Virtual Truffle Hunting—A New Method of Burgundy Truffle (Tuber aestivum Vittad.) Site Typing


Abstract
:1. Introduction
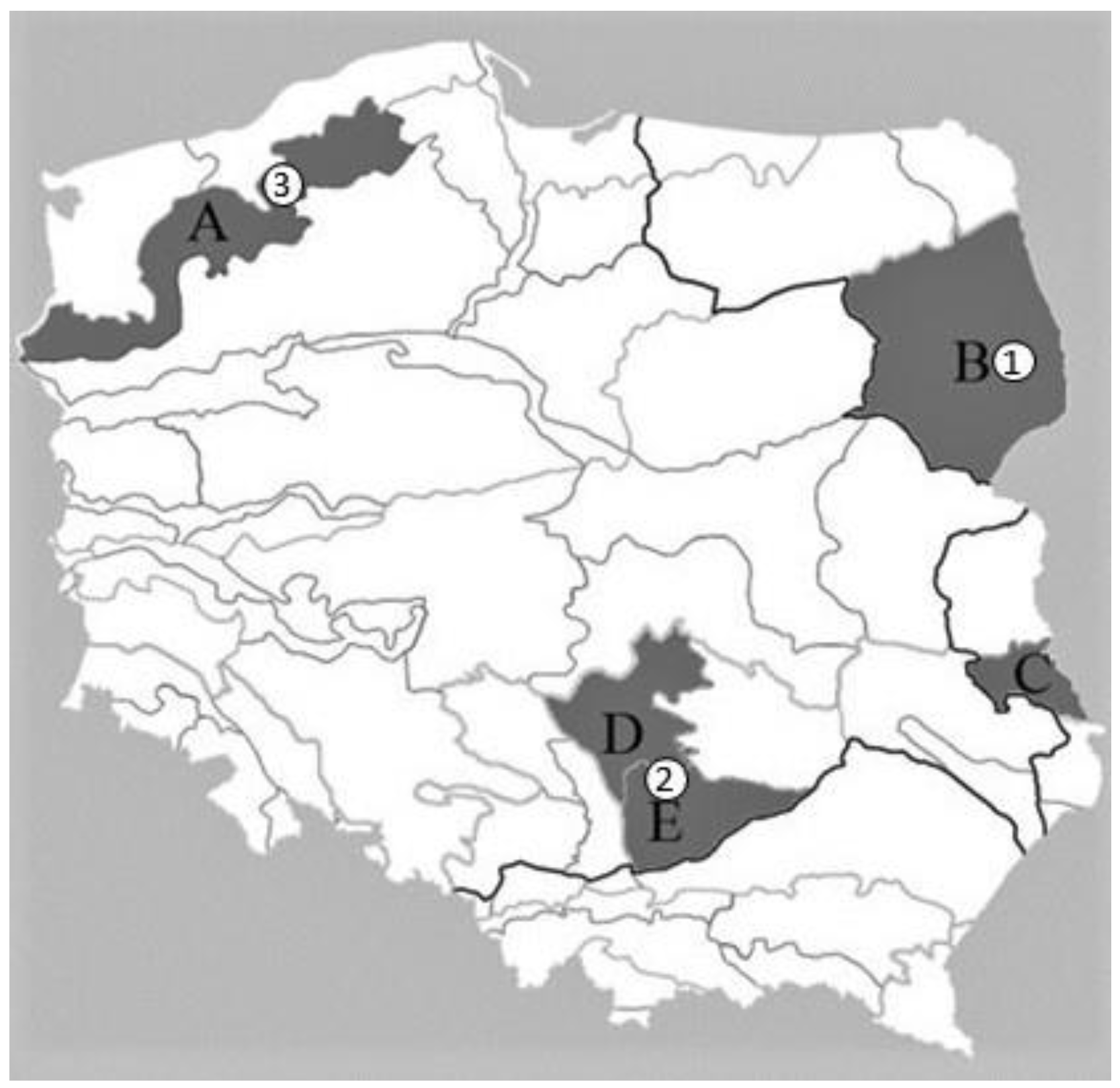
2. Materials and Methods
- bedrock;
- chemical and granulometric composition analysis of soil samples (in the laboratory) [14];
- altitude;
- forest habitat type;
- the structure of the site and the undergrowth layer;
- description of trees and shrubs species that were associated with truffle mycorrhizae;
- information on the presence of orchids and indicator insects included in the method “Virtual Truffle Hunting”.
