
The Vaginal Microbiota, Human Papillomavirus, and Cervical Dysplasia—A Review

Abstract
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
- Analysis of the composition of the vaginal microbiota assessed by molecular methods (next-generation sequencing);
- Groups of reproductive-age and older women with different vaginal microbiota composition, women without HPV infection (healthy) and with HPV infection or cervical intraepithelial neoplasia.
- Conference papers or editorials;
- Duplicated studies.
2.3. Data Analysis
2.4. Study Selection
3. Results
3.1. HPV Prevalence and Integration Mechanism
3.2. Factors Influencing the Composition of the Vaginal Microbiota
3.3. Composition and Functions of the Vaginal Microbiota
3.4. The Role of Lactobacillus Iners
3.5. Altered Vaginal Microbiota and Pathogenic Mechanisms of BV
3.6. The Role of Vaginal Microbiota in Pregnancy Outcomes and Preterm Birth
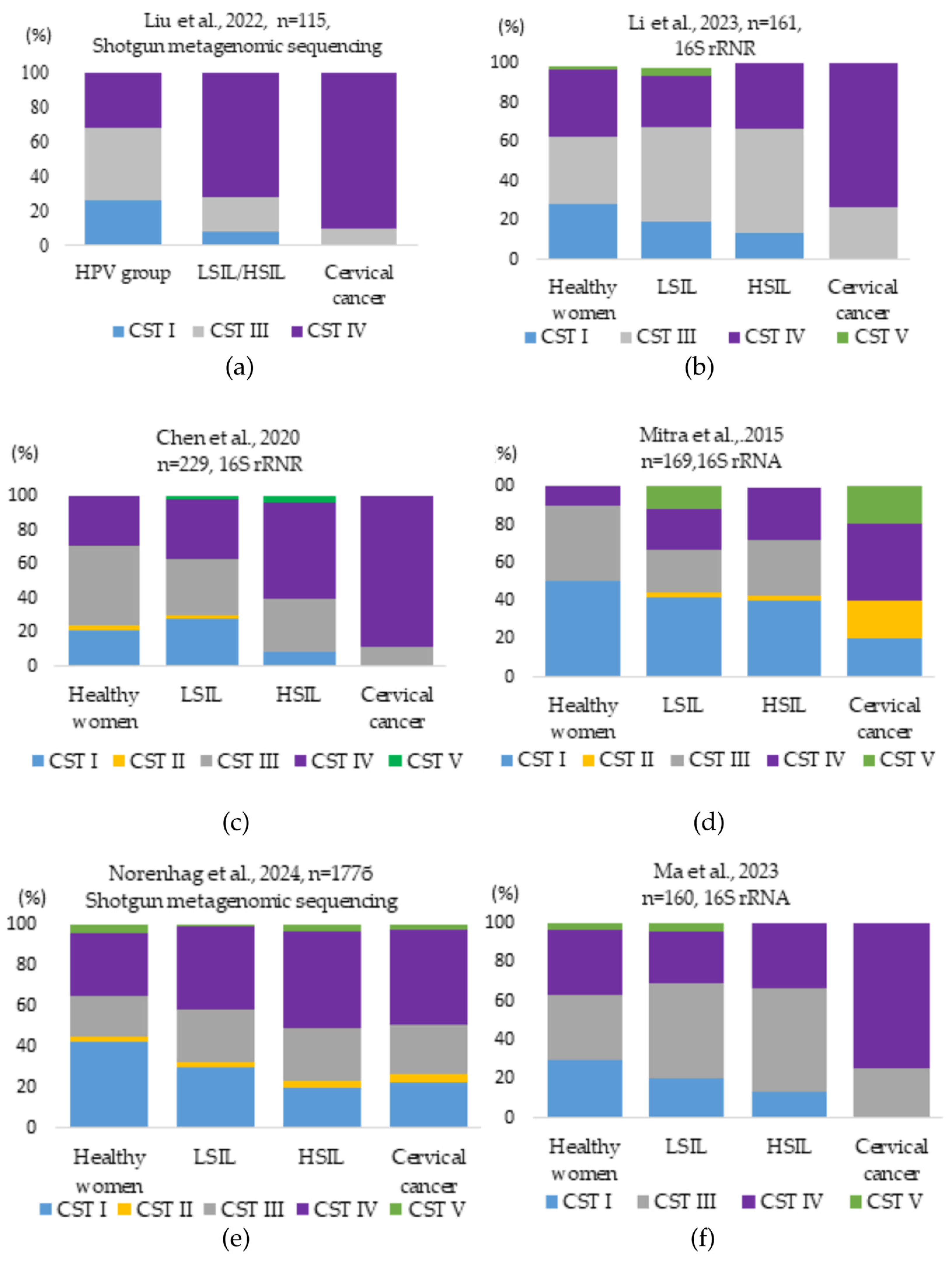
3.7. Relationship Between Vaginal Microbiota, HPV Infection, and Cervical Dysplasia
3.8. Potential for Modeling Vaginal Microbiota to Reduce the Risk of Cervical Lesions: The Role of Probiotics, Antibiotics and Vaccination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Authors, Year | Participants | Sample Types | Analysis Method | Healthy Patients | CIN1 or LSIL | CIN2/3 or HSIL | Cervical Cancer | Key Findings |
|---|---|---|---|---|---|---|---|---|
| Norenhag et al., 2024 [47] | 354 | Cervical and vaginal samples | Shotgun metagenomics sequencing | L. crispatus L. iners | L. iners, G. vaginalis, Peptoniphylus lacrimalis, Megasphaera spp. | Anaerobic bacteria, Gardnerella vaginalis, Prevotella spp., Alloscardovia omnicolens | No data | The CIN group was characterized by microbial diversity, more genes involved in nucleotide biosynthesis, and peptidoglycan biosynthesis. In the control group, the abundance of genes related to amino acid biosynthesis (L-lysine) and sugar degradation was higher. |
| Zheng et al., 2023 [79] | 341 | Cervical cells, tissue and vaginal secretions | Shotgun metagenomic sequencing | L. crispatus L. iners | Gardnerella vaginalis, L. iners | Gardnerella vaginalis, L. iners, S. aureus | Staphylococcus aureus, Phocaeicolavulgatus, Salmonella enterica, G. vaginalis, Bacteroides fragilis | Dysbiosis of the cervical and vaginal microbiota may lead to the enrichment of CpG motifs, which is associated with increased TLR9 expression and may promote the progression of cervical lesions. |
| Liu et al., 2022 [20] | 115 | Cervical cells and vaginal secretions | Shotgun metagenomic sequencing | Lactobacillus spp. | Gardnerella vaginalis, Prevotella bivia | G. vaginalis, Prevotella spp., Anaerobic bacteria | Anaerobic bacteria CST IV | As cervical lesions progressed, Lactobacillus abundance decreased, anaerobic and aerobic bacteria increased, HPV prevalence and E6/E7 oncogene expression were observed. |
