Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a great threat to public health, being a causative pathogen of a deadly coronavirus disease (COVID-19). It has spread to more than 200 countries and infected millions of individuals globally. Although SARS-CoV-2 has structural/genomic similarities with the previously reported SARS-CoV and MERS-CoV, the specific mutations in its genome make it a novel virus. Available therapeutic strategies failed to control this virus. Despite strict standard operating procedures (SOPs), SARS-CoV-2 has spread globally and it is mutating gradually as well. Diligent efforts, special care, and awareness are needed to reduce transmission among susceptible masses particularly elder people, children, and health care workers. In this review, we highlighted the basic genome organization and structure of SARS-CoV-2. Its transmission dynamics, symptoms, and associated risk factors are discussed. This review also presents the latest mutations identified in its genome, the potential therapeutic options being used, and a brief explanation of vaccine development efforts against COVID-19. The effort will not only help readers to understand the deadly SARS-CoV-2 virus but also provide updated information to researchers for their research work.
1. Introduction
The emergence and re-emergence of pathogens is a global human health concern [1]. Coronaviruses are enveloped, their genomes are non-segmented, and they are single-stranded positive-sense RNA (+ssRNA) viruses belonging to the family Coronaviridae and order Nidovirales, which are widely dispersed in humans, animals, and birds. Coronaviruses cause various life-threatening diseases from respiratory infections to hepatic, enteric, and severe neurological diseases [2,3]. Six species of Coronaviruses were known to cause human diseases [4], out of which four (HKU1, NL63, 229E, and OC43) are widespread and responsible for the common cold in individuals with a weak immune response [4]. SARS-CoV-2 is the seventh coronavirus known to infect humans. Its exact origin is unknown; however, it shows homology with the previously identified coronavirus strains SARS-CoV (intermediate host, masked palm civet) and MERS-CoV (intermediate host, dromedary camel) [5,6]. The homology between SARS-CoV-2 and SARS-CoV is 82.45%, and the homology between SARS-CoV-2 and MERS-CoV is 69.58% [7]. SARS-CoV was responsible for SARS outbreaks in 2002–03 in Guangdong Province, China [8,9,10], while MERS-CoV was responsible for respiratory illness in the Middle East in 2012–13 [11]. The mortality rates of MERS and SARS were 37% and 10%, respectively [12,13]. SARS-CoV-2 triggered the COVID-19 pandemic, which spread rapidly worldwide and has become a public health concern [14]. In this review, we focus on SARS-CoV-2 novel genome organization, newly identified mutations in its genome, its transmission dynamics, clinical symptoms, potential treatment strategies, and recent advancements of vaccine production against COVID-19.
2. Insights into Genomic Organization
Coronaviruses, which belong to the Coronaviridae family, are enveloped and pleomorphic viruses [15]. These are positive-sense RNA viruses with a genome size of 30 kb; which appears to be the largest size for a RNA virus, containing a 5′ cap and 3′ poly A-tail. Coronaviruses have a helical and flexible nucleocapsid. The membrane of these viruses contains a membrane glycoprotein, enveloped protein, and spike protein while the RNA is surrounded by nucleocapsid [16,17].
Virus RNA contains 6 open reading frames (ORF1ab, ORF3a, ORF6, ORF7ab, ORF8, and ORF10). Two-thirds of the virus genome comprises 1a/1b ORF and the remaining one-third of the genome code is used for M (membrane), S (spike), N (nucleocapsid), and E (enveloped) viral structural proteins [18,19].
Transcription was carried out by the synthesis of sgRNA (sub-genomic RNA) and replication-transcription complex (RTC), enveloped in double-membrane vesicles. Transcription termination occurred through transcription regulatory sequences that are present in between open reading frames (ORFs). There are 6 ORFs in the SARS-CoV-2 genome, as discussed above [18]. A frameshift mutation in ORF1a and ORF1b produces polypeptides (pp1a and pp1ab), which are further processed by virally encoded proteases such as main proteases (Mpro), chymotrypsin-like proteases (3CLpro), or by papain-like proteases for the production of non-structural proteins (nsps) [20,21]. Besides 1a and 1b open reading frames (ORFs), all other ORFs are responsible for the production of structural proteins (membrane, nucleocapsid, enveloped, and spike proteins), as shown in Figure 1.
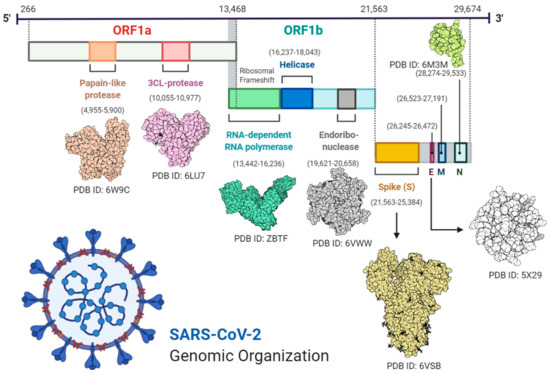
Figure 1.
Complete structural and genomic organization of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [23].
Through sequence analysis of SARS-CoV-2 and SARS-CoV, scientists proposed a mutation in the spike protein responsible for the jumping of the virus from animals to humans [22]. Similarly, some mutations have also been found in protein sequences which lead to the formation of proteins with a change in amino acid residues. For example, at position 723, instead of glycine there is a serine, while at position 1010 there is proline instead of isoleucine [22]. Potential disease recurrence depends on the evolution of the virus due to the accumulation of mutations in the viral genome over time.
2.1. Genome Sequencing
Through genomic sequence analysis, it has been confirmed that although SARS-CoV-2 has many similarities with SARS-CoV and other related coronaviruses, it is a novel virus (Table 1). The virus made a shift in the host organism from animals to humans with a few unique modifications/mutations. Genome sequence analysis suggests that most of the viral contigs/reads had a similarity with the genome of beta-coronavirus. SARS-CoV-2 has 96.20% and 88.00% levels of similarity to the previously published SARSr-CoV (RaTG13) and bat-SL-CoVZC45 genomes, respectively [3]. The sequencing of the SARS-CoV-2 genome from another study indicated 69.58% and 82.45% sequence similarity with MERS-CoV and SARS-CoV genomes, respectively [5,24]. Ten viral genome sequences obtained from 9 patients exhibited 99.98% sequence identity. In another study, sequences from eight patient samples had 99.98% sequence identity with each other across the whole genome [24]. BLASTn search of SARS-CoV-2 sequences has identified matches from the most closely related previously known viruses: SARS-like beta-coronavirus of bat origin, bat-SL-CoVZC45 (sequence identity 88%; query coverage 99%), and bat-SL-CoVZXC21 (sequence identity 88%; query coverage 98%). In 5 gene regions (7, M, N, 14, and E), sequence identity was more than 90% with 98.7% as the highest level for the envelope (E) gene. The Spike (S) gene demonstrated the lowest sequence identity of 75%. However, the sequence identity in 1a and 1b gene regions was 90% and 87%, respectively [24]. The majority of proteins encoded by SARS-CoV-2 were highly similar to proteins encoded by bat-related coronaviruses with a few insertions and deletions [24]. However, protein 13 and the S protein revealed 73.2% and 80% identity with other bat-derived viral proteins, respectively [25]. SARS-CoV-2 encoded a large spike protein, which is a major distinguishing feature among SARS-CoV-2, SARS-CoV, MERS-CoV, and other bat-derived coronaviruses. SARS-CoV-2 exhibits the same genomic organization as bat-SL-CoVZXC21, SARS-CoV, and bat-SL-CoVZC45, as revealed by comparison of predicted coding regions. Ten coding regions were identified including E, M, N, S, 10ab, 9, 8, 7, 3, and 1ab [24].

Table 1.
Sequence homology between SARS-CoV-2 and other coronaviruses strains [7].
2.2. Phylogenetic Analysis
Phylogenetic analysis of SARS-CoV-2 genomes obtained from early patient samples suggested similarity in the sequence organization with beta-coronaviruses such as 5′ UTR (untranslated region), replicase complex (orf1ab), 4 genes (M, N, S, and E), 3′ UTRs (untranslated regions1), and some unidentified non-structural ORFs (open reading frames) [26]. Instead of having sequence similarity with beta-coronaviruses discovered in bats, SARS-CoV-2 is distinct from SARS-CoV, as well as MERS-CoV. Another piece of evidence pointing to its novelty is that the sequence identity in conserved replicase domains (ORF 1ab) is less than 90% between SARS-CoV-2 and other members of beta-coronaviruses and sarbeco-virus sub-genus of the Coronaviridae family [3].
2.3. Conserved Proteins
The S protein is responsible for membrane fusion and receptor binding. It is also critical in controlling virus transmission capacity and host tropism. The S protein of SARS-CoV-2 has two domains, namely the S1 and S2 domains. The S1 domain is responsible for receptor binding, while the S2 domain for membrane fusion [27]. It has been reported that a cellular protease (furin) is responsible for the cleavage of S1/S2 sites and this cleavage is necessary for the entry of virion in human lung cells and S protein facilitated cell fusion [28]. The S1 and S2 domains of SARS-CoV-2 have a sequence similarity of 93% and 68% with bat-SL-CoVZXC21 and bat-SL-CoVZC45, respectively [24,29]. Among sarbeco-coronaviruses, amino acid variations in S protein were identified. Although SARS-CoV and SARS-CoV-2 belong to different clades in the phylogenetic tree, they have 50 conserved amino acids in the S1 domain of the S protein. However, MERS-CoV has mutational differences in S proteins. Most of these mutational events occur in the C-terminal domain [24]. Several other proteases are also involved in different processes, such as entry of the virion, maturation of polyprotein, and assembly of different virion particles [30]. Other than the S protein, a variety of SARS-CoV-2 other proteins show similarity with proteins of other Coronaviridae family members, as shown in Table 2.
Table 2.
Percentage identity between proteins of SARS-CoV-2 and the Coronaviridae family [31].
