Eutypella parasitica and Other Frequently Isolated Fungi in Wood of Dead Branches of Young Sycamore Maple (Acer pseudoplatanus) in Slovenia



Abstract
1. Introduction
2. Materials and Methods
2.1. Definitions of Repeatedly Used Terms
- Sampling site—a site or an area in the forest stand where samples were collected
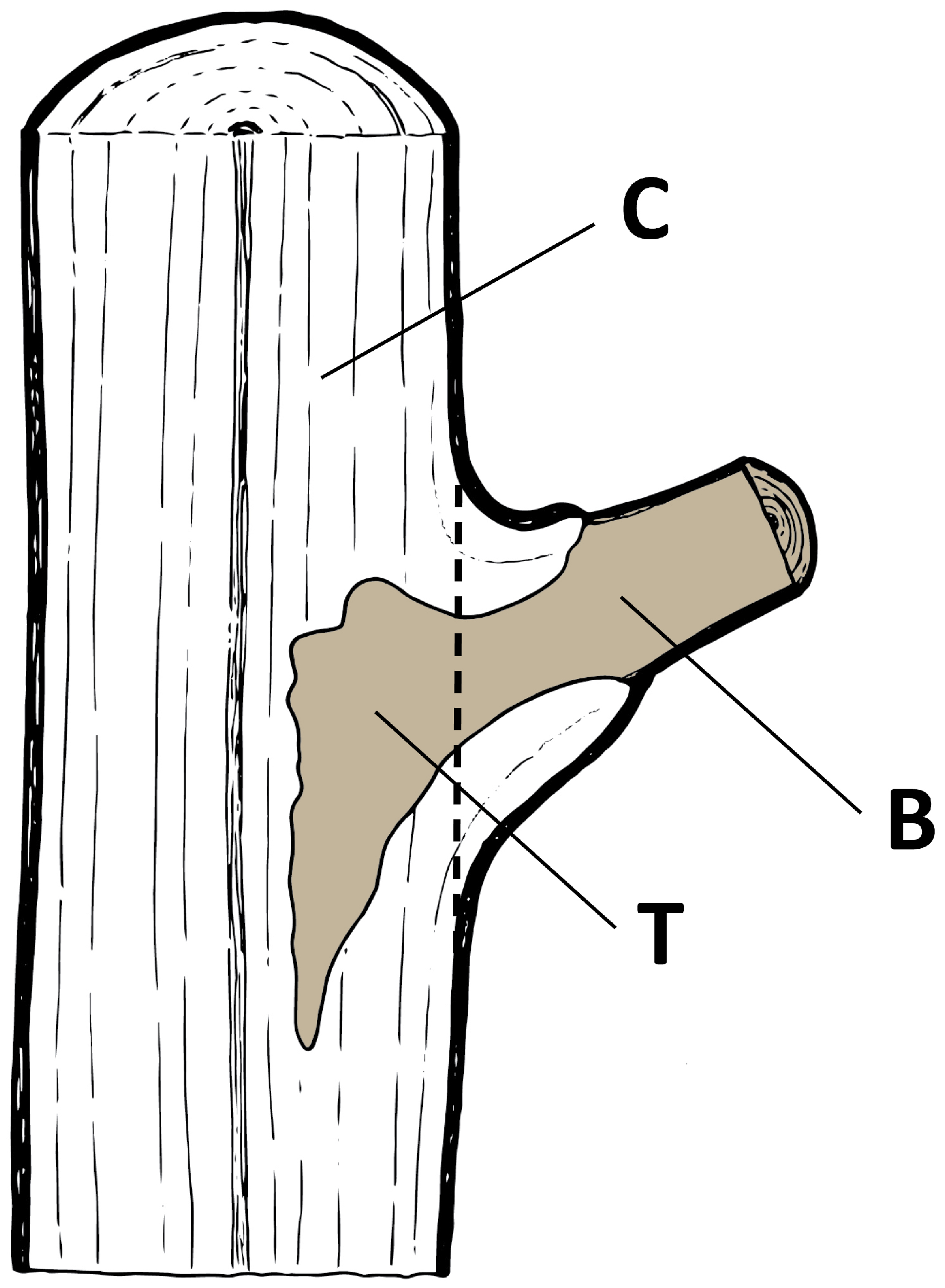
- Sample—a dead A. pseudoplatanus branch with a section of the trunk where it was attached (Figure 1)
- Isolation source—a location in a sample from which subsamples were cut (B—branch; T—trunk; C—control) (Figure 1)
- Subsample—a small piece of approximately 1 × 2 × 2 mm cut from the wood and representing three isolation sources (if possible) in each sample
- Culture—an outgrown mycelium from a subsample
2.2. Inventory of Eutypella Canker of Maple
2.3. Sampling
2.4. Isolation of Fungi
2.5. DNA Extraction, Amplification and Sequencing
2.6. Data Analysis
3. Results
3.1. Eutypella Parasitica
3.2. Colonization Rate, Relative Colonization Frequency and Density Index
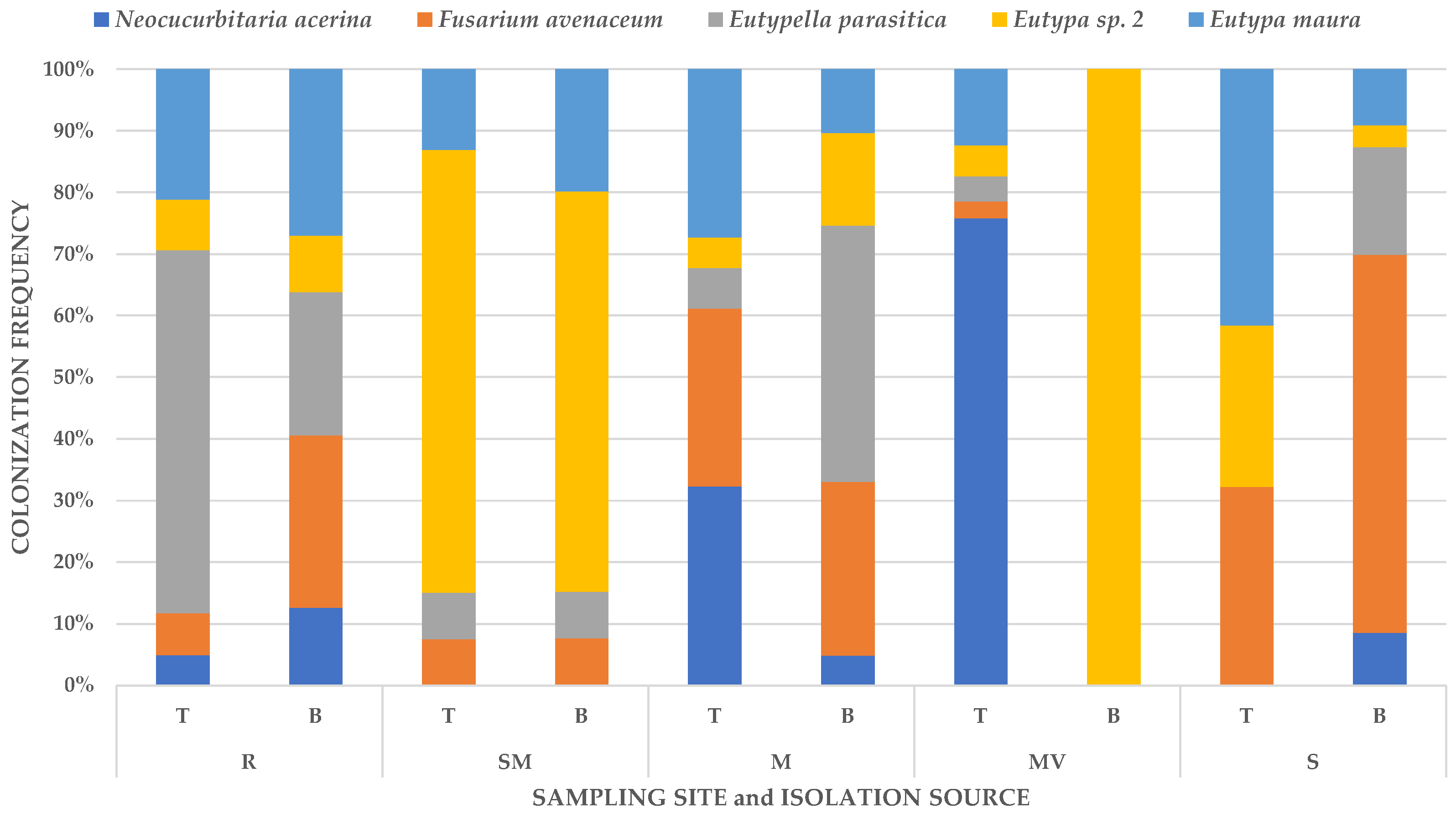
3.3. Community Composition
3.4. Species Diversity
3.5. Branch Diameter and Species Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spiecker, H.; Hein, S.; Makkonen-Spiecker, K.; Thies, M. Valuable Broadleaved Forests in Europe; Brill: Leiden, The Netherlands; Boston, MA, USA, 2009; p. 256. [Google Scholar]
- Rusanen, M.; Myking, T. Euforgen Technical Guidelines for Genetic Conservation and Use for Sycamore (Acer pseudoplatanus); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Hein, S. Distribution of valuable broadleaved forests in Europe, Appendix B. In Valuable Broadleaved Forests in Europe; Spiecker, H., Hein, S., Makkonen-Spiecker, K., Thies, M., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2009; pp. 251–256. [Google Scholar]
- Hein, S.; Collet, C.; Ammer, C.; Le Goff, N.; Skovsgaard, J.P.; Savill, P. A review of growth and stand dynamics of Acer pseudoplatanus L. in Europe: Implications for silviculture. Forestry 2008, 82, 361–385. [Google Scholar] [CrossRef]
- Brus, R. Dendrology for Foresters, 2nd ed.; University of Ljubljana, Biotechnical Faculty, Department of Forestry and Renewable Forest Resources: Ljubljana, Slovenia, 2008; p. 408. [Google Scholar]