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Büntgen, U.; Egli, S.; Camarero, J.J.; Fischer, E.M.; Stobbe, U.; Kauserud, H.; Tegel, W.; Sproll, L.; Stenseth, N.C. Drought-induced decline in Mediterranean truffle harvest. Nat. Clim. Chang. 2012, 2, 827–829. [Google Scholar] [CrossRef]
- Pieroni, A. The changing ethnoecological cobweb of white truffle (Tuber magnatum Pico) gatherers in South Piedmont, NW Italy. J. Ethnobiol. Ethnomed. 2016, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilszczańska, D.; Sierota, Z.; Palenzona, M. New Tuber species found in Poland. Mycorrhiza 2008, 18, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Gruszecka, A.; Hilszczańska, D.; Gil, W.; Kosel, B. Historia i perspektywy użytkowania i badań trufli w Polsce. Sylwan 2017, 16, 320–327. [Google Scholar] [CrossRef]
- Rosa-Gruszecka, A.; Hilszczańska, D.; Gil, W.; Kosel, B. Truffle renaissance in Poland–history, present and prospects. J. Ethnobiol. Ethnomed. 2017, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hilszczańska, D.; Rosa-Gruszecka, A.; Sikora, K.; Szmidla, H. First report of Tuber macrosporum occurrence in Poland. Sci. Res. Essays 2013, 8, 1096–1099. [Google Scholar] [CrossRef] [Green Version]
- Chmiel, M.A. Checklist of Polish larger Ascomycetes. In Biodiversity of Poland; Mirek, Z., Ed.; Szafer Institute of Botany: Cracow, Poland; Polish Academy of Sciences: Cracow, Poland, 2006; Volume 8, p. 152. [Google Scholar]
- Merenyi, Z.; Varga, T.; Geml, J.; Orczán, Á.K.; Chevalier, G.; Bratek, Z. Phylogenyand phylogeography of the Tuber brumale aggr. Mycorrhiza 2014, 24, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Mleczko, P.; Chachuła, P.; Kozak, M. Hypogeous fungi of Pieniny and Gorce, two mountain ranges of the Polish Carpathians. In Proceedings of the XVIIth Congress of European Mycologists, Madeira, Portugal, 21–25 September 2015. [Google Scholar]
- Chachuła, P. Macrofungi in the Dłubnia Landscape Park–preliminary results. Przegląd Przyr. 2018, 29, 58–71. [Google Scholar]
- Ławrynowicz, M. Four Tuber species accompanying T. mesentericum in natural sites in Poland. An. del Jardín Bot. Madr. 2009, 66, 145–149. [Google Scholar] [CrossRef]
- Delmas, J. Tuber spp. In The Biology and Cultivation of Edible Mushrooms; Chang, S.T., Hayes, W.A., Eds.; Academic Press: London, UK, 1978; pp. 645–681. [Google Scholar]
- Lulli, L.; Bragato, G.; Gardin, L. Occurrence of Tuber melanosporum in relation to soil surface layer properties and soil differentiation. Plant Soil 1999, 214, 85–92. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Rosa-Gruszecka, A.; Gawryś, R.; Horak, J. The effect of soil properties and vegetation characteristic in determining the frequency of fruiting bodies of the Burgundy truffle in Southern Poland. Ecoscience 2019, 26, 113–122. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Szmidla, H.; Sikora, K.; Rosa-Gruszecka, A. Soil properties conducive to Tuber aestivum Vitt. fruiting bodies formation in the Nida Basin stands. Pol. J. Environ. Stud. 2019, 28, 1713–1718. [Google Scholar] [CrossRef]
- Pacioni, G.; Ragnelli, A.M.; Miranda, M. Truffle development and interactions with the biotic environment. In Biotechnology of Ectomycorrhizae; Stocchi, V., Bonfante, P., Nuti, M., Eds.; Springer: Boston, MA, USA, 1995; pp. 213–227. [Google Scholar] [CrossRef]
- Stobbe, U.; Egli, S.; Tegel, W.; Peter, M.; Sproll, L.; Büntgen, U. Potential and limitations of Burgundy truffle cultivation. Appl. Microbiol. Biotechnol. 2013, 97, 5215–5224. [Google Scholar] [CrossRef] [PubMed]
- Gažo, J.; Miko, M.; Chevalier, G. First results of inventory research on economically important species of truffles (Tuber) in the Tribec Mountains. Acta Fytotech. Zootech. 2005, 8, 66–71. [Google Scholar]
- Moser, B.; Büntgen, U.; Molinier, V.; Peter, M.; Sproll, L.; Stobbe, U.; Tegel, W.; Egli, S. Ecological indicators of Tuber aestivum habitats in temperate European beech forests. Fungal Ecol. 2017, 29, 59–66. [Google Scholar] [CrossRef]
- Pacioni, G.; Comandini, O. Tuber. In Ectomycorrhizal Fungi: Key Genera in Profile; Cairney, J.W.G.S., Chambers, M., Eds.; Springer: Heidelberg, Germany, 1999; pp. 163–186. [Google Scholar]