2.4. Receptor Binding Domain (RBD)
The RBD of SARS-CoV-2 is found in the C-terminal domain of spike protein as in SARS-CoV, Bat CoV HKU4, and MERS-CoV [32,33]. It was also reported that SARS-CoV-2 uses ACE2 (angiotensin-converting enzyme) as a cell receptor for entry into the human cells [34]. From the phylogenetic analysis, it was found that at genome level, SARS-CoV-2 is closely related to bat-SL-CoVZXC21 and bat-SL-CoVZC45, though the RBD of SARS-CoV-2 is highly similar to SARS-CoV. However, key residues of the receptor-binding domain responsible for the binding of the receptor were different in SARS-CoV-2 as compared to SARS-CoV [24]. From the above studies, it is again established that although SARS-CoV-2 has a great similarity with MERS-CoV, SARS-CoV, and some other bat-derived coronaviruses, it is a novel version of coronavirus and is responsible for an infection that is spreading globally.
3. SARS-CoV-2 Recent Mutations
RNA viruses have a high mutation rate but to survive, they achieve a balance of adaptions via mutations and replication-competent genome. For example, wild type SAR-COV reported showing less than one mutation in a sequenced genome (error frequency of 2 × 10−5), whereas SARS-CoV ExoN(−) revealed 10 mutations in a sequenced genome (error frequency of 3 × 10−4) [35,36]. Like other RNA viruses, SARS-Cov-2 has a high mutation rate and its multiple variants have been reported globally [37]. Research has revealed that 93 mutations were observed over the entire SARS-CoV-2 genome. In all major proteins, except for the envelope protein, 42 missense mutations were identified. There were 29 missense mutations in the polyprotein encoded by ORF1ab, 8 in the glycoprotein of the spike surface, 1 in the matrix protein, and four in the protein of nucleo-capsid. Spike surface glycoprotein has an important role in binding with host cells and regulates the host response [38]. It is also the key target of antibody neutralization [5]. Mutations in the glycoprotein present in the spike surface can induce some important conformational changes that can change antigenicity [39]. In another study, 5775 different genome mutations have been reported, including 2 in-frame insertions, 11 frameshift deletions, 36 stop-gained variants, 66 insertions in the non-coding region, 100 in-frame deletions, 142 deletions in non-coding regions, 484 mutations in non-coding regions, 1965 synonymous mutations, and 2969 missense mutations [40].
The UK faced a dramatic rise in COVID-19 cases that led to an increased number of epidemiological and virological surveys. The viral genome sequence has revealed that a significant number of cases belong to a newly reported phylogenetic cluster. The new variant of SARS-CoV-2 (B.1.1.7) has multiple spike protein mutations, (deletion 69–70, P681H, T716I, S982A, deletion 144, N501Y, D614G, A570D, D1118H) in addition to various mutations in different genomic regions [41]. Although it is clearly understood and predicted that viruses will continue to evolve due to mutations leading to the development of new variants, preliminary research in the UK indicates that this variant is substantially more infectious than its previous variants, having the potential of increasing the reproductive number by 0.4 or higher with increased transmissibility of up to 70 percent. This new variant appeared at the time of the year when social mixing has been increased traditionally. At this stage, there is no evidence of increased severity of infection linked with this new coronavirus variant [41]. This variant is also spreading in different countries throughout the world. Another variant known as B.1.351 has emerged independently in South Africa. It has many similarities with the B.1.1.7 variant and was detected in early October 2020. This variant has also spread to the USA and cases caused by this variant were reported there in early January 2021. A variant known as P.1 emerged in Brazil in early January 2021. It was first discovered in travelers during their regular screening in Japan. This variant has some additional mutations that may affect an antibody’s capability of identifying SARS-CoV-2. These newly identified strains seem to spread more rapidly. This is expected to eventually increase the coronavirus cases worldwide, which may result in a greater burden on hospitals and more deaths (Centers for Disease Control and Prevention).
4. Pathophysiology and Epidemiology
The recently identified SARS-CoV-2 is contagious and spreading globally with many confirmed cases. The infection caused by SARS-CoV-2 is transmitted by person-person contact via respiratory droplets [42]. The invasion of virion in different cells of the vascular system results in different inflammatory changes, such as necrotic changes, edema, and degeneration. It has been reported that SARS-CoV-2 infection also causes hypoxemia. It has been proven that the respiratory system is highly affected by this infection. Many neurologic symptoms have been associated with SARS-CoV-2 infection, such as agitation, dizziness, headache, confusion, and seizure [43].
Millions of people have been affected by SARS-CoV-2 and the mortality rate (number of deaths in a specific population with respect to population size during a specified time interval) is higher in males as compared to females, while individuals >65 years of age have also experienced a higher death ratio in comparison with younger adults and children. According to one study, the mortality rate is 62 times higher among individuals that are 55–65 years or higher, in comparison with individuals less than 54 years of age [44,45]. To date, there have been more than 200 countries affected by SARS-CoV-2 infection. The maximum number of cases has been reported in the USA (>25.3 million) followed by India (>10.7 million), Brazil (>9.9 million), and Russia (>3.7 million), while the death rate is higher in the USA and Brazil as compared to India and Russia (John Hopkins coronavirus Resource Center: https://coronavirus.jhu.edu/map.html (accessed on 1 February 2021). The mortality rate is highly variable among different countries, depending highly on health status, age statistics, and healthcare systems. In some countries, the mortality rate and active cases are increasing exponentially since the outbreak and are still at their peak. More than 100 million cases and more than 2.1 million fatalities have been reported so far across the world (John Hopkins coronavirus Resource Center).
5. Transmission Dynamics
Since the symptoms and features of SARS-CoV-2 were similar to pneumonia, this created confusion in the recognition of this novel coronavirus at the early stages. However, when disease conditions did not improve, physicians and doctors realized that something was wrong with diagnosis and treatment. Gradually, an increase in the patient count was reported, new cases were increasing day by day, and no treatment options were helping. The number of infected people kept on increasing and upon screening and testing, it was confirmed that this novel disease is pneumonia-like but is not pneumonia. There was a different viral agent behind the entire situation. A gradual increase in cases was due to late identification and reporting of diseases.
Over time, it was identified that the virus spreads from human to human through close contact (from breathing, sneezing, and coughing). One study revealed that old people are more prone to infection caused by SARS-CoV-2 than young individuals and children. The age range of 425 early-infected individuals in Wuhan, China was 15–89 years. Out of 425 patients, 240 were males (almost 56%), and the rest were females [46]. The literature revealed the estimated mean incubation period of SARS-CoV-2 was 5.2 days. The mean duration from onset of disease to the first medical visit of patients was 12.5 days. The doubling period and epidemic growth rate was 7.4 days and 0.10/day, respectively [46]. Delayed disease recognition caused a deficiency of isolation systems for patients. Not practicing social distancing has paved the way for SARS-CoV-2 to spread all over the world and for the transmission of infection among millions of people. The death toll has risen, quickly triggering a pandemic.
6. Clinical Symptoms
Clinical symptoms of SARS-CoV-2 infection include coughing, fever, and shortness of breath/difficulty in breathing followed by severe outcomes like sepsis, ARDS (acute respiratory distress syndrome), acute kidney injury, and acute cardiac injury [47]. Clinical manifestations/symptoms of SARS-CoV-2 are divided into 3 groups according to the severity of the disease:
6.1. Mild Disease
Mild disease is characterized by mild pneumonia/non-pneumonia. This situation occurred in 80% of the SARS-CoV-2 cases [48].
6.2. Severe Disease
Characteristics of severe disease are dyspnea, blood oxygen saturation (SpO2) ≤ 93%, a high PaO2/FiO2 ratio or P/F [partial pressure of oxygen, (PaO2), a fraction of inspired oxygen, FiO2)] < 300, and/or lung infiltrates >50% within 24 to 48 h; severe disease occurred in 10% of cases and was affected by patients’ respiratory frequency (>30/min) [49]. The mortality rate of SARS-CoV-2 has varied greatly at 0.7–1.3% and sharply changes from <0.002% for children under 9 years of age to 8% in patients aged over 80 years [50,51].
6.3. Critical Disease
The critical disease is characterized by multi-organ dysfunction/failure, sepsis/septic shock, and respiratory failure. Multi-organ dysfunction occurred in 1.4% of cases [38,39]. Intubation is provided to critical patients that have exerted positive effects on patients and approximately 3.2% of patients of SARS-CoV-2 require intubation or invasive ventilation at some stage of their disease [52,53].
7. Duration of SARS-CoV-2 Replication
The duration and virus replication rate are significant factors to determine the transmission risk and to make the decision pertinent to patient’s isolation [47]. Viral RNA detection is more feasible and a quick option that is adopted well to routine and emergency diagnosis compared to virus isolation, so many laboratories have carried out viral RNA quantification and qualification tests as a possible marker for the identification of any infectious virus [54]. Previously, the duration of SARS-CoV and MERS-CoV RNA detection had been documented. In the case of SARS-CoV, viral RNA was detected in the respiratory sample from 5 days to 28 days of disease onset [55]. Similarly, MERS-CoV viral RNA persisted from 3 days to 21 days in a lower respiratory sample [56,57]. In the case of SARS-CoV-2, viral RNA was present in surviving patients for 14–24 days. However, in non-survivors, viral RNA was sustained until death occurred. This information is very important in deciding a patient’s isolation and medication. One study described the viral replication duration among surviving and non-surviving patients [47]. It was found that the median duration for viral shedding or replication was 20 days among survivors from disease onset. However, virus replication was continuously detectable among non-surviving patients until death. The shortest duration of viral replication observed was 8 days among surviving patients [47]. Similarly, the longest duration of viral replication observed was 37 days in immunosuppressed patients [47]. In severe influenza virus infection, extended viral shedding was associated with fatal outcomes. Delayed antiviral treatments (antiviral drugs or antibodies treatments) were an independent risk factor associated with almost all forms of virus detection for a long time [47,58].