- Savill, P. Future Prospects for the Production of Timber from Valuable Broadleaves. In Valuable Broadleaved Forests in Europe; Spiecker, H., Hein, S., Makkonen-Spiecker, K., Thies, M., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2009; pp. 11–26. [Google Scholar]
- Spiecker, H. Increasing Interest in Valuable Broadleaved Tree Species. In Valuable Broadleaved Forests in Europe; Spiecker, H., Hein, S., Makkonen-Spiecker, K., Thies, M., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2009; pp. 3–6. [Google Scholar]
- ZGS. Forest Funds Database, 2018; Slovenia Forest Service. 2019. Available online: https://www.stat.si/StatWeb/en/News/Index/8384 (accessed on 27 September 2019).
- Wulf, A.; Kehr, R. Diseases, Disorders and Pests of Selected Valuable Broadleaved Tree Species. In Valuable Broadleaved Forests in Europe; Spiecker, H., Hein, S., Makkonen-Spiecker, K., Thies, M., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2009; pp. 61–84. [Google Scholar]
- Jurc, D.; Ogris, N.; Slippers, B.; Stenlid, J. First report of Eutypella canker of Acer pseudoplatanus in Europe. Plant Pathol. 2006, 55, 577. [Google Scholar] [CrossRef]
- Cech, T.L. Erstnachweis von Eutypella parasitica in Österreich. Forstsch. Aktuell 2007, 40, 10–13. [Google Scholar]
- Ogris, N.; Diminić, D.; Piškur, B.; Kraigher, H. First report of Eutypella parasitica causing cankers on field maple (Acer campestre) in Croatia. Plant Pathol. 2008, 57, 785. [Google Scholar] [CrossRef]
- Cech, T.L.; Schwanda, K.; Klosterhuber, M.; Straßer, L.; Kirisits, T. Eutypella canker of maple: First report from Germany and situation in Austria. For. Pathol. 2016, 46, 336–340. [Google Scholar] [CrossRef]
- Jurc, D.; Ogris, N.; Piškur, B.; Csóka, G. First report of Eutypella canker of maple (Eutypella parasitica) in Hungary. Plant Dis. 2016, 100, 1241. [Google Scholar] [CrossRef]
- Černý, K.; Hrabětová, M.; Svobodová, I.; Mrázková, M.; Kowalski, T. Eutypella parasitica naturalised in Bohemian and Polish Silesia. For. Pathol. 2017, 47, e12347. [Google Scholar] [CrossRef]
- Bregant, C. Eutypella parasitica: Primo Rinvenimento e Monitoraggio in Fruili Venezia Giulia. MSc Thesis, Università degli Studi di Padova, Legnaro, Italy, 2018. [Google Scholar]