| Kwon et al., 2019 [80] | 47 | Cervical cells and vaginal secretions | Shotgun metagenomic sequencing | Pseudoalteromonas, Psychrobacter | No data | Lactobacillus, Staphylococcus, Candidatus Endolissoclinum | Alkaliphilus, Pseudothermotoga, Wolbachia | As CIN progresses, peptidoglycan synthesis processes increase, dioxin degradation and 4-oxalocrotonate tautomerase activities decrease. |
| So et al., 2020 [60] | 50 | Cervicovaginal swab specimens | 16S rRNA V3–V4 region | Lactobacillus spp. | L. iners, Sneathia sanguinegens, Gardnerella vaginalis, Prevotella timonensis, Prevotella buccalis | L. iners, G. vaginalis, Sneathia sanguinegens, Atopobium vaginae, Porphyromonas somerae, Prevotella disiens | Fusobacterium spp., Porphyromonas spp. Prevotella timonensis, Escherichia fergusonii, Haemophilus | G. vaginalis is associated with persistent HPV infection and a high risk of developing CIN2/3 and cervical cancer. |
| Teka et al., 2023 [83] | 120 | Cervical cells collected using different brushes | 16S rRNA V4 region | Lactobacillus spp. | L. iners | L. iners | Porphyromonas somerae, Prevotella timonensis, Porphyromonas asaccharolytica, Bacteroides, Anaerococcus | The diversity, composition and relative abundance of the microbiota differ between women with cervical cancer, women with dysplasia, and healthy women. |
| Yang et al., 2024 [106] | 26 | Cervical smear | 16S rRNA V3–V4 region | Lactobacillus spp. | L. iners. Atopobium, Prevotella, Bacteroides | Sneathia Firmicutes Proteobacteria | No data | HPV infection affects metabolic signals, inducing changes in glycolysis, lactate accumulation, and lipid metabolism. Specific metabolites have been identified to distinguish healthy patients from patients with persistent HR-HPV infection. |
| Wu et al., 2021 [87] | 94 | Cervical smear | 16S rRNA V4 region | Lactobacillus spp. | Lactobacillus, Xanthobacter, Thermus, Flavisolibacter Sphingopyxis, Sediminibacterium, Geobacillus | Sneathia, Gardnerella, Megasphera | Porphyromonas, Campylobacter, Prevotella | Identified potential markers of cervical cancer: Porphyromonas and Campylobacter. Functional metabolic pathways associated with Prevotella cause a negative effect, associated with Lactobacillus-positive status. |
| Molina et al., 2022 [49] | 541 | Cervical smear | 16S rRNA V3–V4 region | Lactobacillus spp. Lactobacillus acidophilus | L. iners, Gardnerella vaginalis. L. crispatus | Megasphaera genomosp 1 tipas, Atopobium vaginae, Dialister micraerophilus, Dialister micraerophilus | Megasphaera genomosp 1 type G. vaginalis | The distribution of microbial communities in the microbiome reveals changes in microbial dynamics that may be associated with HR-HPV infection and cervical lesions. |