8. Risk Factors
There are always risk factors associated with the severity of a disease. There are multiple risk factors, which range from age factors to various diseases from which the patient is already suffering [59,60]. This is relevant for SARS-CoV-2. Previous studies related to SARS-CoV and MERS-CoV revealed risk factors associated with these diseases. For example, Sequential Organ Failure Assessment (SOFA) and blood level of d-dimers were major risk factors for the fatal illness of patients infected by SARS [32,33]. SOFA (Sequential Organ Failure Assessment) is a good marker for sepsis/septic shock diagnosis that tells us about the degree and state of multi-organ dysfunction [61,62]. Bacterial infections mainly cause sepsis, but some viral infections also lead to septic shock. Previous studies revealed that 40% of individuals with pneumonia had sepsis [63]. Similarly, age was also a major risk factor in patients affected by MERS and SARS [64,65]. A study was carried out to determine risk factors associated with SARS-CoV-2 [47]. The results suggested that people with a concomitant MERS or SARS status are more susceptible to SARS-CoV-2 than normal individuals. Concomitance was common in half of the patients, with the most common concomitant factor found to be hypertension followed by coronary heart diseases and diabetes [47]. Age is also a major factor since due to faulty function of T-cells and B-cells, excessive production of cytokines in older age leading to poor control on viral replication, and an extended pro-inflammatory response resulting in deadly outcomes [66]. The research included 191 patients, from which 56 died and the remaining patients recovered from the disease. The median age of the abovementioned 191 patients was 56; however, the overall age ranged from 18–87 years for the majority of the male population. It is also determined that > half of patients had sepsis [47]. Carcinoma, chronic kidney diseases, and chronic obstructive lung disease were also present in 2, 2, and 6 patients out of 191, respectively [47]. The risk factor data are summarized in Table 3.
Table 3.
Percentage of risk factors between survivors and non-survivors (associated with the severity of SARS-CoV-2) [47].
Apart from age and comorbidity, lymphopenia, leukocytosis, elevated ALT, procalcitonin, lactate dehydrogenase, creatine kinase, high-sensitivity cardiac troponin I, prothrombin time, d-dimer, creatinine, serum ferritin, and IL-6 were also associated with mortality outcomes. Lymphocyte counts were low in patients with SARS-CoV-2. Surviving patients have a low number of lymphocytes with the onset of the disease but with hospitalization, the lymphocyte number improved. On the other hand, patients who died had severe lymphopenia with a very low number of lymphocytes [47]. Similarly, non-surviving patients had high levels of d-dimer, IL-6, high-sensitivity cardiac troponin I, lactate dehydrogenase, and serum ferritin throughout the clinical course, which increased with the worsening of illness as compared to for patients who survived. It was also seen that a high SOFA number, older age, and a blood d-dimer presence >1 μg/mL were associated with a large number of unexpected deaths [47]. Heart diseases such as new or worsening arrhythmia, myocardial infarction, and new or worsening heart failure were usually seen in patients with pneumonia. In 3% of patients with pneumonia, cardiac arrest occurred [67]. Apart from pneumonia, age, and pre-existing heart problems are usually associated with cardiac failure [68]. In viral respiratory infections and influenza, coronary heart diseases are associated with acute cardiac events [47,69]. In more than half of SARS-CoV-2 patients who died, elevated high-sensitivity cardiac troponin I was found. The reason was that increased d-dimer concentrations (>1 μg/mL) in the blood led to coagulation [47]. Previous studies also suggested that 90% of the patients with pneumonia had high d-dimer concentrations leading to coagulation [70]. High d-dimer levels associated with 28 days of mortality in patients with sepsis was also reported [71]. A summary of these laboratory biomarkers and their relevant levels in survivors and non-survivors is mentioned in Table 4.
Table 4.
Differentiation of various biomarkers among survivors and non-survivors.
9. Treatment
There are some vaccines available that are authorized/approved and protect against coronavirus, but no specific drugs are available for the affected persons to date. However, different combinations of medicines and therapies are used to treat patients. Antiviral and anti-inflammatory treatments have been utilized. Most treatments are supportive and symptomatic. As symptoms of this disease are similar to those of pneumonia, early diagnosis and control of symptoms would lead to valuable results. For critical patients, invasive mechanical ventilation, non-invasive ventilation, continuous renal replacement therapy (CRRT), Oseltamivir therapy, and extracorporeal membrane oxygenation (ECMO) are applied to patients with hypoxemia [59]. Immunoglobulin G and convalescent plasma are also given to patients according to the situation [72]. Patients with severe shortness of breath can be shifted to intensive care units (ICU).
9.1. Antiviral Treatments
Treatments against SARS and MERS will give us guidance for treating coronavirus patients with previously utilized antiviral medicines [73]. To date, no specific drugs have been discovered for treating coronavirus. Previously, for SARS-CoV and MERS-CoV, various antiviral drugs were used to treat the diseases. Examples include ribavirin, methylprednisolone, neuraminidase inhibitors (peramivir, oseltamivir, zanamivir), acyclovir, and ganciclovir [59]. However, these drugs are not proving beneficial for the treatment of SARS-CoV-2. Fortunately, HIV (Human immunodeficiency virus) protease inhibitors and nucleoside analogues have been identified as a potential treatment and can be used to treat SARS-CoV-2 patients until the discovery of a specific drug is achieved [74]. Currently, patients with severe disease symptoms are treated with systemic glucocorticoids (which have anti-inflammatory, vasoconstrictive and antifibrotic effects [75]) and vasopressors (which reduces the ventilation duration and shortens the shock period [59]). In the beginning, oseltamivir (a medicine used to treat influenza) was also used to treat patients with early diagnosis to control symptoms [76] but, it has been discovered now that it has no impact against SARS-CoV-2 [77]. Antibacterial drugs such as moxifloxacin (which interacts with SARS-CoV-2 Main protease enzyme [78]) and azithromycin (which has arrhythmogenic potential, so it is not recommended by WHO but some local organizations have approved it with chloroquine [79]) are also being given to the patients [59]. An antiviral drug, GS-5734 (remdesivir), was also used successfully in the US. Remdesivir is a nucleoside analogue that has antiviral ability against many RNA viruses. As SARS-CoV-2 is an RNA virus, remdesivir is used as a drug to treat patients with severe symptoms. Remdesivir has the potential to block RNA dependent RNA polymerase activity [80]. It also interferes with nsp2 polymerase and the exon proofreading mechanism [81]. Remdesivir grabbed worldwide attention due to its effectiveness against the current outbreak of SARS-CoV-2 in in-vitro experiments [82]. Thus, it was thought that it can be used as a potential drug to treat viral infection caused by SARS-CoV-2, but lately, it has shown no impact in the treatment of SARS-CoV-2 in clinical trials (World Health Organization). Chloroquine is another drug that had been used previously as an anti-malarial as well as for autoimmune disorders [83]. Chloroquine is known to glycosylate viral cell receptors and increase endosomal PH, which disrupt the fusion of viruses with host cells. Chloroquine also has antiviral and immune-modulating activities, acting as an autophagy inhibitor suppressing the release and production of IL-6/ TNF-alpha. This drug is widely distributed in lungs and other parts of the body through an oral intake. One experiment carried out to check chloroquine activity demonstrated that chloroquine could play active role at entry and post-entry stages of SARS-CoV-2. Chloroquine is an economic and safe drug that has been in use for 70 years. It can be a potentially applicable drug to treat SARS-CoV-2 infections [84]. Other research revealed improved clinical outcomes in patients treated with chloroquine [85,86,87]. On the other hand, hydroxychloroquine has proven to be more potent than chloroquine, as no virus strain has been detected later in patients treated with hydroxychloroquine [82,88,89]. Thus, it could also be a potential drug to treat SARS-CoV-2 patients. Lopinavir and ritonavir are protease inhibitors that were used previously to treat HIV [90]. These drugs have been used against SARS-CoV and MERS-CoV, resulting in improved outcomes in patients [91,92]. Ritonavir and lopinavir combinations are also utilized against SARS-CoV-2. Improvements have been seen in Korean patients regarding the decrease in viral loads after intakes of ritonavir and lopinavir [93]. In Shanghai, improvements in pneumonia associated symptoms had also been seen in patients after ritonavir and lopinavir treatment [94]. The mortality rate and viral RNA detection were similar among patients treated with ritonavir and lopinavir as compared to for patients with no such treatment. Instead, ritonavir and lopinavir also show adverse effects in patients, as gastroenteritis was more prevalent in patients treated with these drugs. That is why this treatment was stopped after early trials due to its adverse effects on patients’ health [82]. However, in the same country, patients also recovered after ritonavir and lopinavir administration and a decrease in viral load was observed to the extent that no detectable viral concentrations were observed after treatment. It also helped patients to regain their normal body temperature [82]. Another study suggested that ritonavir and lopinavir treatment did not decrease the viral shedding process [82]. The mechanism behind this process of resistance is not understood yet. Some patients have shown improvement and others have developed different diseases after treatment with ritonavir and lopinavir. There may be mutations in the viral genome which make them resist antiviral treatments or there may be some other reasons for this outcome like patients’ clinical conditions and age factors. Further research is needed to understand the complete process. It has been reported that along with ritonavir and lopinavir, many other compounds might be useful for treating SARS-CoV-2 infection including nucleoside analogues, anti-inflammatory drugs (hormones and other molecules), neuraminidase inhibitors, RNA synthesis inhibitors (3TC and TDF), remdesivir, abidol, peptide (EK1), and Chinese traditional medicines such as LianHuaQingWen and ShuFengJieDu capsules [74].
9.2. Potential Therapeutic Compounds and Drugs
In addition to the drugs mentioned above, various other drugs will be developed against the coronavirus. Clinical trials are in progress to develop specific antiviral drugs and vaccines against SARS-CoV-2. Scientists are working hard on the development of a specific vaccine to control this pandemic. Some viral targeted agents may have the potential to work against SARS-CoV-2 in the future, as mentioned in Table 5. In addition to the abovementioned potential therapeutic agents and drugs, targeting cells inducing inflammatory response can also be an option for the treatment of SARS-CoV-2 patients. Inflammatory response leads to bursting of cytokines upon the onset of infection, consequently activating the innate immune response. One example is of toll-like receptors (TLRs) inducing signaling pathways upon activation, in turn leading to cytokines production and innate immune response. Targeting Toll-like receptors with some kind of antagonist (e.g., anti-TLR4-specific antibodies and TAK-242) would help to control the drawbacks associated with the genetic heterogeneity of SARS-CoV-2 [95,96]. Another option is to use immunosuppressants e.g., corticosteroids to treat the higher stage of infections. However, the use of immunosuppressants has its side effects. For instance, in influenza, immunosuppressants led to a higher death rate, superinfections, as well as long term viral exposure in patients [97]. In the case of MERS-CoV and SARS-CoV, the number of deaths was not increased but the virus was retained for a longer time in patients [98]. A better option is to target proteins that interact with the S (spike) proteins of the virus and allow them to enter the host cell. For example, type II transmembrane serine protease (TMPRSS2) is a host cell protease that activates spike proteins of the SARS virus. Targeting these proteases will lead to the development of the best therapy against SARS-CoV-2. For example, camostat mesylate is a serine protease inhibitor that suppresses and inhibits the enzymatic activity of TMPRSS2 [98]. However, in vivo studies are needed for a complete understanding of the TMPRSS2 antiviral mechanism. These are some of the potential therapies and treatment options that may prove beneficial to treat coronavirus. Diligent efforts are required for identifying an effective treatment against the virus. The vaccine could protect the public from getting infected, but once a person is affected, it could no longer help with treatment. Hence, there is a need to find out therapeutic targets in coronavirus. These targets can be proteins, enzymes, pathways, and receptors which are involved in the host-virus interaction.