- Jurc, D.; Piškur, B.; Ogris, N.; Brglez, A.; Linaldeddu, B.T.; Bregant, C.; Montecchio, L. First Report of Eutypella Canker Caused by Eutypella parasitica on Acer campestre in Italy. Plant Dis. 2020, 104. [Google Scholar] [CrossRef]
- Davidson, R.W.; Lorenz, R.C. Species of Eutypella and Schizoxylon associated with cankers of maple. Phytopathology 1938, 28, 733–745. [Google Scholar]
- Ogris, N.; Jurc, D.; Jurc, M. Spread risk of Eutypella canker of maple in Europe. EPPO Bull. 2006, 36, 475–485. [Google Scholar] [CrossRef]
- French, W.J. Eutypella Canker on Species of Acer in New York State. Ph.D. Thesis, State University College of Forestry at Syracuse University, Syracuse, New York, NY, USA, 1967. [Google Scholar]
- Ogris, N.; Piškur, B.; Jurc, D. Some morphological aspects of Eutypella canker of maple (Eutypella parasitica). In Proceedings of the IUFRO Working Party 7.02.02, Eğirdir, Turkey, 11–16 May 2009; pp. 150–161. [Google Scholar]
- Lachance, D. Discharge and germination of Eutypella parasitica ascospores. Can. J. Bot. 1971, 49, 1111–1118. [Google Scholar] [CrossRef]
- EPPO. Mini Data Sheet on Eutypella parasitica, 2008/028 ed.; European and Mediterranean Plant Protection Organization: Paris, France, 2008; p. 2. [Google Scholar]
- Johnson, D.W. Eutypella Canker of Maple: Ascospore Discharge and Dissemination. Phytopathology 1979, 69, 130. [Google Scholar] [CrossRef]
- Butin, H.; Kowalski, T. Die natürliche Astreinigung und ihre biologischen Voraussetzungen, III. Die Pilzflora von Ahorn, Erle, Birke, Hainbuche und Esche. For. Pathol. 1986, 16, 129–138. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Boil. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Chlebicki, A. Some ascomycetous fungi or their anamorphs occurring on trees in Poland. I. Acta Mycol. 2014, 24, 77–92. [Google Scholar] [CrossRef][Green Version]
- Kowalski, T.; Kehr, R.D. Endophytic fungal colonization of branch bases in several forest tree species. Sydowia 1992, 44, 137–168. [Google Scholar]
- Ellis, M.B.; Ellis, J.P. Microfungi on Land Plants: An Identification Handbook; The Richmond Publishing Co. Ltd.: Slough, UK, 1997; p. 868. [Google Scholar]
