
Intraspecific Growth Response to Drought of Abies alba in the Southeastern Carpathians



Abstract
1. Introduction
2. Materials and Methods
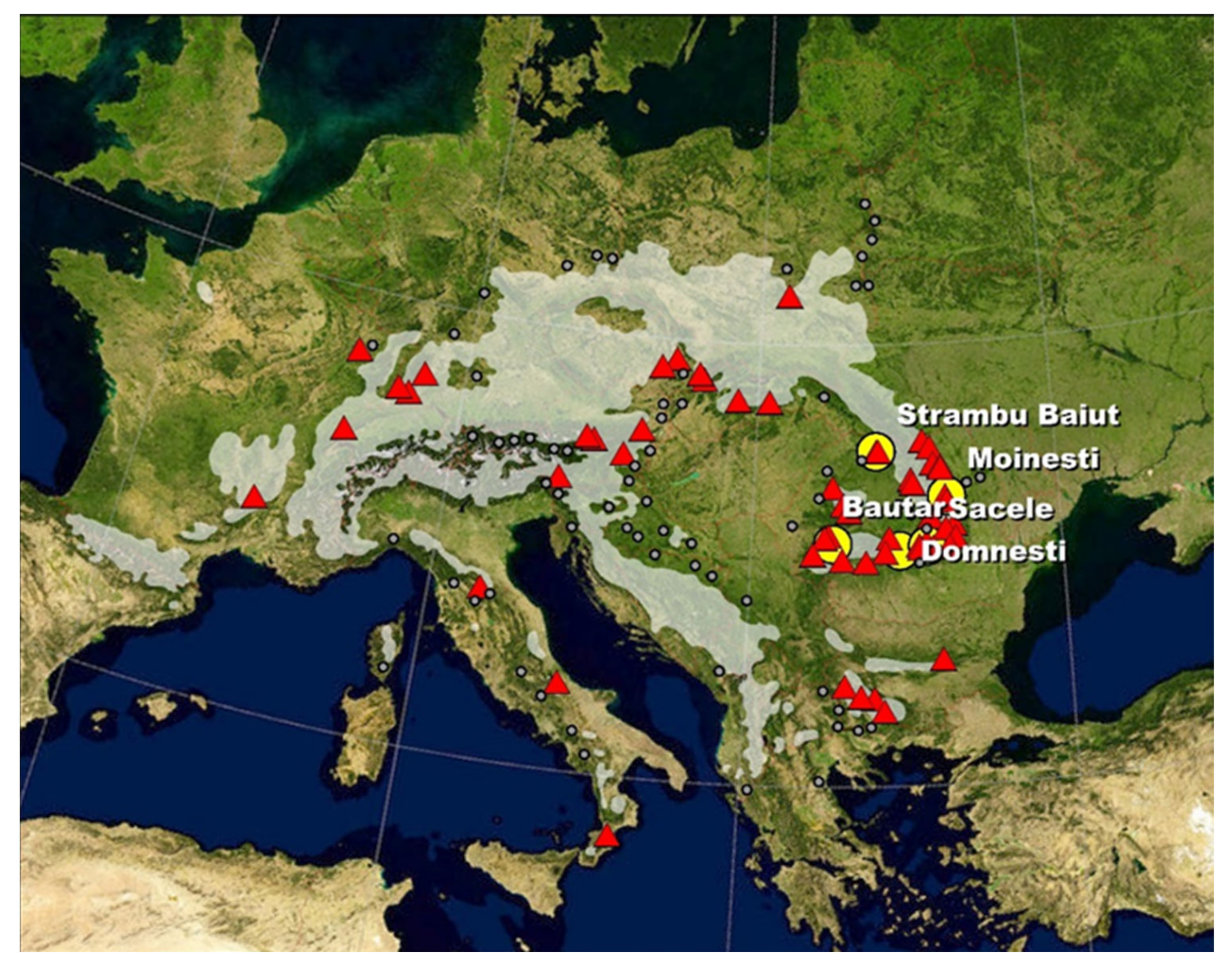
2.1. Trial Site and Plant Material
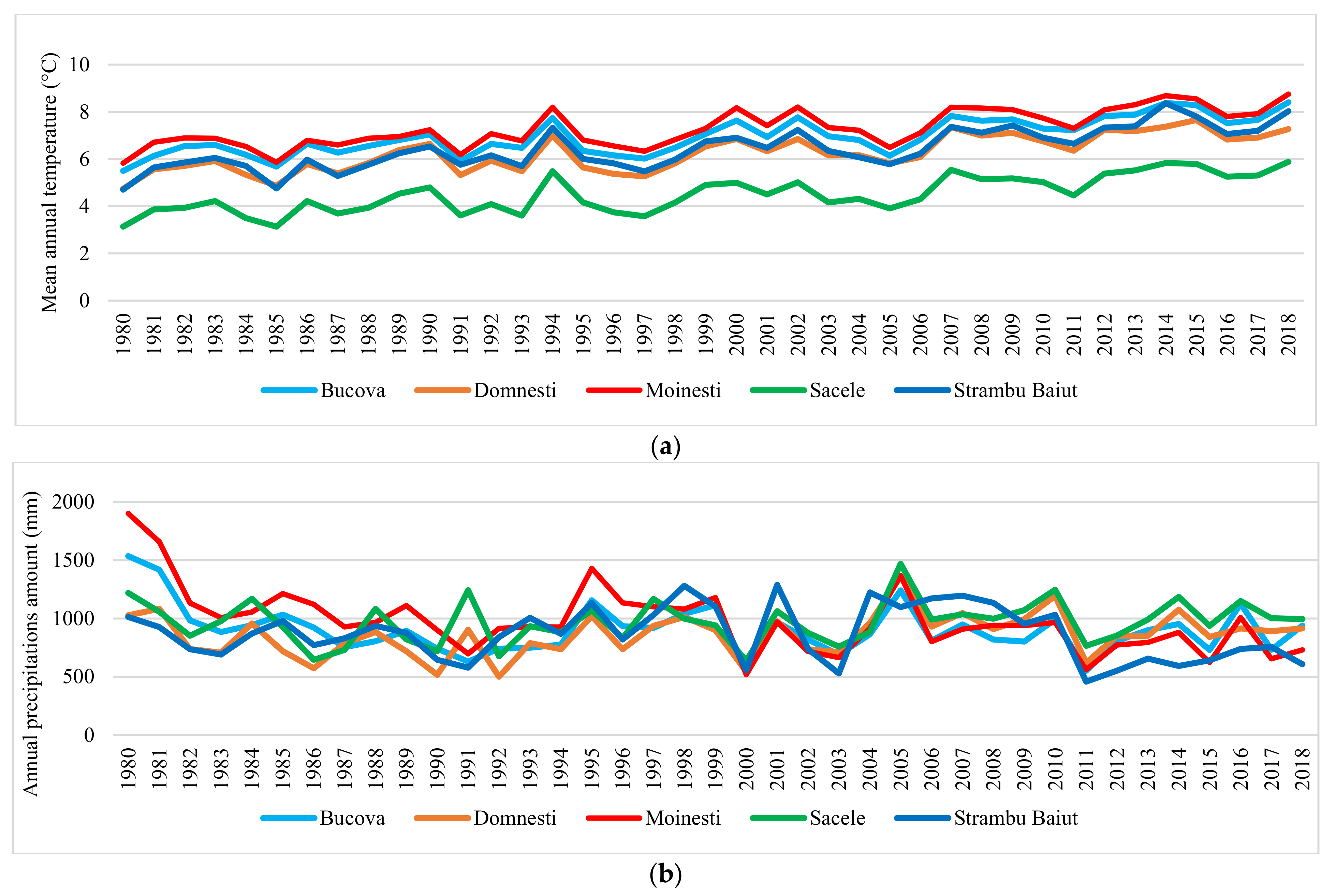
2.2. Phenotypic and Climatic Data
2.3. Determination of Drought Events and Drought Response Parameters
2.4. Data Analysis
3. Results
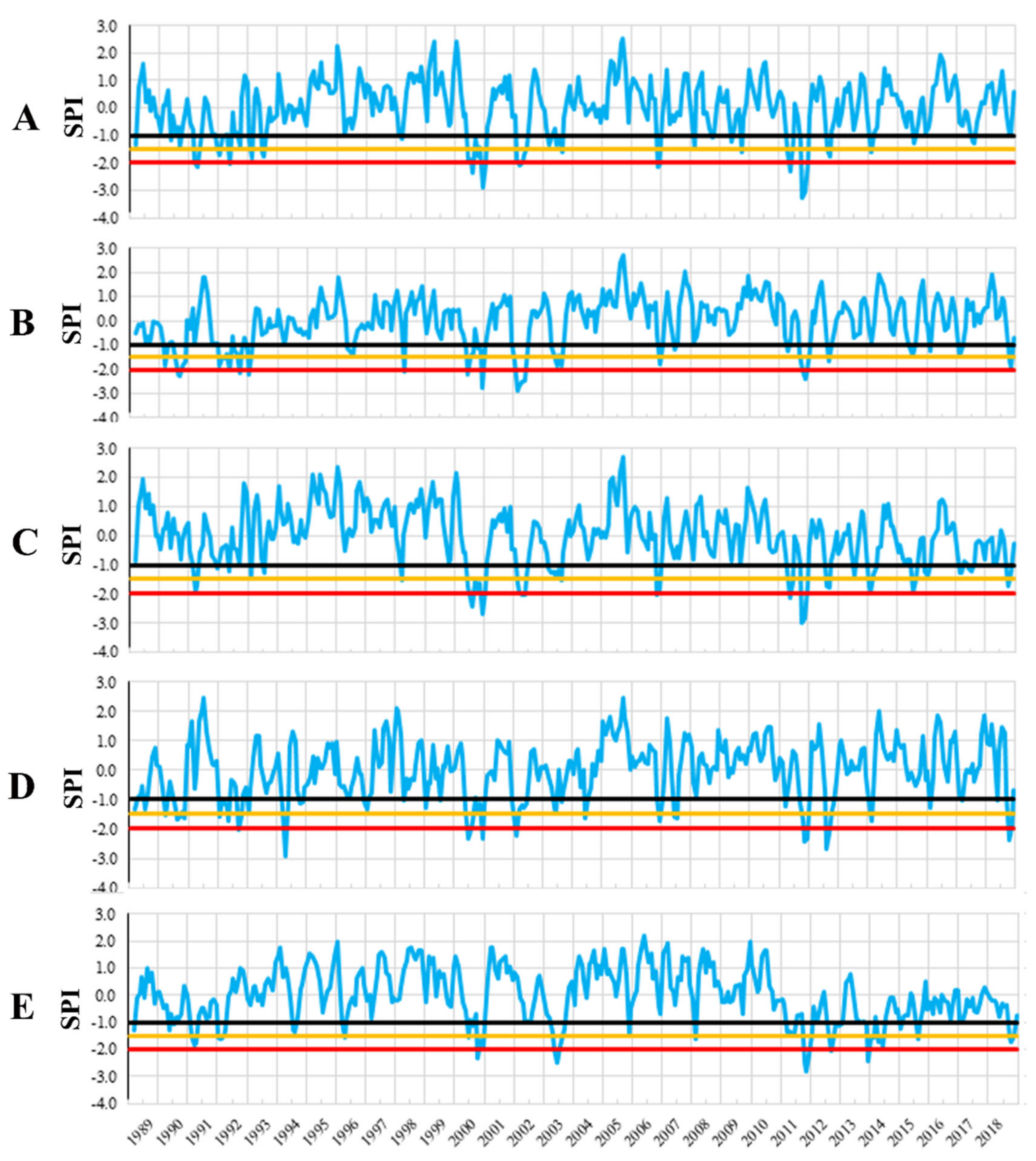
3.1. Identification of Drought Years
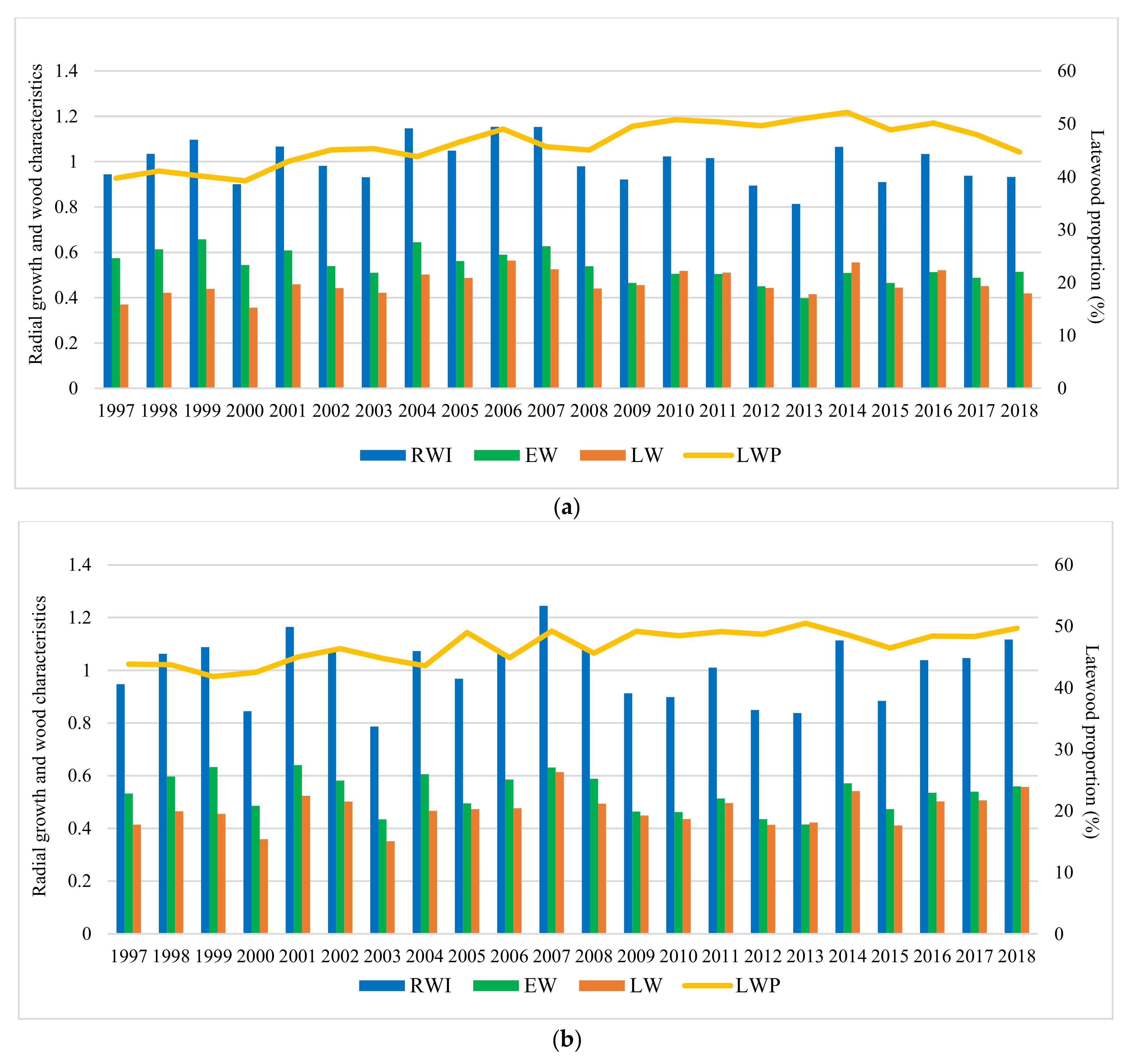
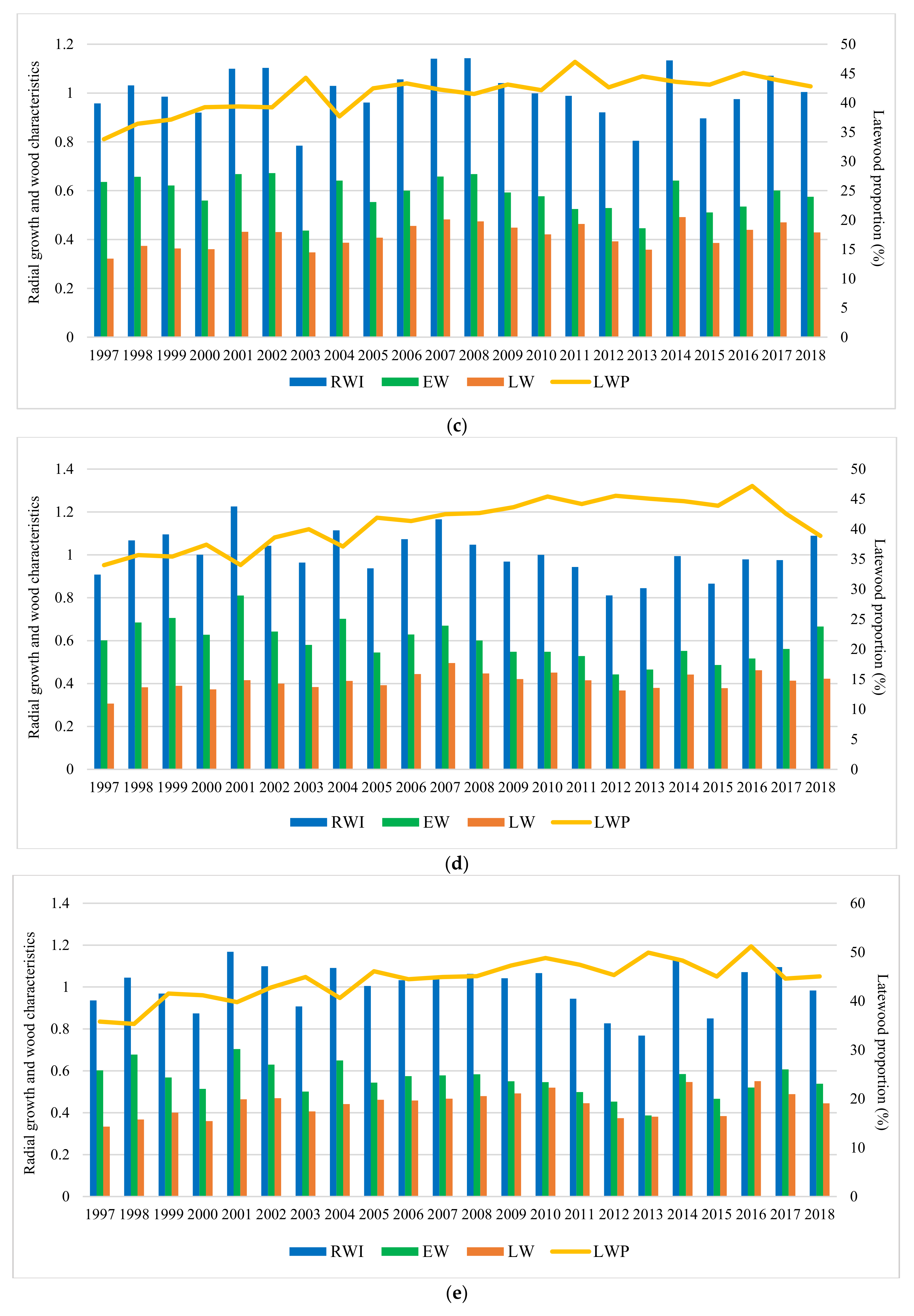
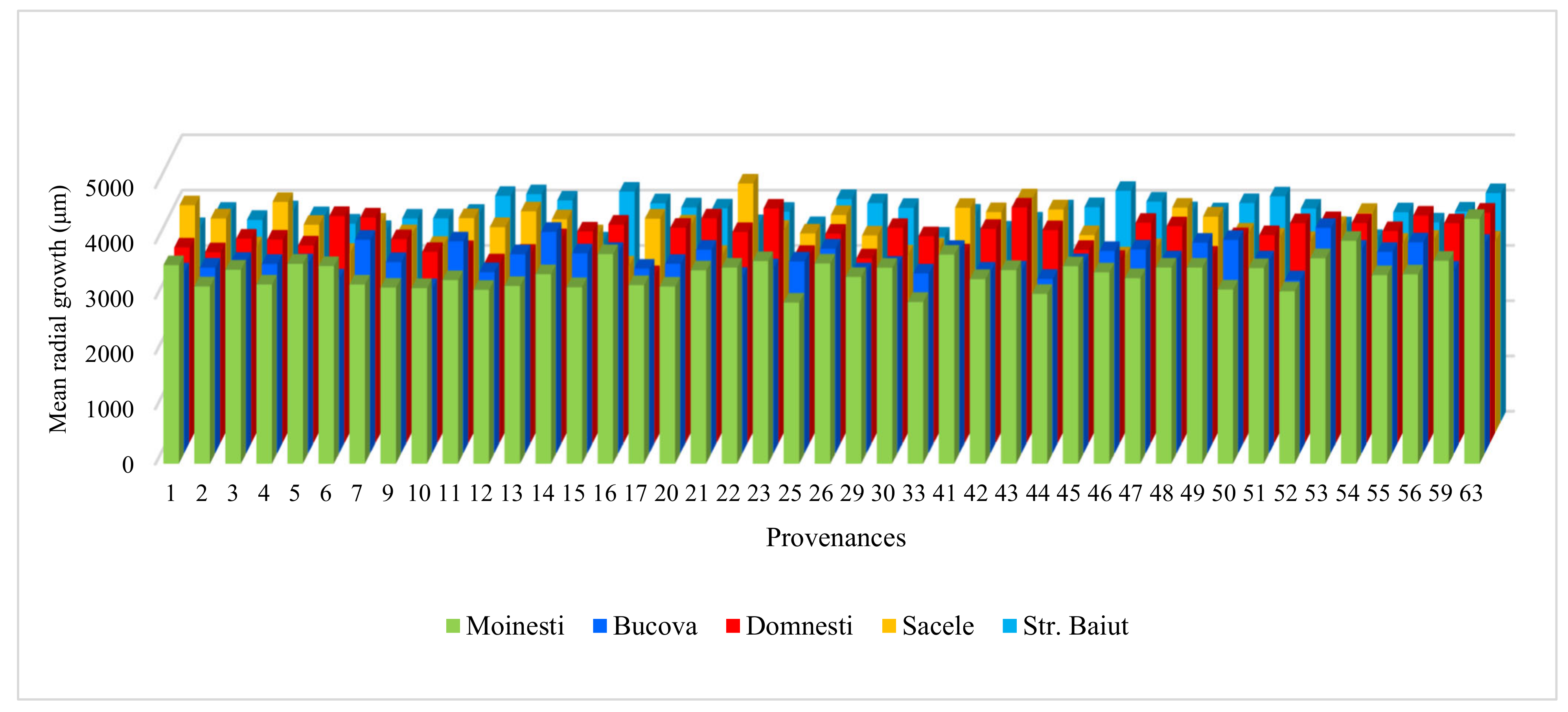
3.2. The Effect of Provenance, Site, and Year on Radial Growth and Wood Characteristics
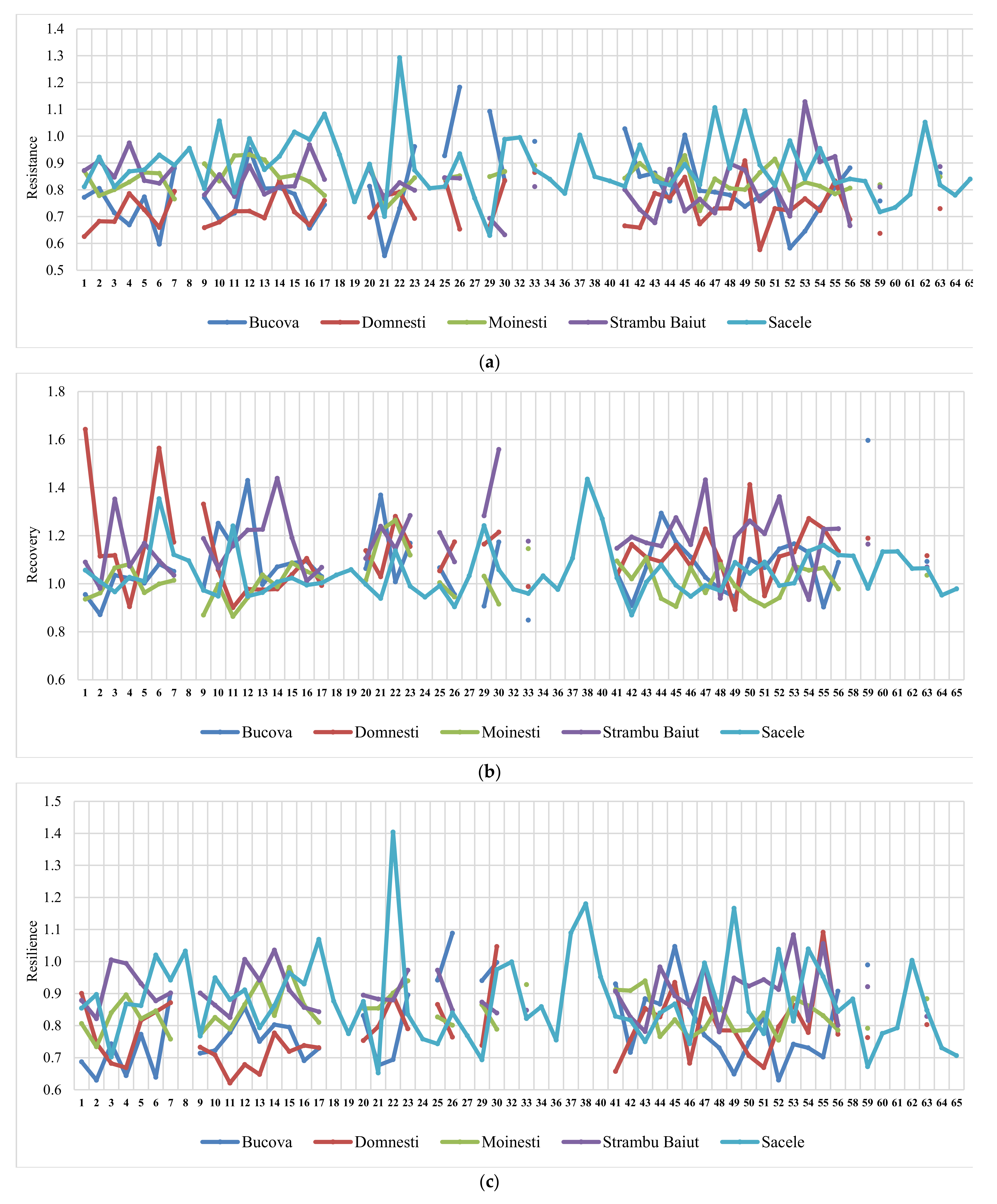
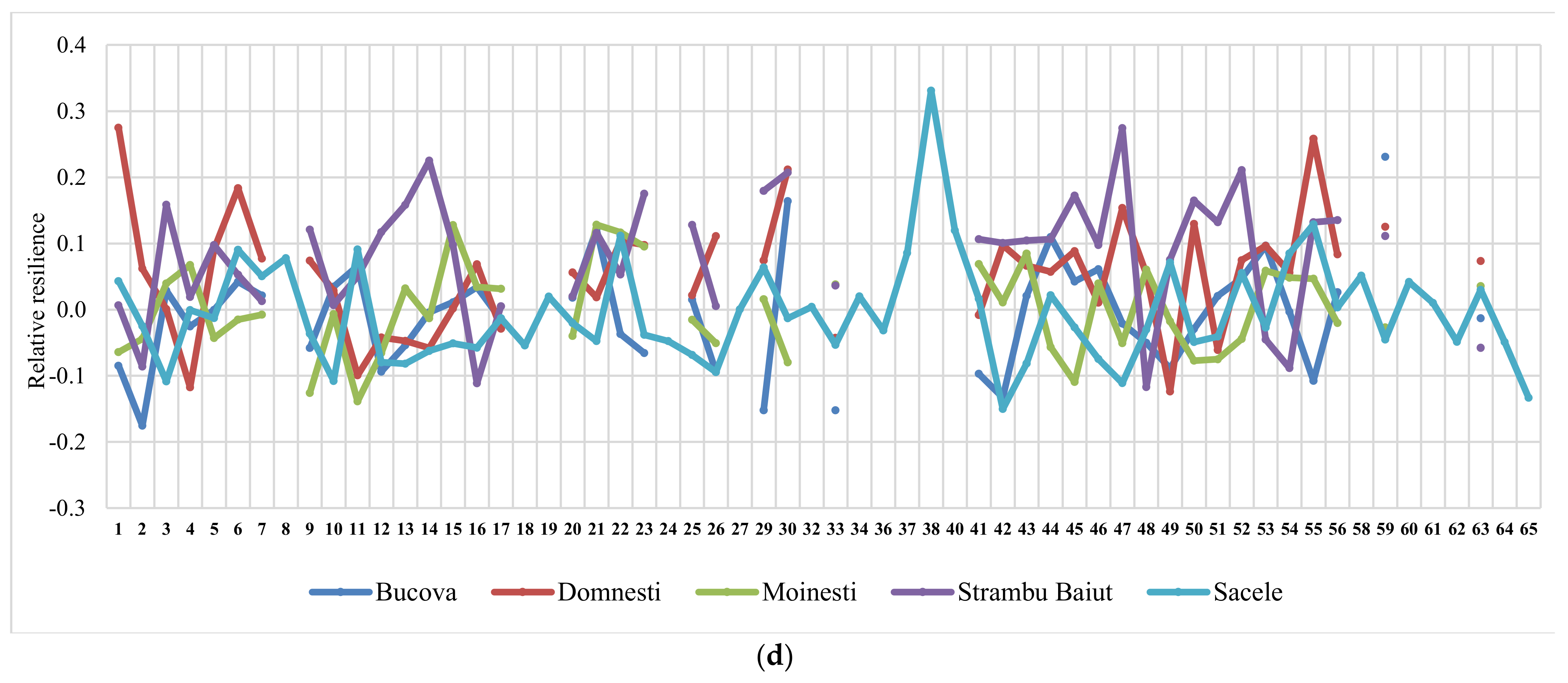
3.3. Genetic Variation in Drought Response
3.4. Phenotypic Correlations
3.5. Growth Response Functions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Prov. | Provenance | Country | Location within Distribution Range | Longitude E | Latitude N | Altitude (m) |
|---|---|---|---|---|---|---|
| 1 | Cheia | Romania | Southeastern edge | 25°55′ | 45°25′ | 950 |
| 2 | Azuga I | Romania | Southeastern edge | 25°35′ | 45°25′ | 1100 |
| 3 | Ghelinta | Romania | Eastern edge | 26°20′ | 45°55′ | 880 |
| 4 | Avrig | Romania | Southeastern edge | 24°30′ | 45°40′ | 660 |
| 5 | Valea Motilor | Romania | Core | 22°45′ | 46°30′ | 750 |
| 6 | Bucium | Romania | Core | 23°10′ | 46°15′ | 910 |
| 7 | Vadul Dobri | Romania | Southeastern edge | 22°35′ | 45°40′ | 1150 |
| 8 | Tismana | Romania | Southeastern edge | 23°00′ | 45°05′ | 950 |
| 9 | Polovragi | Romania | Southeastern edge | 23°48′ | 45°15′ | 1100 |
| 10 | Cozia | Romania | Southeastern edge | 24°20′ | 45°20′ | 1300 |
| 11 | Gura Teghii | Romania | Eastern edge | 26°20′ | 45°35′ | 1100 |
| 12 | Naruja I | Romania | Eastern edge | 26°40′ | 45°40′ | 800 |
| 13 | Soveja | Romania | Eastern edge | 26°40′ | 46°00′ | 750 |
| 14 | Asau | Romania | Eastern edge | 26°25′ | 46°25′ | 1050 |
| 15 | Tusnad | Romania | Eastern edge | 25°50′ | 46°10′ | 650 |
| 16 | Toplita | Romania | Eastern edge | 25°25′ | 46°55′ | 930 |
| 17 | Garcin | Romania | Southeastern edge | 25°45′ | 45°35′ | 1000 |
| 18 | Rasnov | Romania | Southeastern edge | 25°32′ | 45°35′ | 700 |
| 19 | Valiug | Romania | Southeastern edge | 22°10′ | 45°12′ | 600 |
| 20 | Rusca Montana | Romania | Southeastern edge | 22°28′ | 45°35′ | 880 |
| 21 | Azuga II | Romania | Southeastern edge | 25°35′ | 45°25′ | 1125 |
| 22 | Toplita II | Romania | Eastern edge | 23°25′ | 46°55′ | 900 |
| 23 | Rakitovo | Bulgaria | Southeastern edge | 24°05′ | 41°59′ | 1550 |
| 24 | Devin | Bulgaria | Southeastern edge | 24°24′ | 41°42′ | 1500 |
| 25 | Kitilovo | Bulgaria | Southeastern edge | 26°13′ | 42°54′ | 500 |
| 26 | St. Dimitrov | Bulgaria | Southeastern edge | 23°09′ | 42°15′ | 1450 |
| 27 | Raslog | Bulgaria | Southeastern edge | 23°40′ | 42°01′ | 1600 |
| 29 | Vallombrosa | Italy | Southern edge | 11°33′ | 43°45′ | 960 |
| 30 | Paularo | Italy | Core | 13°30′ | 46°31′ | 950 |
| 32 | San Bruno | Italy | Southern edge | 16°20′ | 38°33′ | 1250 |
| 33 | Abeti Soprani | Italy | Southern edge | 14°20′ | 41°52′ | 800 |
| 34 | Trieben | Austria | Core | 14°30′ | 47°28′ | 1125 |
| 36 | Passail | Austria | Core | 15°32′ | 47°13′ | 800 |
| 37 | Liezen | Austria | Core | 14°15′ | 47°31′ | 800 |
| 38 | Hohe Wand | Austria | Core | 16°04′ | 47°49′ | 750 |
| 40 | Todtmoos | Germany | Western edge | 8°05′ | 47°47′ | 320 |
| 41 | Enzklosterle | Germany | Western edge | 8°30′ | 48°16′ | 280 |
| 42 | Sulzburg | Germany | Western edge | 7°43′ | 47°51′ | 560 |
| 43 | Greseuss | France | Western edge | 6°09′ | 48°28′ | 400 |
| 44 | Lepilat | France | Western edge | 4°00′ | 44°40′ | 340 |
| 45 | Le Joux | France | Western edge | 6°15′ | 46°40′ | 260 |
| 46 | Naruja II | Romania | Eastern edge | 26°40′ | 45°40′ | 750 |
| 47 | Moinesti | Romania | Eastern edge | 26°25′ | 46°25′ | 940 |
| 48 | Pangarati | Romania | Eastern edge | 26°10′ | 46°53′ | 860 |
| 49 | Rasca | Romania | Eastern edge | 25°14′ | 47°20′ | 560 |
| 50 | Malini | Romania | Eastern edge | 25°56′ | 47°24′ | 820 |
| 51 | Gura Putnei | Romania | Eastern edge | 25°33′ | 47°47′ | 620 |
