Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint


Abstract
1. Introduction
2. Before and After Aflatoxin Discovery
2.1. Early Knowledge of Fungal Toxigenicity
2.2. Aflatoxins as a Group of Chemically Related Poisons: The Birth of Mycotoxicology
2.3. Causality between Aflatoxins and Liver Cancer
3. Challenges and Prospects for Aflatoxin Research
3.1. Technical Challenges
3.1.1. Analytical Methods and Challenges for Future Development
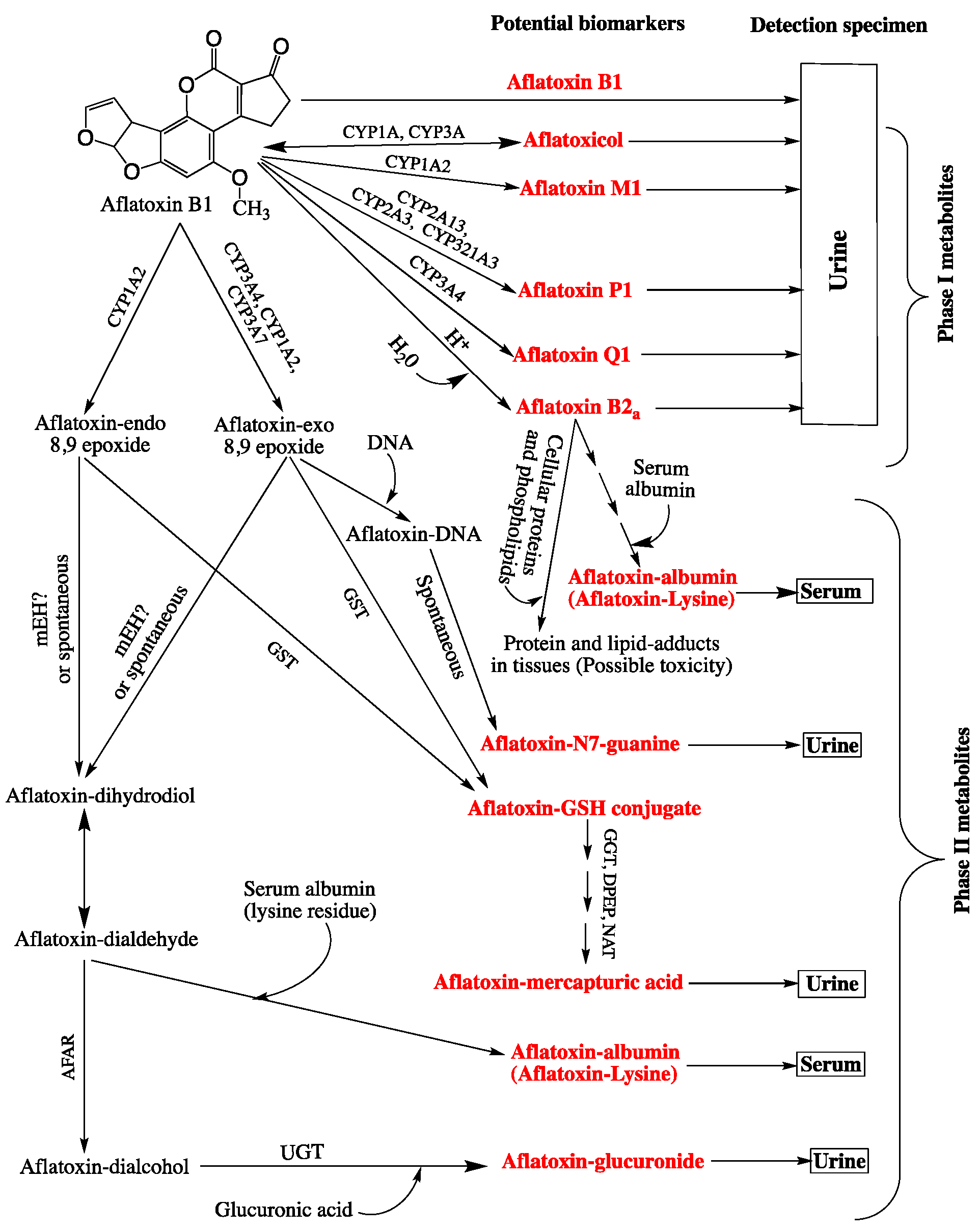
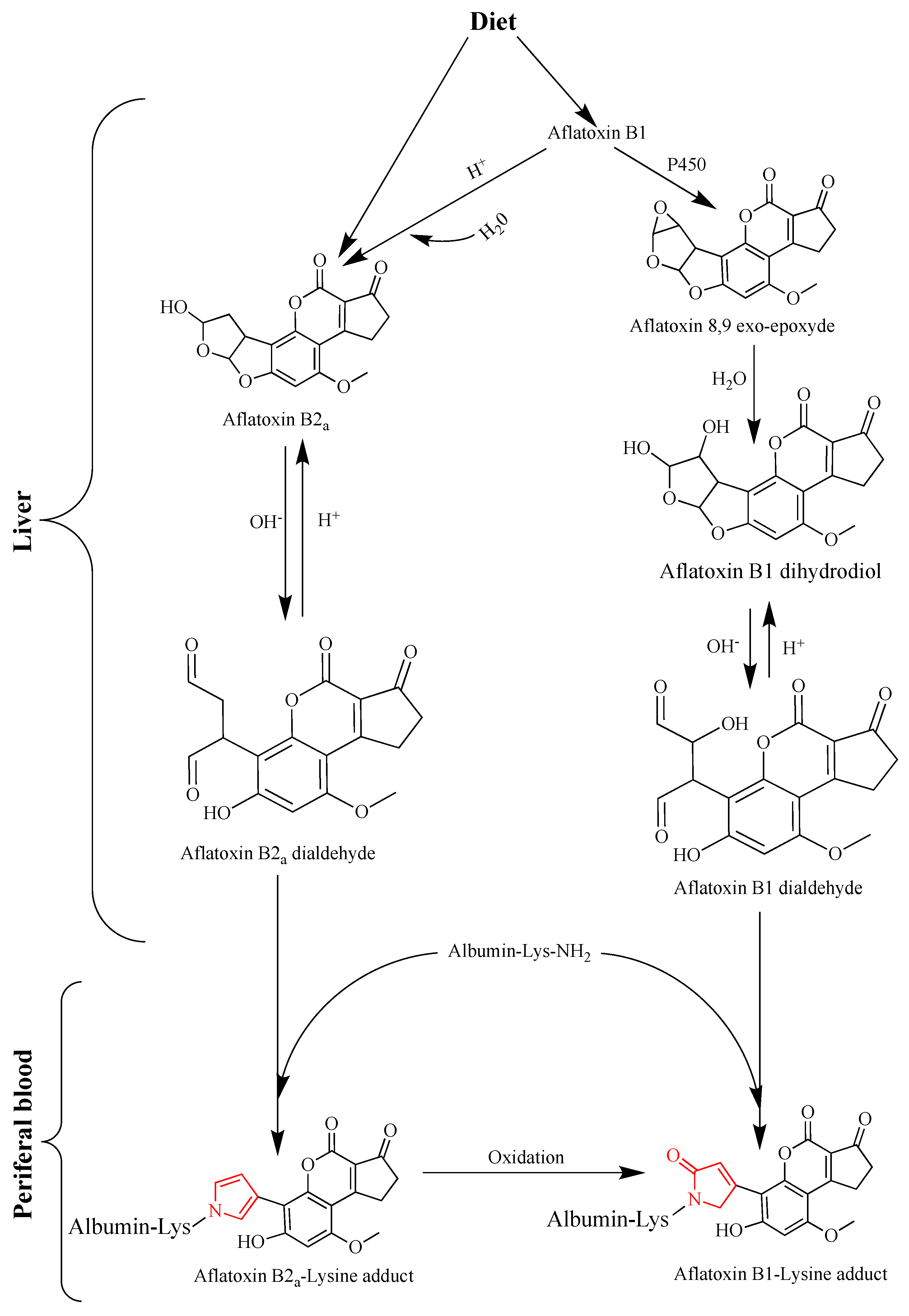
3.1.2. Dosimetry of Aflatoxins and Risk Prediction with Biomarkers
3.2. Natural and Socioeconomic Challenges
3.3. Actions to Mitigate Aflatoxin-Induced Health Risks
3.3.1. At Commodities’ Level: Reducing the Dietary Intake
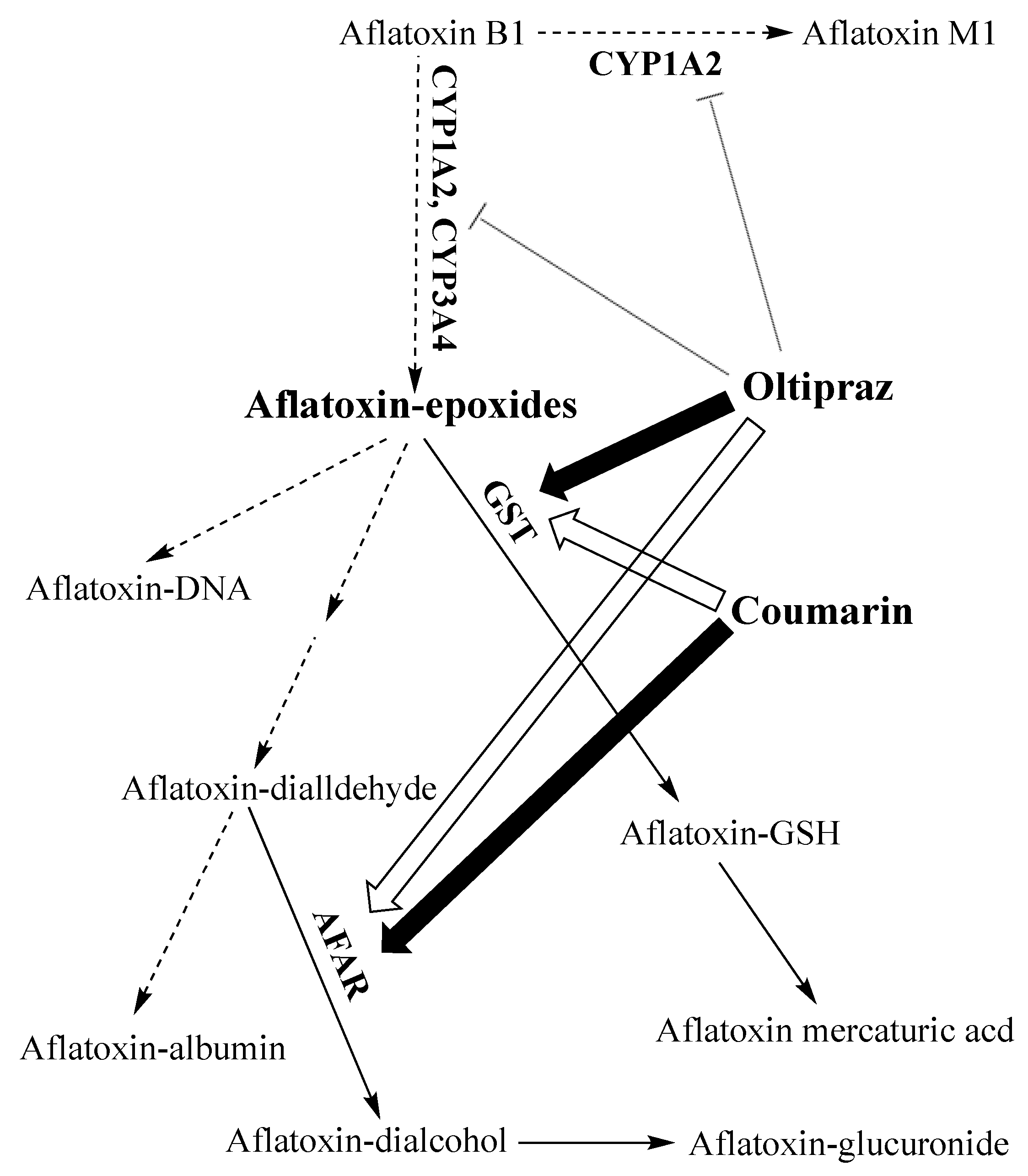
3.3.2. At the Consumer Level: Chemoprotection/Chemoprevention
Reducing the Risk by Interfering with the Toxicity of Aflatoxins
Reducing the Risk by Interfering with the Bioavailability of Aflatoxins
3.4. Legislative Challenges
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Sayiner, M.; Golabi, P.; Younossi, Z.M. Disease burden of hepatocellular carcinoma: A global perspective. Dig. Dis. Sci. 2019, 64, 910–917. [Google Scholar] [CrossRef]
- Blount, W.P. Turkey “x” disease. J. Br. Turk. Fed 1961, 9, 52–54. [Google Scholar]
- Pitt, J.I.; Miller, J.D. A concise history of mycotoxin research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef]
- Yao, H.; Hruska, Z.; Di Mavungu, J.D. Developments in detection and determination of aflatoxins. World Mycotoxin J. 2015, 8, 181–191. [Google Scholar] [CrossRef]
- Scanes, C.G. The neolithic revolution, animal domestication, and early forms of animal agriculture. In Animals and Human Society; Scanes, C.G., Toukhsati, S.R., Eds.; Academic Press: London, UK, 2018; pp. 103–131. [Google Scholar] [CrossRef]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef]
- Schiff, P.L. Ergot and its alkaloids. Am. J. Pharm. Educ. 2006, 70, 98. [Google Scholar] [CrossRef]
- Hofmann, A. Historical view on ergot alkaloids. Pharmacology 1978, 16 (Suppl. 1), 1–11. [Google Scholar] [CrossRef]
- Rosenthal, B. Salem Story: Reading the Witch Trials of 1692; Cambridge University Press: New York, NY, USA, 1993; p. 290. [Google Scholar]
- Chu, F.S. Mycotoxins: Toxicology. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 4096–4108. [Google Scholar] [CrossRef]
- Bové, F.J. The Story of Ergot: The Life Cycle of Ergot; Karger: Basel, Switzerland, 1970; pp. 15–25. [Google Scholar] [CrossRef]
- Wenzell, W.T. An essay on the active constituents of ergot of rye. Am. J. Pharm. 1864, 12, 193–202. [Google Scholar]
- Clark, A.J. Do the pharmacopœal preparations of ergot contain any active principles? Trans. Edinb. Obstet. Soc. 1927, 47, 109–121. [Google Scholar]
- Barger, G.; Carr, F.H. The alkaloids of ergot. J. Chem. Soc. Trans. 1907, 91, 337–353. [Google Scholar] [CrossRef]
- Jacobs, W.A.; Craig, L.C. The ergot alkaloids: VI. Lysergic acid. J. Biol. Chem. 1935, 111, 455–465. [Google Scholar]
- Barger, G. The alkaloids of ergot. Analyst 1937, 62, 340–354. [Google Scholar]
- Ramos, A.; Sanchis, V.; Marín, S. The prehistory of mycotoxins: Related cases from ancient times to the discovery of aflatoxins. World Mycotoxin J. 2011, 4, 101–112. [Google Scholar] [CrossRef]
- Sargeant, K.; Sheridan, A.N.N.; O’Kelly, J.; Carnaghan, R.B.A. Toxicity associated with Certain Samples of Groundnuts. Nature 1961, 192, 1096–1097. [Google Scholar] [CrossRef]
- Loosmore, R.M.; Markson, L.M. Poisoning of cattle by Brazilian groundnut meal. Vet. Rec. 1961, 73, 813–814. [Google Scholar]
- Asplin, F.D.; Carnaghan, R.B.A. The toxicity of certain groundnut meals for poultry with special reference to their effect on ducklings and chickens. Vet. Rec. 1961, 73, 1215–1219. [Google Scholar]
- Loosmore, R.M.; Harding, J.D.J. A toxic factor in Brazilian groundnut causing liver damage in pigs. Vet. Rec. 1961, 73, 1362–1364. [Google Scholar]
- Sargeant, K.; O’Kelly, J.; Carnaghan, R.B.A.; Allcroft, R. The assay of a toxic principle in certain groundnut meals. Vet. Rec. 1961, 73, 1219–1223. [Google Scholar]
- Carnaghan, R.B.A.; Sargeant, K. The toxicity of certain ground nut meals to poultry. Vet. Rec. 1961, 73, 726–727. [Google Scholar]
- Sargeant, K.; Carnaghan, R.B.A. Groundnut toxicity in poultry: Experimental and chemical aspects. Br. Vet. J. 1963, 119, 178–184. [Google Scholar] [CrossRef]
- Allcroft, R.; Carnaghan, R.B.A.; Sargeant, K.; O’Kelly, J. A toxic factor in Brazilian groundnut meal. Vet. Rec. 1961, 73, 428–429. [Google Scholar]
- Campbell, J.G. VI.—An Investigation of the Hepatotoxic effects in the fowl of ragwort (Senecio jacobæa Linn.), with special reference to the induction of liver tumours with seneciphylline. Proc. R. Soc. Edinb. B Biol. Sci. 1956, 66, 111–130. [Google Scholar] [CrossRef]
- Sargeant, K.; Allcroft, R.; Carnaghan, R.B.A. Groundnut toxicity. Vet. Rec. 1961, 73, 865. [Google Scholar]
- Goldblatt, L.A. Aflatoxin: Scientific Background, Control, and Implications; Academic Press Inc.: New York, NY, USA; London, UK, 1969; Chapter 1; pp. 1–11. [Google Scholar]
- Van Der Zijden, A.S.M.; Koelensmid, W.A.A.B.; Boldingh, J.; Barrett, C.B.; Ord, W.O.; Philp, J. Aspergillus flavus and turkey X disease: Isolation in crystalline form of a toxin responsible for turkey X disease. Nature 1962, 195, 1060–1062. [Google Scholar] [CrossRef]
- Nesbitt, B.F.; O’Kelly, J.; Sargeant, K.; Sheridan, A.N.N. Aspergillus flavus and turkey X disease: Toxic metabolites of Aspergillus flavus. Nature 1962, 195, 1062–1063. [Google Scholar] [CrossRef]
- Smith, R.H.; McKernan, W. Hepatotoxic action of chromatographically separated fractions of Aspergillus flavus extracts. Nature 1962, 195, 1301–1303. [Google Scholar] [CrossRef]
- de Iongh, H.; Beerthuis, R.K.; Vles, R.O.; Barrett, C.B.; Ord, W.O. Investigation of the factor in groundnut meal responsible for “turkey X disease”. Biochim. Biophys. Acta 1962, 65, 549–551. [Google Scholar] [CrossRef]
- Hartley, R.D.; Nesbitt, B.F.; O’Kelly, J. Toxic metabolites of Aspergillus flavus. Nature 1963, 198, 1056–1058. [Google Scholar] [CrossRef]
- Asao, T.; Buchi, G.; Abdel-Kader, M.M.; Chang, S.B.; Wick, E.L.; Wogan, G.N. Aflatoxins B and G. J. Am. Chem. Soc. 1963, 85, 1706–1707. [Google Scholar] [CrossRef]
- van Dorp, D.A.; van Der Zijden, A.S.M.; Beerthuis, R.K.; Sparreboom, S.; Ord, W.O.; de Jong, K.; Keuning, R. Dihydro-aflatoxin B, a metabolite of Aspergillus flavus. Remarks on the structure of aflatoxin B. Recl. Trav. Chim. Pays Bas 1963, 82, 587–592. [Google Scholar] [CrossRef]
- Allcroft, R.; Carnaghan, R.B.A. Groundnut toxicity. Aspergillus flavus toxin (aflatoxin) in animal products: Preliminary communication. Vet. Rec. 1962, 74, 863–864. [Google Scholar]
- Allcroft, R.; Carnaghan, R.B.A. Groundnut toxicity: An examination for toxin in human food products from animals fed toxic groundnut meal. Vet. Rec. 1963, 75, 259–263. [Google Scholar]
- Allcroft, R.; Rogers, H.; Lewis, G.; Nabney, J.; Best, P.E. Metabolism of aflatoxin in sheep: Excretion of the “milk toxin”. Nature 1966, 209, 154–155. [Google Scholar] [CrossRef]
- Holzapfel, C.W.; Steyn, P.S.; Purchase, I.F. Isolation and structure of aflatoxins M1 and M2. Tetrahedron Lett. 1966, 25, 2799–2803. [Google Scholar] [CrossRef]