- Unterseher, M.; Otto, P.; Morawetz, W. Species richness and substrate specificity of lignicolous fungi in the canopy of a temperate, mixed deciduous forest. Mycol. Prog. 2005, 4, 117–132. [Google Scholar] [CrossRef]
- Johnová, M. Diversity and ecology of selected lignicolous Ascomycetes in the Bohemian Switzerland National Park (Czech Republic). Czech Mycol. 2009, 61, 81–97. [Google Scholar] [CrossRef]
- Yang, D.-Q. Isolation of wood-inhabiting fungi from Canadian hardwood logs. Can. J. Microbiol. 2005, 51, 1–6. [Google Scholar] [CrossRef]
- Qi, F.-H.; Jing, T.-Z.; Wang, Z.-X.; Zhan, Y.-G. Fungal endophytes from Acer ginnala Maxim: Isolation, identification and their yield of gallic acid. Lett. Appl. Microbiol. 2009, 49, 98–104. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.-D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 2011, 47, 85–95. [Google Scholar] [CrossRef]
- Green, D.J.; Shortle, W.C.; Shigo, A.L. Compartmentalization of Discolored and Decayed Wood in Red Maple Branch Stubs. Forest Sci. 1981, 27, 519–522. [Google Scholar]
- ARSO. Monthly and Yearly Average Temperature in Slovenia for the Period from 1981 to 2010: 12 Monthly and One Yearly Map in Raster Format with 1 km Resolution; Slovenian Environment Agency, Meteorology Office: Ljubljana, Slovene, 2012. [Google Scholar]
- ARSO. Monthly and Yearly Average Sum of Corrected Precipitations in Slovenia for the Period from 1981 to 2010: 12 Monthly and One Yearly Map in Raster Format with 1 km Resolution; Slovenian Environment Agency, Meteorology Office: Ljubljana, Slovene, 2012. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Petrini, O.; Stone, J.; Carroll, F.E. Endophytic fungi in evergreen shrubs in western Oregon: A preliminary study. Can. J. Bot. 1982, 60, 789–796. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing Company: Hoboken, NJ, USA, 2004; p. 256. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, version 3.6.1; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solmyos, P.; et al. Vegan: Community Ecology Package. R package version 2.5–5. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 28 January 2020).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases. Available online: https://nt.ars-grin.gov/fungaldatabases/index.cfm (accessed on 18 February 2020).