| 52 | Solca | Romania | Eastern edge | 24°52′ | 47°40′ | 480 |
| 53 | Botiza | Romania | Core | 23°05′ | 47°40′ | 970 |
| 54 | Strambu Baiut | Romania | Core | 22°55′ | 47°35′ | 760 |
| 55 | Valea Iadului | Romania | Core | 22°40′ | 46°50′ | 800 |
| 56 | Ilisoara Mures | Romania | Eastern edge | 25°08′ | 46°55′ | 1050 |
| 58 | Brezno Michalova | Slovakia | Core | 20°20′ | 48°40′ | 700 |
| 59 | Banska Bystrica | Slovakia | Core | 19°15′ | 48°40′ | 850 |
| 60 | Banska Bystrica | Slovakia | Core | 19°15′ | 48°40′ | 800 |
| 61 | Lidečko | Czech Republic | Core | 18°02′ | 49°05′ | 740 |
| 62 | Vizovice | Czech Republic | Core | 17°52′ | 49°12′ | 650 |
| 63 | Zarovice | Czech Republic | Core | 17°01′ | 49°30′ | 860 |
| 64 | Deblin | Czech Republic | Core | 16°32′ | 49°18′ | 740 |
| 65 | Skarzysko | Poland | Northern edge | 20°50′ | 51°07′ | 130 |

| GLK | RBAR | EPS | SNR | Provenance rbar min | Provenance rbar max | |
|---|---|---|---|---|---|---|
| Bucova | 0.596 | 0.659 | 0.999 | 989.01 | 0.702—Prov. 30 | 0.892—Prov. 17 |
| Domnesti | 0.682 | 0.773 | 0.999 | 1700.15 | 0.752—Prov. 55 | 0.854—Prov. 16 |
| Moinesti | 0.635 | 0.691 | 0.999 | 1096.01 | 0.607—Prov. 33 | 0.905—Prov. 55 |
| Sacele | 0.612 | 0.706 | 0.999 | 1605.15 | 0.633—Prov. 22 | 0.920—Prov. 26 |
| Str. Baiut | 0.627 | 0.689 | 0.999 | 1001.36 | 0.728—Prov. 3 | 0.918—Prov. 50 |
| Trial | Trait | LW | EW | LWP | WD | LAT | LONG | ALT |
|---|---|---|---|---|---|---|---|---|
| SACELE | RW | 0.570 *** | 0.870 *** | −0.258 * | 0.082 | −0.083 | −0.172 | −0.028 |
| LW | - | 0.098 | 0.599 *** | 0.007 | −0.092 | 0.303 * | −0.123 | |
| EW | - | −0.663 *** | 0.090 | −0.043 | −0.389 ** | 0.034 | ||
| LWP | - | 0.059 | −0.126 | 0.522 *** | −0.115 | |||
| WD | - | −0.103 | 0.050 | −0.065 | ||||
| DOMNESTI | RW | 0.885 *** | 0.934 *** | −0.098 | −0.189 | 0.230 | −0.128 | 0.133 |
| LW | - | 0.667 *** | 0.310 * | 0.011 | 0.264 | −0.189 | 0.003 | |
| EW | - | −0.424 ** | −0.292 * | 0.171 | −0.096 | 0.207 | ||
| LWP | - | 0.179 | 0.205 | −0.029 | −0.217 | |||
| WD | - | −0.299 * | −0.173 | −0.170 | ||||
| BUCOVA | RW | 0.495 ** | 0.801 *** | −0.242 | −0.077 | 0.240 | 0.292 * | 0.166 |
| LW | - | −0.123 | 0.709 *** | −0.201 | 0.103 | −0.425 ** | −0.176 | |
| EW | - | −0.764 *** | 0.051 | 0.201 | 0.627 *** | 0.300 * | ||
| LWP | - | −0.172 | −0.059 | −0.723 *** | −0.326 * | |||
| WD | - | −0.240 | 0.050 | −0.031 | ||||
| STRAMBU BAIUT | RW | 0.695 *** | 0.827 *** | −0.071 | −0.304 * | 0.081 | 0.127 | 0.094 |
| LW | - | 0.172 | 0.632 *** | −0.137 | −0.077 | −0.036 | −0.115 | |
| EW | - | −0.594 *** | −0.306 * | 0.166 | 0.198 | 0.230 | ||
| LWP | - | 0.039 | −0.151 | −0.224 | −0.258 | |||
| WD | - | −0.279 * | 0.165 | 0.096 | ||||
| MOINESTI | RW | 0.809 *** | 0.918 *** | −0.192 | −0.362 * | 0.486 ** | −0.100 | 0.072 |
| LW | - | 0.508 *** | 0.397 ** | −0.245 | 0.266 | −0.344 * | −0.005 | |
| EW | - | −0.549 *** | −0.365 * | 0.532 *** | 0.086 | 0.113 | ||
| LWP | - | 0.148 | −0.341 * | −0.382 * | −0.036 | |||
| WD | - | −0.268 | −0.044 | −0.099 |
References
- Field, C.B.; Barros, T.F.V.; Stocker, Q.D.; Dokken, D.J.; Ebi, K.L.; Mastrandrea, M.D.; Mach, K.J.; Plattner, G.K.; Allen, S.K.; Tignor, M. IPCC Report on Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Cambridge University Press: Cambridge, UK, 2012; p. 582. [Google Scholar]
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.O.; Roberts, D.; Skea, J.; Shukla, P.R.; Pirani, A.; Moufouma-Okia, W.; Péan, C.; Pidcock, R. IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Intergovernmental Panel on Climate Change, IPCC: Geneva, Switzerland, 2019; p. 26. [Google Scholar]
- Mishra, A.K.; Singh, V.P. A review of drought concepts. J. Hydrol. 2010, 391, 202–216. [Google Scholar] [CrossRef]
- Naumann, G.; Spinoni, J.; Vogt, J.V.; Barbosa, P. Assessment of drought damages and their uncertainties in Europe. Environ. Res. Lett. 2015, 10, 124013. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Lloret, F.; Escudero, A.; Iriondo, J.M.; Martinez-Vilalta, J.; Valladares, F. Extreme climatic events and vegetation: The role of stabilizing processes. Glob. Chang. Biol. 2012, 18, 797–805. [Google Scholar] [CrossRef]
