
Provenance Variation in Early Survival, Growth, and Carbon Isotope Discrimination of Southwestern Ponderosa Pine Growing in Three Common Gardens across an Elevational Gradient


Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Garden Sites
2.2. Experimental Design and Plant Material
2.3. Survival and Growth Measurements
2.4. Carbon Isotopic Discrimination
2.5. Data Analyses
3. Results
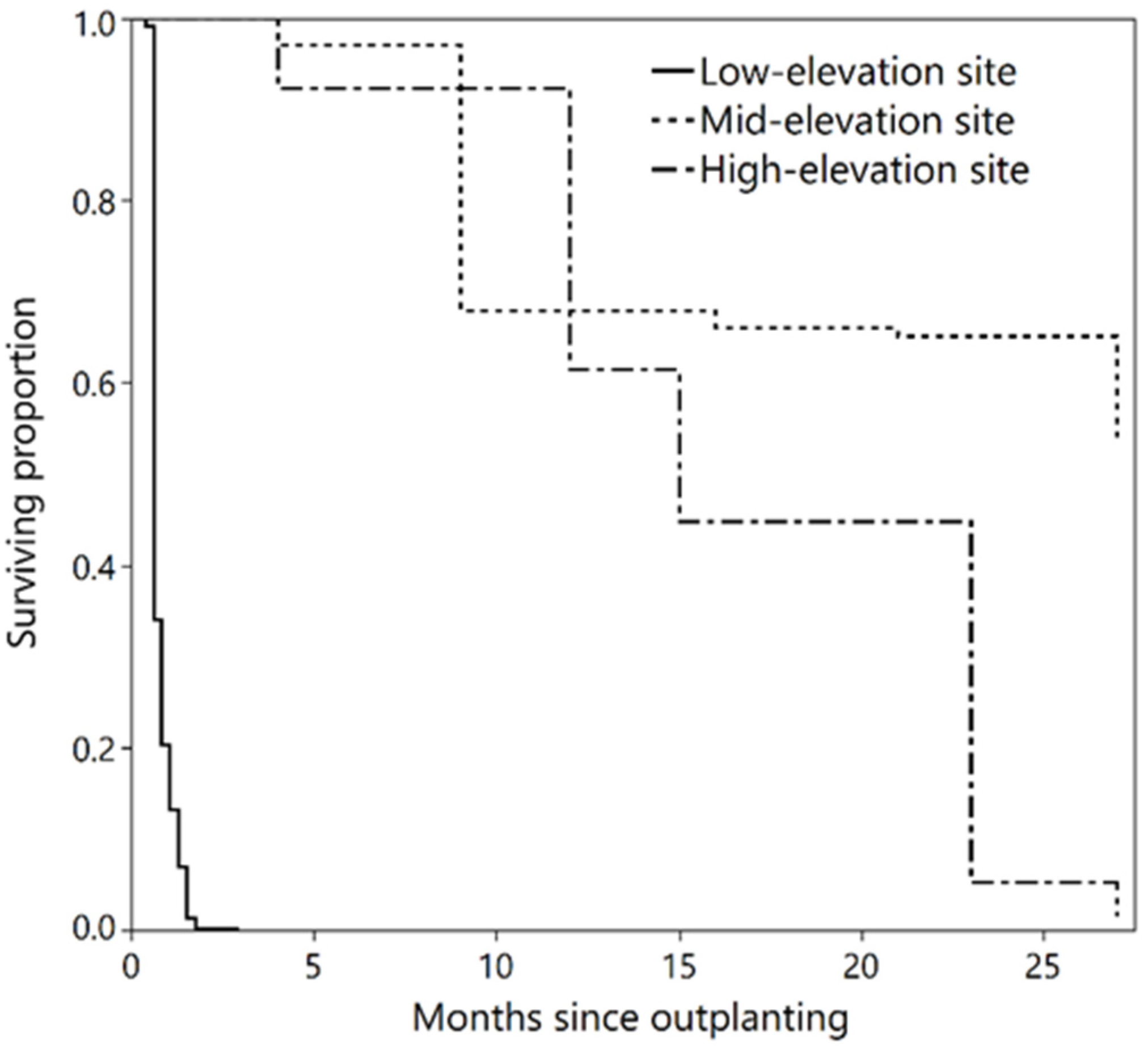
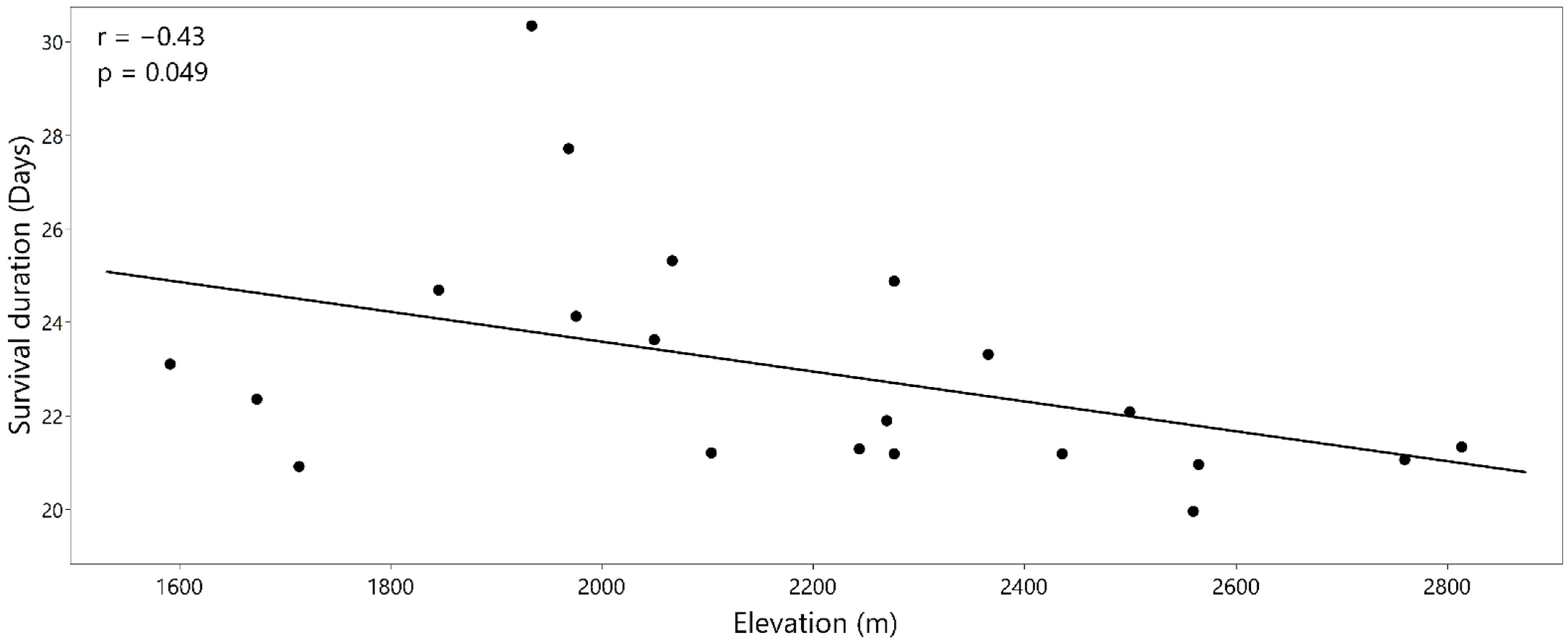
3.1. Survival Analyses
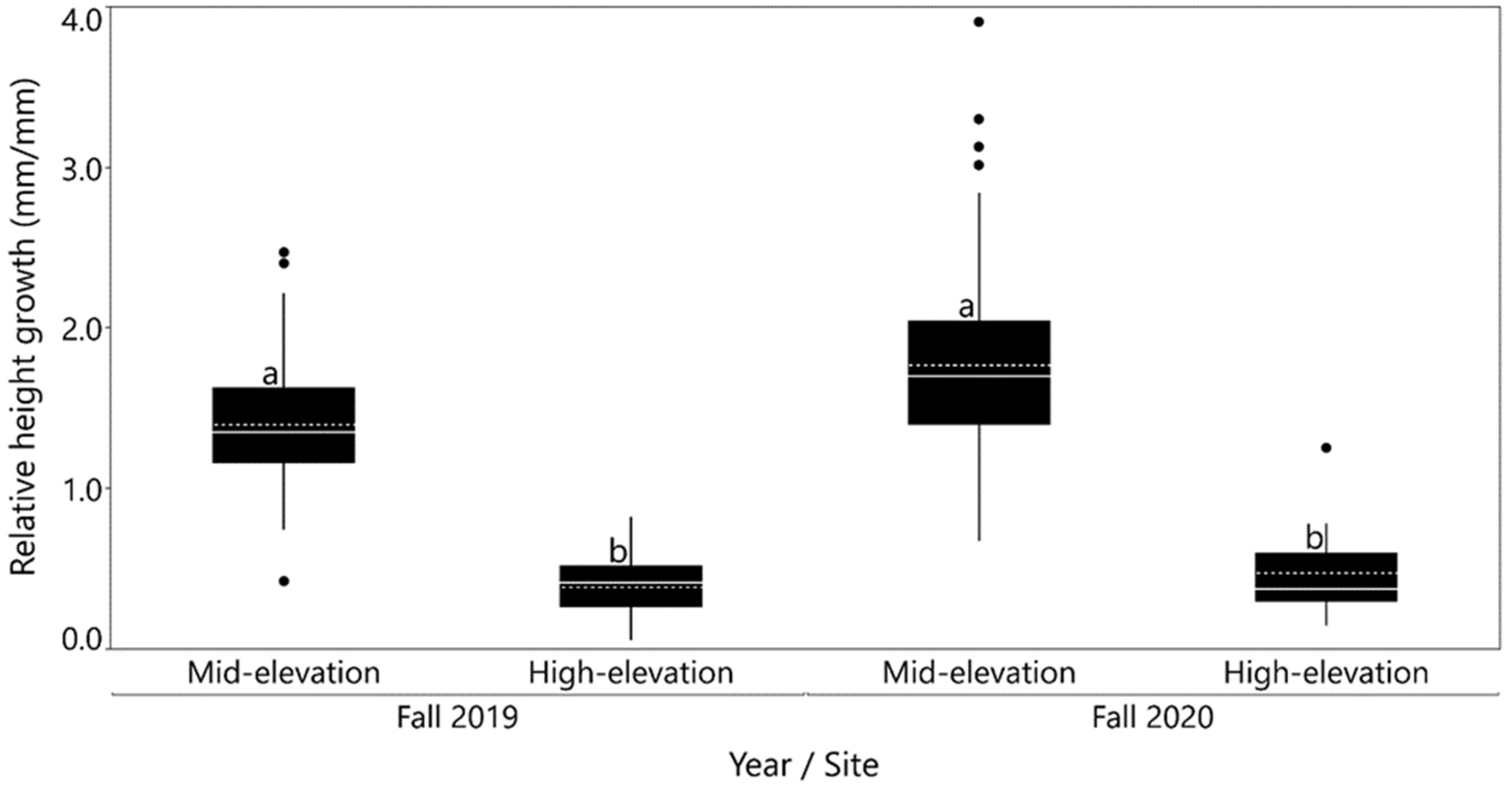
3.2. Growth
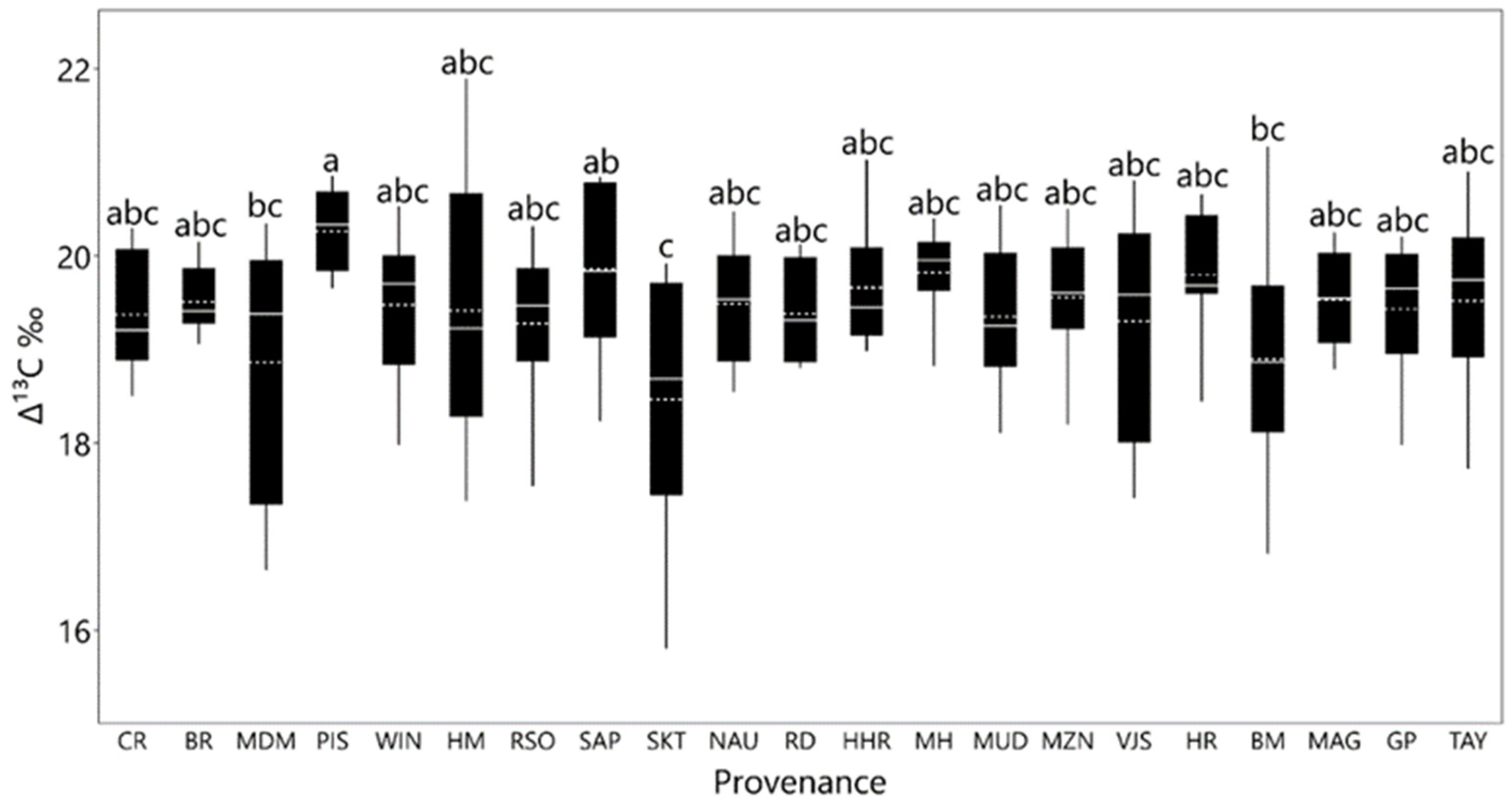
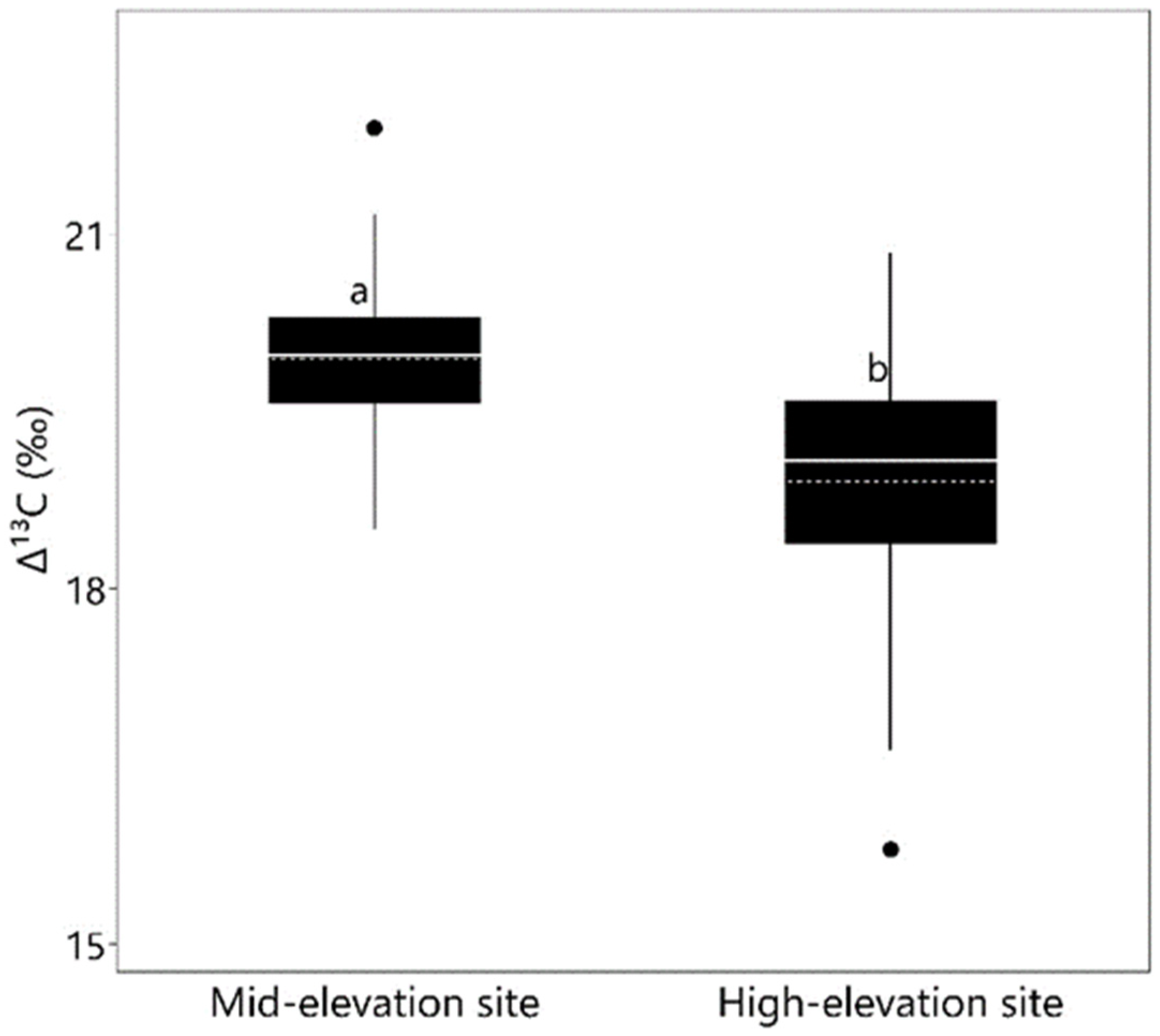
3.3. Carbon Isotope Discrimination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2012, 3, 292–297. [Google Scholar] [CrossRef]