- Wu, B.; Hussain, M.; Zhang, W.; Stadler, M.; Liu, X.; Xiang, M. Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef]
- Danti, R.; Sieber, T.N.; Sanguineti, G. Endophytic mycobiota in bark of European beech (Fagus sylvatica) in the Apennines. Mycol. Res. 2002, 106, 1343–1348. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Herrera, C.S.; Rossman, A.Y.; Samuels, G.J.; Chaverri, P. Pseudocosmospora, a new genus to accommodate Cosmospora vilior and related species. Mycologia 2013, 105, 1287–1305. [Google Scholar] [CrossRef]
- Herrera, C.S. Systematics of the Genus Cosmospora (Nectriaceae, Hypocreales), and Cospeciation of Cosmospora Species. Ph.D. Thesis, Faculty of the Graduate School of the University of Maryland, College Park, MD, USA, 2014. [Google Scholar]
- Glawe, D.A. Bloxamia truncata in Artificial Culture. Mycologia 1984, 76, 741–745. [Google Scholar] [CrossRef]
- Gross, H.L. Impact of Eutypella Canker on the Maple Resource of the Owen Sound and Wingham Forest Districts. For. Chron. 1984, 60, 18–21. [Google Scholar] [CrossRef]
- Unterseher, M.; Tal, O. Influence of small scale conditions on the diversity of wood decay fungi in a temperate, mixed deciduous forest canopy. Mycol. Res. 2006, 110, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Trouillas, F.P.; Gubler, W.D. Host Range, Biological Variation, and Phylogenetic Diversity of Eutypa lata in California. Phytopathology 2010, 100, 1048–1056. [Google Scholar] [CrossRef]
- Wenneker, M.; Van Raak, M.M.J.P.; Van Brouwershaven, I.R.; Martin, W.; Kox, L.F.F. Eutypa lata, the causal agent of dieback in red currant (Ribes rubrum) and gooseberry (R. uva-crispa) in the Netherlands. Eur. J. Plant Pathol. 2011, 131, 441–449. [Google Scholar] [CrossRef]
- Travadon, R.; Baumgartner, K.; Rolshausen, P.E.; Gubler, W.D.; Sosnowski, M.R.; LeComte, P.; Halleen, F.; Péros, J.-P. Genetic structure of the fungal grapevine pathogen Eutypa lata from four continents. Plant Pathol. 2011, 61, 85–95. [Google Scholar] [CrossRef]
- Rolshausen, P.E.; Greve, L.C.; Labavitch, J.M.; Mahoney, N.E.; Molyneux, R.J.; Gubler, W.D. Pathogenesis of Eutypa lata in Grapevine: Identification of Virulence Factors and Biochemical Characterization of Cordon Dieback. Phytopathology 2008, 98, 222–229. [Google Scholar] [CrossRef]
- Rappaz, F. Taxonomie et nomenclature des diatrypacées à aseas octospores. Mycol. Helvetica 1987, 2, 285–648. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org (accessed on 2 September 2019).
- Gennaro, M.; Gonthier, P.; Nicolotti, G. Fungal Endophytic Communities in Healthy and Declining Quercus robur L. and Q. cerris L. Trees in Northern Italy. J. Phytopathol. 2003, 151, 529–534. [Google Scholar] [CrossRef]
- Hanácková, Z.; Havrdová, L.; Černý, L.; Zahradník, D.; Koukol, O. Fungal Endophytes in Ash Shoots-Diversity and Inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Sun, X.; Ding, Q.; Hyde, K.D.; Guo, L.-D. Community structure and preference of endophytic fungi of three woody plants in a mixed forest. Fungal Ecol. 2012, 5, 624–632. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolation Source | Fungal Taxon | |
|---|---|---|
| Butin and Kowalski [25] 1 | dead twigs | Aposphaeria sp. Berk., Diaporthe acerina (Peck) Sacc., Durella atrocyanea (Fr.) Höhn., Durella commutata Fuckel, Eutypa maura (Fr.) Sacc., Fusarium stilboides Wollenw., Pezicula acericola (Peck) Peck ex Sacc. and Berl., Phialocephala sp. W.B. Kendr., Phomopsis pustulata (Sacc.) Died., Prosthecium platanoidis (Pers.) M.E. Barr, Splanchnonema pupula (Fr.) Kuntze |