- Blauhut, V.; Stahl, K.; Stagge, J.H.; Tallaksen, L.M.; de Stefano, L.; Vogt, J. Estimating drought risk across Europe from reported drought impacts, drought indices, and vulnerability factors. Hydrol. Earth Syst. Sci. 2016, 20, 2779–2800. [Google Scholar] [CrossRef]
- Briffa, K.R.; van der Schrier, G.; Jones, P.D. Wet and dry summers in Europe since 1750: Evidence of increasing drought. Int. J. Clim. 2009, 29, 1894–1905. [Google Scholar] [CrossRef]
- Gudmundsson, L.; Seneviratne, S.I. European drought trends. Proc. Int. Assoc. Hydrol. Sci. 2015, 369, 75–79. [Google Scholar] [CrossRef]
- Dumitrescu, A.; Bojariu, R.; Birsan, M.-V.; Marin, L.; Manea, A. Recent climatic changes in Romania from observational data (1961–2013). Theor. Appl. Clim. 2014, 122, 111–119. [Google Scholar] [CrossRef]
- Spinoni, J.; Szalai, S.; Szentimrey, T.; Lakatos, M.; Bihari, Z.; Nagy, A.; Németh, Á.; Kovács, T.; Mihic, D.; Dacic, M.; et al. Climate of the Carpathian Region in the period 1961–2010: Climatologies and trends of 10 variables. Int. J. Clim. 2014, 35, 1322–1341. [Google Scholar] [CrossRef]
- Ionita, M.; Scholz, P.; Chelcea, S. Assessment of droughts in Romania using the Standardized Precipitation Index. Nat. Hazards 2016, 81, 1483–1498. [Google Scholar] [CrossRef]
- Meinshausen, M.; Smit, S.; Calvin, J.H.; Daniel, K.; Kainuma, J.S.; Lamarque, M.L.T.; Matsumoto, J.F.; Montzka, K.; Raper, S.; Riahi, S. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Chang. 2011, 109, 213–241. [Google Scholar] [CrossRef]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Clim. 2018, 38, 1718–1736. [Google Scholar] [CrossRef]
- Hamrick, J. Response of forest trees to global environmental changes. For. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Lindner, M. Developing adaptive forest management strategies to cope with climate change. Tree Physiol. 2000, 20, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Mauri, A.; de Rigo, D.; Caudullo, G. Abies alba in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston, D.T., Mauri, A., Eds.; Publication Office of the European Union: Brusels, Belgium, 2016; pp. 48–49. [Google Scholar]
- Maiorano, L.; Cheddadi, R.; Zimmermann, N.E.; Pellissier, L.; Petitpierre, B.; Pottier, J.; Laborde, H.; Hurdu, B.I.; Pearman, P.B.; Psomas, A.; et al. Building the niche through time: Using 13,000 years of data to predict the effects of climate change on three tree species in Europe. Glob. Ecol. Biogeogr. 2012, 22, 302–317. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How much does climate change threaten European forest tree species distributions? Glog. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Rebetez, M.; Conedera, M.; Augustin, S.; Brang, P.; Tinner, W. What is the potential of silver fir to thrive under warmer and drier climate? Eur. J. For. Res. 2019, 138, 547–560. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.; Carraro, G.; Conedera, M. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Ruosch, M.; Spahni, R.; Joos, F.; Henne, P.D.; Knaap, W.O.; Tinner, W. Past and future evolution of Abies alba forests in Eu-rope—Comparison of a dynamic vegetation model with palaeodata and observations. Glob. Chang. Biol. 2016, 22, 727–740. [Google Scholar] [CrossRef]
- Latreille, A.C.; Pichot, C. Local-scale diversity and adaptation along elevational gradients assessed by reciprocal transplant experiments: Lack of local adaptation in silver fir populations. Ann. For. Sci. 2017, 74, 77. [Google Scholar] [CrossRef]
- Becker, M. The role of climate on present and past vitality of silver fir forests in the Vosges mountains of northeastern France. Can. J. For. Res. 1989, 19, 1110–1117. [Google Scholar] [CrossRef]
- Maaten-Theunissen, M.; Kahle, H.P.; Maaten, E. Drought sensitivity of Norway spruce is higher than that of silver fir along an altitudinal gradient in southwestern Germany. Ann. For. Sci. 2013, 70, 185–193. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian Mountains. Trees 2008, 23, 95–106. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.T.W.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Functional diversity enhances silver fir growth resilience to an extreme drought. J. Ecol. 2016, 104, 1063–1075. [Google Scholar] [CrossRef]