- Hicke, J.A.; Meddens, A.J.H.; Kolden, C.A. Recent Tree Mortality in the Western United States from Bark Beetles and Forest Fires. For. Sci. 2016, 62, 141–153. [Google Scholar] [CrossRef]
- Savage, M.; Nystrom, M.J.F. Double whammy: High-severity fire and drought in ponderosa pine forests of the Southwest. Can. J. For. Res. 2013, 43, 570–583. [Google Scholar] [CrossRef]
- Rodman, K.C.; Veblen, T.T.; Chapman, T.B.; Rother, M.T.; Wion, A.P.; Redmond, M.D. Limitations to recovery following wildfire in dry forests of southern Colorado and northern New Mexico, USA. Ecol. Appl. 2020, 30, e02001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J.S. Empirical Analyses of Plant-Climate Relationships for the Western United States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D. Drought-induced shift of a forest–woodland ecotone: Rapid landscape response to climate variation. Proc. Natl. Acad. Sci. USA 1998, 95, 14839–14842. [Google Scholar] [CrossRef] [Green Version]
- Minott, J.A.; Kolb, T.E. Regeneration patterns reveal contraction of ponderosa forests and little upward migration of pinyon-juniper woodlands. For. Ecol. Manag. 2020, 458, 117640. [Google Scholar] [CrossRef]
- Petrie, M.D.; Bradford, J.B.; Hubbard, R.M.; Lauenroth, W.K.; Andrews, C.M.; Schlaepfer, D.R. Climate change may restrict dryland forest regeneration in the 21st century. Ecology 2017, 98, 1548–1559. [Google Scholar] [CrossRef]
- Kolb, T.E.; Dixit, A.H.; Burney, O. Challenges and opportunities for maintaining ponderosa pine forests in the southwestern United States. Tree Plant. Note 2019, 62, 104–112. [Google Scholar]