Table 5.
Potential therapeutic compounds and drugs for SARS-CoV-2 treatment (in trials).
9.3. Nutritional Treatment
Nutritional treatment is also an option for the treatment of patients infected with any virus. Vaccine/drug discovery and development is a time-consuming process and patients do not have enough time to wait. In the current scenario, nutritional treatment can be given to patients for quick recovery. Vitamin C/ascorbic acid is a powerful compound and is used to treat many viral diseases. Intake of vitamin C in prescribed quantities can help in the cure of disease and help patients to recover quickly. The vitamin C treatment has been successfully used against swine flu, bird flu, Chikungunya, and SARS in the past. It boosts the immune system and directly denatures/kills many human viruses. Vitamin C helps the immune system in fighting viruses, but its dosage is important. Even low doses help patients with a strong immune system. Intake of 200 mg vitamin C per day is reported as favorable for prevention, lessening disease complications and shortening the disease course, and allowing effective management of COVID-19 [104,105]. It also helps in treating the symptoms of severe respiratory illness [98]. Physicians of the Orthomolecular Medicine News Service review board recommended 3000 mg/day of vitamin C in various doses. It can be taken in the form of vinegar (Ascorbic acid) or sodium ascorbate, which is non-acidic. Appropriate agents as antiviral agents can be sprayed with nebulization quickly, thereby eliminating the virus in the pharynx area and wiping up the virus in the rest of the body. A favorable dose of hydrogen peroxide as 3% several times a day is also given as antiviral treatment [106]. Other recommended nutrients are zinc, magnesium, selenium, vitamin A, vitamin B complex, and vitamin D3. Magnesium is a key component of many biochemical pathways, and an intake of 400 mg/day should be included in the daily diet. Due to the lack of magnesium in food nowadays, it is supplemented in the form of chelate, citrate, chloride, or malate form. Viral infections like that of polio can be cured through magnesium more effectively than through vitamin C supplements [107]. Zinc is another nutrient that is essential in many biochemical pathways. It acts as an antioxidant and helps the body to fight against Swine flu, bird flu, and SARS [107]. A total of 20–40 mg/day dose of zinc is recommended for adults [108]. In the presence of selenium, the virus fails to mutate and immunocompetence is improved [109,110]. Nutritional treatment is not just an idea to cure diseases; realistically nutrients are necessary for the body to fight against viruses, as they enhance the overall immunity of the body [111].
10. Platforms for SARS-CoV-2 Vaccine Development
There are various platforms designed to develop vaccines against SARS-CoV-2, for instance, whole virion vaccines (inactivated and live attenuated), nucleic acid vaccines, protein subunit vaccines, and recombinant viral vaccines.
Live attenuated vaccines (LAV) are viruses that can replicate through several passages from cell culture. On the other hand, inactivated vaccines are viruses that are inactivated by exposure to chemicals (e.g., formaldehyde) and heat [112]. Live attenuated vaccines are immunogenic and produce long-lasting immune responses in the host which protect against live pathogens [112,113]. Inactivated vaccines produce a weaker immune response and require more than one dose for effectiveness or some other adjuvants [114]. There are several safety issues regarding the use of attenuated vaccines, including the reactivation in vaccinated individuals, hence their use is not beneficial in the case of highly pathogenic viruses [112]. Attenuated vaccines are not suitable for individuals with a weak immune system, as they are more prone to infection if the virus reverts itself [115]. Moreover, preservation of LAVs requires ultra-cold temperatures and a sustained cold chain distribution system, which may not be available in all countries [114,115,116]. LAVs have undergone preclinical trials [117] and there is a company working on the development of a computationally designed virus that is non-pathogenic [118]. Another company is working on the formalin-inactivated SARS-CoV-2 virus and determined its efficacy in rhesus macaques [119]. This vaccine did not invite any pathological response; on the contrary, in the past, the LAV vaccine against SARS-CoV produced an eosinophil-derived immune response in mice. Histopathological changes were also observed in lungs derived from T helper 2 cells [120,121].
Protein subunit vaccines include antigenic proteins that are produced in vitro. Upon administration with adjuvants, these vaccines produce a strong humoral immune response and high immune memory [122]. Protein subunit vaccines include antigenic proteins and virus-like particles (VLPs), which produce many copies of antigens and produce strong immune responses without the help of adjuvants [122]. Worldwide institutions are working on protein subunit vaccines, since they are an attractive vaccine technology that does not require specific conditions for preservation [114]. However, the costs for mass production, specific mammalian optimization, and cell expression are important considerations [123,124].
Nucleic acid vaccines are produced by choosing specific proteins of pathogens (coding for their epitopes) to initiate an immune response in the host. These proteins are delivered into the host as DNA/RNA plasmid sequences [125,126,127]. Upon injection into host cells, the host cell machinery starts producing pathogen proteins which are recognized by the host immune system, in turn, initiating the production of antibodies against these proteins [127]. If RNA/DNA vaccine is non-capsulated, it will be rejected and removed from host cells soon after the injection. Advanced delivery technologies, such as encapsulation of RNA in liposomes, can be used to avoid the degradation of vaccines by host cells [128]. RNA vaccines produce T cell and antigen-specific antibody response against cancer in clinical trials. A mRNA-based vaccine known as the non-replicating rabies virus glycoprotein (RABV-G) stimulates stable and potent neutralizing antibodies in domestic pigs and mice. It produces functional antibodies against glycoprotein of rabies virus [115,129]. DNA vaccines are also immunogenic in the case of animals; however, in humans, they show a weak immune response and nesed multiple doses with adjuvants [130]. Four DNA vaccines are available for animal use but currently, there are no licensed DNA and RNA vaccines for humans [131]. Nucleic acid vaccines have been developed and some are still under development to protect against SARS-CoV-2 by several institutes such as Moderna (Cambridge, MA, USA), Inovio Pharmaceuticals (Plymouth Meeting, PA, USA), Pfizer (New York, NY, USA), the Imperial College London, Takis Biotech (Rome, Italy), and BIOCAD (St. Petersburg, Russia). The vaccine named BNT162b2 has been approved for delivery and sale in multiple countries. The studies carried out on this RNA vaccine to check its safety measures and immunogenicity in adults (from 18 to 55 years old), who were given 2 doses of vaccine with a 21 days interval, have revealed that neutralizing titers of SARS-CoV-2 and IgG concentrations binding with RBD (receptor binding domain) increased 1.9–4.6 fold in COVID-19 convalescent sera obtained at least 14 days after positive PCR [132]. Results of these studies support the public use of the mRNA vaccine [133]. Nucleic acid vaccines are easy to produce and relatively cheap with the possibility of mass production [134]. The minimum time required for a vaccine development and for its clinical trials is almost 1 year.
Last but not least, another platform for vaccine development is Recombinant viral vector vaccines. These vaccines contain live viruses that can replicate themselves, but they are engineered to carry some extra pathogenic genes of interest. These genes produce proteins after the injection of vaccines into the host. The host immune system produces antibodies against these proteins [135]. Challenges in the development of these vaccines elicit an immune response against the vector instead of an antigen-specific response, as well as the loss of extra viral genes during recombination procedures and during replication inside host cells. However, pre-clinical and clinical trials showed that one dose of such types of vaccines is enough to elicit an effective immune response [136]. Human adenoviruses (hAds) are potential recombinant viral vectors. However, due to their frequent spread in the population, pre-existing immunity makes vaccines less efficient [137,138]. Chimpanzee adenovirus (ChAd) was developed, which is more efficient than hAds due to its low prevalence, hence, acting as a neutralizing antibody [139,140]. Preclinical studies show that ChAd vectors are 100% efficient against emerging viruses and a single dose is sufficient to produce rapid immunity in individuals [138,141,142].
11. Current Status of COVID-19 Vaccine Development
Vaccination produces everlasting immunity in the host by introducing it to antigens and produce a kind of immunologic memory before encountering pathogens. SARS-CoV-2 gains entry into the host by its surface spike (S) protein [143]. This protein attaches with ACE2 (angiotensin-converting enzyme 2) receptor on host cells. These receptors are found abundantly on pulmonary epithelial cells [115]. Currently, vaccines are designed to target S protein on the viral surface. Just after the viral entry in the host, the S protein is recognized by the host immune system, which produces antibodies against both the S protein and nucleoprotein. The first genomic sequence of SARS-CoV-2 was published in January 2020, triggering global activity to develop a vaccine against the contagious virus. The first SARS-CoV-2 vaccine candidate was approved for human clinical testing on 16 March 2020. The Coalition for Epidemic Preparedness Innovations (CEPI) is collaborating with vaccine developers and global health authorities to develop a vaccine against SARS-CoV-2 [144]. On 8 April 2020, the global SARS-CoV-2 vaccine R&D landscape consisted of 115 vaccine candidates. Out of these 115 candidates, 37 were unconfirmed and 78 were confirmed and active. Platforms for vaccine development depend on mRNA and DNA providing potential and flexibility for antigen manipulation. The vaccines based on viral vectors often provide high protein expression, induce a strong immune response, and are highly stable. Vaccine developers (GlaxoSmithKline, Dynavax, and Seqirus) planned to use adjuvants as vaccines because of their ability to enhance immunity and the viability of low doses [144]. According to WHO, six vaccines were approved/authorized against SARS-CoV-2 for public use before 15 December 2020. These vaccines have been designed by using different techniques such as a m-RNA based vaccine (BNT162b2), Peptide vaccine (EpiVacCorona), inactivated virus vaccine (BBIBP-CorV and CoronaVac), and non-replicating viral vector (Sputnik V) vaccine. The SARS-CoV-2 has numerous variants, and it can mutate in an individual and a population over time. Therefore, many questions about the efficacy of vaccines rise with time. It has been reported that the maximum variants have a mutation in the D614G region and the ACE2 binding site is not affected. So, it can be stated that the vaccines developed for SARS-CoV-2 would be effective against its variants [145]. Some of the authorized/approved and candidate vaccines have been discussed in Table 6.