- Bosela, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.A.; Popa, I.; et al. Contrasting effects of environmental change on the radial growth of co-occurring beech and fir trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Klisz, M.; Ukalski, K.; Ukalska, J.; Jastrzębowski, S.; Puchałka, R.; Przybylski, P.; Mionskowski, M.; Matras, J. What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments? Forests 2018, 9, 441. [Google Scholar] [CrossRef]
- Mihai, G.; Bîrsan, M.-V.; Dumitrescu, A.; Alexandru, A.; Mirancea, I.; Ivanov, P.; Stuparu, E.; Teodosiu, M.; Daia, M. Adaptive genetic potential of European silver fir in Romania in the context of climate change. Ann. For. Res. 2018, 61, 95–108. [Google Scholar] [CrossRef]
- Eriksson, G.; Namkoong, G.; Roberds, J.H. Dynamic gene conservation for uncertain futures. For. Ecol. Manag. 1993, 62, 15–37. [Google Scholar] [CrossRef]
- Kremer, A. How well can existing forests withstand climate change. In Climate Change and Forest Genetic Diversity: Implications for Sustainable Forest Management in Europe; Koskela, J., Buck, A., Teissier du Cros, E., Eds.; Bioversity International: Rome, Italy, 2007; pp. 3–17. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia–Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kol-strom, M. Climate change impacts, adaptive capacity and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Mihai, G.; Teodosiu, M.; Birsan, M.-V.; Alexandru, A.-M.; Mirancea, I.; Apostol, E.-N.; Garbacea, P.; Ionita, L. Impact of Climate Change and Adaptive Genetic Potential of Norway Spruce at the South-Eastern Range of Species Distribution. Agric. For. Meteorol. 2020, 291, 108040. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Wykoff, W.R.; Ying, C.C. Physiologic Plasticity, Evolution, and Impacts of a Changing Climate on Pinus Contorta. Clim. Chang. 2001, 50, 355–376. [Google Scholar] [CrossRef]
- Wang, T.; O’Neill, G.A.; Aitken, S.N. Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol. Appl. 2010, 20, 153–163. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ying, C.C.; Spittlehouse, D.L.; Hamilton, D.A. Genetic responses to climate in Pinus contorda: Niche breadth, climate change, and reforestation. Ecol. Monogr. 1999, 69, 375–407. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Kremer, A.; Nagy, L.; Újvári-Jármay, É.; Ducousso, A.; Kóczán-Horváth, A.; Hansen, J.K.; Mátyás, C. Common garden comparisons confirm inherited differences in sensitivity to climate change between forest tree species. PeerJ 2019, 7, e6213. [Google Scholar] [CrossRef] [PubMed]
- Buras, A.; Sass-Klaassen, U.; Verbeek, I.; Copini, P. Provenance selection and site conditions determine growth performance of pedunculate oak. Dendrochronologia 2020, 61, 125705. [Google Scholar] [CrossRef]
- Andalo, C.; Beaulieu, J.; Bousquet, J. The impact of climate change on growth of local white spruce populations in Québec, Canada. For. Ecol. Manag. 2005, 205, 169–182. [Google Scholar] [CrossRef]
- Rweyongeza, D.M.; Yang, R.-C.; Dhir, N.K.; Barnhardt, L.K.; Hansen, C. Genetic Variation and Climatic Impacts on Survival and Growth of White Spruce in Alberta, Canada. Silvae Genet. 2007, 56, 117–127. [Google Scholar] [CrossRef]
- St Clair, J.B.; Mandel, N.L.; Vance-Borland, K.W. Genecology of Douglas fir in western Oregon and Washinton. Ann. Bot. 2005, 96, 1199–1214. [Google Scholar] [CrossRef]
- Chakraborty, D.; Wang, T.; Andre, K.; Konnert, M.; Lexer, M.J.; Matulla, C.; Schueler, S. Selecting Populations for Non-Analogous Climate Conditions Using Universal Response Functions: The Case of Douglas-Fir in Central Europe. PLoS ONE 2015, 10, e0136357. [Google Scholar] [CrossRef] [PubMed]
- Schmidtling, R.C. Use of provenance tests to predict response to climate change: Loblolly pine and Norway spruce. Tree Physiol. 1994, 14, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Gömöry, D.; Longauer, R.; Hlásny, T.; Pacalaj, M.; Strmeň, S.; Krajmerová, D. Adaptation to common optimum in different populations of Norway spruce (Picea abies Karst.). Eur. J. For. Res. 2011, 131, 401–411. [Google Scholar] [CrossRef]