- Granetti, B.; de Angelis, A.; Materozzi, G. Umbria Terra di Tartufi. Regione Umbria: Assessorato Regionale Agricoltura; Foreste, Caccia e Pesca: Perugia, Italy, 2005. [Google Scholar]
- García-Montero, L.G.; Casermeiro, M.A.; Manjón, J.L.; Hernando, I. Impact of active soil carbonate and burn size on the capacity of the rockrose Cistus laurifolius to produce Tuber melanosporum carpophores in truffle culture. Mycol. Res. 2007, 111, 734–739. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Rosa-Gruszecka, A.; Szmidla, H. Characteristic of Tuber spp. localities in natural stands with emphasis on plant species composition. Acta Mycol. 2014, 49, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Selosse, M.A.; Faccio, A.; Scappaticci, G.; Bonfante, P. Chlorophyllous and achlorophyllous specimens of Epipactis microphylla (Neottieae, Orchidaceae) are associated with ectomycorrhizal septomycetes, including truffles. Microb. Ecol. 2004, 47, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Ouanphanivanh, N.; Merényi, Z.; Orczán, Á.K.; Bratek, Z.; Szigeti, Z.; Illyés, Z. Could orchids indicate truffle habitats? Mycorrhizal association between orchids and truffles. Acta Biol. Szeged. 2008, 52, 229–232. [Google Scholar]
- Ellenberg, H.E. Indicator values of plants in Central Europe. Scr. Geobot. 1974, 18, 1–248. [Google Scholar]
- Gryndler, M.; Šmilauer, P.; Šťovíček, V.; Nováková, K.; Hršelová, H.; Jansa, J. Truffle biogeography—A case study revealing ecological niche separation of different Tuber species. Ecol. Evol. 2017, 7, 4275–4288. [Google Scholar] [CrossRef]
- Molina, R.; Trappe, J.M. Patterns of ectomycorrhizal host specificity and potential among Pacific Northwest conifers and fungi. For. Sci. 1982, 28, 423–458. [Google Scholar]
- Benucci, G.M.; Bonito, G.; Falini, L.B.; Bencivenga, M. Mycorrhization of Pecan trees (Carya illinoinensis) with commercial truffle species: Tuber aestivum Vittad. and Tuber borchii Vittad. Mycorrhiza 2012, 22, 383–392. [Google Scholar] [CrossRef]
- Stobbe, U.; Büntgen, U.; Sproll, L.; Tegel, W.; Egli, S.; Fink, S. Spatial distribution and ecological variation of re-discovered German truffle habitats. Fungal Ecol. 2012, 5, 591–599. [Google Scholar] [CrossRef]
- Miko, M.; Gažo, J.; Priecel, J.; Vernarec, J.; Bratek, Z. Soils indicators of suitability for cultivation of burgundy truffle (Tuber aestivum Vitt.) in Slovak Republic. Sci. J. Phytotechny Zootech. 2009, 12, 70–73. [Google Scholar]
- Csorbainé, A.G. Studies on Cultivation Possibilities of Summer Truffle (Tuber aestivum Vittad.) and Smooth Black Truffle (Tuber macrosporum Vittad.) in Hungary. Ph.D. Thesis, Szent Istvan University, Gödöllő, Hungary, 2001. [Google Scholar]
- Wedén, C.; Chevalier, G.; Danell, E. Tuber aestivum (syn. T. uncinatum) biotopes and their history on Gotland, Sweden. Mycol. Res. 2004, 108, 304–310. [Google Scholar] [CrossRef]
- Bruhn, J.; Hall, M. Burgundy Black Truffle Cultivation in an Agroforestry Practice. In Action in Agroforestry; The Center for Agroforestry at the University of Missouri: Columbia, MO, USA, 2011. [Google Scholar]
- Johnson, C.N. Interactions between mammals and ectomycorrhizal fungi. Trends Ecol. Evol. 1996, 11, 503–507. [Google Scholar] [CrossRef]
- Vernes, K.; Cooper, T.; Green, S. Seasonal fungal diets of small mammals in an Australian temperate forest ecosystem. Fungal Ecol. 2015, 18, 107–114. [Google Scholar] [CrossRef]
- Santo, P.D. Interazioni Pianta–Micorriza–Insetto: Il Modello Quercus sp.–Tuber sp.–Leiodes cinnamomea (Panzer). Ph.D. Thesis, University of Molise, Campobasso, Italy, 2013. [Google Scholar]
- Arzone, A. Reperti ecologici ed etologici di Liodes cinnamomea Panzer vivente su Tuber melanosporum Vittadini (Coleoptera Staphylinoidea). Ann. Della Fac. Di Sci. Agrar. Della Univ. Degli Studi Di Torino 1970, 5, 317–357. [Google Scholar]
- Arzone, A. Nuovi reperti sulla biologia di Liodes cinnamomea Panzer in Tuber magnatum Pico (Coleoptera, Staphylinoidea). Allionia Boll. Dell’instituto ed Orto Bot. Dell’università Di Torino 1971, 17, 121–129. [Google Scholar]
- Rosa-Gruszecka, A.; Gange, A.C.; Harvey, D.J.; Jaworski, T.; Hilszczański, J.; Plewa, R.; Konwerski, S.; Hilszczańska, D. Insect-truffle interactions–potential threats to emerging industries? Fungal Ecol. 2017, 25, 59–63. [Google Scholar] [CrossRef]