| Fan et al., 2024 [46] | 125 | Cervical smear | 16S rRNA V4 region | Phylum Firmicutes, Lactobacillus, Streptococcus, Varibaculum (biomarker) | Saccharopolyspora (biomarker), Corynebacterium | Saccharopolyspora (biomarker) Prevotella, Megasphaera, Streptococcus, Sneathia, Dialister, Anaerococcus, Ureaplasma, Peptoniphilus, Mycoplasma | Proteobacteria, Corynebacterium, Coprococcus, Peptococcus, Ruminococcus (biomarkers), Dialister, Peptoniphilus | The microbiota has diagnostic value in distinguishing between healthy women, patients with CIN, and women with cervical lesions. In addition, microorganisms may be potential biomarkers for cervical cancer |
| Ma et al., 2023 [82] | 160 | Cervical smear | 16S rRNA | Lactobacillus spp., L. crispatus, Gardnerella, Prevotella, Fannyhessea vaginae | Lactobacillus spp., Sneathia, Dialister, Gardnerella | Lactobacillus spp., L. iners, Sneathia, Fastidiosipila Megasphaera, Prevotella, Bacteroides. | Prevotella, Bacteroides, Finegoldia, Gardanella, Streptococcus, Fannyhessea vaginae, Peptostreptococcus, Dialister, Veillonella | A gradual decrease in L. crispatus and an increase in anaerobic bacteria are associated with the severity of cervical lesions. HPV 16 infection is associated with a greater diversity of vaginal microbiota and a decrease in Lactobacillus. L. crispatus and L. iners can be considered as biomarkers. |
| Liu et al., 2023 [85] | 692 | Vaginal discharge | 16S rRNA V4 region | Lactobacillus spp. | Ercella, Bacillus, B Lautia, Terrisporobacter, Sporobacter, Romboutsia | Prevotella, Gardnerella, Sneathia, Megasphaera, Dialister, Ureaplasma | No data | CIN lesions are characterized by a decrease in Lactobacillus and Pseudomonas and an increase in Gardnerella, Prevotella and Dialister. High-grade CIN may inhibit the phosphotransferase system, transcription factors, fructose and mannose metabolism, aminosugar and nucleotide metabolism, and galactose metabolism. |
| Wu et al., 2020 [84] | 69 | Cervical smear | 16S rRNA V3–V4 region | Lactobacillus spp., | Lactobacillus, Gardnerella, Atopobium, Sneathia, Streptococcus | Delftia (biomarker), Gardnerella, Prevotella, Atopobium, Sneathia, Streptococcus | No data | The vaginal microbiota is directly or indirectly associated with the progression of CIN. |
References
- Zhang, Y.; Xu, X.; Yu, L.; Shi, X.; Min, M.; Xiong, L.; Pan, J.; Zhang, Y.; Liu, P.; Wu, G.; et al. Vaginal Microbiota Changes Caused by HPV Infection in Chinese Women. Front. Cell. Infect. Microbiol. 2022, 12, 814668. [Google Scholar] [CrossRef]