- Kapeller, S.; Lexer, M.J.; Geburek, T.; Hiebl, J.; Schueler, S. Intraspecific variation in climate response of Norway spruce in the eastern Alpine range: Selecting appropriate provenances for future climate. For. Ecol. Manag. 2012, 271, 46–57. [Google Scholar] [CrossRef]
- Klisz, M.; Ukalska, J.; Koprowski, M.; Tereba, A.; Puchałka, R.; Przybylski, P.; Jastrzębowski, S.; Nabais, C. Effect of provenance and climate on intra-annual density fluctuations of Norway spruce Picea abies (L.) Karst. in Poland. Agric. For. Meteorol. 2019, 269, 145–156. [Google Scholar] [CrossRef]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res. 2009, 39, 1259–1269. [Google Scholar] [CrossRef]
- Burczyk, J.; Giertych, M. Response of Norway Spruce (Picea abies [L] Karst) annual increments to drought for various prov-enances and locations. Silvae Genet. 1991, 40, 146–152. [Google Scholar]
- Arend, M.; Kuster, T.; Günthardt-Goerg, M.S.; Dobbertin, M. Provenance-specific growth responses to drought and air warming in three European oak species (Quercus robur, Q. petraea and Q. pubescens). Tree Physiol. 2011, 31, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Dalla-Salda, G.; Martinez-Meier, A.; Cochard, H.; Rozenberg, P. Genetic variation of xylem hydraulic properties shows that wood density is involved inadaptation to drought in Douglas-fir (Pseudotsuga menziesii (Mirb.)). Ann. For. Sci. 2011, 68, 747–757. [Google Scholar] [CrossRef]
- Eilmann, B.; de Vries, S.M.; Ouden, J.D.; Mohren, G.M.; Sauren, P.; Sass-Klaassen, U. Origin matters! Difference in drought tolerance and productivity of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.)) provenances. For. Ecol. Manag. 2013, 302, 133–143. [Google Scholar] [CrossRef]
- George, J.-P.; Schueler, S.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Inter- and intra-specific variation in drought sensitivity in Abies spec. and its relation to wood density and growth traits. Agric. For. Meteorol. 2015, 214, 430–443. [Google Scholar] [CrossRef]
- George, J.-P.; Grabner, M.; Campelo, F.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Schüler, S. Intra-specific variation in growth and wood density traits under water-limited conditions: Long-term-, short-term-, and sudden responses of four conifer tree species. Sci. Total. Environ. 2019, 660, 631–643. [Google Scholar] [CrossRef]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1996; p. 73. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: San Francisco, CA, USA, 1976; p. 582. [Google Scholar]
- Briffa, K.R.; Jones, P.D. Basic chronology statistics and assessment. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1990; pp. 137–153. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; Indiana State University: Terre Haute, IN, USA, 2009; p. 508. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Dumitriu-Tataranu, I.; Ghelmeziu, N.; Florescu, I.; Milea, I.; Mos, V.; Tocan, M. Estimarea Calitatii Lemnului Prin Metoda Carotelor de Sondaj; Editura Tehnica: Bucuresti, Romania, 1983; p. 347. [Google Scholar]
- Dumitrescu, A.; Birsan, M.-V. ROCADA: A gridded daily climatic dataset over Romania (1961–2013) for nine meteorological variables. Nat. Hazards 2015, 78, 1045–1063. [Google Scholar] [CrossRef]
- McKee, T.B.N.; Doesken, J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 9th Conference on Applied Climatology, Dallas, TX, USA, 15–20 January 1995; pp. 179–184. [Google Scholar]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Sagnard, F.; Barberot, C.; Fady, B. Structure of Genetic diversity in Abies alba Mill. from southwestern Alps: Multivariate analysis of adaptive and non-adaptive traits for conservation in France. For. Ecol. Manag. 2002, 157, 175–189. [Google Scholar] [CrossRef]
- Sindelar, J.; Beran, F. Comparison of some exotic species of Abies genus with chosen silver fir provenances on the plots of town Pisek. Commun. Inst. For. Bohem. 2008, 24, 99–114. [Google Scholar]
- Forner, A.; Aranda, I.; Granier, A.; Valladares, F. Differential impact of the most extreme drought event over the last half century on growth and sap flow in two coexisting Mediterranean trees. Plant Ecol. 2014, 215, 703–719. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Brousseau, L.; Postolache, D.; Lascoux, M.; Drouzas, A.D.; Källman, T.; Leonarduzzi, C.; Liepelt, S.; Piotti, A.; Popescu, F.; Roschanski, A.M.; et al. Local Adaptation in European Firs Assessed through Extensive Sampling across Altitudinal Gradients in Southern Europe. PLoS ONE 2016, 11, e0158216. [Google Scholar] [CrossRef]