| Chlebicki [27] 2 | wood of dead branches | Eutypa maura (Fr.) Sacc., Nectria cinnabarina (Tode) Fr., Prosthecium platanoidis (Pers.) M.E. Barr, Prosthecium pyriforme Jaklitsch and Voglmayr |
| Kowalski and Kehr [28] 3 | living branch bases | Aposphaeria sp. Berk., Diplodina acerina (Pass.) B. Sutton, Mollisia sp. (Fr.) P. Karst., Petrakia irregularis Aa, Pezicula cinnamomea (DC.) Sacc., Phialocephala dimorphospora W.B. Kendr., Phomopsis sp. Sacc. and Roum., Phomopsis pustulata (Sacc.) Died., Splanchnonema pupula (Fr.) Kuntze, Torula sp. Pers. |
| Ellis and Ellis [29] 4 | wood and bark | Eutypa maura (Fr.) Sacc., Phomopsis pustulata (Sacc.) Died., Splanchnonema pupula (Fr.) Kuntze, Xylaria longipes Nitschke |
| Unterseher, Otto and Morawetz [30] 5 | dead canopy twigs | Auricularia auricula-judae (Bull.) Quél., Eutypa maura (Fr.) Sacc., Nectria cinnabarina (Tode) Fr., Peniophora cinerea (Pers.) Cooke, Peniophora lycii (Pers.) Höhn. and Litsch., Prosthecium acerinum Voglmayr and Jaklitsch, Prosthecium pyriforme Jaklitsch and Voglmayr, Schizophyllum commune Fr., Trichoderma viride Pers., Trichoderma spp. Pers. |
| Johnova [31] 4 | decayed trunks, stumps, twigs and branches | Eutypa lata (Pers.) Tul. and C. Tul., Neonectria coccinea (Pers.) Rossman and Samuels, Xylaria longipes Nitschke |
| Sampling Site | Label | Longitude (°E) | Latitude (°N) | Elevation (m a.s.l.) | Relief Aspect | T (°C) 1 | P (mm) 2 |
|---|---|---|---|---|---|---|---|
| Rožnik | R | 14.48723 | 46.06508 | 345 | NE | 10.0 | 1424 |
| Smrekovec | SM | 14.50187 | 45.88679 | 835 | SW | 7.7 | 1652 |
| Mokrc | M | 14.50965 | 45.88135 | 860 | SW | 6.8 | 1702 |
| Mala voda | MV | 14.26736 | 46.04212 | 536 | NW | 9.1 | 1657 |
| Samotorica | S | 14.25378 | 46.03027 | 647 | S | 9.1 | 1636 |
| Sampling Site | DBH (cm) | Tree Height (m) | Branch Height 1 (m) | Branch Length (cm) | Branch Diameter at Branch Base (cm) | Trunk Diameter at Branch Base (cm) |
|---|---|---|---|---|---|---|
| Rožnik | 3.7 (1.0) | 5.42 (1.69) | 2.19 (1.29) | 28.6 (26.9) | 1.0 (0.5) | 3.7 (0.8) |
| Smrekovec | 2.8 (0.6) | 4.64 (0.51) | 1.25 (0.61) | 63.2 (58.7) | 0.8 (0.3) | 3.2 (0.7) |
| Mokrc | 2.7 (0.6) | 4.28 (0.89) | 1.18 (0.79) | 28.1 (25.1) | 0.6 (0.3) | 3.1 (0.7) |
| Mala voda | 4.0 (0.7) | 6.96 (1.66) | 2.49 (1.39) | 10.0 (0.0) | 1.5 (0.7) | 3.8 (1.1) |
| Samotorica | 4.3 (0.9) | 8.49 (2.05) | 3.27 (1.31) | 5.4 (2.6) | 1.5 (0.5) | 3.8 (1.1) |
| Sampling Site | Visually Healthy | Symptomatic |
|---|---|---|
| Rožnik | 213 | 3 |
| Smrekovec | 153 | 2 |
| Mokrc | 92 | 0 |
| Mala voda | 215 | 1 |
| Samotorica | 314 | 0 |
| Sampling Site | Rožnik | Smrekovec | Mokrc | Mala voda | Samotorica | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Isolation Source | T 1 | B 2 | T | B | T | B | T | B | T | B |
| F (%) | 46.77 | 14.52 | 3.23 | 3.23 | 3.23 | 11.29 | 3.23 | 0.00 | 0.00 | 8.06 |
| Taxon | GenBank Accession No. 1 | F (%) | DI | ZLVG No. 2 |
|---|---|---|---|---|
| Acremonium sp. Link | MN244544 MN240814 | 1.41 | 1.83 | 809 808 |
| Alternaria sp. Nees | MN244537 | 0.64 | 1.43 | 781 |
| Aureobasidium pullulans (de Bary and Löwenthal) G. Arnaud | MN244533 | 0.70 | 2.75 | 782 |