- Mitra, A.; MacIntyre, D.A.; Lee, Y.S.; Smith, A.; Marchesi, J.R.; Lehne, B.; Bhatia, R.; Lyons, D.; Paraskevaidis, E.; Li, J.V.; et al. Cervical Intraepithelial Neoplasia Disease Progression Is Associated with Increased Vaginal Microbiome Diversity. Sci. Rep. 2015, 5, 16865. [Google Scholar] [CrossRef] [PubMed]
- Gardella, B.; Pasquali, M.F.; La Verde, M.; Cianci, S.; Torella, M.; Dominoni, M. The Complex Interplay between Vaginal Microbiota, HPV Infection, and Immunological Microenvironment in Cervical Intraepithelial Neoplasia: A Literature Review. Int. J. Mol. Sci. 2022, 23, 7174. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, M.; Atefmehr, L.; Motlaghzadeh, S.; Hejami, F.; Elyasi, F.S.; Zafarian, N.; Taghiabadi, Z.; Aboofazeli, A.; Yarahmady, H.; Modaresi, P.; et al. Prevailing of HPV-16 and 52 Genotype in 2022–2023 in Sanandaj, Iran. Virol. J. 2024, 21, 106. [Google Scholar] [CrossRef] [PubMed]
- Manini, I.; Montomoli, E. Epidemiology and Prevention of Human Papillomavirus. Ann. Ig. Med. Prev. Comunita 2018, 30, 28–32. [Google Scholar] [CrossRef]
- Mukherjee, A.G.; Ramesh Wanjari, U.; Valsala Gopalakrishnan, A.; Jayaraj, R.; Katturajan, R.; Kannampuzha, S.; Murali, R.; Namachivayam, A.; Evan Prince, S.; Vellingiri, B.; et al. HPV-Associated Cancers: Insights into the Mechanistic Scenario and Latest Updates. Med. Oncol. 2023, 40, 212. [Google Scholar] [CrossRef]
- Liu, Y.; Li, T.; Guo, R.; Chen, T.; Wang, S.; Wu, D.; Li, J.; Liu, Z.; Zhao, Y.; Yin, J.; et al. The Vaginal Microbiota among the Different Status of Human Papillomavirus Infection and Bacterial Vaginosis. J. Med. Virol. 2023, 95, e28595. [Google Scholar] [CrossRef]
- Avitabile, E.; Menotti, L.; Croatti, V.; Giordani, B.; Parolin, C.; Vitali, B. Protective Mechanisms of Vaginal Lactobacilli against Sexually Transmitted Viral Infections. Int. J. Mol. Sci. 2024, 25, 9168. [Google Scholar] [CrossRef]
- Huang, R.; Liu, Z.; Sun, T.; Zhu, L. Cervicovaginal Microbiome, High-Risk HPV Infection and Cervical Cancer: Mechanisms and Therapeutic Potential. Microbiol. Res. 2024, 287, 127857. [Google Scholar] [CrossRef]
- Janicka-Kośnik, M.; Sarecka-Hujar, B.; Jakiel, G.; Słabuszewska-Jóźwiak, A. Cervicovaginal Microbiome after Cervical Intraepithelial Neoplasia Treatment. A Protocol for Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2021, 18, 9050. [Google Scholar] [CrossRef]
- Kyrgiou, M.; Moscicki, A.B. Vaginal Microbiome and Cervical Cancer. Semin. Cancer Biol. 2022, 86, 189–198. [Google Scholar] [CrossRef]
- Tossas, K.Y.; Zhu, B.; Perera, R.A.; Serrano, M.G.; Sullivan, S.; Sayeed, S.; Strauss, J.F.; Winn, R.A.; Buck, G.A.; Seewaldt, V.L. Does the Vaginal Microbiome Operate Differently by Race to Influence Risk of Precervical Cancer? J. Women’s Health 2023, 32, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X. Vaginal Microbiome Distinction in Women with HPV+, Cervical Intraepithelial Neoplasia, and Cervical Cancer, a Retrospective Study. Front. Cell. Infect. Microbiol. 2025, 14, 1483544. [Google Scholar] [CrossRef]