- Heer, K.; Behringer, D.; Piermattei, A.; Bassler, C.; Brandl, R.; Fady, B.; Jehl, H.; Liepelt, S.; Lorch, S.; Piotti, A. Linking dendroecology and association genetics in natural populations: Stress responses archived in tree rings associate with SNP genotypes in silver fir (Abies alba Mill.). Mol. Ecol. 2018, 27, 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Sutherland, R.A. Stomatal control of xylem embolism. Plant Cell Environ. 1991, 14, 607–612. [Google Scholar] [CrossRef]
- St Clair, J.B.; Howe, G.T. Genetic maladaptation of coastal Douglas-fir seedlings to future climates. Glob. Chang. Biol. 2007, 13, 1441–1454. [Google Scholar] [CrossRef]
- Rosner, S.; Světlík, J.; Andreassen, K.; Børja, I.; Dalsgaard, L.; Evans, R.; Karlsson, B.; Tollefsrud, M.; Solberg, S. Wood density as a screening trait for drought sensitivity in Norway spruce. Can. J. For. Res. 2014, 44, 154–161. [Google Scholar] [CrossRef]







| Trials | Prov. Region | Altitude m | TMA °C | TMVEG °C | SAP mm | PMVEG °C | SPVEG mm | De Martonne Index | De Martonne IndexVEG |
|---|---|---|---|---|---|---|---|---|---|
| Bucova | D2 | 650 | 7.37 | 13.46 | 878 | 92 | 550 | 50 | 23 |
| Domnesti | C2 | 880 | 6.68 | 12.83 | 916 | 104 | 622 | 55 | 27 |
| Moinesti | A2 | 815 | 7.72 | 13.71 | 868 | 91 | 544 | 49 | 23 |
| Sacele | B1 | 1225 | 4.92 | 10.91 | 1001 | 113 | 678 | 67 | 32 |
| Strambu-Baiut | A1 | 890 | 6.90 | 13.24 | 879 | 87 | 521 | 52 | 22 |
| Abbreviation | Wood and Climate Characteristics |
|---|---|
| RW | Ring width |
| EW | Earlywood width |
| LW | Latewood width |
| LWP | Latewood proportion |
| DW | Wood density |
| MAT | Mean annual temperature |
| MTVEG | Mean temperature during growing season (April to September) |
| MTJAN | Mean temperature for January (the coldest month) |
| MTJUL | Mean temperature for July (the warmest month) |
| MTOCT-DEC | Mean temperature from October to December of previous year |
| MTOCT-MAR | Mean temperature from October of previous year to March of current year |
| MTJAN-MAR | Mean temperature from January to March of current year |
| MAP | Mean annual precipitation amount |
| MPVEG | Mean precipitation during growing season |
| MPJAN | Mean precipitation of the coldest month |
| MPJUL | Mean precipitation of the warmest month |
| MPOCT-DEC | Mean precipitation from October to December of previous year |
| MPOCT-MAR | Mean precipitation from October of previous year to March of current year |
| MPJAN-MAR | Mean precipitation from January to March of the current year |
| SAP | Annual precipitation amount |
| SPOCT-DEC | Precipitation amount in autumn-winter of previous year |
| SPOCT-MAR | Precipitation amount from October of previous year to March of current year |
| SPJAN-MAR | Precipitation amount from January to March of current year |
| Trial | 2000 | 2002/2003 (1) | 2011 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Extreme Drought | Severe Drought | Total Months | Extreme Drought | Severe Drought | Total Months | Extreme Drought | Severe Drought | Total Months | |
| Bucova | I, II, VIII, XII | VI, VII, X | 7 | III, IV | II, V | 4 | V, X, XI, XII | IV | 5 |
| Domnesti | VI, XII | VII | 3 | II, III, IV, V | I | 5 | X, XI, XII | - | 3 |
| Moinesti | I, VII, VIII, XII | II, VI, IX, X, XI | 9 | IV, V | II, III | 4 | V, XI | VI, XII | 4 |
| Sacele | VI, VII, XII | VIII, X | 5 | II | I | 2 | XI, XII | X | 3 |
| Strambu Baiut | X, XII | VI, XI | 4 | V, VI, VII | VIII | 4 | X, XI, XII | - | 3 |
| Trial | Source of Variation | DF | Variance (s2) | ||||
|---|---|---|---|---|---|---|---|
| RW | LW | EW | LWP | WD | |||
| SACELE | Provenance (P) | 59 | 8750.95 *** | 2054.92 *** | 5973.12 *** | 0.071 *** | 0.002 *** |
| Repetition (B) | 2 | 0.010 | 0.079 | 0.115 | 0.098 | 0.001 | |
| Year (Y) | 21 | 1.880 *** | 0.289 *** | 1.422 *** | 0.290 *** | - | |
| Interaction P × Y | 1239 | 0.038 *** | 0.011 | 0.018 * | 0.004 | - | |