| Bloxamia sp. Berk. And Broome | MN251064 | 0.83 | 2.60 | 810 |
| Cadophora sp. Lagerb. And Melin | MN251055 | 0.90 | 3.50 | 783 |
| Cerrena sp. Gray | MN223745 | 1.28 | 2.50 | 801 |
| Clonostachys sp. Corda | MN244536 | 0.51 | 2.67 | 784 |
| Coprinellus sp. P. Karst. | MN240808 MN244538 MN240810 | 2.75 | 1.79 | 813 812 811 |
| Cosmospora sp. Rabenh. | MN251063 | 0.70 | 1.83 | 785 |
| Cytospora sp. Ehrenb. | MN251061 | 0.26 | 1.00 | 786 |
| Daldinia sp. Ces. And De Not. | MN244534 MN244541 | 1.28 | 2.50 | 814 815 |
| Dendryphion europaeum Crous and R.K. Schumach. | MN251057 | 0.32 | 1.67 | 787 |
| Diaporthe spp. Nitschke | 4.22 | 2.20 | ||
| Diaporthe sp. 1 | MN240809 MN244550 MN244548 | 2.05 | 2.46 | 818 819 816 |
| Diaporthe sp. 2 | MN240816 | 2.18 | 2.00 | 788 |
| Epicoccum nigrum Link | MN216311 MN244547 | 2.37 | 2.31 | 820 817 |
| Eutypa lata (Pers.) Tul. and C. Tul. | MN252417 MN252418 MN252420 | 2.69 | 3.50 | 821 822 824 |
| Eutypa maura (Fr.) Sacc. | MN252421 MN252423 | 8.77 | 2.91 | 789 826 |
| Eutypa spp. Tul. and C. Tul. | 11.77 | 2.83 | ||
| Eutypa sp. 1 | MN252415 MN252411 | 2.24 | 2.69 | 828 790 |
| Eutypa sp. 2 | MN252405 | 7.36 | 3.03 | 831 |
| Eutypa sp. 3 | MN252406 | 1.60 | 1.56 | 832 |
| Eutypa sp. 4 | MN252416 | 0.38 | 1.00 | 829 |
| Eutypa sp. 5 | MN252408 | 0.19 | 1.00 | 833 |
| Eutypella parasitica R.W. Davidson and R.C. Lorenz | MN252407 | 3.97 | 3.26 | 791 |
| Fusarium acuminatum Ellis and Everh. | MN976065 | 1.28 | 1.43 | 843 |
| Fusarium avenaceum (Fr.) Sacc. | MN976063 | 5.50 | 2.00 | 844 |
| Fusarium lateritium Nees | MN240811 MN976066 | 1.92 | 3.00 | 845 846 |
| Fusarium merismoides Corda | MN976064 | 2.50 | 1.86 | 847 |
| Nectria sp. (Fr.) Fr. | MN244545 | 0.38 | 1.50 | 793 |
| Neocucurbitaria acerina Wanas., Camporesi, E.B.G. Jones and K.D. Hyde | MN216310 | 4.54 | 1.92 | 848 |
| Neocucurbitaria sp. Wanas., E.B.G. Jones and K.D. Hyde | MN251052 | 0.45 | 1.40 | 850 |
| Neonectria coccinea (Pers.) Rossman and Samuels | MN242704 | 1.92 | 1.58 | 851 |
| Neonectria sp. Wollenw. | MN252412 | 3.84 | 3.00 | 795 |
| Neosetophoma sp. Gruyter, Aveskamp and Verkley | MN244543 | 0.38 | 1.20 | 796 |
| Nigrograna obliqua Jaklitsch and Voglmayr | MN244540 | 2.69 | 1.83 | 878 |
| Paraphaeosphaeria neglecta Verkley, Riccioni and Stielow | MN240812 MN244542 | 1.47 | 2.30 | 853 854 |
| Parathyridaria sp. Jaklitsch and Voglmayr | MN244551 | 0.32 | 1.67 | 879 |
| Penicillium brevicompactum Dierckx | MN242710 | 0.19 | 1.00 | 855 |
| Peniophora incarnata (Pers.) P. Karst. | MN223746 | 2.82 | 2.00 | 797 |
| Petrakia irregularis Aa | MN216309 | 3.58 | 2.80 | 856 |
| Petrakia sp. Syd. And P. Syd. | MN240815 | 0.90 | 2.33 | 857 |
| Phaeosphaeriaceae sp. M.E. Barr | MN251068 | 2.37 | 1.61 | 858 |
| Phialemonium sp. W. Gams and McGinnis | MN251053 MN251056 | 0.90 | 1.27 | 860 861 |
| Phialocephala sp. W.B. Kendr. | MN242702 | 1.73 | 1.69 | 862 |
| Phomopsis pustulata (Sacc.) Died. | MN251066 | 3.13 | 2.13 | 799 |
| Phomopsis velata (Sacc.) Traverso | MN244546 | 0.58 | 1.50 | 865 |
| Pleosporales sp. Luttr. Ex M.E. Barr | MN251054 | 0.45 | 1.40 | 866 |
| Prosthecium sp. Fresen. | MN251051 | 0.32 | 1.67 | 803 |
| Pseudocosmospora rogersonii C.S. Herrera and P. Chaverri | MN242705 | 2.18 | 2.43 | 867 |
| Sarocladium sp. W. Gams and D. Hawksw. | MN251067 | 1.22 | 1.36 | 800 |
| Splanchnonema pupula (Fr.) Kuntze | MN251065 | 1.15 | 1.64 | 802 |