- Fargione, J.; Haase, D.L.; Burney, O.T.; Kildisheva, O.A.; Edge, G.; Cook-Patton, S.C.; Chapman, T.; Rempel, A.; Hurteau, M.D.; Davis, K.T.; et al. Challenges to the Reforestation Pipeline in the United States. Front. For. Glob. Chang. 2021, 4, 8. [Google Scholar] [CrossRef]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Jaquish, B.C.; Sáenz-Romero, C.; Joyce, D.G.; Leites, L.P.; Bradley St Clair, J.; López-Upton, J. Comparative genetic responses to climate in the varieties of Pinus ponderosa and Pseudotsuga menziesii: Reforestation. For. Ecol. Manag. 2014, 1324, 47–157. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.; Ronce, O.; Robledo-Arnuncio, J.J.; Guillaume, F.; Bohrer, G.; Nathan, R.; Bridle, J.R.; Gomulkiewicz, R.; Klein, E.K.; Ritland, K.; et al. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garfin, A.; Jardine, A.; Merideth, R.; Black, M.; LeRoy, S. Assessment of Climate Change in the Southwest United States: A Report Prepared for the National Climate Assessment; Island Press: Washington, DC, USA, 2013; 531p. [Google Scholar]
- Grady, K.C.; Kolb, T.E.; Ikeda, D.H.; Whitham, T.G. A bridge too far: Cold and pathogen constraints to assisted migration of riparian forests. Restor. Ecol. 2015, 23, 811–820. [Google Scholar] [CrossRef]
- Bucharova, A. Assisted migration within species range ignores biotic interactions and lacks evidence. Restor. Ecol. 2017, 25, 14–18. [Google Scholar] [CrossRef]
- Pearson, G.A. Management of Ponderosa Pine in the Southwest; Agriculture Monograph No. 6; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1950; 218p.
- Schubert, G.H.; Heidmann, L.J.; Larson, M.M. Artificial Reforestation Practices for the Southwest; U.S. Department of Agriculture, Handbook: Washington, DC, USA, 1970; Volume 370, 25p.