- Hochberg, M.E.; Bertault, G.; Poitrineau, K.; Janssen, A. Olfactory orientation of the truffle beetle, Leiodes cinnamomea. Entomol. Exp. Appl. 2003, 109, 147–153. [Google Scholar] [CrossRef]
- Fogel, R.; Peck, S.B. Ecological Studies of Hypogeous Fungi, I. Coleoptera associated with sporocarps. Mycologia 1975, 67, 741–747. [Google Scholar] [CrossRef]
- Newton, A.F. Mycophagy in Staphylinoidea (Coleoptera). In Fungus–Insect Relationships: Perspectives in Ecology and Evolution; Wheeler, Q., Blackwell, M., Eds.; Columbia University Press: New York, NY, USA, 1984; pp. 302–353. [Google Scholar]
- Pacioni, G.; Bologna, M.A.; Laurenzi, M. Insect attraction by Tuber: A chemical explanation. Mycol. Res. 1991, 95, 1359–1363. [Google Scholar] [CrossRef]
- Bratek, Z.; Papp, L.; Merkl, O.; Takács, V. Föld alatti gombákon élő rovarok. Mikológiai Közlemények 1993, 31, 55–65. [Google Scholar]
- Byk, A.; Mokrzycki, T.; Rosa-Gruszecka, A.; Tylkowski, S.; Zamojski, M. Grzybolcowate (Bolboceratidae) i wygonakowate (Ochodaeidae) -aktywność, wymagania ekologiczne i metody obserwacji. SiM CEPL 2016, 49, 124–141. [Google Scholar]
- Büntgen, U.; Tegel, W.; Egli, S.; Stenseth, N.C. Truffles and climate change. Front. Ecol. Environ. 2011, 9, 150–151. [Google Scholar] [CrossRef]
- Ponce, R.A.; Ágreda, T.; Águeda, B.; Aldea, J.; Martínez-Peña, F.; Modrego, M.P. Soil physical properties influence “black truffle” fructification in plantations. Mycorrhiza 2014, 24, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Benucci, G.M.; Bonito, G.; Falini, L.B.; Bencivenga, M.; Donnini, D. Truffles, timber, food, and fuel: Sustainable approaches for multi-cropping truffles and economically important plants. In Edible Ectomycorrhizal Mushrooms; Zambonelli, A., Bonito, G.M., Eds.; Springer: Heidelberg, Germany, 2012; pp. 265–280. [Google Scholar] [CrossRef]
- Gardin, L. I tartufi minori della Toscana. In Gli Ambienti di Crescita Dei Tartufi Marzuolo e Scorzone; ARSIA: Florence, Italy, 2005; p. 55. [Google Scholar]
- Wedén, C.; Danell, E.; Tibell, L. Species recognition in the truffle genus Tuber–the synonyms Tuber aestivum and Tuber uncinatum. Environ. Microbiol. 2005, 7, 1535–1546. [Google Scholar] [CrossRef]
- Wedén, C.; Pettersson, L.; Danell, E. Truffle cultivation in Sweden: Results from Quercus robur and Corylus avellana field trials on the island of Gotland. Scand. J. For. Res. 2009, 24, 37–53. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Szmidla, H.; Horak, J.; Rosa-Gruszecka, A. Ectomycorrhizal communities in a Tuber aestivum Vittad.orchard in Poland. Open Life Sci. 2016, 11, 348–357. [Google Scholar] [CrossRef]
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L.; Trappe, M.J. Diversity, Ecology, and Conservation of Truffle Fungi in Forests of the Pacific Northwest; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2009. [CrossRef] [Green Version]
- Streiblová, E.; Gryndlerová, H.; Gryndler, M. Truffle brûlé: An efficient fungal life strategy. FEMS Microbiol. Ecol. 2012, 80, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Notivoli, R.; Martín-Santafé, M.; Sánchez, S.; Barriuso, J.J. Cultivation potentiality of black truffle in Zaragoza province (Northeast Spain). J. Maps 2016, 12, 994–998. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Barreda, S.; Sánchez, S.; Marco, P.; Serrano-Notivoli, R. Agro-climatic zoning of Spanish forests naturally producing black truffle. Agric. For. Meteorol. 2019, 269, 231–238. [Google Scholar] [CrossRef]
- Ghetti, P.F.; Chierici, E. Indice Biotico Esteso (IBE): I Macroinvertebrati Nel Controllo Della Qualita Degli Ambienti di Acque Correnti: Manuale di Applicazione; Agenzia provinciale per la protezione dell’ambiente: Trento, Italy, 1997. [Google Scholar]
- Central Geological Database. Available online: https://geolog.pgi.gov.pl/ (accessed on 25 March 2021).
- Forest Data Bank. Available online: https://www.bdl.lasy.gov.pl (accessed on 25 March 2021).
- Virtual Truffle Hunting (Software). Available online: https://github.com/AleksandraRosa/Virtual-Truffle-Hunting/blob/main/VirtualTruffleHunting.exe (accessed on 27 April 2021).
