Prolongation of Tick-Borne Encephalitis Cycles in Warmer Climatic Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Epidemiological Data
2.2. Statistical Analysis
3. Results
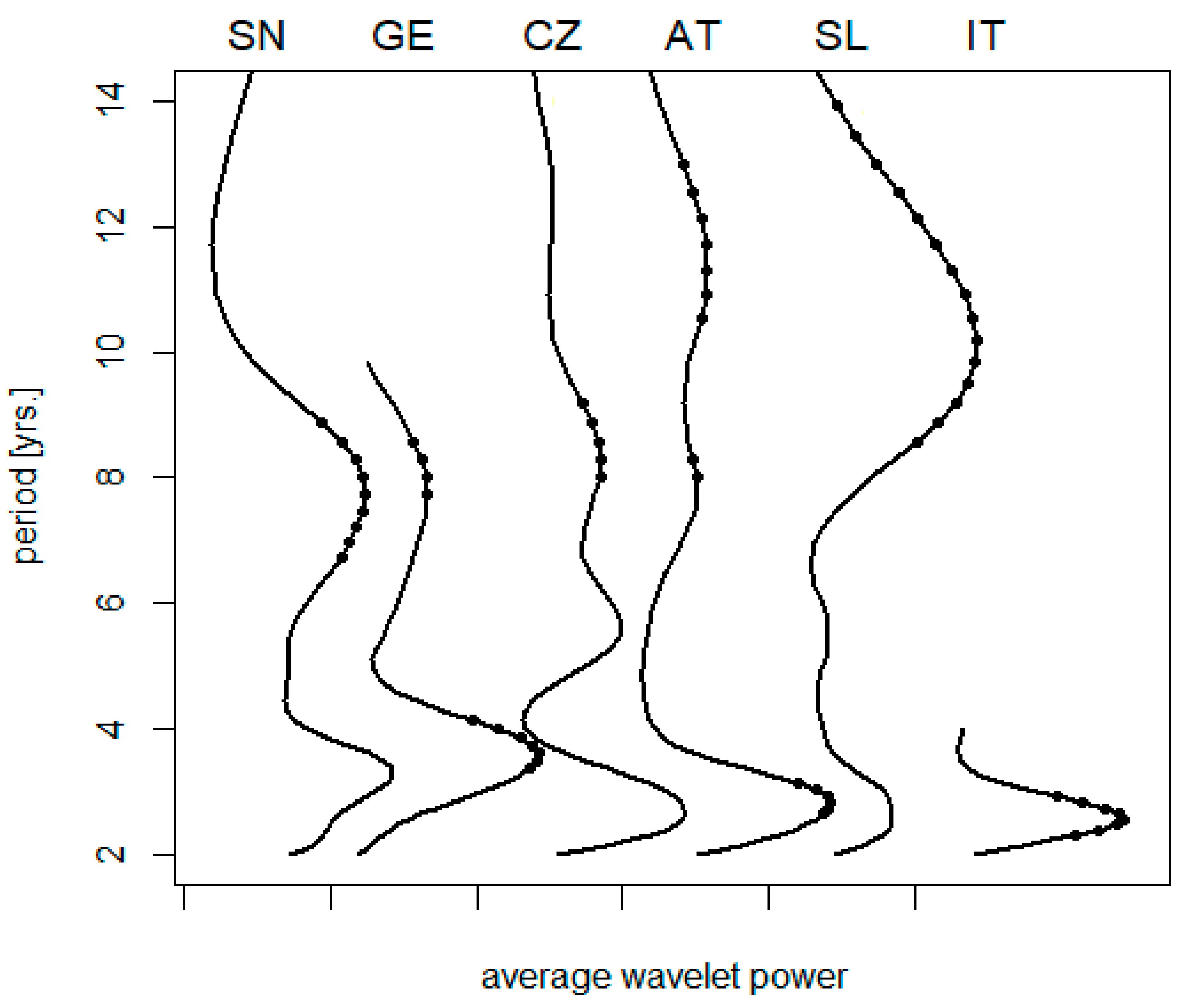
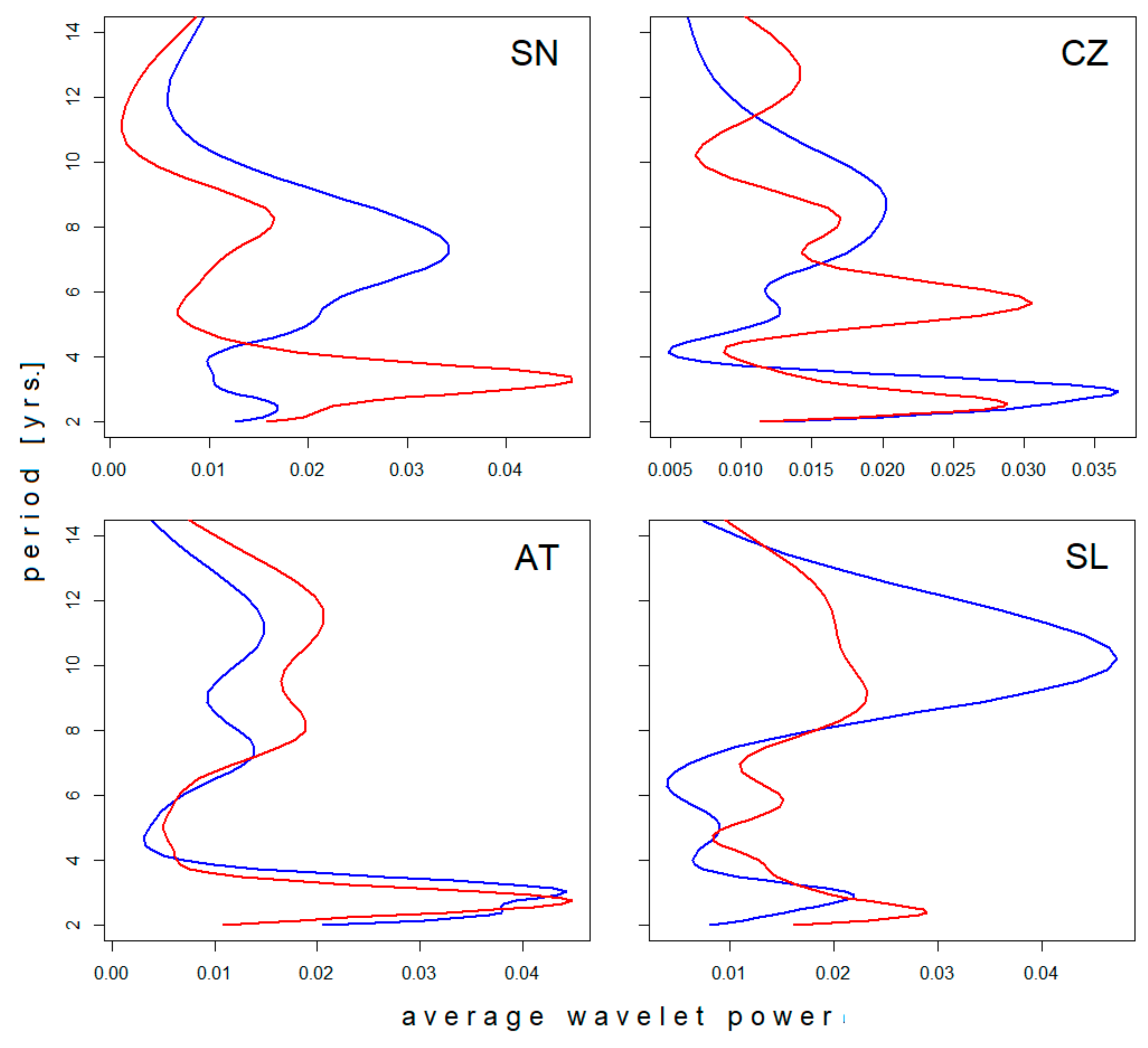
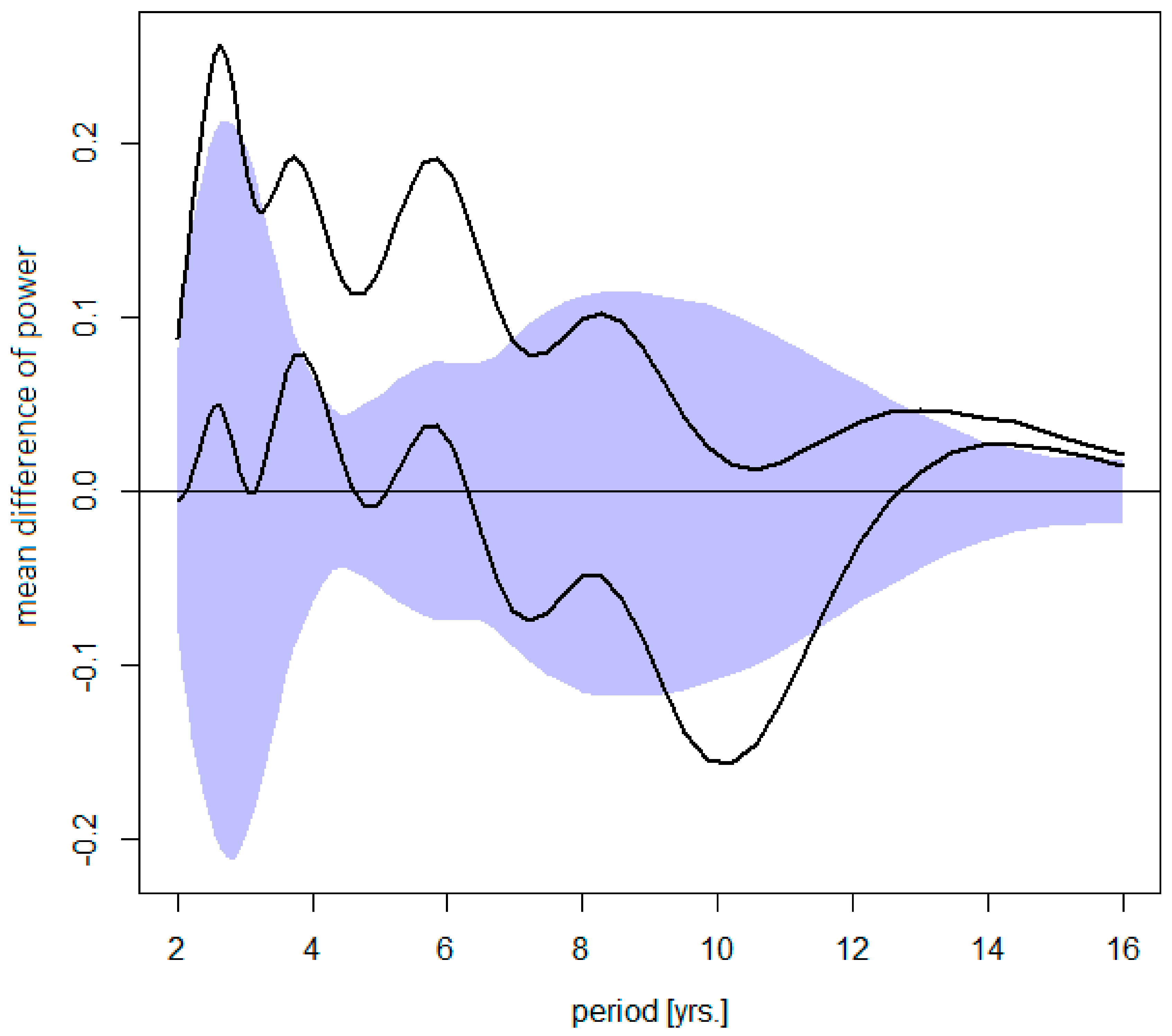
3.1. Effects of Latitude
3.2. Longitudinal Shifts
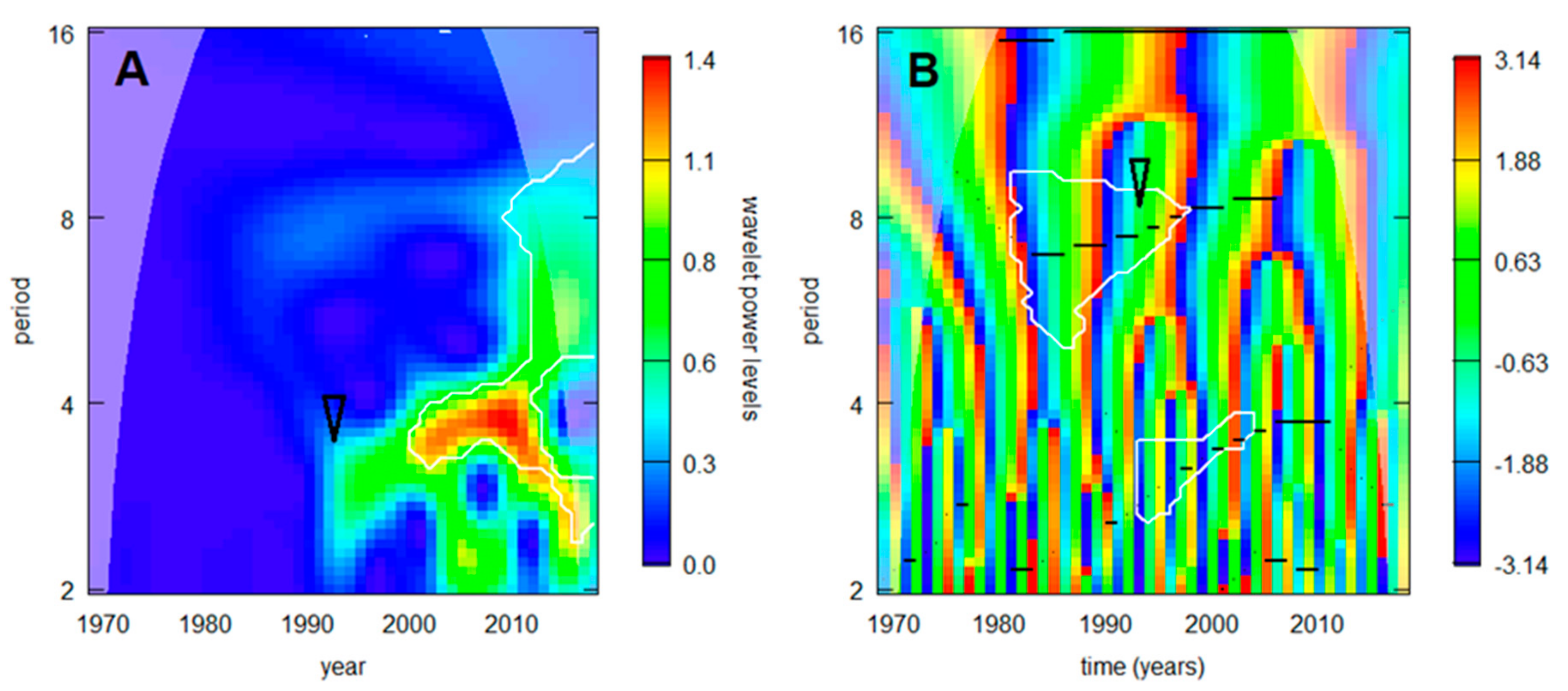
3.3. Local Effects
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
References
- Kaiser, R. Tick-borne encephalitis. Infect. Dis. Clin. N. Am. 2008, 22, 561–575. [Google Scholar] [CrossRef]
- Beauté, J.; Spiteri, G.; Warns-Petit, E.; Zeller, H. Tick-borne encephalitis in Europe, 2012 to 2016. Euro Surveill. 2018, 23, 1800201. [Google Scholar] [CrossRef] [PubMed]