- Kolb, T.E.; Flathers, K.; Bradford, J.B.; Andrews, C.; Asherin, L.A.; Moser, W.K. Stand density, drought, and herbivory constrain ponderosa pine regeneration pulse. Can. J. For. Res. 2020, 50, 862–871. [Google Scholar] [CrossRef]
- Rehfeldt, G.E. Genetic Variation in the Ponderosae of the Southwest. Am. J. Bot. 1993, 80, 330–343. [Google Scholar] [CrossRef]
- Dixit, A.; Kolb, T. Variation in seedling budburst phenology and structural traits among southwestern ponderosa pine provenances. Can. J. For. Research 2020, 50, 872–879. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change—Evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- Kolb, T.E.; Grady, K.C.; McEttrick, M.P.; Herrero, A. Local-Scale Drought Adaptation of Ponderosa Pine Seedlings at Habitat Ecotones. For. Sci. 2016, 62, 641–651. [Google Scholar] [CrossRef]
- Kerr, K.L.; Meinzer, F.C.; McCulloh, K.A.; Woodruff, D.R.; Marias, D.E. Expression of functional traits during seedling establishment in two populations of Pinus ponderosa from contrasting climates. Tree Physiol. 2015, 35, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Moran, E.; Lauder, J.; Musser, C.; Stathos, A.; Shu, M. The genetics of drought tolerance in conifers. New Phytol. 2017, 216, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the Relationship Between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Funct. Plant Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Cregg, B.M.; Olivas-García, J.M.; Hennessey, T.C. Provenance variation in carbon isotope discrimination of mature ponderosa pine trees at two locations in the Great Plains. Can. J. For. Res. 2000, 30, 428–439. [Google Scholar] [CrossRef]
- Olivas-García, J.M.; Cregg, B.M.; Hennessey, T.C. Genotypic variation in carbon isotope discrimination and gas exchange of ponderosa pine seedlings under two levels of water stress. Can. J. For. Res. 2000, 30, 10. [Google Scholar] [CrossRef]
- Howe, A.A.; Landhäusser, S.M.; Burney, O.T.; Long, J.N.; Violett, R.D.; Mock, K.E. Exploring seedling-based aspen (Populus tremuloides) restoration near range limits in the Intermountain West, USA. For. Ecol. Manag. 2020, 476, 118470. [Google Scholar] [CrossRef]
- Dixit, A.; Kolb, T.; Burney, O. Provenance Geographical and Climatic Characteristics Influence Budburst Phenology of Southwestern Ponderosa Pine Seedlings. Forests 2020, 11, 1067. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Ratner, B. The correlation coefficient: Its values range between +1/−1, or do they? J. Target Meas. Anal. Mark. 2009, 17, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Berdeja, A.; Hamilton, J.A.; Bontemps, A.; Schmitt, J.; Wright, J.W. Evidence for population differentiation among Jeffrey and Ponderosa pines in survival, growth and phenology. For. Ecol. Manag. 2019, 434, 40–48. [Google Scholar] [CrossRef]
- Burney, O.T.; Jacobs, D.F. Ungulate herbivory of boreal and temperate forest regeneration in relation to seedling mineral nutrition and secondary metabolites. New For. 2013, 44, 753–768. [Google Scholar] [CrossRef]
- Shepperd, W.D.; Mata, S.A. Planting Aspen to Rehabilitate Riparian Areas: A Pilot Study; United States Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005.
- Howe, A.A. Assessment of a Seedling-Based Approach to Aspen Restoration in the Intermountain West. Ph.D. Thesis, Utah State University, Logan, UT, USA, 2018. Available online: https://digitalcommons.usu.edu/etd/7062 (accessed on 4 November 2021).
- Oliet, J.A.; Blasco, R.; Valenzuela, P.; de Blas, M.M.; Puértolas, J. Should we use meshes or solid tube shelters when planting in Mediterranean semiarid environments? New For. 2019, 50, 267–282. [Google Scholar] [CrossRef]