- De Laurentis, G.; Spinelli, S. Carta Della Vocazionalità Tartuficola Della Regione Abruzzo Risultati Dell’indagine Conclusiva; ARSSA: Lanciano, Italy, 2009. [Google Scholar]
- Laruccia, N.; Marletto, V.; Leonardi, P.; Puliga, F.; Zambonelli, A. Map of suitability for the spontaneous growth of Tuber magnatum in Emilia-Romagna (Italy). Ital. J. Mycol. 2020, 49, 38–53. [Google Scholar] [CrossRef]
- Hilszczańska, D. Polskie Trufle. In Skarb Odzyskany. O hodowli i Kulinariach Podziemnego Przysmaku; CILP: Warsaw, Poland, 2016. [Google Scholar]
- Serrano-Notivoli, R.; Incausa-Ginés, A.; Barriuso-Vargas, J.; Sánchez-Durán, S.; Martín-Santafé, M. Modelización espacial del hábitat potencial de la trufa negra (Tuber melanosporum Vittad.) en la provincia de Huesca (España). ITEA-Inf. Técnica Económica Agrar. 2015, 111, 227–246. [Google Scholar] [CrossRef]
- Pogoda i klimat. Available online: https://meteomodel.pl/ (accessed on 29 April 2021).
- Zawadzka, D.; Kwiecień, E. Puszcze i lasy Polski. In Encyklopedia Ilustrowana; Oficyna Wydawnicza Multico: Warsaw, Poland, 2011. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Possibility of Truffle Fruiting Body Occurrence On Site (Preference Level) | ||||
|---|---|---|---|---|
| Characteristics of Forest Site | High (Optimal Conditions) 3 pts | Medium (Conditions within Tolerance Limit) 2 pts | Low (Conditions at the Tolerance Limit) 1 pts | None (Conditions Outside the Tolerance Limit) 0 pts |
| Bedrock | Chalk Corallian limestones Gypsum Limestone Lithothamnium Limestones Marly limestone Oolite Pelagic limestones Shelly limestones | Encrinite Marlstone Nummulite limestone Seekreide Terra rossa Travertine | Calcareous gaizes Dolomite Marly schist Other rocks containing calcium | Rocks not containing any form of calcium |
| Soil pH in H2O | >7.2 | 6.8–7.1 | 6.7–5.7 | <5.7 |
| Ca [cmol(+)/kg] | 30–60 | 15–30 or >60 | <15 | Not present |
| CaCO3 [%] | >5 | 1–5 | <1 | Not present |
| C/N | >15 | 10–15 | 5–10 | <5 |
| Soil structure | Clay loam Loam Sandy clay loam Silty clay Silty clay loam | Clay | Sandy loam Sandy clay Silt loam | Sand Loamy sand Silt |
| Altitude (m.a.s.l.) | <600 | 600–1000 | 1000–1600 | >1600 |
| Forest habitat type (Fr.—fresh, Mo.—moist, U—upland, BF—broadleaved forest, M—mixed) | Fr. BF; Fr. UBF | Fr. MBF; Fr. UMBF | Mo. BF; Mo. UBF; Mo. MBF; Mo. UMBF | Other forest habitat types |
| Forest structure | Open canopy closure, presence of gaps, paths or tracks | Moderate canopy closure | Full canopy closure, some shrubs are present | Full canopy closure, dense shrub layer |
| Tree and shrub mycorrhizal partners of Tuber aestivum | More than 2 species in tree layer, presence of Corylus avellana | At least 1 species in tree layer and at least one tree or shrub species in lower forest layer | At least 1 species of tree or shrub in tree or shrub layer. | Lack of tree or shrub mycorrhizal partners of Tuber aestivum |
| Orchids presence (Cephalanthera, Cypripedium or Epipactis genera) | More than one species (3 pts) | One species (2 pts) | No species of Cephalanthera, Cypripedium or Epipactis genera (0 pts) | |
| Insects associated with truffle | Three points (3 pts) should be added to the final equation, if at least one of the insect species occurs in the subarea (not dependent on number of species or their abundance):
| |||
| Trees and Shrubs Taxa | |
|---|---|
| Angiospermae | Gymnospermae |
| Betula pendula Carpinus betulus Corylus avellana Fagus sylvatica Quercus spp. Populus spp. Tilia spp. Ulmus spp. | Abies alba Picea abies Pinus nigra Pinus sylvestris |
| Plot No. | Macroregion | Bedrock (Age) | Altitude (m.a.s.l.) | Tree Species Dominant (Age) and Accompanying Tree Species | Forest Habitat Type |
|---|---|---|---|---|---|
| 1. | West Pomeranian Lake District | Glacial till (Weichselian glaciation) | 20–75 | Fagus sylvatica (155 yr.), Quercus petraea, Acer pseudoplatanus, Pinus sylvestris | Fr. BF |