- de Iongh, H.; Vles, R.O.; van Pelt, J.G. Milk of mammals fed an aflatoxin-containing diet. Nature 1964, 202, 466–467. [Google Scholar] [CrossRef]
- Dutton, M.F.; Heathcote, J.G. The structure, biochemical properties and origin of the aflatoxins B2a and G2a. Chem. Ind. 1968, 13, 418–421. [Google Scholar]
- Lillehoj, E.B.; Ciegler, A. Biological activity of aflatoxin B2a. Appl. Microbiol. 1969, 17, 516–519. [Google Scholar]
- Dalezios, J.; Wogan, G.N.; Weinreb, S.M. Aflatoxin P1: A new aflatoxin metabolite in monkeys. Science 1971, 171, 584. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Wendiro, D.; Vuzi, P.C.; Hawumba, J.F. Methods for detection of aflatoxins in agricultural food crops. J. Appl. Chem. 2014, 2014, 706291. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef]
- Oettlé, A.G. Cancer in africa, especially in regions South of the sahara. J. Natl. Cancer Inst. 1964, 33, 383–439. [Google Scholar]
- Harington, J.S.; McGlashan, N.D.; Bradshaw, E.; Geddes, E.W.; Purves, L.R. A spatial and temporal analysis of four cancers in African gold miners from Southern Africa. Br. J. Cancer 1975, 31, 665–678. [Google Scholar] [CrossRef]
- Keen, P.; Martin, P. Is aflatoxin carcinogenic in man? The evidence in Swaziland. Trop. Geogr. Med. 1971, 23, 44–53. [Google Scholar]
- Lopez, A.; Crawford, M.A. Aflatoxin content of groundnuts sold for human consumption in uganda. Lancet 1967, 290, 1351–1354. [Google Scholar] [CrossRef]
- Barnes, J.M. Aflatoxin as a health hazard. J. Appl. Bacteriol. 1970, 33, 285–298. [Google Scholar] [CrossRef]
- Payet, M.; Cros, J.; Quenum, C.; Sankale, M.; Moulanier, M. Deux observations d’enfants ayant consommé de façon prolongée des farines souillées par Aspergillus flavus. La Presse Médicale 1966, 74, 649–651. [Google Scholar]
- Dao, M. La malnutrition infantile en AOF. Le cas du Sénégal 1930–1960. Ann. Fac. Lett. Sci. Hum. 2004, 34, 01304682. [Google Scholar]
- FAO. Perspective on mycotoxins. Selected documents of the Joint FAO/WHO/UNEP conference on mycotoxins. FAO Food Nutr. Pap. 1979, 13, 1–167. [Google Scholar]
- Serck-Hanssen, A. Aflatoxin-induced fatal hepatitis? A case report from Uganda. Arch. Environ. Health 1970, 20, 729–731. [Google Scholar] [CrossRef]
- Alpert, E.; Serck-Hanssen, A.; Rajagopolan, B. Aflatoxin-induced hepatic injury in the African monkey. Arch. Environ. Health 1970, 20, 723–728. [Google Scholar] [CrossRef]
- IARC (International Agency for Research on Cancer). Some inorganic substances, chlorinated hydrocarbons, aromatic amines, N-Nitroso compounds, and natural products. In IARC Monogr. Eval. Carcinog. Risk Chem. Hum.; World Health Organization: Lyon, France, 1972; Volume 1, pp. 145–156. [Google Scholar]
- Stoloff, L.; Friedman, L. Information bearing on the evaluation of the hazard to man from aflatoxin ingestion. PAG Bull. 1976, 6, 21–32. [Google Scholar]
- Stoloff, L. Aflatoxin as a cause of primary liver-cell cancer in the United States: A probability study. Nutr. Cancer 1983, 5, 165–186. [Google Scholar] [CrossRef]
- Bruce, R.D. Risk assessment for aflatoxin: II. Implications of human epidemiology data. Risk Anal. 1990, 10, 561–569. [Google Scholar] [CrossRef]
- Zhang, K.; Wong, J.W.; Krynitsky, A.J.; Trucksess, M.W. Perspective on advancing FDA regulatory monitoring for mycotoxins in foods using liquid chromatography and mass spectrometry (Review). J. AOAC Int. 2016, 99, 890–894. [Google Scholar] [CrossRef]
- IARC (International Agency for Research on Cancer). Some naturally occurring substances. In IARC Monogr. Eval. Carcinog. Risks Hum.; World Health Organization: Lyon, France, 1976; Volume 10, pp. 51–72. [Google Scholar]
- IARC (International Agency for Research on Cancer). Overall evaluations of carcinogenicity: An updating of IARC monographs. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans Supplement 7; World Health Organization: Lyon, France, 1987; Volume 1–42, pp. 83–87. [Google Scholar]
- Ross, R.K.; Yu, M.C.; Henderson, B.E.; Yuan, J.M.; Qian, G.S.; Tu, J.T.; Gao, Y.T.; Wogan, G.N.; Groopman, J.D. Urinary aflatoxin biomarkers and risk of hepatocellular carcinoma. Lancet 1992, 339, 943–946. [Google Scholar] [CrossRef]
- Lasky, T.; Magder, L. Hepatocellular carcinoma p53 G > T transversions at codon 249: The fingerprint of aflatoxin exposure? Environ. Health Perspect. 1997, 105, 392–397. [Google Scholar]
- Qian, G.S.; Ross, R.K.; Yu, M.C.; Yuan, J.M.; Gao, Y.T.; Henderson, B.E.; Wogan, G.N.; Groopman, J.D. A follow-up study of urinary markers of aflatoxin exposure and liver cancer risk in Shanghai, People’s Republic of China. Cancer Epidemiol. Biomark. Prev. 1994, 3, 3–10. [Google Scholar]
- IARC (International Agency for Research on Cancer). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monogr. Eval. Carcinog. Risks Hum.; World Health Oorganization: Lyon, France, 1993; Volume V56, pp. 51–72. [Google Scholar]
- IARC (International Agency for Research on Cancer). Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. In IARC Monogr. Eval. Carcinog. Risks Hum.; World Health Organisation: Lyon, France, 2002; Volume 82, pp. 1–556. [Google Scholar]
- Probst, C.; Njapau, H.; Cotty, P.J. Outbreak of an acute aflatoxicosis in Kenya in 2004: Identification of the causal agent. Appl. Environ. Microbiol. 2007, 73, 2762–2764. [Google Scholar] [CrossRef]
- CDC (Centers for Disease Control and Prevention). Outbreak of aflatoxin poisoning—Eastern and central provinces, Kenya, January–July 2004. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 790–793. [Google Scholar]
- Azziz-Baumgartner, E.; Lindblade, K.; Gieseker, K.; Rogers, H.S.; Kieszak, S.; Njapau, H.; Schleicher, R.; McCoy, L.F.; Misore, A.; DeCock, K.; et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environ. Health Perspect. 2005, 113, 1779–1783. [Google Scholar] [CrossRef]
- Bressac, B.; Kew, M.; Wands, J.; Ozturk, M. Selective G to T mutations of p53 gene in hepatocellular carcinoma from southern Africa. Nature 1991, 350, 429–431. [Google Scholar] [CrossRef]
- Hussain, S.P.; Schwank, J.; Staib, F.; Wang, X.W.; Harris, C.C. TP53 mutations and hepatocellular carcinoma: Insights into the etiology and pathogenesis of liver cancer. Oncogene 2007, 26, 2166. [Google Scholar] [CrossRef]
- Staib, F.; Perwez Hussain, S.; Hofseth, L.J.; Wang, X.W.; Harris, C.C. TP53 and liver carcinogenesis. Hum. Mutat. 2003, 21, 201–216. [Google Scholar] [CrossRef]
- IARC (International Agency for Research on Cancer). Chemical agents and related occupations. Aflatoxins. In IARC Monogr. Eval. Carcinog. Risks Hum.; World Health Organisation: Lyon, France, 2012; Volume 100F, pp. 225–248. [Google Scholar]
- Kew, M.C. Synergistic interaction between aflatoxin B1 and hepatitis B virus in hepatocarcinogenesis. Liver Int. 2003, 23, 405–409. [Google Scholar] [CrossRef]
- Stern, M.C.; Umbach, D.M.; Yu, M.C.; London, S.J.; Zhang, Z.Q.; Taylor, J.A. Hepatitis B, aflatoxin B(1), and p53 codon 249 mutation in hepatocellular carcinomas from Guangxi, People’s Republic of China, and a meta-analysis of existing studies. Cancer Epidemiol. Biomark. Prev. 2001, 10, 617–625. [Google Scholar]