- Leon-Gomez, P.; Romero, V.I. Human Papillomavirus, Vaginal Microbiota and Metagenomics: The Interplay between Development and Progression of Cervical Cancer. Front. Microbiol. 2025, 15, 1515258. [Google Scholar] [CrossRef] [PubMed]
- Musa, J.; Maiga, M.; Green, S.J.; Magaji, F.A.; Maryam, A.J.; Okolo, M.; Nyam, C.J.; Cosmas, N.T.; Silas, O.A.; Imade, G.E.; et al. Vaginal Microbiome Community State Types and High-Risk Human Papillomaviruses in Cervical Precancer and Cancer in North-Central Nigeria. BMC Cancer 2023, 23, 683. [Google Scholar] [CrossRef]
- Ye, J.; Zheng, L.; He, Y.; Qi, X. Human Papillomavirus Associated Cervical Lesion: Pathogenesis and Therapeutic Interventions. MedComm 2023, 4, e368. [Google Scholar] [CrossRef]
- Wolf, J.; Kist, L.F.; Pereira, S.B.; Quessada, M.A.; Petek, H.; Pille, A.; Maccari, J.G.; Mutlaq, M.P.; Nasi, L.A. Human Papillomavirus Infection: Epidemiology, Biology, Host Interactions, Cancer Development, Prevention, and Therapeutics. Rev. Med. Virol. 2024, 34, e2537. [Google Scholar] [CrossRef] [PubMed]
- Tosado-Rodríguez, E.; Mendez, L.B.; Espino, A.M.; Dorta-Estremera, S.; Aquino, E.E.; Romaguera, J.; Godoy-Vitorino, F. Inflammatory Cytokines and a Diverse Cervicovaginal Microbiota Associate with Cervical Dysplasia in a Cohort of Hispanics Living in Puerto Rico. PLoS ONE 2023, 18, e0284673. [Google Scholar] [CrossRef]
- Chen, Y.; Qiu, X.; Wang, W.; Li, D.; Wu, A.; Hong, Z.; Di, W.; Qiu, L. Human Papillomavirus Infection and Cervical Intraepithelial Neoplasia Progression Are Associated with Increased Vaginal Microbiome Diversity in a Chinese Cohort. BMC Infect. Dis. 2020, 20, 629. [Google Scholar] [CrossRef]
- Liu, H.; Liang, H.; Li, D.; Wang, M.; Li, Y. Association of Cervical Dysbacteriosis, HPV Oncogene Expression, and Cervical Lesion Progression. Microbiol. Spectr. 2022, 10, e0015122. [Google Scholar] [CrossRef]
- Magalhães, G.M.; Vieira, É.C.; Garcia, L.C.; De Carvalho-Leite, M.d.L.R.; Guedes, A.C.M.; Araújo, M.G. Update on Human Papilloma Virus—Part I: Epidemiology, Pathogenesis, and Clinical Spectrum. An. Bras. Dermatol. 2021, 96, 1–16. [Google Scholar] [CrossRef]
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global Burden of Cancer Attributable to Infections in 2018: A Worldwide Incidence Analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef]
- Parama, D.; BharathwajChetty, B.; Jayaprakash, S.; Lee, E.H.C.; Khatoon, E.; Alqahtani, M.S.; Abbas, M.; Kumar, A.P.; Kunnumakkara, A.B. The Emerging Role of Human Papillomavirus in Lung Cancer. Life Sci. 2024, 351, 122785. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Chang, J.; Wei, K. Correlation between Breast Cancer and Human Papillomavirus (HPV) Infection. Heliyon 2024, 10, e37027. [Google Scholar] [CrossRef]