| 2. | West Pomeranian Lake District | Terminal moraine till (Weichselian glaciation) | 135–158 | Fagus sylvatica (148 yr.), Carpinus betulus, Pinus sylvestris, Picea abies | Fr. MBF |
| 3. | West Pomeranian Lake District | Travertine (Late Carboniferous) | 96–118 | Fagus sylvatica (98 yr.), Quercus robur | Fr. BF |
| 4. | North Podlasie Lowland | Sandur sand and gravel (Riss glaciation) | 174–175 | Tilia cordata (150 yr.), Picea abies | Fr. MBF |
| 5. | North Podlasie Lowland | Glacial till (Riss glaciation) | 179–180 | Picea abies (175 yr.), Quercus robur, Pinus sylvestris | Fr. MBF |
| 6. | Volhynia Polesie | Chalk (Late Cretaceous) | 241–243 | Quercus robur (98 yr.), Carpinus betulus, Pinus sylvestris | Fr. BF |
| 7. | Volhynia Polesie | Chalk (Late Cretaceous) | 219–230 | Quercus robur (73 yr.), Carpinus betulus | Fr. BF |
| 8. | Przedbórz Upland | Limestone (Late Jurassic) | 295–320 | Quercus robur (23 yr.), Fagus sylvatica, Pinus sylvestris, Betula pendula | Fr. UMBF |
| 9. | Przedbórz Upland | Glacial till (Riss glaciation) | 210–215 | Pinus sylvestris (100 yr.), Quercus robur | Fr. BF |
| 10. | Nida Basin | Gypsum (Miocene) | 244–246 | Quercus petraea (126 yr.), Carpinus betulus, Tilia cordata | Fr. BF |
| 11. | Nida Basin | Gypsum (Miocene) | 219–228 | Fagus sylvatica (48 yr.), Quercus petraea, Carpinus betulus | Fr. BF |
| 12. | Nida Basin | Marly limestone (Late Cretaceous) | 301–303 | Quercus robur (92 yr.), Carpinus betulus | Fr. UBF |
| 13. | Nida Basin | Marly limestone (Late Cretaceous) | 300–305 | Quercus robur (84 yr.), Pinus sylvestris, Carpinus betulus | Fr. UBF |
| 14. | Nida Basin | Marly limestone (Late Cretaceous) | 297–301 | Acer pseudoplatanus (32 yr.), Tilia cordata, Carpinus betulus | Fr. UBF |
| 15. | Nida Basin | Lithothamnium limestones (Miocene) | 291–295 | Quercus robur (65 yr.), Carpinus betulus | Fr. MBF |
| 16. | Nida Basin | Lithothamnium limestones (Miocene) | 300–301 | Quercus robur (70 yr.), Carpinus betulus | Fr. MBF |
| 17. | Nida Basin | Lithothamnium limestones (Miocene) | 294–295 | Carpinus betulus (65 yr.), Quercus robur | Fr. MBF |
| 18. | Nida Basin | Lithothamnium limestones (Miocene) | 295–296 | Quercus robur (77 yr.) | Fr. UBF |
| 19. | Nida Basin | Marly limestone (Late Cretaceous) | 251–252 | Quercus robur (147 yr.) | Fr. UBF |
| 20. | Nida Basin | Marly limestone (Late Cretaceous) | 250–251 | Quercus robur (147 yr.), Tilia cordata, Carpinus betulus, Acer pseudoplatanus | Fr. UBF |
| Characteristics of Forest Sites | Site Number | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Bedrock | Glacial till (0 pts) | Terminal moraine till (0 pts) | Travertine (2 pts) | Sandur sand and gravel (0 pts) | Glacial till (0 pts) |
| Soil pH in H2O | 7.8 (3 pts) | 7.7 (3 pts) | 7.7 (3 pts) | 7.5 (3 pts) | 5.3 (0 pts) |
| Ca [cmol(+)/kg] | 18.87 (2 pts) | 32.25 (3 pts) | 45.59 (3 pts) | 9.24 (1 pts) | 1.63 (1 pts) |
| CaCO3 [%] | 7.23 (2 pts) | 42.7 (3 pts) | 60.24 (2 pts) | 3.78 (2 pts) | 0 (0 pts) |
| C/N | 16.2 (2 pts) | 22.1 (1 pts) | 25.2 (1 pts) | 13.7 (3 pts) | 13.9 (3 pts) |
| Soil structure | Sandy loam (1 pts) | Sand (0 pts) | Loamy sand (0 pts) | Loamy sand (0 pts) | Loamy sand (0 pts) |
| Altitude (m.a.s.l.) | 20–75 (3 pts) | 135–158 (3 pts) | 96–118 (3 pts) | 174–175(3 pts) | 179–180 (3 pts) |
| Forest habitat type | Fr. BF (3 pts) | Fr. MBF (2 pts) | Fr. BF (3 pts) | Fr. MBF(2 pts) | Fr. MBF (2 pts) |
| Forest structure | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Moderate canopy closure (2 pts) | Full canopy closure, some shrubs are present (1 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Full canopy closure, dense shrub layer (0 pts) |
| Tree and shrubs mycorrhizal partners of Tuber aestivum | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | At least 1species in tree layer and at least one tree or shrub species in lower forest layer (2 pts) | At least 1 species of tree or shrub in tree or shrub layer (1 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) |
