing (HTS) methods. Can. J. Plant Pathol. 2018, 40, 378–386. [Google Scholar] [CrossRef]
- Huse, S.M.; Welch, D.M.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ. Microbiol. 2010, 12, 1889–1898. [Google Scholar] [CrossRef] [Green Version]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Christopherson, V.; Tremblay, J.-P.; Gagné, P.N.; Bérubé, J.; St-Laurent, M.H. Meeting caribou in the alpine: Do moose compete with caribou for food? Glob. Ecol. Conserv. 2019, 20, e00733. [Google Scholar] [CrossRef]
- Masella, A.; Bartram, A.; Truszkowski, J.; Brown, D.; Neufeld, J. PANDAseq: Paired-end assembler for Illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagné, P.; Bérubé, J.A. Illumicut, a C++ Program Specially Designed to Efficiently Detect and Remove forward and Reverse Sequencing Primers in Paired-End Reconstructed Sequences. Available online: https://github.com/Patg13/Illumicut (accessed on 22 November 2017).
- Gagné, P.; Bérubé, J.A. HomopRemover, a Program Designed to Efficiently Remove Sequences Containing very Long Homopolymers. Available online: https://github.com/Patg13/HomopRemover (accessed on 22 November 2017).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Valentini, A.; Pompanon, F.; Taberlet, P. DNA barcoding for ecologists. Trends Ecol. Evol. 2009, 24, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, R.H.; Bok, G.; Ryberg, M.; Kristiansson, E.; Hallenberg, N. A software pipeline for processing and identification of fungal ITS sequences. Source Code Biol. Med. 2009, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Meyers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T. BLAST plus: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ait Baamrane, M.A.; Shehzad, W.; Ouhammou, A.; Abbad, A.; Naimi, M.; Coissac, E.; Taberlet, P.; Znari, M. Assessment of the food habits of the Moroccan dorcas gazelle in M’Sabih Talaa, west central Morocco, using the trnL approach. PLoS ONE 2012, 7, e35643. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Schmitt, J. Monographie de l’Ile d’Anticosti (Golfe Saint-Laurent); Hermann: Paris, France, 1904; 370p. [Google Scholar]
- McCaffery, K.R.; Tranetzki, J.; Piechura, J. Summer foods of deer in northern Wisconsin. J. Wild. Manag. 1974, 38, 215–219. [Google Scholar] [CrossRef]
- Korschgen, L.J.; Porath, W.R.; Torgerson, O. Spring and summer foods of deer in the Missouri Ozarks. J. Wildl. Manag. 1980, 44, 89–97. [Google Scholar] [CrossRef]
- Crawford, H.S. Seasonal food selection and digestibility by tame white-tailed deer in Central Maine. J. Wild. Manag. 1982, 46, 974–982. [Google Scholar] [CrossRef]
- Daigle, C.; Crêtes, M.; Lesage, L.; Ouellet, J.P.; Huot, J. Summer diet of two white-tailed deer, Odocoileus virginianus, populations living at low and high density in Southern Quebec. Can. Field-Nat. 2004, 118, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
- Kalač, P. A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Towards chemical and nutritional inventory of Portuguese wild edible mushrooms in different habitats. Food Chem. 2012, 130, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Sawaya, W.N.; Al-Shalhat, A.; Al-Sogair, A.; Al-Mohammad, M. Chemical Composition and Nutritive Value of Truffles of Saudi Arabia. J. Food Sci. 1985, 50, 450–453. [Google Scholar] [CrossRef]
- Beluhan, S.; Ranogajec, A. Chemical composition and non-volatile components of Croatian wild edible mushrooms. Food Chem. 2011, 124, 1076–1082. [Google Scholar] [CrossRef]