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-borne encephalitis. Antivir. Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Suess, J. Epidemiology and ecology of TBE relevant to the production of effective vaccines. Vaccine 2003, 21, S19–S35. [Google Scholar] [CrossRef]
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Ruzek, D.; Avsic-Zupanc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Haemig, P.D.; de Luna, S.S.; Grafström, A.; Lithner, S.; Lundkvist, A.; Waldenström, J.; Kindberg, J.; Stedt, J.; Olsén, B. Forecasting risk of tick-borne encephalitis (TBE): Using data from wildlife and climate to predict next year’s number of human victims. Scand. J. Infect. Dis. 2011, 43, 366–372. [Google Scholar] [CrossRef]
- Palo, R.T. Tick-borne encephalitis transmission risk: Its dependence on host population dynamics and climate effects. Vector Borne Zoon. Dis. 2014, 14, 346–352. [Google Scholar] [CrossRef]
- Brugger, K.; Walter, M.; Chitimia-Dobler, L.; Dobler, G.; Rubel, F. Forecasting next season’s Ixodes ricinus nymphal density: The example of southern Germany 2018. Exp. Appl. Acarol. 2018, 75, 281–288. [Google Scholar] [CrossRef]
- Zeman, P. Cyclic patterns in the central European tick-borne encephalitis incidence series. Epidemiol. Infect. 2017, 145, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Zeman, P. Predictability of tick-borne encephalitis fluctuations. Epidemiol. Infect. 2017, 145, 2781–2786. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cazelles, B.; Chavez, M.; McMichael, A.J.; Hales, S. Nonstationary influence of El Niño on the synchronous dengue epidemics in Thailand. PLoS Med. 2005, 2, e106. [Google Scholar] [CrossRef]