- Wang, S.S.; O’Neill, J.P.; Qian, G.S.; Zhu, Y.R.; Wang, J.B.; Armenian, H.; Zarba, A.; Wang, J.S.; Kensler, T.W.; Cariello, N.F.; et al. Elevated HPRT mutation frequencies in aflatoxin-exposed residents of daxin, Qidong county, People’s Republic of China. Carcinogenesis 1999, 20, 2181–2184. [Google Scholar] [CrossRef]
- Wang, H.; Liao, P.; Zeng, S.X.; Lu, H. It takes a team: A gain-of-function story of p53-R249S. J. Mol. Cell. Biol. 2019, 1, 277–283. [Google Scholar] [CrossRef]
- Vidal, A.; Mengelers, M.; Yang, S.; De Saeger, S.; De Boevre, M. Mycotoxin biomarkers of exposure: A comprehensive review. Comp. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef]
- Groopman, J.D.; Johnson, D.; Kensler, T.W. Aflatoxin and hepatitis B virus biomarkers: A paradigm for complex environmental exposures and cancer risk. Cancer Biomark. 2005, 1, 5–14. [Google Scholar] [CrossRef]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Watanabe, M.F.; Park, H.D.; Chen, G.C.; Chen, G.; Yu, S.Z. Detection of microcystins, a blue-green algal hepatotoxin, in drinking water sampled in Haimen and Fusui, endemic areas of primary liver cancer in China, by highly sensitive immunoassay. Carcinogenesis 1996, 17, 1317–1321. [Google Scholar] [CrossRef]
- Buechi, G.; Foulkes, D.M.; Kurono, M.; Mitchell, G.F.; Schneider, R.S. Total synthesis of racemic aflatoxin B1. J. Am. Chem. Soc. 1967, 89, 6745–6753. [Google Scholar] [CrossRef]
- FDA (U.S. Food and Drug Administration). CPG Sec 527.400. Whole Milk, Lowfat Milk, Skim Milk–Aflatoxin M1. Available online: http://www.fda.gov/ICECI/ComplianceManuals/CompliancePolicyGuidanceManual/ucm074482.htm (accessed on 3 July 2019).
- Wang, J.S.; Qian, G.S.; Zarba, A.; He, X.; Zhu, Y.R.; Zhang, B.C.; Jacobson, L.; Gange, S.J.; Munoz, A.; Kensler, T.W.; et al. Temporal patterns of aflatoxin-albumin adducts in hepatitis B surface antigen-positive and antigen-negative residents of Daxin, Qidong County, People’s Republic of China. Cancer Epidemiol. Biomark. Prev. 1996, 5, 253–261. [Google Scholar]
- Chakraborty, S.; Baral, S.; Paul, D.; Steinbach, P.J.; Rice, P.A.; Min, J.-H.; Ansari, A. Evidence for conformational capture mechanism for damage recognition by DNA repairprotein Rad4. Biophys. J. 2019, 116, 499a. [Google Scholar] [CrossRef]
- Ren, X.; Zhang, Q.; Wu, W.; Yan, T.; Tang, X.; Zhang, W.; Yu, L.; Li, P. Anti-idiotypic nanobody-phage display-mediated real-time immuno-PCR for sensitive, simultaneous and quantitative detection of total aflatoxins and zearalenone in grains. Food Chem. 2019, 297, 124912. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins (Basel) 2019, 11, 375. [Google Scholar] [CrossRef]
- Al-Jaal, B.A.; Jaganjac, M.; Barcaru, A.; Horvatovich, P.; Latiff, A. Aflatoxin, fumonisin, ochratoxin, zearalenone and deoxynivalenol biomarkers in human biological fluids: A systematic literature review, 2001–2018. Food Chem. Toxicol. 2019, 129, 211–228. [Google Scholar] [CrossRef]
- Maragos, C.M. Emerging technologies for mycotoxin detection. J. Toxicol. Toxin Rev. 2004, 23, 317–344. [Google Scholar] [CrossRef]
- Maragos, C.M. Multiplexed biosensors for mycotoxins. J. AOAC Int. 2016, 99, 849–860. [Google Scholar] [CrossRef]
- Krska, R.; Sulyok, M.; Berthiller, F.; Schuhmacher, R. Mycotoxin testing: From multi-toxin analysis to metabolomics. JSM Mycotoxins 2017, 67, 11–16. [Google Scholar] [CrossRef]
- Keener, L.; Nicholson-Keener, S.M.; Koutchma, T. Harmonization of legislation and regulations to achieve food safety: US and Canada perspective. J. Sci. Food Agric. 2014, 94, 1947–1953. [Google Scholar] [CrossRef]
- Winickoff, D.E.; Bushey, D.M. Science and power in global food regulation: The rise of the codex alimentarius. Sci. Technol. Hum. Values 2009, 35, 356–381. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize: An extensive survey. Toxins (Basel) 2016, 8, 363. [Google Scholar] [CrossRef]
- Berthiller, F.; Brera, C.; Iha, M.H.; Krska, R.; Lattanzio, V.M.T.; MacDonald, S.; Malone, R.J.; Maragos, C.; Solfrizzo, M.; Stranska-Zachariasova, M.; et al. Developments in mycotoxin analysis: An update for 2015–2016. World Mycotoxin J. 2017, 10, 5–29. [Google Scholar] [CrossRef]
- Panasiuk, L.; Jedziniak, P.; Pietruszka, K.; Piatkowska, M.; Bocian, L. Frequency and levels of regulated and emerging mycotoxins in silage in Poland. Mycot. Res. 2019, 35, 17–25. [Google Scholar] [CrossRef]
- Schwab, C.; Kovalsky, P. Why Advanced Multiple Mycotoxin Detection Matters. BIOMIN Holding GmbH. Getzersdorf, Austria. Available online: http://www. biomin.net/en/articles/why-advanced-multiple-mycotoxin -detection-matters/ (accessed on 29 June 2019).
- Giovati, L.; Magliani, W.; Ciociola, T.; Santinoli, C.; Conti, S.; Polonelli, L. AFM1 in milk: Physical, biological, and prophylactic methods to mitigate contamination. Toxins (Basel) 2015, 7, 4330–4349. [Google Scholar] [CrossRef]
- Nakazato, M.; Saito, K.; Kikuchi, Y.; Ibe, A.; Fujinuma, K.; Nishijima, M.; Nishima, T.; Morozumi, S.; Wauke, T.; Hitokoto, H. Aflatoxicol formation by Aspergillus flavus and A. parasiticus. Food Hyg. Saf. Sci. 1985, 26, 380–384. [Google Scholar] [CrossRef]
- Stubblefield, R.D.; Shotwell, O.L.; Shannon, G.M.; Weisleder, D.; Rohwedder, W.K. Parasiticol: A new metabolite from Aspergillus parasiticus. Agric. Food Chem. 1970, 18, 391–393. [Google Scholar] [CrossRef]
- Dunn, J.J.; Lee, L.S.; Ciegler, A. Mutagenicity and toxicity of aflatoxin precursors. Environ. Mutagen. 1982, 4, 19–26. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- Meyer, H.; Skhosana, D.Z.; Motlanthe, M.; Louw, W.; Rohwer, E. Long term monitoring (2014–2018) of multi-mycotoxins in South African commercial maize and wheat with a locally developed and validated LC-MS/MS method. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Michlmayr, H.; Malachova, A.; Koutnik, A.; Maloku, I.; Berthiller, F.; Adam, G.; Krska, R.; et al. Biotransformation of the mycotoxin deoxynivalenol in fusarium resistant and susceptible near isogenic wheat lines. PLoS ONE 2015, 10, e0119656. [Google Scholar] [CrossRef]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Berthiller, F.; Häubl, G.; Jaunecker, G.; Adam, G.; Krska, R.; Schuhmacher, R. Stable isotopic labelling-assisted untargeted metabolic profiling reveals novel conjugates of the mycotoxin deoxynivalenol in wheat. Anal. Bioanal. Chem. 2013, 405, 5031–5036. [Google Scholar] [CrossRef]
- Varga, E.; Glauner, T.; Berthiller, F.; Krska, R.; Schuhmacher, R.; Sulyok, M. Development and validation of a (semi-)quantitative UHPLC-MS/MS method for the determination of 191 mycotoxins and other fungal metabolites in almonds, hazelnuts, peanuts and pistachios. Anal. Bioanal. Chem. 2013, 405, 5087–5104. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Jani, D.; Allinson, J.; Berisha, F.; Cowan, K.J.; Devanarayan, V.; Gleason, C.; Jeromin, A.; Keller, S.; Khan, M.U.; Nowatzke, B.; et al. Recommendations for use and fit-for-purpose validation of biomarker multiplex ligand binding assays in drug development. AAPS J. 2016, 18, 1–14. [Google Scholar] [CrossRef]
- FAO (Food and Agricultural Organzation of the United Nations). Agreement on the Application of Sanitary and Phytosanitary Measures (SPS) and Agreement on Technical Barriers to Trade (TBT). Available online: http://www.fao.org/3/x7354e/X7354e02.htm (accessed on 17 July 2019).