- Engemann, R.M.; Anthony, R.M.; Barnes, V.G., Jr.; Krupa, H.W.; Evans, J. Evaluations of plastic mesh tubes for protecting conifer seedlings from pocket gophers in three western States. West. J. Appl. For. 1999, 14, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.S.; Loewenstein, E.F.; Chappelka, A.H. Effect of animal browse protection and fertilizer application on the establishment of planted Nuttall oak seedlings. New For. 2006, 32, 133–143. [Google Scholar] [CrossRef]
- Hultine, K.R.; Marshall, J.D. Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia 2000, 123, 32–40. [Google Scholar] [CrossRef]
- Zhang, J.; Cregg, B.M. Growth and physiological responses to varied environments among populations of Pinus ponderosa. For. Ecol. Manag. 2005, 219, 1–12. [Google Scholar] [CrossRef]
- Soolanayakanahally, R.Y.; Guy, R.D.; Silim, S.N.; Drewes, E.C.; Schroeder, W.R. Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L.). Plant Cell Environ. 2009, 32, 1821–1832. [Google Scholar] [CrossRef]
- Goodrich, B.A.; Waring, K.M.; Kolb, T.E. Genetic variation in Pinus strobiformis growth and drought tolerance from southwestern US populations. Tree Physiol. 2016, 36, 1219–1235. [Google Scholar] [CrossRef]
- Willyard, A.; Gernandt, D.S.; Potter, K.; Hipkins, V.; Marquardt, P.; Mahalovich, M.F.; Langer, S.K.; Telewski, F.W.; Cooper, B.; Douglas, C.; et al. Pinus ponderosa: A checkered past obscured four species. Am. J. Bot. 2017, 104, 161–181. [Google Scholar] [CrossRef] [Green Version]
- Sáenz-Romero, C.; O’Neill, G.; Aitken, S.N.; Lindig-Cisneros, R. Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges. Forests 2021, 12, 9. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latitude/Longitude | Elevation (m) | MAT (°C) | MAP (mm) | |
|---|---|---|---|---|
| Provenance (code) | ||||
| Cherry Road (CR) | 34.586/–112.057 | 1592 | 14.2 | 408 |
| Blue River (BR) | 33.555/–109.193 | 1674 | 11.8 | 581 |
| Mesa Del Medio (MDM) | 35.116/–105.217 | 1714 | 12.3 | 397 |
| Prescott—Iron Springs Road (PIS) | 34.585/–112.559 | 1846 | 11.0 | 546 |
| Townsend Winona (WIN) | 35.254/–111.415 | 1934 | 10.2 | 398 |
| Hualapai Mountains (HM) | 35.084/–113.875 | 1969 | 11.5 | 403 |
| Ruidoso Service Office (RSO) | 33.350/–105.583 | 1976 | 11.1 | 506 |
| Sapello Rt. 3 (SAP) | 35.700/–105.250 | 2050 | 9.8 | 456 |
| South Kaibab Tusayan Dist. (SKT) | 35.939/–112.084 | 2067 | 8.9 | 410 |
| Northern Arizona University (NAU) | 35.182/–111.655 | 2104 | 8.4 | 540 |
| Rim District (RD) | 34.487/–111.343 | 2244 | 9.4 | 767 |
| HH Ranch (HHR) | 34.183/–107.525 | 2270 | 9.2 | 364 |
| Mineral Hill (MH) | 35.633/–105.461 | 2277 | 8.5 | 528 |
| Mud Springs (MUD) | 36.463/–106.859 | 2277 | 6.9 | 443 |
| Manzano Mountains (MZN) | 34.623/–106.400 | 2366 | 8.6 | 638 |
| Vallecitos-Jemez Springs (VJS) | 35.809/–106.589 | 2436 | 6.8 | 571 |
| Hartman Ridge (HR) | 35.550/–105.533 | 2500 | 8.8 | 526 |
| Borrego Mesa (BM) | 35.990/–105.794 | 2560 | 6.3 | 470 |
| Magdalena Mountains (MAG) | 34.006/–107.215 | 2565 | 8.9 | 512 |
| Green’s Peak (GP) | 34.126/–109.535 | 2760 | 5.7 | 671 |
| Mount Taylor (TAY) | 35.266/–107.633 | 2814 | 6.1 | 722 |
| Common Garden Site | ||||
| Low-elevation | 35.586/–111.969 | 1930 | 9.7 | 482 |
| Mid-elevation | 35.160/–111.730 | 2200 | 7.6 | 556 |
| High-elevation | 37.620/–113.025 | 2780 | 4.9 | 685 |
| Provenance | Fall 2019 Relative Height Growth (mm/mm) | Fall 2019 GLD (mm) |
|---|---|---|
| Mid-elevation site | ||
| CR | 1.0 (0.1) a | 7.66 (0.41) a |
| BR | 1.3 (0.2) a | 7.96 (0.45) a |
| MDM | 1.2 (0.0) a | 6.24 (0.06) a |
| PIS | 1.4 (0.0) a | 7.27 (0.23) a |