| Orchids presence (Cephalanthera, Epipactis or Cypripedium genera) | 1 species (Cephalanthera damasonium) (2 pts) | Not present (0 pts) | Not present (0 pts) | Not present (0 pts) | Not present (0 pts) |
| Insects associated with truffle | Not reported (0 pts) | Not reported (0 pts) | Not reported (0 pts) | Odonteus armiger (3 pts) | Odonteus armiger (3 pts) |
| Final note (possibility of occurrence) | 26/36 (medium) | 20/36 (low) | 20/36 (low) | 21/36 (low) | 15/36 (none) |
| Presence of hypogeous fungi | Rhizopogon roseolus (1) | Tuber ferrugineum (1) | Elaphomyces muricatus (2) | Melanogaster broomeianus (19) | Not found |
| Characteristics of Forest Sites | Site Number | ||||
|---|---|---|---|---|---|
| 6 | 7 | 8 | 9 | 10 | |
| Bedrock | Chalk (3 pts) | Chalk (3 pts) | Limestone (3 pts) | Glacial till (0 pts) | Gypsum (3 pts) |
| Soil pH in H2O | 7.1 (2 pts) | 7.2 (3 pts) | 7.9 (3 pts) | 4.3 (0 pts) | 7.6 (3 pts) |
| Ca [cmol(+)/kg] | 7.85 (1 pts) | 12.72 (1 pts) | 2.36 (1 pts) | 0.83 (1 pts) | 38.88 (3 pts) |
| CaCO3 [%] | 0.25 (1 pts) | 0.17 (1 pts) | 1.93 (2 pts) | 0 (0 pts) | 0.88 (1 pts) |
| C/N | 13.3 (3 pts) | 13.6 (3 pts) | 11.6 (3 pts) | 16 (2 pts) | 10.9 (3 pts) |
| Soil structure | Sandy loam (1 pts) | Loamy sand (0 pts) | Loamy sand (0 pts) | Loamy sand (0 pts) | Loam (3 pts) |
| Altitude (m.a.s.l.) | 241–243 (3 pts) | 219–230 (3 pts) | 295–320 (3 pts) | 210–215 (3 pts) | 244–246 (3 pts) |
| Forest habitat type | Fr. BF (3 pts) | Fr. BF (3 pts) | Fr. BF (3 pts) | Fr. BF (3 pts) | Fr. BF (3 pts) |
| Forest structure | Full canopy closure, some shrubs are present (1 pts) | Full canopy closure, some shrubs are present (1 pts) | Full canopy closure, dense shrub layer (0 pts) | Full canopy closure, dense shrub layer (0 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) |
| Tree and shrubs mycorrhizal partners of Tuber aestivum | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) |
| Orchids presence (Cephalanthera, Epipactis or Cypripedium genera) | More than 1 species (Cypripedium calceolus, Cephalanthera damasonium) (3 pts) | 1 species (Cephalanthera damasonium) (2 pts) | Not present (0 pts) | Not present (0 pts) | More than 1 species (Cypripedium calceolus, Cephalanthera damasonium) (3 pts) |
| Insects associated with truffle | Not reported (0 pts) | Leiodes oblonga (3 pts) | Not reported (0 pts) | Not reported (0 pts) | Leiodes oblonga; L. cinnamomea (3 pts) |
| Final note (possibility of occurrence) | 24/36 (medium) | 26/36 (medium) | 21/36 (low) | 12/36 (none) | 34/36 (high) |
| Presence of hypogeous fungi | Tuber excavatum (17), Genea verrucosa (1), Elaphomyces sp. (1) | Tuber rufum (5), Genea spp. (4), Hymenogaster sp. (1) | Not found | Elaphomyces sp. (2) | Tuber aestivum (6) |
| Characteristics of Forest Sites | Site Number | ||||
|---|---|---|---|---|---|
| 11 | 12 | 13 | 14 | 15 | |
| Bedrock | Gypsum (3 pts) | Marly limestone (3 pts) | Marly limestone (3 pts) | Marly limestone (3 pts) | Lithothamnium limestones (3 pts) |
| Soil pH in H2O | 7.5 (3 pts) | 5.6 (0 pts) | 7.5 (3 pts) | 7.5 (3 pts) | 7.3 (3 pts) |
| Ca [cmol(+)/kg] | 40.6 (3 pts) | 20.78 (2 pts) | 33.56 (3 pts) | 30.35 (3 pts) | 30.63 (3 pts) |
| CaCO3 [%] | 20.24 (3 pts) | 0.33 (1 pts) | 16.77 (3 pts) | 38.04 (3 pts) | 6.77 (3 pts) |
| C/N | 16.3 (2 pts) | 15.3 (2 pts) | 17.4 (2 pts) | 23.7 (1 pts) | 12.7 (3 pts) |
| Soil structure | Sandy loam (1 pts) | Silt loam (1 pts) | Loam (3 pts) | Silt loam (1 pts) | Sandy loam (1 pts) |
| Altitude (m.a.s.l.) | 219–228 (3 pts) | 301–303 (3 pts) | 300–305 (3 pts) | 297–301 (3 pts) | 291–295 (3 pts) |
| Forest habitat type | Fr. BF (3 pts) | Fr. UBF (3 pts) | Fr. UBF (3 pts) | Fr. UBF (3 pts) | Fr. MBF (2 pts) |
| Forest structure | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Full canopy closure, some shrubs are present (1 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) |
| Tree and shrubs mycorrhizal partners of Tuber aestivum | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) |