- Groopman, J.D.; Kensler, T.W. The light at the end of the tunnel for chemical-specific biomarkers: Daylight or headlight? Carcinogenesis 1999, 20, 1–11. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Wild, C.P.; Hudson, G.J.; Sabbioni, G.; Chapot, B.; Hall, A.J.; Wogan, G.N.; Whittle, H.; Montesano, R.; Groopman, J.D. Dietary intake of aflatoxins and the level of albumin-bound aflatoxin in peripheral blood in The Gambia, West Africa. Cancer Epidemiol. Biomark. Prev. 1992, 1, 229. [Google Scholar]
- Gan, L.S.; Skipper, P.L.; Peng, X.C.; Groopman, J.D.; Chen, J.S.; Wogan, G.N.; Tannenbaum, S.R. Serum albumin adducts in the molecular epidemiology of aflatoxin carcinogenesis: Correlation with aflatoxin B1 intake and urinary excretion of aflatoxin M1. Carcinogenesis 1988, 9, 1323–1325. [Google Scholar] [CrossRef]
- Turner, P.C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica 2013. [Google Scholar] [CrossRef]
- Kensler, T.W.; Gange, S.J.; Egner, P.A.; Dolan, P.M.; Munoz, A.; Groopman, J.D.; Rogers, A.E.; Roebuck, B.D. Predictive value of molecular dosimetry: Individual versus group effects of oltipraz on aflatoxin-albumin adducts and risk of liver cancer. Cancer Epidemiol. Biomark. Prev. 1997, 6, 603–610. [Google Scholar]
- Rushing, B.R.; Selim, M.I. Structure and oxidation of pyrrole adducts formed between aflatoxin B2a and biological amines. Chem. Res. Toxicol. 2017, 30, 1275–1285. [Google Scholar] [CrossRef]
- Groopman, J.D.; Hall, A.J.; Whittle, H.; Hudson, G.J.; Wogan, G.N.; Montesano, R.; Wild, C.P. Molecular dosimetry of aflatoxin-N7-guanine in human urine obtained in The Gambia, West Africa. Cancer Epidemiol. Biomark. Prev. 1992, 1, 221. [Google Scholar]
- Bennett, R.A.; Essigmann, J.M.; Wogan, G.N. Excretion of an aflatoxin-guanine adduct in the urine of aflatoxin B1-treated rats. Cancer Res. 1981, 41, 650–654. [Google Scholar]
- Chou, M.W.; Chen, W. Food restriction reduces aflatoxin B1 (AFB1)-DNA adduct formation, AFB1-Glutathione conjugation, and DNA damage in AFB1-treated male F344 rats and B6C3F1 mice. J. Nutr. 1997, 127, 210–217. [Google Scholar] [CrossRef]
- Wild, C.P.; Turner, P.C. The toxicology of aflatoxins as a basis for public health decisions. Mutagenesis 2002, 17, 471–481. [Google Scholar] [CrossRef]
- Carvajal, M. Aflatoxin-DNA adducts as biomarkers of cancer: Nature, formation, kinds of AF-DNA adducts, methodology, effects, and control. In Food Contaminants; American Chemical Society: Washington, DC, USA, 2008; Volume 1001, pp. 13–55. [Google Scholar]
- Groopman, J.D.; Donahue, P.R.; Zhu, J.Q.; Chen, J.S.; Wogan, G.N. Aflatoxin metabolism in humans: Detection of metabolites and nucleic acid adducts in urine by affinity chromatography. Proc. Natl. Acad. Sci. USA 1985, 82, 6492. [Google Scholar] [CrossRef]
- Yu, M.W.; Lien, J.P.; Chiu, Y.H.; Santella, R.M.; Liaw, Y.F.; Chen, C.J. Effect of aflatoxin metabolism and DNA adduct formation on hepatocellular carcinoma among chronic hepatitis B carriers in Taiwan. J. Hepatol. 1997, 27, 320–330. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Zhang, L.S.; Hu, X.; Xiao, Y.; Chen, J.S.; Xu, Y.C.; Fremy, J.; Chu, F.S. Correlation of dietary aflatoxin B1 levels with excretion of aflatoxin M1 in human urine. Cancer Res. 1987, 47, 1848–1852. [Google Scholar]
- Kensler, T.W.; He, X.; Otieno, M.; Egner, P.A.; Jacobson, L.P.; Chen, B.; Wang, J.S.; Zhu, Y.R.; Zhang, B.C.; Wang, J.B.; et al. Oltipraz chemoprevention trial in Qidong, People’s Republic of China: Modulation of serum aflatoxin albumin adduct biomarkers. Cancer Epidemiol. Biomark. Prev. 1998, 7, 127–134. [Google Scholar]
- Johnson, D.N.; Egner, P.A.; Obrian, G.; Glassbrook, N.; Roebuck, B.D.; Sutter, T.R.; Payne, G.A.; Kensler, T.W.; Groopman, J.D. Quantification of urinary aflatoxin B1 dialdehyde metabolites formed by aflatoxin aldehyde reductase using isotope dilution tandem mass spectrometry. Chem. Res. Toxicol. 2008, 21, 752–760. [Google Scholar] [CrossRef]
- Kelly, V.P.; Ellis, E.M.; Manson, M.M.; Chanas, S.A.; Moffat, G.J.; McLeod, R.; Judah, D.J.; Neal, G.E.; Hayes, J.D. Chemoprevention of aflatoxin; hepatocarcinogenesis by coumarin, a natural benzopyrone that is a potent inducer of aflatoxin-aldehyde reductase, the glutathione-transferase A5 and P1 subunits, and NAD(P)H:quinone oxidoreductase in rat liver. Cancer Res. 2000, 60, 957. [Google Scholar]
- Kwak, M.K.; Egner, P.A.; Dolan, P.M.; Ramos-Gomez, M.; Groopman, J.D.; Itoh, K.; Yamamoto, M.; Kensler, T.W. Role of phase 2 enzyme induction in chemoprotection by dithiolethiones. Mutat. Res. 2001, 480–481, 305–315. [Google Scholar] [CrossRef]
- Díaz de León-Martínez, L.; Díaz-Barriga, F.; Barbier, O.; Ortíz, D.L.G.; Ortega-Romero, M.; Pérez-Vázquez, F.; Flores-Ramírez, R. Evaluation of emerging biomarkers of renal damage and exposure to aflatoxin-B1 in Mexican indigenous women: A pilot study. Environ. Sci. Pollut. Res. 2019, 26, 12205–12216. [Google Scholar] [CrossRef]
- Li, S.; Muhammad, I.; Yu, H.; Sun, X.; Zhang, X. Detection of Aflatoxin adducts as potential markers and the role of curcumin in alleviating AFB1-induced liver damage in chickens. Ecotoxicol. Environ. Saf. 2019, 176, 137–145. [Google Scholar] [CrossRef]
- McMillan, A.; Renaud, J.B.; Burgess, K.M.N.; Orimadegun, A.E.; Akinyinka, O.O.; Allen, S.J.; Miller, J.D.; Reid, G.; Sumarah, M.W. Aflatoxin exposure in Nigerian children with severe acute malnutrition. Food Chem. Toxicol. 2018, 111, 356–362. [Google Scholar] [CrossRef]
- Egner, P.A.; Groopman, J.D.; Wang, J.S.; Kensler, T.W.; Friesen, M.D. Quantification of aflatoxin-B1-N7-Guanine in human urine by high-performance liquid chromatography and isotope dilution tandem mass spectrometry. Chem. Res. Toxicol. 2006, 19, 1191–1195. [Google Scholar] [CrossRef]
- Groopman, J.D.; Wild, C.P.; Hasler, J.; Junshi, C.; Wogan, G.N.; Kensler, T.W. Molecular epidemiology of aflatoxin exposures: Validation of aflatoxin-N7-guanine levels in urine as a biomarker in experimental rat models and humans. Environ. Health Perspect. 1993, 99, 107–113. [Google Scholar] [CrossRef]
- Jangampalli Adi, P.; Matcha, B. Analysis of aflatoxin B1 in contaminated feed, media, and serum samples of Cyprinus carpio L. by high-performance liquid chromatography. Food Qual. Saf. 2018, 2, 199–204. [Google Scholar] [CrossRef]
- Hendrickse, R.G.; Maxwell, S.M.; Young, R. Aflatoxins and heroin. BMJ (Clin. Res. Ed.) 1989, 299, 492–493. [Google Scholar] [CrossRef]
- Yourtee, D.M.; Bean, T.A.; Kirk-Yourtee, C.L. Human aflatoxin B1 metabolism: An investigation of the importance of aflatoxin Q1 as a metabolite of hepatic post-mitochondrial fraction. Toxicol. Lett. 1987, 38, 213–224. [Google Scholar] [CrossRef]
- Raney, K.D.; Shimada, T.; Kim, D.H.; Groopman, J.D.; Harris, T.M.; Guengerich, F.P. Oxidation of aflatoxins and sterigmatocystin by human liver microsomes: Significance of aflatoxin Q1 as a detoxication product of aflatoxin B1. Chem. Res. Toxicol. 1992, 5, 202–210. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Effect of dietary acids on the formation of aflatoxin B2a as a means to detoxify aflatoxin B1. Food Addit. Contam.: Part A 2016, 33, 1456–1467. [Google Scholar] [CrossRef]
- Johnson, W.W.; Guengerich, F.P. Reaction of aflatoxin B1 exo-8,9-epoxide with DNA: Kinetic analysis of covalent binding and DNA-induced hydrolysis. Proc. Natl. Acad. Sci. USA 1997, 94, 6121–6125. [Google Scholar] [CrossRef]
- Brown, K.L.; Bren, U.; Stone, M.P.; Guengerich, F.P. Inherent stereospecificity in the reaction of aflatoxin B(1) 8,9-epoxide with deoxyguanosine and efficiency of DNA catalysis. Chem. Res. Toxicol. 2009, 22, 913–917. [Google Scholar] [CrossRef]
- McCoy, L.F.; Scholl, P.F.; Sutcliffe, A.E.; Kieszak, S.M.; Powers, C.D.; Rogers, H.S.; Gong, Y.Y.; Groopman, J.D.; Wild, C.P.; Schleicher, R.L. Human aflatoxin albumin adducts quantitatively compared by ELISA, HPLC with fluorescence detection, and HPLC with isotope dilution mass spectrometry. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1653–1657. [Google Scholar] [CrossRef]
- Tang, L.; Xu, L.; Afriyie-Gyawu, E.; Liu, W.; Wang, P.; Tang, Y.; Wang, Z.; Huebner, H.J.; Ankrah, N.A.; Ofori-Adjei, D.; et al. Aflatoxin–albumin adducts and correlation with decreased serum levels of vitamins A and E in an adult Ghanaian population. Food Addit. Contam.: Part A 2009, 26, 108–118. [Google Scholar] [CrossRef]
- Kensler, T.W. Chemoprevention by inducers of carcinogen detoxication enzymes. Environ. Health Perspect. 1997, 105 (Suppl. 4), 965–970. [Google Scholar]
- Wang, J.S.; Shen, X.; He, X.; Zhu, Y.R.; Zhang, B.C.; Wang, J.B.; Qian, G.S.; Kuang, S.Y.; Zarba, A.; Egner, P.A.; et al. Protective alterations in phase 1 and 2 metabolism of aflatoxin B1 by oltipraz in residents of Qidong, People’s Republic of China. J. Natl. Cancer Inst. 1999, 91, 347–354. [Google Scholar] [CrossRef]
- Miraglia, N.; Assennato, G.; Clonfero, E.; Fustinoni, S.; Sannolo, N. Biologically effective dose biomarkers. G Ital. Med. Lav. Ergon. 2004, 26, 298–301. [Google Scholar]
- Mykkänen, H.; Zhu, H.; Salminen, E.; Juvonen, R.O.; Ling, W.; Ma, J.; Polychronaki, N.; Kemiläinen, H.; Mykkänen, O.; Salminen, S.; et al. Fecal and urinary excretion of aflatoxin B1 metabolites (AFQ1, AFM1 and AFB-N7-guanine) in young Chinese males. Int. J. Cancer 2005, 115, 879–884. [Google Scholar] [CrossRef]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef]
- Strosnider, H.; Azziz-Baumgartner, E.; Banziger, M.; Bhat Ramesh, V.; Breiman, R.; Brune, M.-N.; DeCock, K.; Dilley, A.; Groopman, J.; Hell, K.; et al. Workgroup report: Public health strategies for reducing aflatoxin exposure in developing countries. Environ. Health Perspect. 2006, 114, 1898–1903. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, J.; Wang, G.; Groopman, J.D.; Kensler, T.W. Qidong: A crucible for studies on liver cancer etiology and prevention. Cancer Biol. Med. 2019, 16, 24–37. [Google Scholar]
- Shephard, G.S.; Burger, H.M.; Gambacorta, L.; Gong, Y.Y.; Krska, R.; Rheeder, J.P.; Solfrizzo, M.; Srey, C.; Sulyok, M.; Visconti, A.; et al. Multiple mycotoxin exposure determined by urinary biomarkers in rural subsistence farmers in the former Transkei, South Africa. Food Chem. Toxicol. 2013, 62, 217–225. [Google Scholar] [CrossRef]
- Cotty, P.J.; Jaime-Garcia, R. Influences of climate on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol. 2007, 119, 109–115. [Google Scholar] [CrossRef]
- Baluka, S.A.; Schrunk, D.; Imerman, P.; Kateregga, J.N.; Camana, E.; Wang, C.; Rumbeiha, W.K. Mycotoxin and metallic element concentrations in peanut products sold in Ugandan markets. Cogent Food Agric. 2017, 3, 1313925. [Google Scholar] [CrossRef]
- Mutegi, C.; Wagacha, M.; Kimani, J.; Otieno, G.; Wanyama, R.; Hell, K.; Christie, M.E. Incidence of aflatoxin in peanuts (Arachis hypogaea Linnaeus) from markets in Western, Nyanza and Nairobi Provinces of Kenya and related market traits. J. Stored Prod. Res. 2013, 52, 118–127. [Google Scholar] [CrossRef]
- Mutegi, C.K.; Cotty, P.J.; Bandyopadhyay, R. Prevalence and mitigation of aflatoxins in Kenya (1960-to date). World Mycotoxin J. 2018, 11, 341–357. [Google Scholar] [CrossRef]
- Villers, P. Food safety and aflatoxin control. J. Food Res. 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef]
- Krishnamachari, K.A.V.R.; Nagarajan, V.; Bhat, R.; Tilak, T.B.G. Hepatitis due to aflatoxicosis: An outbreak in Western India. Lancet 1975, 305, 1061–1063. [Google Scholar] [CrossRef]
- Stepman, F. Scaling-up the impact of aflatoxin research in Africa. The role of social sciences. Toxins (Basel) 2018, 10, 136. [Google Scholar] [CrossRef]
- FAO/WHO. Risk Communication Applied to Food Safety Handbook. Available online: http://www.fao.org/3/a-i5863e.pdf (accessed on 18 September 2019).