- Kreppel, K.S.; Caminade, C.; Telfer, S.; Rajerison, M.; Rahalison, L.; Morse, A.; Baylis, M. A Non-Stationary Relationship between Global Climate Phenomena and Human Plague Incidence in Madagascar. PLoS Negl. Trop. Dis. 2014, 8, e3155. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Yu, P.; Cazelles, B.; Xu, L.; Tan, H.; Yang, J.; Huang, S.; Xu, B.; Cai, J.; Ma, C.; et al. Interannual cycles of Hantaan virus outbreaks at the human–animal interface in Central China are controlled by temperature and rainfall. PNAS 2017, 114, 8041–8046. [Google Scholar] [CrossRef]
- Ims, R.A.; Henden, J.-A.; Killengreen, S.T. Collapsing population cycles. Trends Ecol. Evol. 2007, 23, 79–86. [Google Scholar] [CrossRef]
- ISW-TBE. International Scientific Working Group on TBE. Available online: http://www.tbe-info.com (accessed on 15 April 2002).
- Lundkvist, A.; Wallensten, A.; Vene, S.; Hjertqvist, M. Tick-borne encephalitis increasing in Sweden, 2011. Euro Surveill. 2011, 16, 19981. [Google Scholar] [CrossRef]
- Suess, J.; Schrader, C.; Falk, U.; Wohanka, N. Tick-borne encephalitis (TBE) in Germany—Epidemiological data, development of risk areas and virus prevalence in field-collected ticks and in ticks removed from humans. Int. J. Med. Microbiol. 2004, 293, 69–79. [Google Scholar] [CrossRef]
- PFIZER. FSME Erkrankungsfälle in Österreich. Available online: https://www.pfizer.at/fileadmin/content/Content/04_Presse/04.3_Pressemappen/FSME_in_Oesterreich/FSME_Situationsbericht_2017_Endversion.pdf (accessed on 11 March 2018).