- Palanisamy, M.; Gil-Ramírez, A.; Ruiz-Rodríguez, A.; Marín, F.R.; Reglero, G.; Soler-Rivas, C. Testing edible mushrooms to inhibit the pancreatic lipase activity by an in vitro digestion model. Int. J. Food. Sci. Technol. 2012, 47, 1004–1010. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H.; Guinness, F.E.; Albon, S.D. Red Deer: Behavior and Ecology of Two Sexes; University of Chicago Press: Chicago, IL, USA, 1982. [Google Scholar]
- Barboza, P.S.; Bowyer, R.T. Sexual segregation in dimorphic deer: A new gastrocentic hypothesis. J. Mammal. 2000, 81, 473–489. [Google Scholar] [CrossRef] [Green Version]
- Akyüz, M. Nutritive value, flavonoid content and radical scavenging activity of the truffe (Terfezia boudieri Chatin). J. Soil Sci. Plant Nutr. 2013, 13, 143–151. [Google Scholar]
- Cohen, N.; Cohen, J.; Asatiani, M.; Varshney, V.; Yu, H.-T.; Yang, Y.-C.; Li, Y.-H.; Mau, J.-L.; Wasser, S. Chemical Composition and Nutritional and Medicinal Value of Fruit Bodies and Submerged Cultured Mycelia of Culinary-Medicinal Higher Basidiomycetes Mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Colak, A.; Faiz, O.; Sesli, E. Nutritional composition of some wild edible mushrooms. Turk. J. Biochem. 2009, 34, 25–31. [Google Scholar]
| Sample Size | Males | Females | |||
|---|---|---|---|---|---|
| Juveniles | Adults | Juveniles | Adults | Total | |
| 2014 | 0 | 15 | 2 | 30 | 47 |
| 2015 | 5 | 8 | 3 | 32 | 48 |
| Total | 5 | 23 | 5 | 62 | 95 |
| Species | Total Reads |
|---|---|
| Hypholoma capnoides | 1,111,546 |
| Lactarius deliciosus | 946,384 |
| Cantharellus lutescens | 837,572 |
| Armillaria ostoyae1 | 331,324 |
| Suillus pictus | 305,891 |
| Russula nauseosa1 | 304,270 |
| Suillus bresadolae | 299,284 |
| Cortinarius alboviolaceus | 263,161 |
| Paxillus involutus | 259,158 |
| Craterellus tubaeformis | 257,369 |
| Species | Total Occurrence |
|---|---|
| Lactarius deliciosus | 199 |
| Cantharellus lutescens | 159 |
| Hypholoma capnoides | 155 |
| Cortinarius alboviolaceus | 144 |
| Craterellus tubaeformis | 122 |
| Russula cessans | 108 |
| Suillus bresadolae | 106 |
| Hebeloma velutipes | 106 |
| Cortinarius caninus | 105 |
| Suillus sp. | 98 |
| Models | Explanatory Variables | β | SE | t | CI5% | CI95% |
|---|---|---|---|---|---|---|
| Females | ||||||
| Age + Year | Intercept | 65.4 | 5.1 | 12.9 | 55.5 | 75.4 |
| Age (juvenile) | −29.4 | 13.2 | −2.2 | −55.2 | −3.6 | |
| Year (2015) | 33.7 | 6.9 | 4.9 | 20.1 | 47.2 | |
| Males | ||||||
| Age + Year | Intercept | 60.1 | 5.7 | 10.6 | 49.0 | 71.1 |
| Age (juvenile) | −29.2 | 12.5 | −2.3 | −53.6 | −4.7 | |
| Year (2015) | 20.3 | 9.6 | 2.1 | 1.5 | 39.1 |
| Models | Explanatory Variables | β | SE | t | CI5% | CI95% |
|---|---|---|---|---|---|---|
| Adult females | ||||||
| Year + Lactation | Intercept | 51.1 | 8.1 | 6.3 | 35.2 | 67.0 |
| Year (2015) | 37.0 | 8.6 | 4.3 | 20.3 | 53.8 | |
| Lactation (lactating) | 19.2 | 9.0 | 2.1 | 1.6 | 36.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadotte, M.; Richard, J.H.; Bérubé, J.A.; Côté, S.D. Mycophagy of White-Tailed Deer (Odocoileus virginianus Zimmermann) in the Boreal Forest. Forests 2021, 12, 1247. https://doi.org/10.3390/f12091247
Cadotte M, Richard JH, Bérubé JA, Côté SD. Mycophagy of White-Tailed Deer (Odocoileus virginianus Zimmermann) in the Boreal Forest. Forests. 2021; 12(9):1247. https://doi.org/10.3390/f12091247
Chicago/Turabian StyleCadotte, Myriam, Julien H. Richard, Jean A. Bérubé, and Steeve D. Côté. 2021. "Mycophagy of White-Tailed Deer (Odocoileus virginianus Zimmermann) in the Boreal Forest" Forests 12, no. 9: 1247. https://doi.org/10.3390/f12091247
APA StyleCadotte, M., Richard, J. H., Bérubé, J. A., & Côté, S. D. (2021). Mycophagy of White-Tailed Deer (Odocoileus virginianus Zimmermann) in the Boreal Forest. Forests, 12(9), 1247. https://doi.org/10.3390/f12091247