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef]
- Chauhan, Y.S.; Wright, G.C.; Rachaputi, R.C.N.; Holzworth, D.; Broome, A.; Krosch, S.; Robertson, M.J. Application of a model to assess aflatoxin risk in peanuts. J. Agric. Sci. 2010, 148, 341–351. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S.; Hammer, G.L.; Probert, M.E.; Robertson, M.J.; Holzworth, D.; Huth, N.I.; Hargreaves, J.N.G.; Meinke, H.; Hochman, Z.; et al. An overview of APSIM, a model designed for farming systems simulation. Eur. J. Agron. 2003, 18, 267–288. [Google Scholar] [CrossRef]
- Chauhan, Y.; Tatnell, J.; Krosch, S.; Karanja, J.; Gnonlonfin, B.; Wanjuki, I.; Wainaina, J.; Harvey, J. An improved simulation model to predict pre-harvest aflatoxin risk in maize. Field Crop. Res. 2015, 178, 91–99. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hammer, G.L.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Chang. 2013, 3, 497. [Google Scholar] [CrossRef]
- Pitt, R.E. A Descriptive Model of Mold Growth and aflatoxin formation as affected by environmental conditions. J. Food Prot. 1993, 56, 139–146. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Predicting mycotoxins in foods: A review. Food Microbiol. 2009, 26, 757–769. [Google Scholar] [CrossRef]
- Gqaleni, N.; Smith, J.E.; Lacey, J.; Gettinby, G. Effects of Temperature, Water Activity, and Incubation Time on Production of Aflatoxins and Cyclopiazonic Acid by an Isolate of Aspergillus flavus in Surface Agar Culture. Appl. Environ. Microbiol. 1997, 63, 1048. [Google Scholar]
- Molina, M.; Giannuzzi, L. Modelling of aflatoxin production by Aspergillus parasiticus in a solid medium at different temperatures, pH and propionic acid concentrations. Food Res. Int. 2002, 35, 585–594. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef]
- Mousa, W.; Ghazali, F.M.; Jinap, S.; Ghazali, H.M.; Radu, S. Modelling the effect of water activity and temperature on growth rate and aflatoxin production by two isolates of Aspergillus flavus on paddy. J. Appl. Microbiol. 2011, 111, 1262–1274. [Google Scholar] [CrossRef]
- Astoreca, A.; Vaamonde, G.; Dalcero, A.; Ramos, A.J.; Marín, S. Modelling the effect of temperature and water activity of Aspergillus flavus isolates from corn. Int. J. Food Microbiol. 2012, 156, 60–67. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; de Voil, P.G.; Zurcher, E.J.; Herrmann, N.I.; McLean, G.; Chenu, K.; van Oosterom, E.J.; Snow, V.; Murphy, C.; et al. APSIM – Evolution towards a new generation of agricultural systems simulation. Environ. Model. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Chauhan, Y.S.; Wright, G.C.; Rachaputi, N.C. Modelling climatic risks of aflatoxin contamination in maize. Aust. J. Exp. Agric. 2008, 48, 358–366. [Google Scholar] [CrossRef]
- Anyamba, A.; Chretien, J.-P.; Britch, S.C.; Soebiyanto, R.P.; Small, J.L.; Jepsen, R.; Forshey, B.M.; Sanchez, J.L.; Smith, R.D.; Harris, R.; et al. Global disease outbreaks associated with the 2015–2016 El Niño event. Sci. Rep. 2019, 9, 1930. [Google Scholar] [CrossRef]
- Chao, T.C.; Maxwell, S.M.; Wong, S.Y. An outbreak of aflatoxicosis and boric acid poisoning in Malaysia: A clinicopathological study. J. Pathol. 1991, 164, 225–233. [Google Scholar] [CrossRef]
- Kamala, A.; Shirima, C.; Jani, B.; Bakari, M.; Sillo, H.; Rusibamayila, N.; De Saeger, S.; Kimanya, M.; Gong, Y.Y.; Simba, A. Outbreak of an acute aflatoxicosis in Tanzania during 2016. World Mycotoxin J. 2018, 11, 311–320. [Google Scholar] [CrossRef]
- Reddy, B.N.; Raghvender, C.N. Outbreaks of aflatoxicosis in India. Afr. J. Food Agric. Nutr. Dev. 2007, 7, 1–15. [Google Scholar]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef]
- Khlangwiset, P.; Wu, F. Costs and efficacy of public health interventions to reduce aflatoxin-induced human disease. Food Addit. Contam.: Part A 2010, 27, 998–1014. [Google Scholar] [CrossRef]
- Pandey, K.M.; Kumar, R.; Pandey, K.A.; Soni, P.; Gangurde, S.S.; Sudini, K.H.; Fountain, C.J.; Liao, B.; Desmae, H.; Okori, P.; et al. Mitigating aflatoxin contamination in groundnut through a combination of genetic resistance and post-harvest management practices. Toxins (Basel) 2019, 11, 315. [Google Scholar] [CrossRef]
- Grace, D.; Mahuku, G.; Hoffmann, V.; Atherstone, C.; Upadhyaya, H.D.; Bandyopadhyay, R. International agricultural research to reduce food risks: Case studies on aflatoxins. Food Secur. 2015, 7, 569–582. [Google Scholar] [CrossRef]
- Benkerroum, N. Mycotoxins in dairy products: A review. Int. Dairy J. 2016, 62, 63–75. [Google Scholar] [CrossRef]
- Abrar, M.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Randhawa, M.A.; Saeed, F.; Waqas, K. Aflatoxins: Biosynthesis, occurrence, toxicity, and remedies. Crit. Rev. Food Sci. Nutr. 2013, 53, 862–874. [Google Scholar] [CrossRef]
- Jalili, M. A review on aflatoxins reduction in food. Iran. J. Health Saf. Environ. 2016, 3, 445–459. [Google Scholar]
- Doyle, M.P.; Applebaum, R.S.; Brackett, R.E.; Marth, E.H. Physical, chemical and biological degradation of mycotoxins in foods and agricultural commodities. J. Food Prot. 1982, 45, 964–971. [Google Scholar] [CrossRef]
- Verheecke, C.; Liboz, T.; Mathieu, F. Microbial degradation of aflatoxin B1: Current status and future advances. Int. J. Food Microbiol. 2016, 237, 1–9. [Google Scholar] [CrossRef]
- Hayes, J.D.; McLeod, R.; Ellis, E.M.; Pulford, D.J.; Ireland, L.S.; McLellan, L.I.; Judah, D.J.; Manson, M.M.; Neal, G.E. Regulation of glutathione S-transferases and aldehyde reductase by chemoprotectors: Studies of mechanisms responsible for inducible resistance to aflatoxin B1. IARC Sci. Publ. 1996, 175–187. [Google Scholar]
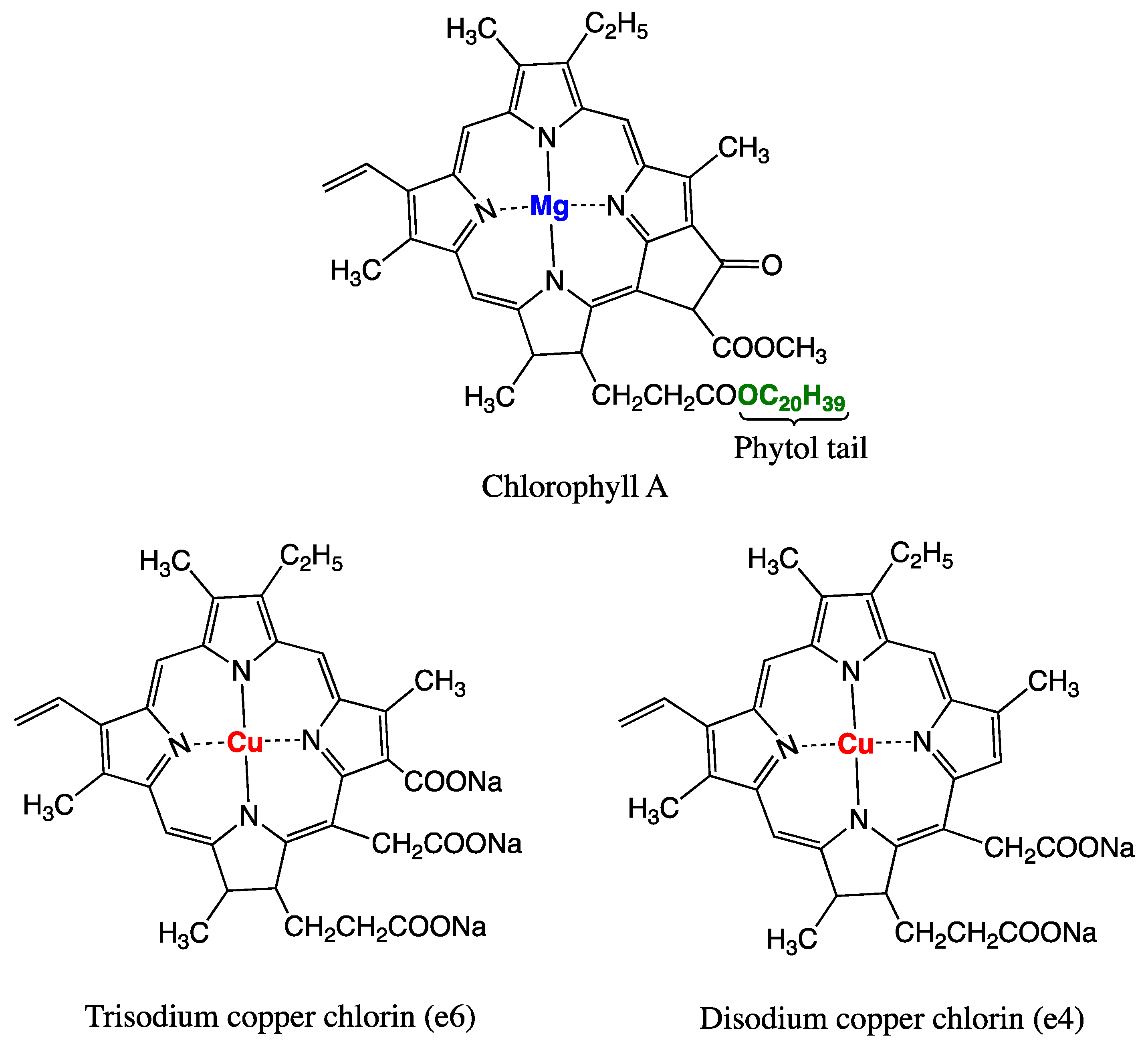
- Fahey, J.W.; Stephenson, K.K.; Dinkova-Kostova, A.T.; Egner, P.A.; Kensler, T.W.; Talalay, P. Chlorophyll, chlorophyllin and related tetrapyrroles are significant inducers of mammalian phase 2 cytoprotective genes. Carcinogenesis 2005, 26, 1247–1255. [Google Scholar] [CrossRef]
- Li, Y.; Su, J.-J.; Qin, L.-L.; Egner, P.A.; Wang, J.-S.; Groopman, J.D.; Kensler, T.W.; Roebuck, B.D. Reduction of aflatoxin B1 adduct biomarkers by oltipraz in the tree shrew (Tupaia belangeri chinensis). Cancer Lett. 2000, 154, 79–83. [Google Scholar]
- Roebuck, B.; Liu, Y.-L.; Rogers, A.E.; Groopman, J.D.; Kensler, T.W. Protection against aflatoxin B1-induced hepatocarcinogenesis in F344 rats by 5-(2-pyrazinyl)-4-methyl-1, 2-dithiole-3-thione (oltipraz): Predictive role for short-term molecular dosimetry. Cancer Res. 1991, 51, 5501–5506. [Google Scholar]
- Bammler, T.K.; Slone, D.H.; Eaton, D.L. Effects of dietary oltipraz and ethoxyquin on aflatoxin B1 biotransformation in non-human primates. Toxicol. Sci. 2000, 54, 30–41. [Google Scholar] [CrossRef]
- Bolton, M.G.; Muñoz, A.; Jacobson, L.P.; Groopman, J.D.; Maxuitenko, Y.Y.; Roebuck, B.; Kensler, T.W. Transient intervention with oltipraz protects against aflatoxin-induced hepatic tumorigenesis. Cancer Res. 1993, 53, 3499–3504. [Google Scholar]
- Primiano, T.; Egner, P.A.; Sutter, T.R.; Kelloff, G.J.; Roebuck, B.; Kensler, T.W. Intermittent dosing with oltipraz: Relationship between chemoprevention of aflatoxin-induced tumorigenesis and induction of glutathione S-transferases. Cancer Res. 1995, 55, 4319–4324. [Google Scholar]
- Langouet, S.; Coles, B.; Morel, F.; Becquemont, L.; Beaune, P.; Guengerich, F.P.; Ketterer, B.; Guillouzo, A. Inhibition of CYP1A2 and CYP3A4 by oltipraz results in reduction of aflatoxin B1 metabolism in human hepatocytes in primary culture. Cancer Res. 1995, 55, 5574–5579. [Google Scholar]
- Yates, M.S.; Kwak, M.-K.; Egner, P.A.; Groopman, J.D.; Bodreddigari, S.; Sutter, T.R.; Baumgartner, K.J.; Roebuck, B.D.; Liby, K.T.; Yore, M.M.; et al. Potent Protection against Aflatoxin-Induced Tumorigenesis through Induction of Nrf2-Regulated Pathways by the Triterpenoid 1-[2-Cyano-3-,12-Dioxooleana-1,9(11)-Dien-28-Oyl]Imidazole. Cancer Res. 2006, 66, 2488. [Google Scholar] [CrossRef]
- Johnson, N.M.; Egner, P.A.; Baxter, V.K.; Sporn, M.B.; Wible, R.S.; Sutter, T.R.; Groopman, J.D.; Kensler, T.W.; Roebuck, B.D. Complete protection against aflatoxin B(1)-induced liver cancer with a triterpenoid: DNA adduct dosimetry, molecular signature, and genotoxicity threshold. Cancer Prev. Res. (Phila.) 2014, 7, 658–665. [Google Scholar] [CrossRef]