Table 6.
Some of authorized/approved and candidate vaccines against SARS-CoV-2 (more details can be found at: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/covid-19-vaccines (accessed on 1 February 2021).
12. Recommended Preventive Measures
Since SARS-CoV-2 is of a zoonotic origin, extensive measures are required to control person to person transfer and the rapid spread of the disease. Despite strict regulations, SARS-CoV-2 has spread very quickly globally. Special attention, care, and efforts are needed to reduce transmission among susceptible populations, including elder people, children, and health care workers. Preventive measures are the current master plan to reduce the number of active cases. A guideline was published for health care providers, researchers, medical staff, and the general public [146]. The early spread of disease was among the elderly population due to their weak immune systems and rapid onset of illness [147,148]. Various public services have provided decontaminating agents to people for hand cleansing on a routine basis. Care should be taken among health care societies while care should be exercised in dealing with virus samples such as urine and fecal samples that provide an alternative route of transmission [149,150]. Major preventive control measures have been implemented like the screening of travelers to minimize disease spreading all over the world.
Some general recommendations are issued by WHO as well as other organizations:
- Regular use of face masks [151].
- Wash your hands frequently and sanitize them after close contact with objects and patients. Isolate patients in a separate room and minimize visits to patients [152].
- Avoid personal contact with farm and wild animals [152].
- Avoid close contact with people that have any respiratory illness or symptoms [153].
- Specifically, people with weak immune systems should avoid public gatherings and healthy people should also avoid gatherings to minimize the chances of getting the disease [154].
- People with flu and a cough should avoid close contact with healthy people. While coughing and sneezing they should use disposable tissue/cloth and dispose of them properly. Afterward, wash hands frequently and use sanitizer [155].
- Strict hygiene rules should be followed in hospitals and other health care departments to avoid the spread of disease and to prevent infection [156].
- Some of the vaccines like Pfizer-BioNTech COVID-19 Vaccine and Moderna COVID-19 Vaccine are approved for emergency use; however, they have not been fully evaluated for efficacy against SARS-CoV-2 variants that recently emerged in the UK and South Africa [157].
The most important strategies which people should follow is to avoid public gatherings, use portable hand sanitizers, and avoid contact with the face, mouth, and nose after visiting contaminated areas such as hospitals and other health care units. For health care workers, care should be taken by wearing gloves, masks (FFP3 and N95), gowns, and eye protection to avoid transmission of the virus.
Since there is no proper treatment of SARS-CoV-2 available to everyone yet, only by following SOPs and by exercising personal care can the chances of getting an infection can be minimized. The general public should take vitamin-rich foods to enhance body immunity.
13. Conclusions
Re-emergence of the virus after modifications in its genome for stable adaptation is a serious concern to human health. SARS-CoV-2 has a zoonotic origin and changed its host from bats and animals to humans. In 2019, it appeared for the very first time, transmitted from human to human, and gradually spread to over 200 countries. This virus spread through close human contact by coughing, sneezing, and aerosols. The transmission was so rapid and widespread that controlling it became a difficult task. Through sequencing and phylogenetic analysis, researchers found that this virus is novel and there is no cure for its infection. Its genome has similarities with those of SARS-CoV and MERS-CoV but due to mutations somewhere in the genome, it became a novel virus. It causes infections with mild symptoms (fever, flu, and cough) to severe symptoms and clinical outcomes (ARDS, septic shock, respiratory failure, and Multi-organ dysfunction). There are multiple risk factors involved regarding the severity of disease like comorbidities, organ failures, diabetes, etc. The ratio of illness and deaths has been high among elder people due to having a weaker immune system. There is no proper treatment and no specific drugs against this virus to date. Although some vaccines have been approved/authorized against the virus, preventive strategies should still be adopted by the public to minimize the chance of getting the disease.
Author Contributions
Conceptualization, I.M., S.A., T.A., and M.T.u.Q.; data curation, I.M., and A.B.; writing—original draft preparation, I.M., M.I., and S.A.; writing—review and editing, T.A., A.B., A.A. (Asma Abro), K.S.A., A.A. (Ahmad Almatroudi), and M.T.u.Q.; supervision, A.A. (Ahmad Almatroudi), and M.T.u.Q.; project administration, A.A. (Asma Abro), T.A., A.A. (Ahmad Almatroudi), and M.T.u.Q.; funding acquisition, K.S.A. and A.A. (Ahmad Almatroudi). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article.
Acknowledgments
Authors would like to thank three anonymous reviewers for their constructive comments and valuable suggestions, which facilitated us in improving our work. In addition, authors apologize to all the researchers whose relevant work could not be cited due to space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sohrab, S.S.; Azhar, E.I. Genetic diversity of MERS-CoV spike protein gene in Saudi Arabia. J. Infect. Public Health 2020, 13, 709–717. [Google Scholar] [CrossRef]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus Pathogenesis. Adv. Virus Res. 2011, 81, 85–164. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Yu, F.; Du, L.; Ojcius, D.M.; Pan, C.; Jiang, S. Measures for diagnosing and treating infections by a novel coronavirus responsible for a pneumonia outbreak originating in Wuhan, China Fei. Microbes Infect. 2020, 22, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Cui, W.; Tian, B. The potential intermediate hosts for SARS-CoV-2. Front. Microbiol. 2020, 11, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Singh, R.; Dar, Z.; Bijarnia, R.K.; Dhingra, N.; Kaur, T. Genetic comparison among various coronavirus strains for the identification of potential vaccine targets of SARS-CoV2. Infect. Genet. Evol. 2020, 89, 104490. [Google Scholar] [CrossRef]
- Zhong, N.; Zheng, B.; Li, Y.; Poon, L.; Lancet, Z.X.-T. Epidemiology and Cause of Severe Acute Respiratory Syndrome (SARS) in Guangdong, People’s Republic of China, in February. Lancet 2003, 362, 1353–1358. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Günther, S.; Preiser, W.; Van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef]
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- WHO. Summary of Probable SARS Cases with Onset of Illness from 1 November 2002 to 31 July 2003. Available online: https://www.ci.nii.ac.jp (accessed on 1 January 2021).
- Assiri, A.; McGeer, A.; Perl, T.M.; Price, C.S.; Al Rabeeah, A.A.; Cummings, D.A.T.; Alabdullatif, Z.N.; Assad, M.; Almulhim, A.; Makhdoom, H.; et al. Hospital outbreak of middle east respiratory syndrome coronavirus. N. Engl. J. Med. 2013, 369, 407–416. [Google Scholar] [CrossRef]
- Wong, G.; Liu, W.; Liu, Y.; Zhou, B.; Bi, Y.; Microbe, G.G.-C. MERS, SARS, and Ebola: The Role of Super-Spreaders in Infectious Disease. Cell Host Micro. 2015, 18, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Guan, Y.; Yuen, K.Y. Severe acute respiratory syndrome. Nat. Med. 2004, 10, S88–S97. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Adair, B.D.; Yoshioka, C.; Quispe, J.D.; Orca, G.; Kuhn, P.; Milligan, R.A.; Yeager, M.; Buchmeier, M.J. Supramolecular Architecture of Severe Acute Respiratory Syndrome Coronavirus Revealed by Electron Cryomicroscopy. J. Virol. 2006, 80, 7918–7928. [Google Scholar] [CrossRef]
- Barcena, M.; Oostergetel, G.T.; Bartelink, W.; Faas, F.G.A.; Verkleij, A.; Rottier, P.J.M.; Koster, A.J.; Bosch, B.J. Cryo-electron tomography of mouse hepatitis virus: Insights into the structure of the coronavirion. Proc. Natl. Acad. Sci. USA 2009, 106, 582–587. [Google Scholar] [CrossRef]
- Tsai, P.H.; Wang, M.L.; Yang, D.M.; Liang, K.H.; Chou, S.J.; Chiou, S.H.; Lin, T.H.; Wang, C.T.; Chang, T.J. Genomic variance of Open Reading Frames (ORFs) and Spike protein in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). J. Chin. Med. Assoc. 2020, 83, 725–732. [Google Scholar] [CrossRef]
- Alamri, M.A.; Qamar, M.T.U.; Mirza, M.U.; Alqahtani, S.M.; Froeyen, M.; Chen, L.-L. Discovery of human coronaviruses pan-papain-like protease inhibitors using computational approaches. J. Pharm. Anal. 2020, 10, 546–559. [Google Scholar] [CrossRef]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef]