- Rezza, G.; Farchi, F.; Pezzott, P.; Ruscio, M.; Lo Presti, A.; Ciccozzi, M.; Mondardini, V.; Paternoster, C.; Bassetti, M.; Merelli, M.; et al. Tick-borne encephalitis in north-east Italy: A 14-year retrospective study, January 2000 to December 2013. Euro Surveill. 2015, 20, 30034. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.E.; Shen, Z.; Long, S.R.; Wu, M.C.; Shih, H.H.; Zheng, Q.; Yen, N.-C.; Tung, C.C.; Liu, H.H. The empirical mode decomposition and the Hilbert spectrum for nonlinear and non-stationary time series analysis. Proc. R. Soc. Lond. Ser. A 1998, 454, 903–995. [Google Scholar] [CrossRef]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Donghoh, K.; Hee-Seok, O. EMD: A package for empirical mode decomposition and Hilbert spectrum. R J. 2009, 1, 40–46. [Google Scholar]
- Roesch, A.; Schmidbauer, H. WaveletComp 1.1: A guided tour through the R package. Available online: http://www.hs-stat.com/projects/WaveletComp/WaveletComp_guided_tour.pdf (accessed on 18 March 2018).
- EDJNet. Europe one degree warmer. Available online: https://www.onedegreewarmer.eu (accessed on 23 August 2019).
- Randolph, S.E. Human activities predominate in determining changing incidence of tick-borne encephalitis in Europe. Euro Surveill. 2010, 15, 24–31. [Google Scholar] [CrossRef]
- Jaenson, T.G.T.; Petersson, E.H.; Jaenson, D.G.E.; Kindberg, J.; Pettersson, J.H.O.; Hjertqvist, M.; Medlock, J.M.; Bengtsson, H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: Incidence of human TBE correlates with abundance of deer and hares. Parasit. Vectors 2018, 11, 477. [Google Scholar] [CrossRef]
- Hubalek, Z. North Atlantic weather oscillation and human infectious diseases in the Czech Republic, 1951-2003. Eur. J. Epidemiol. 2005, 20, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Dizij, A.; Kurtenbach, K. Clethrionomys glareolus, but not Apodemus flavicollis, acquires resistance to lxodes ricinus L, the main European vector of Borrelia burgdorferi. Parasit. Immunol. 1995, 17, 177–183. [Google Scholar] [CrossRef]
- Awerbuch, T.E.; Sandberg, S. Trends and oscillations in tick population dynamics. J. Theor. Biol. 1995, 175, 511–516. [Google Scholar] [CrossRef]
- Fan, G.; Thieme, H.R.; Zhu, H. Delay differential systems for tick population dynamics. J. Math. Biol. 2015, 71, 1017–1048. [Google Scholar] [CrossRef]
- Grzybek, M.; Alsarraf, M.; Tolkacz, K.; Behnke-Borowczyk, J.; Biernat, B.; Stanczak, J.; Strachecka, A.; Guz, L.; Szczepaniak, K.; Paleolog, P.; et al. Seroprevalence of TBEV in bank voles from Poland—A long-term approach. Emerg. Microb. Infect. 2018, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gerth, H.-J.; Grimshandl, D.; Stage, B.; Döller, G.; Kunz, C. Roe deer as sentinels for endemicity of tick-borne encephalitis virus. Epidemiol. Infect. 1995, 115, 355–365. [Google Scholar] [CrossRef]
- Martello, E.; Mannelli, A.; Ragagli, C.; Ambrogi, C.; Selmi, M.; Ceballos, L.A.; Tomassone, L. Range expansion of Ixodes ricinus to higher altitude, and co-infestation of small rodents with Dermacentor marginatus in the Northern Apennines, Italy. Ticks Tick Borne Dis. 2014, 5, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.T.; Jaenson, D.G.E.; Eisen, L.; Petersson, E.; Lindgren, E. Changes in the geographical distribution and abundance of the tick Ixodes ricinus during the past 30 years in Sweden. Parasit. Vectors 2012, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Peters, W.; Hebblewhite, M.; Mysterud, A.; Spitz, D.; Focardi, S.; Urbano, F.; Morellet, N.; Heurich, M.; Kjellander, P.; Linnell, J.D.C.; et al. Migration in geographic and ecological space by a large herbivore. Ecol. Monogr. 2017, 87, 297–320. [Google Scholar] [CrossRef]