| Error | 2640 | 0.032 | 0.010 | 0.017 | 0.004 | 0.001 | |
| DOMNESTI | Provenance (P) | 42 | 5568.98 *** | 1543.09 *** | 2011.95 *** | 0.015 *** | 0.003 ** |
| Repetition (B) | 2 | 0.003 | 0.036 | 0.035 | 0.036 | 0.009 | |
| Year (Y) | 21 | 1.868 *** | 0.497 *** | 0.623 *** | 0.088 *** | - | |
| Interaction P × Y | 882 | 0.023 *** | 0.007 *** | 0.007 *** | 0.001 | - | |
| Error | 2838 | 0.015 | 0.005 | 0.006 | 0.002 | 0.001 | |
| BUCOVA | Provenance (P) | 42 | 4107.42 *** | 1495.19 *** | 3182.79 *** | 0.072 *** | 0.001 |
| Repetition (B) | 2 | 0.002 | 0.234 | 0.265 | 0.244 | 0.001 | |
| Year (Y) | 21 | 1.080 *** | 0.387 *** | 0.568 *** | 0.201 *** | - | |
| Interaction P × Y | 882 | 0.024 | 0.008 | 0.012 | 0.003 | - | |
| Error | 2838 | 0.023 | 0.009 | 0.011 | 0.004 | 0.001 | |
| STRAMBU BAIUT | Provenance (P) | 42 | 3543.39 *** | 1139.49 *** | 1930.62 *** | 0.036 *** | 0.005 |
| Repetition (B) | 2 | 0.003 | 0.136 | 0.100 | 0.125 | 0.006 | |
| Year (Y) | 21 | 1.426 *** | 0.467 *** | 0.708 *** | 0.213 *** | - | |
| Interaction P × Y | 882 | 0.020 *** | 0.008 | 0.011 ** | 0.004 | - | |
| Error | 2838 | 0.014 | 0.007 | 0.010 | 0.005 | 0.004 | |
| MOINESTI | Provenance (P) | 42 | 5347 *** | 1137 *** | 2497 *** | 0.023 *** | 0.002 * |
| Repetition (B) | 2 | 0.006 | 0.304 | 0.329 | 0.289 | 0.001 | |
| Year (Y) | 21 | 1.241 *** | 0.305 *** | 0.617 *** | 0.133 *** | - | |
| Interaction P × Y | 882 | 0.013 | 0.005 | 0.009 | 0.003 | - | |
| Error | 2838 | 0.015 | 0.006 | 0.010 | 0.004 | 0.001 | |
| Source of Variation | DF | Variance (s2) | ||||
|---|---|---|---|---|---|---|
| RW | LW | EW | LWP | WD | ||
| Provenance (P) | 42 | 5894.39 *** | 1476.69 *** | 2743.90 *** | 0.032 *** | 0.003 ** |
| Site (S) | 4 | 62,418.47 *** | 44,090.87 *** | 30,738.85 *** | 1.616 *** | 0.009 *** |
| Year (Y) | 21 | 5.405 *** | 1.369 *** | 2.860 *** | 0.471 *** | - |
| Interaction P × S | 168 | 50,470.00 *** | 14,551.71 *** | 27,404.64 *** | 0.042 *** | 0.001 |
| Interaction P × Y | 882 | 0.024 | 0.008 | 0.011 | 0.004 | - |
| Interaction S × Y | 84 | 0.396 *** | 0.118 *** | 0.153 *** | 0.056 *** | - |
| Error | 13,200 | 0.023 | 0.009 | 0.013 | 0.005 | 0.001 |
| Trial | Source of Variation | DF | Variance (s2) | |||
|---|---|---|---|---|---|---|
| Res | Rec | Rsl | rRsl | |||
| SACELE | Provenance (P) | 59 | 0.119 *** | 0.100 | 0.138 *** | 0.049 |
| Repetition (B) | 2 | 0.288 | 0.622 | 0.602 | 0.209 | |
| Extreme drought year (DrY) | 3 | 2.418 *** | 4.893 *** | 4.272 *** | 2.863 *** | |
| Interaction P x DrY | 177 | 0.078 | 0.093 | 0.110 ** | 0.050 | |
| Error | 2436 | 0.068 | 0.103 | 0.079 | 0.052 | |
| DOMNESTI | Provenance (P) | 42 | 0.038 | 0.108* | 0.087 ** | 0.033 |
| Repetition (B) | 2 | 0.260 | 0.154 | 0.110 | 0.082 | |
| Extreme drought year (DrY) | 2 | 6.318 *** | 16.716 *** | 0.520 *** | 9.681 *** | |
| Interaction P x DrY | 86 | 0.080 *** | 0.119 ** | 0.066 * | 0.061 *** | |
| Error | 1368 | 0.049 | 0.081 | 0.052 | 0.034 | |
| BUCOVA | Provenance (P) | 42 | 0.074 | 0.121 | 0.071 | 0.049 |
| Repetition (B) | 2 | 0.304 | 0.858 | 0.034 | 0.148 | |
| Extreme drought year (DrY) | 3 | 3.873 *** | 8.484 *** | 0.307 *** | 4.845 *** | |
| Interaction P x DrY | 126 | 0.099 * | 0.162 | 0.089 * | 0.075 | |
| Error | 1856 | 0.081 | 0.185 | 0.073 | 0.063 | |
| STRAMBU BAIUT | Provenance (P) | 42 | 0.081 * | 0.125 | 0.081 *** | 0.063 |
| Repetition (B) | 2 | 0.330 | 0.168 | 0.342 | 0.108 | |
| Extreme drought year (DrY) | 4 | 0.911 *** | 9.110 *** | 3.391 *** | 3.856 *** | |
| Interaction P x DrY | 168 | 0.052 | 0.102 | 0.037 | 0.050 | |
| Error | 2055 | 0.061 | 0.098 | 0.042 | 0.054 | |
| MOINESTI | Provenance (P) | 42 | 0.029 | 0.048 | 0.065 ** | 0.027 |
| Repetition (B) | 2 | 0.069 | 0.111 | 0.291 | 0.076 | |
| Extreme drought year (DrY) | 2 | 4.277 *** | 6.144 *** | 0.950 *** | 6.171 *** | |
| Interaction P x DrY | 86 | 0.044 | 0.065 ** | 0.035 | 0.048 ** | |
| Error | 1368 | 0.035 | 0.043 | 0.039 | 0.030 | |
| Trial | Drought Parameters | s2 | |||
|---|---|---|---|---|---|
| 2000 | 2002/2003 (1) | 2011 | 2012 | ||
| SACELE | Resistance | 0.143 * | 0.082 | 0.066 * | 0.064 |
| Recovery | 0.123 | 0.102 | 0.082 | 0.074 | |
| Resilience | 0.207 *** | 0.085 | 0.078 * | 0.098 * | |