- Alamri, M.A.; Qamar, M.T.U.; Mirza, M.U.; Bhadane, R.; Alqahtani, S.M.; Muneer, I.; Froeyen, M.; Salo-Ahen, O.M.H. Pharmacoinformatics and molecular dynamics simulation studies reveal potential covalent and FDA-approved inhibitors of SARS-CoV-2 main protease 3CLpro. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef]
- Angeletti, S.; Benvenuto, D.; Bianchi, M.; Giovanetti, M.; Pascarella, S.; Ciccozzi, M. COVID-2019: The role of the nsp2 and nsp3 in its pathogenesis. J. Med. Virol. 2020, 92, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Qamar, M.T.U.; Rehman, A.; Tusleem, K.; Ashfaq, U.A.; Qasim, M.; Zhu, X.; Fatima, I.; Shahid, F.; Chen, L.L. Designing of a next generation multiepitope based vaccine (MEV) against SARS-COV-2: Immunoinformatics and in silico approaches. PLoS ONE 2020, 15, e0244176. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef]
- Qamar, M.T.U.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Muhseen, Z.T.; Hameed, A.R.; Al-Hasani, H.M.H.; Qamar, M.T.U.; Li, G. Promising terpenes as SARS-CoV-2 spike receptor-binding domain (RBD) attachment inhibitors to the human ACE2 receptor: Integrated computational approach. J. Mol. Liq. 2020, 320, 114493. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784. [Google Scholar] [CrossRef]
- Ren, L.L.; Wang, Y.M.; Wu, Z.Q.; Xiang, Z.C.; Guo, L.; Xu, T.; Jiang, Y.Z.; Xiong, Y.; Li, Y.J.; Li, X.W.; et al. Identification of a novel coronavirus causing severe pneumonia in human: A descriptive study. Chin. Med. J. 2020, 133, 1015–1024. [Google Scholar] [CrossRef]
- Gioia, M.; Ciaccio, C.; Calligari, P.; Simone, G.D.; Sbardella, D.; Tundo, G.; Francesco, G.; Di, A.; Di, D. Role of proteolytic enzymes in the COVID-19 infection and promising therapeutic approaches. Biochem. Pharmacol. 2020, 182, 1–22. [Google Scholar] [CrossRef]
- Ceraolo, C.; Giorgi, F.M. Genomic variance of the 2019-nCoV coronavirus. J. Med. Virol. 2020, 92, 522–528. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Li, K.S.M.; Tsang, A.K.L.; Lam, C.S.F.; Ahmed, S.; Chen, H.; Chan, K.-H.; Woo, P.C.Y.; Yuen, K.-Y. Genetic Characterization of Betacoronavirus Lineage C Viruses in Bats Reveals Marked Sequence Divergence in the Spike Protein of Pipistrellus Bat Coronavirus HKU5 in Japanese Pipistrelle: Implications for the Origin of the Novel Middle East Respiratory Sy. J. Virol. 2013, 87, 8638–8650. [Google Scholar] [CrossRef]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.J.; Jiang, S. The spike protein of SARS-CoV—A target for vaccine and therapeutic development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Pachetti, M.; Marini, B.; Benedetti, F.; Giudici, F.; Mauro, E.; Storici, P.; Masciovecchio, C.; Angeletti, S.; Ciccozzi, M.; Gallo, R.C.; et al. Emerging SARS-CoV-2 mutation hot spots include a novel RNA-dependent-RNA polymerase variant. J. Transl. Med. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Sevajol, M.; Subissi, L.; Decroly, E.; Canard, B.; Imbert, I. Insights into RNA synthesis, capping, and proofreading mechanisms of SARS-coronavirus. Virus Res. 2014, 194, 90–99. [Google Scholar] [CrossRef]
- Pathan, R.K.; Biswas, M.; Khandaker, M.U. Time series prediction of COVID-19 by mutation rate analysis using recurrent neural network-based LSTM model. Chaos Solitons Fractals 2020, 138, 110018. [Google Scholar] [CrossRef]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–560. [Google Scholar] [CrossRef]
- Phan, T. Genetic diversity and evolution of SARS-CoV-2. Infect. Genet. Evol. 2020, 81, 104260. [Google Scholar] [CrossRef]
- Rouchka, E.C.; Chariker, J.H.; Chung, D. Variant analysis of 1040 SARS-CoV-2 genomes. PLoS ONE 2020, 15, e0241535. [Google Scholar] [CrossRef]
- Wise, J. Covid-19: New coronavirus variant is identified in UK. BMJ 2020, 371, m4857. [Google Scholar] [CrossRef]
- Qamar, M.T.U.; Shahid, F.; Aslam, S.; Ashfaq, U.A.; Aslam, S.; Fatima, I.; Fareed, M.M.; Zohaib, A.; Chen, L.-L. Reverse vaccinology assisted designing of multiepitope-based subunit vaccine against SARS-CoV-2. Infect. Dis. Poverty 2020, 9, 1–14. [Google Scholar]
- Azer, S.A. COVID-19: Pathophysiology, diagnosis, complications and investigational therapeutics. New Microbes New Infect. 2020, 37, 100738. [Google Scholar] [CrossRef] [PubMed]
- Worldometer Coronavirus Cases. Available online: https://www.worldometers.info/coronavirus/ (accessed on 1 January 2021).
- Yanez, N.D.; Weiss, N.S.; Romand, J.A.; Treggiari, M.M. COVID-19 mortality risk for older men and women. BMC Public Health 2020, 20, 1–7. [Google Scholar] [CrossRef]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Ko, J.-H.; Joo, E.-J.; Park, S.-J.; Baek, J.Y.; Kim, W.D.; Jee, J.; Kim, C.J.; Jeong, C.; Kim, Y.-J.; Shon, H.J. Neutralizing antibody production in asymptomatic and mild COVID-19 patients, in comparison with pneumonic COVID-19 patients. J. Clin. Med. 2020, 9, 2268. [Google Scholar] [CrossRef]
- Mokhtari, T.; Hassani, F.; Ghaffari, N.; Ebrahimi, B.; Yarahmadi, A.; Hassanzadeh, G. COVID-19 and multiorgan failure: A narrative review on potential mechanisms. J. Mol. Histol. 2020, 51, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Hiyoshi, A.; Montgomery, S. COVID-19 case-fatality rate and demographic and socioeconomic influencers: Worldwide spatial regression analysis based on country-level data. BMJ Open 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases from the Chinese Center for Disease Control and Prevention. JAMA J. Am. Med. Assoc. 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Gandhi, A.; Sokhi, J.; Lockie, C.; Ward, P.A. Emergency Tracheal Intubation in Patients with COVID-19: Experience from a UK Centre. Anesthesiol. Res. Pract. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Meng, L.; Qiu, H.; Wan, L.; Ai, Y.; Xue, Z.; Guo, Q.; Deshpande, R.; Zhang, L.; Meng, J.; Tong, C.; et al. Intubation and Ventilation amid the COVID-19 Outbreak: Wuhan’s Experience. Anesthesiology 2020, 132, 1317–1332. [Google Scholar] [CrossRef] [PubMed]
- Storch, G.A. Diagnostic virology. Clin. Infect. Dis. 2000, 31, 739–751. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Z.; Jin, L.; Chu, F.; Mao, Y.; Wang, H.; Liu, M.; Wang, M.; Zhang, L.; Gao, G.F.; et al. Persistent shedding of viable SARS-CoV in urine and stool of SARS patients during the convalescent phase. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 165–171. [Google Scholar] [CrossRef]
- Oh, M.; Park, W.B.; Choe, P.G.; Choi, S.-J.; Kim, J.-I.; Chae, J.; Park, S.S.; Kim, E.-C.; Oh, H.S.; Kim, E.J.; et al. Viral Load Kinetics of MERS Coronavirus Infection. N. Engl. J. Med. 2016, 375, 1303–1305. [Google Scholar] [CrossRef]
- Corman, V.M.; Albarrak, A.M.; Omrani, A.S.; Albarrak, M.M.; Farah, M.E.; Almasri, M.; Muth, D.; Sieberg, A.; Meyer, B.; Assiri, A.M.; et al. Viral Shedding and Antibody Response in 37 Patients With Middle East Respiratory Syndrome Coronavirus Infection. Clin. Infect. Dis. 2015, 62, civ951. [Google Scholar] [CrossRef]
- Writing Committee of the WHO Consultation on Clinical Aspects of Pandemic (H1N1) 2009 Influenza. Clinical aspects of pandemic 2009 influenza A (H1N1) virus infection. N. Engl. J. Med. 2010, 362, 1708–1719. [Google Scholar] [CrossRef]
- Di Castelnuovo, A.; Bonaccio, M.; Costanzo, S.; Gialluisi, A.; Antinori, A.; Berselli, N.; Blandi, L.; Bruno, R.; Cauda, R.; Guaraldi, D.; et al. Common cardiovascular risk factors and in-hospital mortality in 3,894 patients with COVID-19: Survival analysis and machine learning-based findings from the multicentre Italian CORIST Study. Nutr. Metabol. Cardiovas. Dis. 2020, 30, 1899–1913. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef]
- Sprung, C.L.; Schein, R.M.H.; Balk, R.A. The new sepsis consensus definitions: The good, the bad and the ugly. Intensive Care Med. 2016, 42, 2024–2026. [Google Scholar] [CrossRef] [PubMed]
- Lopes Ferreira, F.; Peres Bota, D.; Bross, A.; Mélot, C.; Vincent, J.L. Serial evaluation of the SOFA score to predict outcome in critically ill patients. J. Am. Med. Assoc. 2001, 286, 1754–1758. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Wang, Y.; Liu, Y.; Liu, X.; Gu, L.; Zhang, X.; Pu, Z.; Yang, G.; Liu, B.; Mie, Q.; et al. Disease Severity and Clinical Outcomes of Community Acquired Pneumonia Caused by Non-Influenza Respiratory Viruses: A Multicenter Prospective Registry Study from CAP-China Network. Eur. Resp. J. 2019, 54, 1802406. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.W.M.; Ng, C.K.; Chan, Y.H.; Mok, Y.W.; Lee, S.; Chu, Y.; Law, W.L.; Lee, M.P. Short term outcome and risk factors for adverse clinical outcomes in adults with severe acute respiratory syndrome (SARS). Thorax 2003, 58, 686–689. [Google Scholar] [CrossRef]
- Arabi, Y.M.; Balkhy, H.H.; Hayden, F.G.; Bouchama, A.; Luke, T.; Baillie, J.K.; Al-Omari, A.; Hajeer, A.H.; Senga, M.; Denison, M.R.; et al. Middle east respiratory syndrome. N. Engl. J. Med. 2017, 376, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; Girard, T.D.; Wesley Ely, E. The Immunopathogenesis of Sepsis in Elderly Patients. Clin. Infec. Dis. 2005, 41, S504–S512. [Google Scholar] [CrossRef] [PubMed]