| WIN | 1.3 (0.0) a | 7.53 (0.45) a |
| HM | 1.5 (0.1) a | 7.00 (0.31) a |
| RSO | 1.4 (0.2) a | 7.38 (0.33) a |
| SAP | 1.4 (0.1) a | 7.21 (0.21) a |
| SKT | 1.0 (0.0) a | 7.80 (0.23) a |
| NAU | 1.1 (0.1) a | 7.75 (0.34) a |
| RD | 1.6 (0.1) a | 8.46 (0.45) a |
| HHR | 1.2 (0.0) a | 7.49 (0.20) a |
| MH | 1.5 (0.1) a | 7.55 (0.46) a |
| MUD | 1.4 (0.2) a | 6.50 (0.25) a |
| MZN | 1.2 (0.2) a | 7.86 (0.36) a |
| VJS | 1.3 (0.1) a | 7.90 (0.24) a |
| HR | 1.5 (0.2) a | 7.80 (0.22) a |
| BM | 1.6 (0.2) a | 7.41 (0.32) a |
| MAG | 1.1 (0.1) a | 6.40 (0.52) a |
| GP | 1.7 (0.3) a | 7.45 (0.58) a |
| TAY | 1.7 (0.1) a | 7.19 (0.19) a |
| High-elevation site | ||
| CR | 0.08 (NA) a | 5.15 (NA) ab |
| BR | 0.3 (0.0) a | 5.32 (0.01) ab |
| MDM | 0.3 (0.1) a | 4.46 (0.20) ab |
| PIS | 0.4 (0.0) a | 5.39 (0.26) ab |
| WIN | 0.5 (0.0) a | 5.58 (0.59) ab |
| HM | 0.3 (0.1) a | 5.10 (0.09) ab |
| RSO | 0.5 (0.2) a | 6.01 (0.07) a |
| SAP | 0.4 (0.0) a | 5.05 (0.84) ab |
| SKT | 0.4 (NA) a | 5.32 (NA) ab |
| NAU | 0.3 (0.0) a | 5.75 (0.22) ab |
| RD | 0.2 (NA) a | 6.42 (NA) a |
| HHR | 0.3 (NA) a | 3.80 (NA) ab |
| MH | 0.3 (NA) a | 4.58 (NA) ab |
| MUD | 0.2 (NA) a | 3.90 (NA) ab |
| MZN | 0.3 (0.0) a | 5.70 (0.12) ab |
| VJS | 0.6 (NA) a | 4.72 (NA) ab |
| HR | 0.2 (0.0) a | 4.14 (0.56) b |
| BM | 0.4 (0.0) a | 4.50 (0.05) ab |
| MAG | 0.0 (NA) a | 5.80 (NA) ab |
| GP | 0.6 (NA) a | 6.45 (NA) a |
| TAY | 0.3 (0.2) a | 5.60 (0.40) ab |
| Provenance | Site | Provenance × Site | |
|---|---|---|---|
| 2019 relative height growth | 0.926 | <0.0001 | 0.982 |
| 2019 GLD | 0.008 | 0.001 | 0.143 |
| ∆13C | 0.006 | 0.0001 | 0.239 |
| Latitude | Longitude | Elevation | MAT | MAP | |
|---|---|---|---|---|---|
| Mid-elevation site | |||||
| 2019 relative height growth (mm/mm) | 0.09 (0.668) | 0.13 (0.561) | 0.56 (0.007) | −0.51 (0.015) | 0.54 (0.010) |
| 2019 GLD (mm) | −0.12 (0.586) | −0.26 (0.244) | 0.001 (0.993) | −0.04 (0.843) | 0.43 (0.051) |
| ∆13C (‰) | 0.32 (0.154) | −0.05 (0.806) | −0.001 (0.996) | 0.03 (0.864) | −0.09 (0.683) |
| High-elevation site | |||||
| 2019 relative height growth (mm/mm) | 0.09 (0.708) | 0.007 (0.973) | 0.10 (0.657) | −0.30 (0.175) | 0.04 (0.850) |
| 2019 GLD (mm) | −0.53 (0.014) | −0.40 (0.066) | 0.06 (0.789) | 0.01 (0.960) | 0.60 (0.004) |
| ∆13C (‰) | −0.42 (0.054) | 0.01 (0.942) | 0.02 (0.936) | 0.09 (0.699) | 0.31 (0.161) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dixit, A.; Kolb, T.; Burney, O.; Mock, K.; Grady, K. Provenance Variation in Early Survival, Growth, and Carbon Isotope Discrimination of Southwestern Ponderosa Pine Growing in Three Common Gardens across an Elevational Gradient. Forests 2021, 12, 1561. https://doi.org/10.3390/f12111561
Dixit A, Kolb T, Burney O, Mock K, Grady K. Provenance Variation in Early Survival, Growth, and Carbon Isotope Discrimination of Southwestern Ponderosa Pine Growing in Three Common Gardens across an Elevational Gradient. Forests. 2021; 12(11):1561. https://doi.org/10.3390/f12111561
Chicago/Turabian StyleDixit, Aalap, Thomas Kolb, Owen Burney, Karen Mock, and Kevin Grady. 2021. "Provenance Variation in Early Survival, Growth, and Carbon Isotope Discrimination of Southwestern Ponderosa Pine Growing in Three Common Gardens across an Elevational Gradient" Forests 12, no. 11: 1561. https://doi.org/10.3390/f12111561
APA StyleDixit, A., Kolb, T., Burney, O., Mock, K., & Grady, K. (2021). Provenance Variation in Early Survival, Growth, and Carbon Isotope Discrimination of Southwestern Ponderosa Pine Growing in Three Common Gardens across an Elevational Gradient. Forests, 12(11), 1561. https://doi.org/10.3390/f12111561