| Rel. resilience | 0.071 | 0.044 | 0.039 | 0.044 | |
| DOMNESTI | Resistance | 0.060 | 0.072 ** | 0.067 ** | - |
| Recovery | 0.272 * | 0.023 | 0.050 * | - | |
| Resilience | 0.109 * | 0.048 * | 0.063 * | - | |
| Rel. resilience | 0.090 ** | 0.022 | 0.042 * | - | |
| BUCOVA | Resistance | 0.203 ** | 0.049 | 0.046 | - |
| Recovery | 0.251 | 0.217 | 0.059 | - | |
| Resilience | 0.159 * | 0.072 | 0.062 | - | |
| Rel. resilience | 0.084 | 0.058 | 0.051 | - | |
| STRAMBU BAIUT | Resistance | 0.085 | 0.035 | 0.053 | 0.033 |
| Recovery | 0.174 * | 0.078 | 0.060 | 0.081 | |
| Resilience | 0.054 | 0.043 | 0.032 | 0.044 | |
| Rel. resilience | 0.090 | 0.025 | 0.042 | 0.046 | |
| MOINESTI | Resistance | 0.027 | 0.061 * | 0.028 | |
| Recovery | 0.080 ** | 0.051 | 0.049 * | ||
| Resilience | 0.041 | 0.049 | 0.047 * | ||
| Rel. resilience | 0.050 * | 0.039 | 0.035 | ||
| Trial | Trait | RW | LW | EW | LWP | WD |
|---|---|---|---|---|---|---|
| SACELE | Resistance | 0.211 | 0.301 * | 0.097 | 0.188 | 0.262 |
| Recovery | 0.077 | 0.068 | 0.063 | −0.014 | 0.065 | |
| Resilience | 0.261 * | 0.320 * | 0.144 | 0.139 | 0.145 | |
| Rel. resilience | 0.129 | 0.091 | 0.100 | −0.053 | 0.096 | |
| DOMNESTI | Resistance | 0.301 * | 0.317 * | 0.255 | 0.121 | 0.137 |
| Recovery | 0.244 | 0.206 | 0.207 | 0.014 | −0.101 | |
| Resilience | 0.389 ** | 0.362 * | 0.337 * | 0.090 | 0.052 | |
| Rel. resilience | 0.295 * | 0.214 | 0.281 | −0.038 | −0.086 | |
| BUCOVA | Resistance | 0.094 | 0.513 *** | −0.242 | 0.465 ** | 0.094 |
| Recovery | 0.131 | 0.170 | 0.030 | 0.033 | −0.167 | |
| Resilience | 0.316 * | 0.698 *** | −0.115 | 0.473 ** | −0.123 | |
| Rel. resilience | 0.255 | 0.162 | 0.181 | −0.058 | −0.243 | |
| STRAMBU BAIUT | Resistance | −0.170 | −0.150 | −0.122 | −0.084 | 0.265 |
| Recovery | 0.225 | 0.252 | 0.106 | 0.126 | 0.027 | |
| Resilience | 0.130 | 0.223 | −0.004 | 0.119 | 0.249 | |
| Rel. resilience | 0.308 * | 0.402 ** | 0.106 | 0.231 | 0.043 | |
| MOINESTI | Resistance | 0.336 * | 0.240 | 0.329 * | −0.084 | 0.106 |
| Recovery | 0.126 | −0.095 | 0.249 | −0.365 * | 0.094 | |
| Resilience | 0.298 * | 0.109 | 0.363 * | −0.263 | 0.114 | |
| Rel. resilience | 0.110 | −0.055 | 0.198 | −0.262 | 0.068 |
| Trial | Growth Response Model | Signif. | R2 | Partial R2 | |
|---|---|---|---|---|---|
| Temp. | Precip. | ||||
| Bucova | 10,632.399—41.943 MT2VEG + 0.154 MP2JAN-MAR | *** | 0.458 | 0.412 | 0.043 |
| Domnesti | 15,399.754—55.908 MT2VEG—46.317 MPOCT-MAR | *** | 0.496 | 0.475 | 0.154 |
| Moinesti | 8780.790—29.468 MT2VEG + 0.060 MP2JAN-MAR | *** | 0.372 | 0.292 | 0.017 |
| Sacele | 11,985.713—49.258 MT2VEG—41.649 MPOCT-MAR | *** | 0.415 | 0.326 | 0.093 |
| S. Baiut | 9154.895—36.635 MT2VEG + 17.794 MPOCT-MAR | *** | 0.503 | 0.326 | 0.062 |
| Trial | Growth Response Model | Signif. | R2 | Partial R2 | |
|---|---|---|---|---|---|
| Temp. | Precip. | ||||
| Bucova | 32.271 + 0.212 MAT2 + 0.027 MPVEG | *** | 0.091 | 0.092 | 0.008 |
| Domnesti | 26.927 + 0.087 MT2VEG + 0.052 MPVEG | *** | 0.222 | 0.206 | 0.111 |
| Moinesti | 30.329 + 0.069 MT2VEG − 0.002 MAP | *** | 0.136 | 0.066 | 0.006 |
| Sacele | 16.864 + 0.147 MT2VEG + 0.057 MPVEG | *** | 0.154 | 0.152 | 0.052 |
| S. Baiut | 32.411 + 0.081 MT2VEG − 0.003 MAP | *** | 0.150 | 0.058 | 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihai, G.; Alexandru, A.M.; Stoica, E.; Birsan, M.V. Intraspecific Growth Response to Drought of Abies alba in the Southeastern Carpathians. Forests 2021, 12, 387. https://doi.org/10.3390/f12040387
Mihai G, Alexandru AM, Stoica E, Birsan MV. Intraspecific Growth Response to Drought of Abies alba in the Southeastern Carpathians. Forests. 2021; 12(4):387. https://doi.org/10.3390/f12040387
Chicago/Turabian StyleMihai, Georgeta, Alin Madalin Alexandru, Emanuel Stoica, and Marius Victor Birsan. 2021. "Intraspecific Growth Response to Drought of Abies alba in the Southeastern Carpathians" Forests 12, no. 4: 387. https://doi.org/10.3390/f12040387
APA StyleMihai, G., Alexandru, A. M., Stoica, E., & Birsan, M. V. (2021). Intraspecific Growth Response to Drought of Abies alba in the Southeastern Carpathians. Forests, 12(4), 387. https://doi.org/10.3390/f12040387