- Marrie, T.J.; Shariatzadeh, M.R. Community-Acquired Pneumonia Requiring Admission to an Intensive Care Unit. Medicine 2007, 86, 103–111. [Google Scholar] [CrossRef]
- Corrales-Medina, V.F.; Musher, D.M.; Shachkina, S.; Chirinos, J.A. Acute pneumonia and the cardiovascular system. Lancet 2013, 381, 496–505. [Google Scholar] [CrossRef]
- Lei, J.; Kusov, Y.; Hilgenfeld, R. Nsp3 of coronaviruses: Structures and functions of a large multi-domain protein. Antivir. Res. 2018, 149, 58–74. [Google Scholar] [CrossRef]
- Milbrandt, E.B.; Reade, M.C.; Lee, M.; Shook, S.L.; Angus, D.C.; Kong, L.; Carter, M.; Yealy, D.M.; Kellum, J.A. Prevalence and significance of coagulation abnormalities in community-acquired pneumonia. Mol. Med. 2009, 15, 438–445. [Google Scholar] [CrossRef]
- Rodelo, J.R.; De La Rosa, G.; Valencia, M.L.; Ospina, S.; Arango, C.M.; Gómez, C.I.; García, A.; Nuñez, E.; Jaimes, F.A. D-dimer is a significant prognostic factor in patients with suspected infection and sepsis. Am. J. Emerg. Med. 2012, 30, 1991–1999. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, J.; Bao, L.; Shi, Y. Convalescent plasma as a potential therapy for COVID-19. Lancet Infect. Dis. 2020, 20, 398–400. [Google Scholar] [CrossRef]
- Khalili, J.S.; Zhu, H.; Mak, N.S.A.; Yan, Y.; Zhu, Y. Novel coronavirus treatment with ribavirin: Groundwork for an evaluation concerning COVID-19. J. Med. Virol. 2020, 92, 740–746. [Google Scholar] [CrossRef]
- Lu, H. Drug treatment options for the 2019-new coronavirus (2019-nCoV). Biosci. Trends 2020, 14, 69–71. [Google Scholar] [CrossRef]
- Prescott, H.C.; Rice, T.W. Corticosteroids in COVID-19 ARDS: Evidence and Hope during the Pandemic. JAMA J. Am. Med. Assoc. 2020, 324, 1292–1295. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Tan, Q.; Duan, L.; Ma, Y.; Wu, F.; Huang, Q.; Mao, K.; Xiao, W.; Xia, H.; Zhang, S.; Zhou, E.; et al. Is oseltamivir suitable for fighting against COVID-19: In silico assessment, in vitro and retrospective study. Bioorg. Chem. 2020, 104, 104257. [Google Scholar] [CrossRef] [PubMed]
- Marciniec, K.; Beberok, A.; Pęcak, P.; Boryczka, S.; Wrześniok, D. Ciprofloxacin and moxifloxacin could interact with SARS-CoV-2 protease: Preliminary in silico analysis. Pharmacol. Rep. 2020, 72, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Sultana, J.; Cutroneo, P.M.; Crisafulli, S.; Puglisi, G.; Caramori, G.; Trifirò, G. Azithromycin in COVID-19 Patients: Pharmacological Mechanism, Clinical Evidence and Prescribing Guidelines. Drug Saf. 2020, 43, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Gotte, M. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus. J. Biol. Chem. 2020, 295, 4773–4779. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus susceptibility to the antiviral remdesivir (GS-5734) is mediated by the viral polymerase and the proofreading exoribonuclease. MBio 2018, 9, e00221-18. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Yan, Y.; Zou, Z.; Sun, Y.; Li, X.; Xu, K.F.; Wei, Y.; Jin, N.; Jiang, C. Anti-malaria drug chloroquine is highly effective in treating avian influenza A H5N1 virus infection in an animal model. Cell Res. 2013, 23, 300–302. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Zhonghua, J.; He, H.; Hu, X.; Za, Z. Expert consensus on chloroquine phosphate for the treatment of novel coronavirus pneumonia. Bibl. Nac. Med. 2020, 43, 185–188. [Google Scholar]
- Gautret, P.; Lagier, J.-C.; Parola, P.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; Dupont, H.T. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, A.; Shaikh, A.; Singh, R.; Misra, A. Chloroquine and hydroxychloroquine in the treatment of COVID-19 with or without diabetes: A systematic search and a narrative review with a special reference to India and other developing countries. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Ye, F.; Zhang, M.; Cui, C.; Huang, B.; Niu, P.; Liu, X.; Zhao, L.; Dong, E.; Song, C. In vitro antiviral activity and projection of optimized dosing design of hydroxychloroquine for the treatment of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Clin. Infect. Dis. 2020, 71, 732–739. [Google Scholar] [CrossRef]
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cvetkovic, R.S.; Goa, K.L. Lopinavir/ritonavir: A review of its use in the management of HIV infection. Drugs 2003, 63, 769–802. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.M.; Cheng, C.C.; Hung, F.N.; Wong, M.M.L.; Chan, H.; Chan, S.; Kao, Y.T.; Poon, L.L.M.; Wong, L.P.; Guan, Y.; et al. Role of lopinavir/ritonavir in the treatment of SARS: Initial virological and clinical findings. Thorax 2004, 59, 252–256. [Google Scholar] [CrossRef]
- Arabi, Y.M.; Asiri, A.Y.; Assiri, A.M.; Aziz Jokhdar, H.A.; Alothman, A.; Balkhy, H.H.; AlJohani, S.; Al Harbi, S.; Kojan, S.; Al Jeraisy, M.; et al. Treatment of Middle East respiratory syndrome with a combination of lopinavir/ritonavir and interferon-β1b (MIRACLE trial): Statistical analysis plan for a recursive two-stage group sequential randomized controlled trial. Trials 2020, 21, 8. [Google Scholar] [CrossRef]
- Lim, J.; Jeon, S.; Shin, H.Y.; Kim, M.J.; Seong, Y.M.; Lee, W.J.; Choe, K.W.; Kang, Y.M.; Lee, B.; Park, S.J. Case of the index patient who caused tertiary transmission of coronavirus disease 2019 in Korea: The application of lopinavir/ritonavir for the treatment of COVID-19 pneumonia monitored by quantitative RT-PCR. J. Korean Med. Sci. 2020, 35, e79. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, X.; Lu, Y.; Chen, F.; Zhang, W. Clinical characteristics and therapeutic procedure for four cases with 2019 novel coronavirus pneumonia receiving combined Chinese and Western medicine treatment. Biosci. Trends 2020, 14, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Piao, W.; Shirey, K.A.; Ru, L.W.; Lai, W.; Szmacinski, H.; Snyder, G.A.; Sundberg, E.J.; Lakowicz, J.R.; Vogel, S.N.; Toshchakov, V.Y. A Decoy Peptide that Disrupts TIRAP Recruitment to TLRs Is Protective in a Murine Model of Influenza. Cell Rep. 2015, 11, 1941–1952. [Google Scholar] [CrossRef]
- Perrin-Cocon, L.; Aublin-Gex, A.; Sestito, S.E.; Shirey, K.A.; Patel, M.C.; André, P.; Blanco, J.C.; Vogel, S.N.; Peri, F.; Lotteau, V. TLR4 antagonist FP7 inhibits LPS-induced cytokine production and glycolytic reprogramming in dendritic cells, and protects mice from lethal influenza infection. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Lansbury, L.; Rodrigo, C.; Leonardi-Bee, J.; Nguyen-Van-Tam, J.; Lim, W.S. Corticosteroids as adjunctive therapy in the treatment of influenza. Cochrane Database Syst. Rev. 2019. [Google Scholar] [CrossRef]
- Baughn, L.B.; Sharma, N.; Elhaik, E.; Sekulic, A.; Bryce, A.H.; Fonseca, R. Targeting TMPRSS2 in SARS-CoV-2 Infection. Mayo Clin. Proc. 2020, 95, 1989–1999. [Google Scholar] [CrossRef]
- Li, G.; De Clercq, E. Therapeutic options for the 2019 novel coronavirus (2019-nCoV). Nat. Rev. Drug Discov. 2020, 19, 149–150. [Google Scholar] [CrossRef]
- Zumla, A.; W Chan, J.F.; Azhar, E.I.; C Hui, D.S.; Yuen, K.-Y. Coronaviruses—Drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A.; et al. First case of 2019 novel coronavirus in the United States. N. Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef]
- Beigel, J.H.; Nam, H.H.; Adams, P.L.; Krafft, A.; Ince, W.L.; El-Kamary, S.S.; Sims, A.C. Advances in Respiratory Virus Therapeutics–A Meeting Report from the 6th Isirv Antiviral Group Conference. Antivir. Res. 2019, 167, 45–67. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.; Griffin, I.; Tucker, C.; Smith, D.; Oechsle, O.; Phelan, A.; Stebbing, J. Baricitinib as potential treatment for 2019-nCoV acute respiratory disease. Lancet 2020, 395, e30–e31. [Google Scholar] [CrossRef]
- Hoang, B.X.; Shaw, D.G.; Fang, W.; Han, B. A Possible application of high dose vitamin C in the prevention and therapy for Coronavirus Infections. J. Glob. Antimicrob. Resist. 2020, 23, 256–262. [Google Scholar] [CrossRef]
- Abobaker, A.; Alzwi, A.; Alraied, A.H.A. Overview of the possible role of vitamin C in management of COVID-19. Pharmacol. Rep. 2020, 72, 1517–1528. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T. The human microbiome: Implications for health and disease, including HIV infection. Top. Antivir. Med. 2018, 26, 75–78. [Google Scholar] [PubMed]
- Saul, A.W. Nutritional treatment of coronavirus. Orthomol. Med. News Serv. 2020, 16, 22. [Google Scholar]
- Skrajnowska, D.; Bobrowska-Korczak, B. Role of zinc in immune system and anti-cancer defense mechanisms. Nutrients 2019, 11, 2273. [Google Scholar] [CrossRef]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2120. [Google Scholar] [CrossRef] [PubMed]
- Harthill, M. Review: Micronutrient selenium deficiency influences evolution of some viral infectious diseases. Biol. Trace Elem. Res. 2011, 143, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Stachowska, E.; Folwarski, M.; Jamioł-Milc, D.; Maciejewska, D.; Skonieczna-Żydecka, K. Nutritional support in coronavirus 2019 disease. Medicina (B Aires) 2020, 56, 289. [Google Scholar] [CrossRef]