- Smela, M.E.; Currier, S.S.; Bailey, E.A.; Essigmann, J.M. The chemistry and biology of aflatoxin B(1): From mutational spectrometry to carcinogenesis. Carcinogenesis 2001, 22, 535–545. [Google Scholar] [CrossRef]
- Williams, D.E. The rainbow trout liver cancer model: Response to environmental chemicals and studies on promotion and chemoprevention. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 121–127. [Google Scholar] [CrossRef]
- Bechtel, D.H. Molecular dosimetry of hepatic aflatoxin B1-DNA adducts: Linear correlation with hepatic cancer risk. Regul. Toxicol. Pharmacol. 1989, 10, 74–81. [Google Scholar] [CrossRef]
- Eaton, D.L.; Schaupp, C.M. Of mice, rats, and men: Could Nrf2 activation protect against aflatoxin heptocarcinogenesis in humans? Cancer Prev. Res. (Phila.) 2014, 7, 653–657. [Google Scholar] [CrossRef]
- Olden, K.; Vulimiri, S.V. Laboratory to community: Chemoprevention is the answer. Cancer Prev. Res. 2014, 7, 648. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef]
- Gregory, J.F., III; Edds, G.T. Effect of dietary selenium on the metabolism of aflatoxin B1 in turkeys. Food Chem. Toxicol. 1984, 22, 637–642. [Google Scholar] [CrossRef]
- Shi, C.Y.; Chua, S.C.; Lee, H.P.; Ong, C.N. Inhibition of aflatoxin B1-DNA binding and adduct formation by selenium in rats. Cancer Lett. 1994, 82, 203–208. [Google Scholar] [CrossRef]
- Chen, K.; Fang, J.; Peng, X.; Cui, H.; Chen, J.; Wang, F.; Chen, Z.; Zuo, Z.; Deng, J.; Lai, W.; et al. Effect of selenium supplementation on aflatoxin B1-induced histopathological lesions and apoptosis in bursa of Fabricius in broilers. Food Chem. Toxicol. 2014, 74, 91–97. [Google Scholar] [CrossRef]
- Shi, D.; Guo, S.; Liao, S.; Su, R.; Pan, J.; Lin, Y.; Tang, Z. Influence of selenium on hepatic mitochondrial antioxidant capacity in ducklings intoxicated with aflatoxin B(1). Biol. Trace Elem. Res. 2012, 145, 325–329. [Google Scholar] [CrossRef]
- Shi, D.; Liao, S.; Guo, S.; Li, H.; Yang, M.; Tang, Z. Protective effects of selenium on aflatoxin B1-induced mitochondrial permeability transition, DNA damage, and histological alterations in duckling liver. Biol. Trace Elem. Res. 2015, 163, 162–168. [Google Scholar] [CrossRef]
- Wang, F.; Shu, G.; Peng, X.; Fang, J.; Chen, K.; Cui, H.; Chen, Z.; Zuo, Z.; Deng, J.; Geng, Y.; et al. Protective effects of sodium selenite against aflatoxin B1-induced oxidative stress and apoptosis in broiler spleen. Int. J. Environ. Res. Public Health 2013, 10, 2834–2844. [Google Scholar] [CrossRef]
- Diaz, G.J.; Murcia, H.W.; Cepeda, S.M. Cytochrome P450 enzymes involved in the metabolism of aflatoxin B1 in chickens and quail. Poult. Sci. 2010, 89, 2461–2469. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, N.; Zhu, M.K.; Zhao, L.; Zhou, J.-C.; Qi, D.S. Prevention of aflatoxin B1 hepatoxicity by dietary selenium is associated with inhibition of cytochrome P450 isozymes and up-regulation of 6 selenoprotein genes in chick liver. J. Nutr. 2016, 146, 655–661. [Google Scholar] [CrossRef]
- Weng, M.W.; Lee, H.W.; Choi, B.; Wang, H.T.; Hu, Y.; Mehta, M.; Desai, D.; Amin, S.; Zheng, Y.; Tang, M.S. AFB1 hepatocarcinogenesis is via lipid peroxidation that inhibits DNA repair, sensitizes mutation susceptibility and induces aldehyde-DNA adducts at p53 mutational hotspot codon 249. Oncotarget 2017, 8, 18213–18226. [Google Scholar]
- MacFarquhar, J.K.; Broussard, D.L.; Melstrom, P.; Hutchinson, R.; Wolkin, A.; Martin, C.; Burk, R.F.; Dunn, J.R.; Green, A.L.; Hammond, R.; et al. Acute selenium toxicity associated with a dietary supplement. Arch. Intern. Med. 2010, 170, 256–261. [Google Scholar] [CrossRef]
- Hoffmann, V.; Moser, C.; Saak, A. Food safety in low and middle-income countries: The evidence through an economic lens. World Dev. 2019, 123, 104611. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Cheng, S.J.; Zhou, Z.C.; Athar, M.; Khan, W.A.; Bickers, D.R.; Mukhtar, H. Antimutagenic activity of green tea polyphenols. Mutat. Res.-Genet. Toxicol. 1989, 223, 273–285. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Lim, S.-B. Antioxidant and anticancer activities of broccoli by-products from different cultivars and maturity stages at harvest. Prev. Nutr. Food Sci. 2015, 20, 8–14. [Google Scholar] [CrossRef]
- Alumkal, J.J.; Slottke, R.; Schwartzman, J.; Cherala, G.; Munar, M.; Graff, J.N.; Beer, T.M.; Ryan, C.W.; Koop, D.R.; Gibbs, A.; et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Investig. New Drugs 2015, 33, 480–489. [Google Scholar] [CrossRef]
- Techapiesancharoenkij, N.; Fiala, J.L.A.; Navasumrit, P.; Croy, R.G.; Wogan, G.N.; Groopman, J.D.; Ruchirawat, M.; Essigmann, J.M. Sulforaphane, a cancer chemopreventive agent, induces pathways associated with membrane biosynthesis in response to tissue damage by aflatoxin B1. Toxicol. Appl. Pharmacol. 2015, 282, 52–60. [Google Scholar] [CrossRef]
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403. [Google Scholar] [CrossRef]
- Qin, G.; Gopalan-Kriczky, P.; Su, J.; Ning, Y.; Lotlikar, P.D. Inhibition of aflatoxin B1-induced initiation of hepatocarcinogenesis in the rat by green tea. Cancer Lett. 1997, 112, 149–154. [Google Scholar] [CrossRef]
- Nakachi, K.; Matsuyama, S.; Miyake, S.; Suganuma, M.; Imai, K. Preventive effects of drinking green tea on cancer and cardiovascular disease: Epidemiological evidence for multiple targeting prevention. Biofactors 2000, 13, 49–54. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M.; Imai, K.; Nakachi, K. Green tea: Cancer preventive beverage and/or drug. Cancer Lett. 2002, 188, 9–13. [Google Scholar] [CrossRef]
- Tang, L.; Tang, M.; Xu, L.; Luo, H.; Huang, T.; Yu, J.; Zhang, L.; Gao, W.; Cox, S.B.; Wang, J.-S. Modulation of aflatoxin biomarkers in human blood and urine by green tea polyphenols intervention. Carcinogenesis 2008, 29, 411–417. [Google Scholar] [CrossRef]
- Lu, W.; Kelly, A.L.; Miao, S. Emulsion-based encapsulation and delivery systems for polyphenols. Trends Food Sci. Technol. 2016, 47, 1–9. [Google Scholar] [CrossRef]
- Tantoush, Z.; Apostolovic, D.; Kravic, B.; Prodic, I.; Mihajlovic, L.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Green tea catechins of food supplements facilitate pepsin digestion of major food allergens, but hampers their digestion if oxidized by phenol oxidase. J. Funct. Foods 2012, 4, 650–660. [Google Scholar] [CrossRef]
- Ganai, S.A. Histone deacetylase inhibitor sulforaphane: The phytochemical with vibrant activity against prostate cancer. Biomed. Pharmacother. 2016, 81, 250–257. [Google Scholar] [CrossRef]
- Fahey, J.W.; Holtzclaw, W.D.; Wehage, S.L.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Sulforaphane bioavailability from glucoraphanin-rich broccoli: Control by active endogenous myrosinase. PLoS ONE 2015, 10, e0140963. [Google Scholar] [CrossRef]
- Fahey, J.W.; Wade, K.L.; Wehage, S.L.; Holtzclaw, W.D.; Liu, H.; Talalay, P.; Fuchs, E.; Stephenson, K.K. Stabilized sulforaphane for clinical use: Phytochemical delivery efficiency. Mol. Nutr. Food Res. 2017, 61, 1600766. [Google Scholar] [CrossRef]
- Fahey, J.W.; Wehage, S.L.; Holtzclaw, W.D.; Kensler, T.W.; Egner, P.A.; Shapiro, T.A.; Talalay, P. Protection of humans by plant glucosinolates: Efficiency of conversion of glucosinolates to isothiocyanates by the gastrointestinal microflora. Cancer Prev. Res. 2012, 5, 603. [Google Scholar] [CrossRef]
- Amjad, A.I.; Parikh, R.A.; Appleman, L.J.; Hahm, E.-R.; Singh, K.; Singh, S.V. Broccoli-derived sulforaphane and chemoprevention of prostate cancer: From bench to bedside. Curr. Pharmacol. Rep. 2015, 1, 382–390. [Google Scholar] [CrossRef]
- Hsu, C.-Y.; Chen, Y.-H.; Chao, P.-Y.; Chen, C.-M.; Hsieh, L.-L.; Hu, S.-P. Naturally occurring chlorophyll derivatives inhibit aflatoxin B1-DNA adduct formation in hepatoma cells. Mutat. Res.-Genet. Toxicol. Environ. 2008, 657, 98–104. [Google Scholar] [CrossRef]
- Wang, Y.-T.; Yang, C.-H.; Huang, T.-Y.; Tai, M.; Hsueh; Sie, R.-H.; Shaw, J.-F. Cytotoxic effects of chlorophyllides in ethanol crude extracts from plant leaves. Evid. Based Complement. Altern. Med. 2019, 2019, 13. [Google Scholar] [CrossRef]
- Hibasami, H.; Kyohkon, M.; Ohwaki, S.; Katsuzaki, H.; Imai, K.; Nakagawa, M.; Ishi, Y.; Komiya, T. Pheophorbide a, a moiety of chlorophyll a, induces apoptosis in human lymphoid leukemia molt 4B cells. Int. J. Mol. Med. 2000, 6, 277–279. [Google Scholar] [CrossRef]