| Trichoderma atroviride P. Karst. | MN242707 | 1.66 | 1.44 | 880 |
| Trichoderma citrinoviride Bissett | MN242706 | 1.15 | 1.38 | 881 |
| Trichoderma harzianum Rifai | MN242709 | 0.06 | 1.00 | 882 |
| Trichoderma sp. Pers. | MN242708 | 0.13 | 2.00 | 883 |
| Typhula sp. (Pers.) Fr. | MN251062 | 0.32 | 1.67 | 870 |
| Valsa sp. Fr. | MN244549 | 0.38 | 2.00 | 804 |
| Xylaria longipes Nitschke | MN251059 | 0.51 | 2.00 | 868 |
| Xylaria sp. Hill ex Schrank | MN240813 MN251060 | 1.73 | 2.45 | 872 869 |
| Sampling Site | Total Number of Fungal Taxa 3 | Average Number of Fungal Taxa per Sample | Colonization Rate (%) 1 | Shannon Diversity (H’) | Species Evenness (J’) | |||
|---|---|---|---|---|---|---|---|---|
| T | B | C | Average 2 | |||||
| Rožnik | 42 | 4.64 | 92.02 | 100.00 | 2.50 | 96.01 | 3.30 | 0.88 |
| Smrekovec | 35 | 3.30 | 86.11 | 93.21 | 1.88 | 89.66 | 3.01 | 0.85 |
| Mokrc | 35 | 3.30 | 95.31 | 84.72 | 0.63 | 90.02 | 3.15 | 0.88 |
| Mala voda | 40 | 3.62 | 90.82 | 87.50 | 0.00 | 89.16 | 3.16 | 0.86 |
| Samotorica | 42 | 3.63 | 92.36 | 91.33 | 0.00 | 91.84 | 3.21 | 0.86 |
| Isolation Source 1 | |||
|---|---|---|---|
| T | B | C | |
| Shannon diversity (H’) | 3.51 | 3.59 | 1.15 |
| Species evenness (J’) | 0.87 | 0.91 | 0.83 |
| Sampling Site | Rožnik | Smrekovec | Mokrc | Mala voda |
|---|---|---|---|---|
| Smrekovec | 0.55 | |||
| Mokrc | 0.63 | 0.60 | ||
| Mala voda | 0.62 | 0.51 | 0.60 | |
| Samotorica | 0.65 | 0.51 | 0.57 | 0.64 |
| Fungal Taxa | Branch Base Diameter | |||||
|---|---|---|---|---|---|---|
| 0.2–0.7 cm | 0.8–1.2 cm | 1.3–3.2 cm | ||||
| n 1 | % 2 | n | % | n | % | |
| Coprinellus sp. | 6 | 25.00 | 13 | 54.17 | 5 | 20.83 |
| Eutypa maura | 9 | 19.15 | 25 | 53.19 | 13 | 27.66 |
| Eutypa sp. 2 | 11 | 29.73 | 17 | 45.95 | 9 | 24.32 |
| Eutypella parasitica | 7 | 36.84 | 9 | 47.37 | 3 | 15.79 |
| Fusarium avenaceum | 20 | 46.51 | 12 | 27.91 | 11 | 25.58 |
| Neocucurbitaria acerina | 18 | 48.65 | 10 | 27.03 | 9 | 24.32 |
| Neonectria sp. | 6 | 30.00 | 8 | 40.00 | 6 | 30.00 |
| Peniophora incarnata | 11 | 50.00 | 5 | 22.73 | 6 | 27.27 |
| Petrakia irregularis | 7 | 35.00 | 9 | 45.00 | 4 | 20.00 |
| Phomopsis pustulata | 13 | 56.52 | 7 | 30.43 | 3 | 13.04 |
| OI 3 | 3.60 | 3.75 | 3.80 | |||
| Shannon diversity index (H’) | 3.56 | 3.71 | 3.69 | |||
| Species evenness (J’) | 0.93 | 0.92 | 0.94 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brglez, A.; Piškur, B.; Ogris, N. Eutypella parasitica and Other Frequently Isolated Fungi in Wood of Dead Branches of Young Sycamore Maple (Acer pseudoplatanus) in Slovenia. Forests 2020, 11, 467. https://doi.org/10.3390/f11040467
Brglez A, Piškur B, Ogris N. Eutypella parasitica and Other Frequently Isolated Fungi in Wood of Dead Branches of Young Sycamore Maple (Acer pseudoplatanus) in Slovenia. Forests. 2020; 11(4):467. https://doi.org/10.3390/f11040467
Chicago/Turabian StyleBrglez, Ana, Barbara Piškur, and Nikica Ogris. 2020. "Eutypella parasitica and Other Frequently Isolated Fungi in Wood of Dead Branches of Young Sycamore Maple (Acer pseudoplatanus) in Slovenia" Forests 11, no. 4: 467. https://doi.org/10.3390/f11040467
APA StyleBrglez, A., Piškur, B., & Ogris, N. (2020). Eutypella parasitica and Other Frequently Isolated Fungi in Wood of Dead Branches of Young Sycamore Maple (Acer pseudoplatanus) in Slovenia. Forests, 11(4), 467. https://doi.org/10.3390/f11040467