| Orchids presence (Cephalanthera, Epipactis or Cypripedium genera) | Not present (0 pts) | Not present (0 pts) | 1 species (Cypripedium calceolus) (2 pts) | More than 1 species (Cypripedium calceolus, Epipactis sp., Cephalanthera damasonium) (3 pts) | More than 1 species (Cypripedium calceolus, Epipactis sp., Cephalanthera damasonium) (3 pts) |
| Insects associated with truffle | Leiodes oblonga, L. cinnamomea (3 pts) | Leiodes oblonga (3 pts) | Leiodes oblonga (3 pts) | Leiodes oblonga (3 pts) | Not reported (0 pts) |
| Final note (possibility of occurrence) | 30/36 (medium) | 24/36 (low) | 34/36 (high) | 30/36 (medium) | 30/36 (medium) |
| Presence of hypogeous fungi | Tuber aestivum (38) | Tuber aestivum (7) | Tuber aestivum (29) | Tuber aestivum (3), T. excavatum (24) | Tuber aestivum (20) |
| Characteristics of Forest Sites | Site Number | ||||
|---|---|---|---|---|---|
| 16 | 17 | 18 | 19 | 20 | |
| Bedrock | Lithothamnium limestones (3 pts) | Lithothamnium limestones (3 pts) | Lithothamnium limestones (3 pts) | Marly limestone (3 pts) | Marly limestone (3 pts) |
| Soil pH in H2O | 7.3 (3 pts) | 7.5 (3 pts) | 7.5 (3 pts) | 5.9 (1 pts) | 7.4 (3 pts) |
| Ca [cmol(+)/kg] | 45.3 (3 pts) | 30.63 (3 pts) | 28.02 (2 pts) | 31.07 (3 pts) | 46.37 (3 pts) |
| CaCO3 [%] | 17.76 (3 pts) | 6.77 (3 pts) | 3.14 (2 pts) | 1.82 (2 pts) | 14.62 (3 pts) |
| C/N | 15.3 (2 pts) | 13.0 (3 pts) | 13.2 (3 pts) | 12.1 (3 pts) | 14.5 (3 pts) |
| Soil structure | Sandy loam (1 pts) | Loamy sand (0 pts) | Sand (0 pts) | Sandy loam (1 pts) | Loam (3 pts) |
| Altitude (m.a.s.l.) | 300–301 (3 pts) | 294–295 (3 pts) | 295–296 (3 pts) | 251–252 (3 pts) | 250–251 (3 pts) |
| Forest habitat type | Fr. MBF (2 pts) | Fr. MBF (2 pts) | Fr. UBF (3 pts) | Fr. UBF (3 pts) | Fr. UBF (3 pts) |
| Forest structure | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Open canopy closure, presence of gaps, paths or tracks (3 pts) | Moderate canopy closure (2 pts) |
| Tree and shrubs mycorrhizal partners of Tuber aestivum | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) | At least 1species in tree layer and at least one tree or shrub species in lower forest layer (2 pts) | At least 1species in tree layer and at least one tree or shrub species in lower forest layer (2 pts) | More than 2 species in tree layer, presence of Corylus avellana (3 pts) |
| Orchids presence (Cephalanthera, Epipactis or Cypripedium genera) | More than 1 species (Cypripedium calceolus, Epipactis sp., Cephalanthera sp.) (3 pts) | 1 species (Cephalanthera damasonium) (2 pts) | 1 species (Cephalanthera damasonium) (2 pts) | 1 species (Cephalanthera damasonium) (2 pts) | 1 species (Cephalanthera damasonium) (2 pts) |
| Insects associated with truffle | Not reported (0 pts) | Not reported (0 pts) | Not reported (0 pts) | Leiodes oblonga (3 pts) | Leiodes oblonga (3 pts) |
| Final note (possibility of occurrence) | 29 /36 (medium) | 28/36 (medium) | 26/36 (medium) | 29/36 (medium) | 34/36 (high) |
| Presence of hypogeous fungi | Tuber excavatum (14), T. aestivum (13) | Tuber excavatum (6), T. aestivum (1), Genea sp. (2) | Tuber aestivum (6) | Tuber aestivum (3) | Tuber aestivum (24) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa-Gruszecka, A.; Hilszczańska, D.; Pacioni, G. Virtual Truffle Hunting—A New Method of Burgundy Truffle (Tuber aestivum Vittad.) Site Typing. Forests 2021, 12, 1239. https://doi.org/10.3390/f12091239
Rosa-Gruszecka A, Hilszczańska D, Pacioni G. Virtual Truffle Hunting—A New Method of Burgundy Truffle (Tuber aestivum Vittad.) Site Typing. Forests. 2021; 12(9):1239. https://doi.org/10.3390/f12091239
Chicago/Turabian StyleRosa-Gruszecka, Aleksandra, Dorota Hilszczańska, and Giovanni Pacioni. 2021. "Virtual Truffle Hunting—A New Method of Burgundy Truffle (Tuber aestivum Vittad.) Site Typing" Forests 12, no. 9: 1239. https://doi.org/10.3390/f12091239
APA StyleRosa-Gruszecka, A., Hilszczańska, D., & Pacioni, G. (2021). Virtual Truffle Hunting—A New Method of Burgundy Truffle (Tuber aestivum Vittad.) Site Typing. Forests, 12(9), 1239. https://doi.org/10.3390/f12091239