- Breinholt, V.; Hendricks, J.; Pereira, C.; Arbogast, D.; Bailey, G. Dietary chlorophyllin is a potent inhibitor of aflatoxin B1 hepatocarcinogenesis in rainbow trout. Cancer Res. 1995, 55, 57–62. [Google Scholar]
- Lai, C.-N.; Butler, M.A.; Matney, T.S. Antimutagenic activities of common vegetables and their chlorophyll content. Mutat. Res.-Genet. Toxicol. 1980, 77, 245–250. [Google Scholar] [CrossRef]
- Simonich, M.T.; Egner, P.A.; Roebuck, B.D.; Orner, G.A.; Jubert, C.; Pereira, C.; Groopman, J.D.; Kensler, T.W.; Dashwood, R.H.; Williams, D.E.; et al. Natural chlorophyll inhibits aflatoxin B 1 -induced multi-organ carcinogenesis in the rat. Carcinogenesis 2007, 28, 1294–1302. [Google Scholar] [CrossRef]
- Simonich, M.T.; McQuistan, T.; Jubert, C.; Pereira, C.; Hendricks, J.D.; Schimerlik, M.; Zhu, B.; Dashwood, R.H.; Williams, D.E.; Bailey, G.S. Low-dose dietary chlorophyll inhibits multi-organ carcinogenesis in the rainbow trout. Food Chem. Toxicol. 2008, 46, 1014–1024. [Google Scholar] [CrossRef][Green Version]
- McQuistan, T.J.; Simonich, M.T.; Pratt, M.M.; Pereira, C.B.; Hendricks, J.D.; Dashwood, R.H.; Williams, D.E.; Bailey, G.S. Cancer chemoprevention by dietary chlorophylls: A 12,000-animal dose-dose matrix biomarker and tumor study. Food Chem. Toxicol. 2012, 50, 341–352. [Google Scholar] [CrossRef]
- Chiu, L.C.; Kong, C.K.; Ooi, V.E. The chlorophyllin-induced cell cycle arrest and apoptosis in human breast cancer MCF-7 cells is associated with ERK deactivation and Cyclin D1 depletion. Int. J. Mol. Med. 2005, 16, 735–740. [Google Scholar]
- Xia, J.; Ni, L.; Han, J.; Wang, Y.; Li, Y.; Li, Y.; Tian, Y. Simultaneous aqueous two-phase flotation of sodium chlorophyllin and removal of sugars from saponified solution of bamboo leaves. Chem. Eng. Process. 2016, 101, 41–49. [Google Scholar] [CrossRef]
- Egner, P.A.; Wang, J.B.; Zhu, Y.R.; Zhang, B.C.; Wu, Y.; Zhang, Q.N.; Qian, G.S.; Kuang, S.Y.; Gange, S.J.; Jacobson, L.P.; et al. Chlorophyllin intervention reduces aflatoxin-DNA adducts in individuals at high risk for liver cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 14601–14606. [Google Scholar] [CrossRef]
- Afriyie-Gyawu, E.; Mackie, J.; Dash, B.; Wiles, M.; Taylor, J.; Huebner, H.; Tang, L.; Guan, H.; Wang, J.-S.; Phillips, T. Chronic toxicological evaluation of dietary NovaSil clay in sprague-dawley rats. Food Addit. Contam.: Part B 2005, 22, 259–269. [Google Scholar] [CrossRef]
- Mitchell, N.J.; Kumi, J.; Johnson, N.M.; Dotse, E.; Marroquin-Cardona, A.; Wang, J.-S.; Jolly, P.E.; Ankrah, N.-A.; Phillips, T.D. Reduction in the urinary aflatoxin M1 biomarker as an early indicator of the efficacy of dietary interventions to reduce exposure to aflatoxins. Biomarkers 2013, 18, 391–398. [Google Scholar] [CrossRef]
- Phillips, T.D.; Afriyie-Gyawu, E.; Williams, J.; Huebner, H.; Ankrah, N.A.; Ofori-Adjei, D.; Jolly, P.; Johnson, N.; Taylor, J.; Marroquin-Cardona, A.; et al. Reducing human exposure to aflatoxin through the use of clay: A review. Food Addit. Contam.: Part A 2008, 25, 134–145. [Google Scholar] [CrossRef]
- Wiles, M.; Huebner, H.; Afriyie-Gyawu, E.; Taylor, R.; Bratton, G.; Phillips, T. Toxicological evaluation and metal bioavailability in pregnant rats following exposure to clay minerals in the diet. J. Toxicol. Environ. Health A 2004, 67, 863–874. [Google Scholar] [CrossRef]
- Chung, T.K.; Baker, D.H. Phosphorus utilization in chicks fed hydrated sodium calcium aluminosilicate. J. Anim. Sci. 1990, 68, 1992–1998. [Google Scholar] [CrossRef]
- Hernandez-Mendoza, A.; Garcia, H.S.; Steele, J.L. Screening of Lactobacillus casei strains for their ability to bind aflatoxin B1. Food Chem. Toxicol. 2009, 47, 1064–1068. [Google Scholar] [CrossRef]
- Gratz, S.; Täubel, M.; Juvonen, R.O.; Viluksela, M.; Turner, P.C.; Mykkänen, H.; El-Nezami, H. Lactobacillus rhamnosus strain GG modulates intestinal absorption, fecal excretion, and toxicity of aflatoxin B(1) in rats. Appl. Environ. Microbiol. 2006, 72, 7398–7400. [Google Scholar] [CrossRef]
- El-Nezami, H.; Mykkanen, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of Lactobacillus and Propionibacterium strains to remove aflatoxin B, from the chicken duodenum. J. Food Prot. 2000, 63, 549–552. [Google Scholar] [CrossRef]
- El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem. Toxicol. 1998, 36, 321–326. [Google Scholar] [CrossRef]
- Fazeli, M.R.; Hajimohammadali, M.; Moshkani, A.; Samadi, N.; Jamalifar, H.; Khoshayand, M.R.; Vaghari, E.; Pouragahi, S. Aflatoxin B1 binding capacity of autochthonous strains of lactic acid bacteria. J. Food Prot. 2009, 72, 189–192. [Google Scholar] [CrossRef]
- Topcu, A.; Bulat, T.; Wishah, R.; Boyaci, I.H. Detoxification of aflatoxin B1 and patulin by Enterococcus faecium strains. Int. J. Food Microbiol. 2010, 139, 202–205. [Google Scholar] [CrossRef]
- Kabak, B.; Brandon, E.F.A.; Var, I.; Blokland, M.; Sips, A.J.A.M. Effects of probiotic bacteria on the bioaccessibility of aflatoxin B1 and ochratoxin A using an in vitro digestion model under fed conditions. J. Environ. Sci. Health B 2009, 44, 472–480. [Google Scholar] [CrossRef]
- Ahlberg, S.; Randolph, D.; Okoth, S.; Lindahl, J. Aflatoxin binders in foods for human consumption—Can this be promoted safely and ethically? Toxins (Basel) 2019, 11, 410. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Liu, Z.; Shi, Y.; Liu, J.; Xu, X.; Hao, S.; Mu, P.; Deng, F.; Deng, Y. Aflatoxin B1 degradation and detoxification by Escherichia coli CG1061 isolated from chicken cecum. Front. Pharmacol. 2019, 9. [Google Scholar] [CrossRef]
- Xu, L.; Eisa Ahmed, F.M.; Sangare, L.; Zhao, Y.; Selvaraj, N.J.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel aflatoxin-degrading enzyme from Bacillus shackletonii L7. Toxins (Basel) 2017, 9, 36. [Google Scholar] [CrossRef]
- Wang, C.; Li, Z.; Wang, H.; Qiu, H.; Zhang, M.; Li, S.; Luo, X.; Song, Y.; Zhou, H.; Ma, W.; et al. Rapid biodegradation of aflatoxin B1 by metabolites of Fusarium sp. WCQ3361 with broad working temperature range and excellent thermostability. J. Sci. Food Agric. 2017, 97, 1342–1348. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Yan, H.; Yin, C.; Liu, Y.; Xu, Q.; Liu, X.; Zhang, Z. Effective biodegradation of aflatoxin B1 using the Bacillus licheniformis (BL010) strain. Toxins (Basel) 2018, 10, 497. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- El-Nezami, H.S.; Polychronaki, N.N.; Ma, J.; Zhu, H.; Ling, W.; Salminen, E.K.; Juvonen, R.O.; Salminen, S.J.; Poussa, T.; Mykkänen, H.M. Probiotic supplementation reduces a biomarker for increased risk of liver cancer in young men from Southern China. Am. J. Clin. Nutr. 2006, 83, 1199–1203. [Google Scholar] [CrossRef]
- Petrick, J.L.; McGlynn, K.A. The changing epidemiology of primary liver cancer. Curr. Epidemiol. Rep. 2019, 6, 104–111. [Google Scholar] [CrossRef]
- Sirma, A.J.; Lindahl, J.F.; Makita, K.; Senerwa, D.; Mtimet, N.; Kang’ethe, E.K.; Grace, D. The impacts of aflatoxin standards on health and nutrition in sub-Saharan Africa: The case of Kenya. Glob. Food Secur. 2018, 18, 57–61. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Worldwide Regulations for Mycotoxins in Food and Feed in 2003; FAO Food Nutr. Pap. 81; FAO: Rome, Italy, 2004. [Google Scholar]
- Garcia-Alvarez-Coque, J.-M.; Taghouti, I.; Martinez-Gomez, V. Changes in aflatoxin standards: Implications for EU border controls of nut imports. Appl. Econ. Perspect. Policy 2019. [Google Scholar] [CrossRef]
- van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef]
- Fernandes, T.H.; Ferrão, J.; Bell, V. Mycotoxins, food and health. J. Nutr. Health Food Sci. 2017, 5, 1–10. [Google Scholar]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Effect on public health of a possible increase of the maximum level for ‘aflatoxin total’ from 4 to 10 μg/kg in peanuts and processed products thereof, intended for direct human consumption or use as an ingredient in foodstuffs. EFSA J. 2018, 16, e05175. [Google Scholar]
- World Health Organization. Evaluation of Certain Contaminants in Food: Eighty-Third Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- JECFA (Codex Committee on Contaminants in Foods). Proposed Draft Maximum Level for Total Aflatoxins in Ready-to-Eat Peanuts and Associated Sampling Plan. Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FMeetings%252FCX-735-12%252FCRDs%252Fcf12_CRD10x.pdf (accessed on 8 August 2019).