- Davis, M.L.; Stephens, P.A.; Kjellander, P. Beyond climate envelope projections: Roe deer survival and environmental change. J. Wildl. Manag. 2016, 80, 452–464. [Google Scholar] [CrossRef]
- Gaillard, J.-M.; Hewison, A.J.M.; Klein, F.; Plard, F.; Douhard, M.; Davison, R.; Bonenfant, C. How does climate change influence demographic processes of widespread species? Lessons from the comparative analysis of contrasted populations of roe deer. Ecol. Lett. 2013, 16, 48–57. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Centroid Northing | Centroid Easting | Period | Cases | Authority | References |
|---|---|---|---|---|---|---|
| Sweden | 59.5 | 15.0 | 1969–2018 | 5369 | Public Health Agency of Sweden (Folkhälsomyndigheten) | [16,17] |
| E. Germany 1 | 52.5 | 13.4 | 1993–2017 | 201 | Robert Koch Institute | [18] |
| Czech Rep. | 49.8 | 15.5 | 1971–2018 | 22,716 | National Institute of Public Health (SZU) | |
| Austria | 47.7 | 14.9 | 1970–2018 | 9746 | Zentrum für Virologie, Med. Universität Wien | [19] |
| Slovenia | 46.2 | 14.9 | 1972–2018 | 8710 | National Institute of Public Health (NIJZ) | [16] |
| NE Italy 2 | 46.2 | 12.2 | 2000–2013 | 367 | n.a. | [20] |
| Region/Country | Site | Temperature | No. Freezing Days/y | ||||
|---|---|---|---|---|---|---|---|
| 1900–1999 | 2000–2018 | Increase | 1900–1999 | 2000–2018 | Decrease | ||
| Sweden | Uppsala | 5.9 | 7 | 1.1 | 77.5 | 60.6 | 16.9 |
| Stockholm | 6.4 | 7.5 | 1.1 | 63.3 | 49.4 | 13.9 | |
| E Germany | Berlin | 9.2 | 10.2 | 1 | 35.6 | 27.2 | 8.4 |
| Dresden | 7.9 | 9.1 | 1.2 | 48.2 | 37.2 | 11 | |
| Czech Rep. | Prague | 8 | 9.1 | 1.1 | 49.1 | 41.1 | 8 |
| Brno | 8.6 | 9.6 | 1 | 46.2 | 40.3 | 5.9 | |
| Austria | Vienna | 9 | 10.1 | 1.1 | 43.1 | 35.3 | 7.8 |
| Graz | 8.1 | 8.9 | 0.8 | 53.4 | 46.3 | 7.1 | |
| Slovenia | Maribor | 9.5 | 10.4 | 0.9 | 40.1 | 34.6 | 5.5 |
| Ljubljana | 8.8 | 9.9 | 1.1 | 46.9 | 36.8 | 10.1 | |
| NE Italy | Belluno | 7.8 | 8.9 | 1.1 | 51.3 | 37.1 | 14.2 |
| Trieste | 11.4 | 12.6 | 1.2 | 20.9 | 9.8 | 11.1 | |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeman, P. Prolongation of Tick-Borne Encephalitis Cycles in Warmer Climatic Conditions. Int. J. Environ. Res. Public Health 2019, 16, 4532. https://doi.org/10.3390/ijerph16224532
Zeman P. Prolongation of Tick-Borne Encephalitis Cycles in Warmer Climatic Conditions. International Journal of Environmental Research and Public Health. 2019; 16(22):4532. https://doi.org/10.3390/ijerph16224532
Chicago/Turabian StyleZeman, Petr. 2019. "Prolongation of Tick-Borne Encephalitis Cycles in Warmer Climatic Conditions" International Journal of Environmental Research and Public Health 16, no. 22: 4532. https://doi.org/10.3390/ijerph16224532
APA StyleZeman, P. (2019). Prolongation of Tick-Borne Encephalitis Cycles in Warmer Climatic Conditions. International Journal of Environmental Research and Public Health, 16(22), 4532. https://doi.org/10.3390/ijerph16224532