- Lauring, A.S.; Jones, J.O.; Andino, R. Rationalizing the development of live attenuated virus vaccines. Nat. Biotechnol. 2010, 28, 573–579. [Google Scholar] [CrossRef]
- Pulendran, B.; Ahmed, R. Immunological mechanisms of vaccination. Nat. Immunol. 2011, 12, 509–517. [Google Scholar] [CrossRef]
- Lee, J.; Arun Kumar, S.; Jhan, Y.Y.; Bishop, C.J. Engineering DNA vaccines against infectious diseases. Acta Biomater. 2018, 80, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, H.R.; Gilbride, C.; Allen, E.; Belij-Rammerstorfer, S.; Bissett, C.; Ewer, K.; Lambe, T. The early landscape of coronavirus disease 2019 vaccine development in the UK and rest of the world. Immunology 2020, 160, 223–232. [Google Scholar] [CrossRef]
- Haque, A.; Pant, A.B. Efforts at COVID-19 Vaccine Development: Challenges and Successes. Vaccines 2020, 8, 739. [Google Scholar] [CrossRef]
- Luo, F.; Liao, F.L.; Wang, H.; Tang, H.B.; Yang, Z.Q.; Hou, W. Evaluation of Antibody-Dependent Enhancement of SARS-CoV Infection in Rhesus Macaques Immunized with an Inactivated SARS-CoV Vaccine. Virol. Sin. 2018, 33, 201–204. [Google Scholar] [CrossRef]
- Lin, J.T.; Zhang, J.S.; Su, N.; Xu, J.G.; Wang, N.; Chen, J.T.; Chen, X.; Liu, Y.X.; Gao, H.; Jia, Y.P.; et al. Safety and immunogenicity from a Phase I trial of inactivated severe acute respiratory syndrome coronavirus vaccine. Antivir. Ther. 2007, 12, 1107–1113. [Google Scholar]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.T.; Sbrana, E.; Iwata-Yoshikawa, N.; Newman, P.C.; Garron, T.; Atmar, R.L.; Peters, C.J.; Couch, R.B. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Bolles, M.; Deming, D.; Long, K.; Agnihothram, S.; Whitmore, A.; Ferris, M.; Funkhouser, W.; Gralinski, L.; Totura, A.; Heise, M.; et al. A Double-Inactivated Severe Acute Respiratory Syndrome Coronavirus Vaccine Provides Incomplete Protection in Mice and Induces Increased Eosinophilic Proinflammatory Pulmonary Response upon Challenge. J. Virol. 2011, 85, 12201–12215. [Google Scholar] [CrossRef]
- Karch, C.; Burkhard, P. Vaccine Technologies: From Whole Organisms to Rationally Designed Protein Assemblies. Biochem. Pharmacol. 2016, 120, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, S.; Robinson, J.; Cunningham, G.; Vaccine, R.I. The Complexity and Cost of Vaccine Manufacturing–An Overview. Vaccine 2017, 35, 4064–4071. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Cannon, G.; Weissman, D. RNA based vaccines. DNA Cell Biol. 2002, 21, 953–961. [Google Scholar] [CrossRef]
- Blakney, A.K.; McKay, P.F.; Christensen, D.; Yus, B.I.; Aldon, Y.; Follmann, F.; Shattock, R.J. Effects of Cationic Adjuvant Formulation Particle Type, Fluidity and Immunomodulators on Delivery and Immunogenicity of saRNA. J. Control. Rel. 2019, 304, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Geall, A.; Mandl, C.; Ulemr, J.B. RNA: The New Revolution in Nucleic Acid Vaccines. Sem. Immunol. 2013, 25, 152–159. [Google Scholar] [CrossRef]
- Ulmer, J.; Mason, P.; Geall, A.; Vaccine, C.M. RNA-Based Vaccines. Vaccine 2012, 30, 4414–4418. [Google Scholar] [CrossRef] [PubMed]
- Schnee, M.; Vogel, A.B.; Voss, D.; Petsch, B.; Baumhof, P.; Kramps, T.; Stitz, L. An mRNA vaccine encoding rabies virus glycoprotein induces protection against lethal infection in mice and correlates of protection in adult and newborn pigs. PLoS Negl. Trop. Dis. 2016, 10, e0004746. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Louder, M.; Holman, L.; Vaccine, I.G. A SARS DNA Vaccine Induces Neutralizing Antibody and Cellular Immune Responses in Healthy Adults in a Phase I Clinical Trial. Vaccine 2008, 26, 6338–6343. [Google Scholar] [CrossRef]
- Kramps, T.; Elbers, K. Introduction to RNA vaccines. In Methods in Molecular Biology; Humana Press Inc.: New York, USA, 2017; Volume 1499, pp. 1–11. [Google Scholar]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Lee, L.Y.Y.; Izzard, L.; Hurt, A.C. A review of DNA vaccines against influenza. Front. Immunol. 2018, 9, 1568. [Google Scholar] [CrossRef]
- Bull, J.J.; Nuismer, S.L.; Antia, R. Recombinant vector vaccine evolution. PLoS Comput. Biol. 2019, 15, e1006857. [Google Scholar] [CrossRef] [PubMed]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef]
- Jartti, T.; Jartti, L.; Ruuskanen, O.; Söderlund-Venermo, M. New Respiratory Viral Infections. Curr. Opin. Pulm. Med. 2012, 18, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Z.; Gao, H.; Li, Y.; Hou, L.; Yao, H.; Wu, S.; Liu, J.; Wang, L.; Zhai, Y.; et al. Open-label phase I clinical trial of Ad5-EBOV in Africans in China. Hum. Vaccin. Immunother. 2017, 13, 2078–2085. [Google Scholar] [CrossRef]
- Dudareva, M.; Andrews, L.; Gilbert, S.; Bejon, P.; Vaccine, K.M. Prevalence of Serum Neutralizing Antibodies against Chimpanzee Adenovirus 63 and Human Adenovirus 5 in Kenyan Children, in the Context of Vaccine Vector. Vaccine 2009, 27, 3501–3504. [Google Scholar] [CrossRef]
- Morris, S.J.; Sebastian, S.; Spencer, A.J.; Gilbert, S.C. Simian adenoviruses as vaccine vectors. Future Virol. 2016, 11, 649–659. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Haddock, E.; Feldmann, F.; Meade-White, K.; Bushmaker, T.; Fischer, R.J.; Okumura, A.; Hanley, P.W.; Saturday, G.; Edwards, N.J.; et al. A Single Dose of ChAdOx1 MERS Provides Protective Immunity in Rhesus Macaques. Sci. Adv. 2020, 6, eaba8399. [Google Scholar] [CrossRef]
- Stanley, D.A.; Honko, A.N.; Asiedu, C.; Trefry, J.C.; Lau-Kilby, A.W.; Johnson, J.C.; Hensley, L.; Ammendola, V.; Abbate, A.; Grazioli, F.; et al. Chimpanzee adenovirus vaccine generates acute and durable protective immunity against ebolavirus challenge. Nat. Med. 2014, 20, 1126–1129. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Gui, M.; Wang, X.; Xiang, Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLoS Pathog. 2018, 14, e1007236. [Google Scholar] [CrossRef]
- Le, T.T.; Andreadakis, Z.; Kumar, A.; Gómez Román, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Dearlove, B.; Lewitus, E.; Bai, H.; Li, Y.; Reeves, D.B.; Joyce, M.G.; Scott, P.T.; Amare, M.F.; Vasan, S.; Michael, N.L.; et al. A SARS-CoV-2 vaccine candidate would likely match all currently circulating variants. Proc. Natl. Acad. Sci. USA 2020, 117, 23652–23662. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.H.; Cai, L.; Cheng, Z.S.; Cheng, H.; Deng, T.; Fan, Y.P.; Fang, C.; Huang, D.; Huang, L.Q.; Huang, Q.; et al. A rapid advice guideline for the diagnosis and treatment of 2019 novel coronavirus (2019-nCoV) infected pneumonia (standard version). Mil. Med. Res. 2020, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.L.; McNamara, M.S.; Sinclair, D.A. Why does COVID-19 disproportionately affect older people? Aging 2020, 12, 9959. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tang, J.; Wei, F. Updated understanding of the outbreak of 2019 novel coronavirus (2019-nCoV) in Wuhan, China. J. Med. Virol. 2020, 92, 441–447. [Google Scholar] [CrossRef]
- Lee, N.; Hui, D.; Wu, A.; Chan, P.; Cameron, P.; Joynt, G.M.; Ahuja, A.; Yung, M.Y.; Leung, C.B.; To, K.F.; et al. A major outbreak of severe acute respiratory syndrome in Hong Kong. N. Engl. J. Med. 2003, 348, 1986–1994. [Google Scholar] [CrossRef]
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F.; et al. Epidemiological, Demographic, and Clinical Characteristics of 47 Cases of Middle East Respiratory Syndrome Coronavirus Disease from Saudi Arabia: A Descriptive Study. Lancet Infec. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef]
- Eikenberry, S.E.; Mancuso, M.; Iboi, E.; Phan, T.; Eikenberry, K.; Kuang, Y.; Kostelich, E.; Gumel, A.B. To mask or not to mask: Modeling the potential for face mask use by the general public to curtail the COVID-19 pandemic. Infect. Dis. Model. 2020, 5, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Finset, A.; Bosworth, H.; Butow, P.; Gulbrandsen, P.; Hulsman, R.L.; Pieterse, A.H.; Street, R.; Tschoetschel, R.; van Weert, J. Effective health communication—A key factor in fighting the COVID-19 pandemic. Patient Educ. Couns. 2020, 103, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Graham Carlos, W.; Dela Cruz, C.S.; Cao, B.; Pasnick, S.; Jamil, S. Novel Wuhan (2019-NCoV) coronavirus. Am. J. Respir. Crit. Care Med. 2020, 201, P7–P8. [Google Scholar] [CrossRef]
- Chakraborty, I.; Maity, P. COVID-19 outbreak: Migration, effects on society, global environment and prevention. Sci. Total Environ. 2020, 728, 138882. [Google Scholar] [CrossRef]
- Ali, I.; Alharbi, O.M.L. COVID-19: Disease, management, treatment, and social impact. Sci. Total Environ. 2020, 728, 138861. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Maqsood, A.; Abduljabbar, T.; Vohra, F. Tobacco smoking a potential risk factor in transmission of COVID-19 infection. Pakistan J. Med. Sci. 2020, 36, S104–S107. [Google Scholar] [CrossRef]
- Knoll, M.D.; Wonodi, C. Oxford–AstraZeneca COVID-19 vaccine efficacy. Lancet 2021, 397, 72–74. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).