- Tomašević, I.; Petrović, J.; Jovetić, M.; Raičević, S.; Milojević, M.; Miočinović, J. Two year survey on the occurrence and seasonal variation of aflatoxin M1 in milk and milk products in Serbia. Food Control 2015, 56, 64–70. [Google Scholar] [CrossRef]
- Li, Y.; Xiong, B.; Beghin, J.C. The political economy of food standard determination: International evidence from maximum residue limits. Econ. Work. Pap. (2002–2016) 2014, 47, 239–267. Available online: http://lib.dr.iastate.edu/econ_las_workingpapers/47 (accessed on 8 August 2019).
- Saha Turna, N.; Wu, F. Risk assessment of aflatoxin-related liver cancer in Bangladesh. Food Addit. Contam.: Part A 2019, 36, 320–326. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aflatoxin | Crystallization Solvents | Fluorescence under UV Light (360 nm) | Fluorescence Spectrum | MW | Melting Point °C | Chemical Formula | |
|---|---|---|---|---|---|---|---|
| Excitation Wavelength (nm) | Emission Wavelength (nm) | ||||||
| Aflatoxin B1 | Trichloroethylene/chloroform | Blue | 365 | 425 | 312 | 265–270 | C17H12O6 |
| Aflatoxin B2 | Methanol | Blue | 365 | 425 | 314 | 305–309 | C17H14O6 |
| Aflatoxin G1 | Methanol | Green | 365 | 450 | 328 | 247–250 | C17H12O7 |
| Aflatoxin G2 | Ethanol | Green-Blue | 365 | 450 | 330 | 237–240 | C17H14O7 |
| Year | Study | Action/Outcome | References |
|---|---|---|---|
| 1960 | Outbreak of turkey “X” disease in the Eastern and Southern regions of London (England) poultry farms | Starting a wide investigation to understand the disease | [2] |
| 1961 | Association of the X disease to the imported Brazilian groundnut meal | Removal of Brazilian groundnut from rations and reduction of the disease incidence | [2] |
| In vivo confirmation of the toxicity of Brazilian groundnut to other birds and animals | Characterization of typical symptoms and organ lesions caused by the disease | [2,18,19,20,21,22,23] | |
| Preparation of a concentrate of the toxic principle in an aqueous suspension | Development of reliable quantitative biological toxicity assay | [22] | |
| Association of Aspergillus flavus contamination with the toxicity of groundnut | Establishing the relation between toxic fractions from cultures of A. flavus and fluorescence under UV light | [18] | |
| 1962 | Improving separation and purification techniques of A. flavus culture extracts by thin-layer chromatography (TLC) | New separation methods for aflatoxin purification Identification of aflatoxins B and G | [30] |
| Developing crystallization methods Studies on physicochemical properties of aflatoxins | Preparation of crystalline aflatoxins Isolation and chemical characterization of aflatoxin B1 | [29,31,32] | |
| 1962 | Animal trials for the fate of aflatoxin after ingestion of contaminated feed | Detection of a toxic principle, “milk toxin”, in milk drawn from cows fed on toxic groundnut | [36] |
| 1963 | Purification and crystallization of aflatoxins | Identification and chemical characterization of aflatoxins B1, B2, G1 and G2 | [33] |
| First chemical synthesis of aflatoxins G1 and G2 | |||
| Structural studies of aflatoxins B and G | Elucidation of the chemical structures of aflatoxins B and G as difuranocoumarin derivatives | [34,35] | |
| 1964 | Purification and toxicity testing of the “milk toxin” in dry milk | Partial characterization of the “milk toxin” and its relatedness to aflatoxin B1 | [40] |
| 1965 | Setting safety standard by the United States Food and Drug Administration (US FDA) | First regulation of total aflatoxins (ML of 30 ppb in foods) in the USA. | [60] |
| 1966 | Sheep fed on a mixture of aflatoxins to determine organs and/or body fluids secretions where the “milk toxin” is accumulated or secreted | Detection of “milk toxin” in the milk, urine, kidney, and liver Designation of the “milk toxin” as “aflatoxin M” | [38] |
| Purification and fractionation of Aflatoxin M from sheep urine | Separation and physicochemical characterization of aflatoxins M1 and M2 as hydroxylated metabolites of aflatoxins B1 and B2, respectively | [39] | |
| Feeding malnourished African children with meals supplemented with peanut as part of an initiative of the United Nations Food and Agricultural Organization (FAO) to control kwashiorkor | Liver damage in most children having been beneficiary participants in the initiative | [51] | |
| 1967 | Studies on in vitro chemical characterization of aflatoxins | Complete chemical synthesis of aflatoxin B1 | [82] |
| 1969 | Revising standard levels by the US FDA | Action level for total aflatoxins in foods lowered from 30 ppb to 20 ppb | [60] |
| 1970 | A case-control study on liver failure leading to the death of a teenager fed on mouldy cassava in Uganda | Circumstantial evidence of the implication of aflatoxins in acute intoxication | [54] |
| 1971 | First review by the International Agency for Research on Cancer (IARC) working group of the available studies on the possible relationship between aflatoxin intake and liver cancer | Studies reviewed were considered to provide a circumstantial evidence for the carcinogenicity of aflatoxins in humans | [56] |
| 1975 | Second review by the IARC working group of the previous and newly generated data on the causality between aflatoxins and liver cancer | Confirmation of the previous status of “circumstantial evidence” for carcinogenicity in humans | [61] |
| 1977 | Regulatory action guidelines of the US FDA | Action level of 0.5 ppb for aflatoxin M1 in milk | [83] |
| 1979 | Building evidence for a link between liver damage and aflatoxin intake | The FAO establishes the first provisional acceptable limit of 30 mg aflatoxin per kg meal | [53] |
| 1987 | Third review of new available data generated from better-designed studies to address previous recommendations of the IARC working group | Classification of naturally occurring mixtures of aflatoxins in group 1 carcinogens | [62] |
| 1991 | Development of mechanistic studies to demonstrate the carcinogenicity of aflatoxins at the molecular level | Demonstration of the genotoxicity of aflatoxin by induction of point mutation in codon 249 of TP53 tumor suppressor gene | [71] |
| 1992 | High-quality design of epidemiological and mechanistic studies on the carcinogenicity of aflatoxins | Establishment of an almost linear relationship between AFB1 intake and liver cancer Demonstration of synergistic action between dietary intake of aflatoxins and hepatitis virus B hepatocellular carcinoma First use of Aflatoxin P1, Aflatoxin M1, and DNA-adduct in urine as a biomarker for the exposure assessment | [63] |
| Review of the newly generated data by the IARC working group | Addition of Aflatoxin B1 to the group 1 carcinogens | [66] | |
| 1997 | Cohort studies taking into account available biomarkers to confirm the carcinogenicity of Aflatoxin B1 by | Recommendation of wider use of biomarkers as reliable tools to assess exposure to aflatoxins and for aflatoxicosis diagnostic | [64,65] |
| 2002 | Review of new and previous data on the carcinogenicity of aflatoxins by the IARC working group | Confirmation of the previous status of aflatoxins | [67] |
| 2004 | Investigations on a large aflatoxicosis that occurred in Kenya | The first use of aflatoxin-albumin adduct in blood serum as a biomarker for aflatoxin exposure | [70] |
| 2012 | Review of previous and new data on aflatoxin carcinogenicity with an emphasis on mechanistic studies on the genotoxicity of aflatoxins and biomarkers | Revision of the previous classification to consider aflatoxins, implicitly including Aflatoxins B1, B2, G1, G2, and M1 in group 1 carcinogens * | [74] |
| Biomarker | Limitations | Strengths | Detection Specimen | Validation Status | References |
|---|---|---|---|---|---|
| Aflatoxin B1 (parent) * | No correlation with the ingested amount of the aflatoxin | Useful when used along with other biomarkers | Urine and serum | No | [65,133,134] |
| Phase I Metabolites | |||||
| Aflatoxicol * | Lack of correlation with aflatoxin intake | May be useful when used along with other biomarkers | Urine | No | [135] |
| Aflatoxin M1 * | Significance for short term exposure only | Major aflatoxin B1 metabolite excreted in the urine. Highly correlated with aflatoxin B1 dietary intake. Evidence for a dose-response relationship with hepatocellular carcinoma (HCC) | Urine | Yes | [65,122,123] |
| Aflatoxin P1 * | Significance for short term exposure only. Lack of correlation with aflatoxin B1 intake | Useful when associated with other biomarkers in providing information on the risk of disease onset and diagnosis | Urine | No | [63,65,117] |
| Aflatoxin Q1 * | Significance short term exposure only. Rarely detected in the urine after exposure to aflatoxin B1 | May be useful if used along with other biomarkers | Urine | No | [65,136,137] |
| Aflatoxin B2 a * | Does not necessarily reflect the DNA damaging effect of aflatoxin B1. Lack of correlation with aflatoxin B1 intake | One of the major metabolites of aflatoxin B1 which may inform on acute toxicity (forms adducts with proteins and phospholipids) | Serum, Urine | No | [116,138] |
| Aflatoxin B1-8,9-dihydro-diol ** | Not excreted in biofluids; no easily accessible or available samples | May be a good indicator for acute toxicity, as it leads to the formation of adducts with functional proteins | Liver (in vitro) | No | [117] |
| Aflatoxin B1-exo-8,9-epoxide ** | Not excreted in biofluids; no easily accessible or available samples. Short-lived, very unstable intermediate metabolite (difficult to quantitate accurately any time after ingestion) | Directly related to toxicity mechanism; best risk marker for the aflatoxin intake, dose/response determinations, and prediction of the disease onset | Liver (in vitro) | No | [72,139,140] |
| Phase II Metabolites (Aflatoxin-Protein Adducts) | |||||
| Aflatoxin B1-lysine ** | Limited value regarding quantitative risk assessment and the prediction of HCC at early stages | Chronic exposure (stable for more than 3 months in serum). Highly correlated with aflatoxin B1 dietary intake Best used to associate aflatoxin intake and child growth impairment | Blood serum | Yes | [84,113,129,131,141,142] |
| Phase II Metabolites (Aflatoxin DNA Adducts) | |||||
| Aflatoxin B1-N7-Guanine ** | Not valid for a long-term exposure or to predict the onset of liver cancer | Linear proportionality with ingested aflatoxin B1. Confirmation of the etiology of aflatoxins in HCC | Urine | Yes | [122,130] |
| Aflatoxin B1-FAPY ** | Not excreted in biofluids; no easily accessible or available samples | Directly implicated in DNA mutations leading to cancer; suitable marker for dose-response determination and disease outcome | Liver, kidney, viscera (in vitro) | No | [119] |
| Aflatoxin B1-8,9-dihydro-diol ** | Idem as aflatoxin B1-FAPY | May be a good indicator for acute toxicity, as it leads to the formation of adducts with functional proteins | Liver (in vitro) | No | [117] |
| Aflatoxin B-exo-8,9-epoxide ** | Idem as aflatoxinB1-FAPY Short-lived, very unstable intermediate metabolite | Directly related to toxicity mechanism; best risk marker for the aflatoxin intake, dose/response determinations, and prediction of the disease onset | Liver (in vitro) | No | [72,139,140] |
| Aflatoxin-glutathione | Idem as aflatoxin B1-FAPY. Unavailability for sampling; only detected in bile | Best indicator for the detoxification of aflatoxins | Bile | No | [143] |
| Aflatoxin-mercapturic acid * | Low level in urine does not necessarily indicate a shift towards the genotoxic pathway if the phase I enzymes are inhibited (low levels of epoxides) | Good risk biomarker to monitor the outcome of chemoprotective actions; reflects the induction of the detoxification pathway via aflatoxin B1–GST conjugate formation (phase II metabolites activation) | Urine | No | [120,125,144] |
| Aflatoxin-glucuronide | Insufficiently studied | Indicator of the of aflatoxin aldehyde reductase (AFAR) activity; potential use to monitor chemoprotection action by administering AFAR inducers | Urine | No | [80] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benkerroum, N. Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint. Int. J. Environ. Res. Public Health 2019, 16, 3633. https://doi.org/10.3390/ijerph16193633
Benkerroum N. Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint. International Journal of Environmental Research and Public Health. 2019; 16(19):3633. https://doi.org/10.3390/ijerph16193633
Chicago/Turabian StyleBenkerroum, Noreddine. 2019. "Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint" International Journal of Environmental Research and Public Health 16, no. 19: 3633. https://doi.org/10.3390/ijerph16193633
APA StyleBenkerroum, N. (2019). Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint. International Journal of Environmental Research and Public Health, 16(19), 3633. https://doi.org/10.3390/ijerph16193633