- Hong, L.J.; Huynh, S.; Kim, J.; Denham, L.; Momeni, M.; Ioffe, Y.J.M. Margin Status Post Cervical Conization Predicts Residual Adenocarcinoma In Situ (AIS) and Occult Adenocarcinoma in a Predominantly Hispanic Population. Diagnostics 2021, 11, 1889. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.M.; Wentzensen, N.; Castle, P.E.; Schiffman, M.; Zuna, R.; Arend, R.C.; Clarke, M.A. Racial and Ethnic Disparities in Cervical Cancer Incidence, Survival, and Mortality by Histologic Subtype. J. Clin. Oncol. 2022, 41, 1059. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, A.A.; Gargano, J.W.; Park, I.U.; Griffin, M.R.; Niccolai, L.M.; Powell, M.; Bennett, N.M.; Saadeh, K.; Pemmaraju, M.; Higgins, K.; et al. Cervical Adenocarcinoma in Situ: Human Papillomavirus Types and Incidence Trends in Five States, 2008–2015. Int. J. Cancer 2019, 146, 810. [Google Scholar] [CrossRef]
- Smith, A.J.B.; Beavis, A.L.; Rositch, A.F.; Levinson, K. Disparities in Diagnosis and Treatment of Cervical Adenocarcinoma Compared to Squamous Cell Carcinoma: An Analysis of the National Cancer Database, 2004–2017. J. Low. Genit. Tract Dis. 2022, 27, 29. [Google Scholar] [CrossRef]
- Tergas, A.; Byron, S.; Zhang, G.; Izatt, T.; Zhang, J.; Schork, N.; Rodriguez, L. YIA24-007: Genetic Ancestry and Genomic Alterations of Cervical Cancer in a Diverse Patient Population. J. Natl. Compr. Cancer Netw. 2024, 22. [Google Scholar] [CrossRef]
- Massad, L.S.; Clarke, M.A.; Perkins, R.B.; Garcia, F.; Chelmow, D.; Cheung, L.C.; Darragh, T.M.; Egemen, D.; Lorey, T.S.; Nayar, R.; et al. Applying Results of Extended Genotyping to Management of Positive Cervicovaginal Human Papillomavirus Test Results: Enduring Guidelines. J. Low. Genit. Tract Dis. 2025, 29, 134. [Google Scholar] [CrossRef]
- Frąszczak, K.; Barczyński, B.; Kondracka, A. Does Lactobacillus Exert a Protective Effect on the Development of Cervical and Endometrial Cancer in Women? Cancers 2022, 14, 4909. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; MacIntyre, D.A.; Marchesi, J.R.; Lee, Y.S.; Bennett, P.R.; Kyrgiou, M. The Vaginal Microbiota, Human Papillomavirus Infection and Cervical Intraepithelial Neoplasia: What Do We Know and Where Are We Going Next? Microbiome 2016, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Ehret, A.; Bark, V.N.; Mondal, A.; Fehm, T.N.; Hampl, M. Regression Rate of High-Grade Cervical Intraepithelial Lesions in Women Younger than 25 Years. Arch. Gynecol. Obstet. 2022, 307, 981. [Google Scholar] [CrossRef]
- Pangarkar, M.A. The Bethesda System for Reporting Cervical Cytology. CytoJournal 2022, 19, 28. [Google Scholar] [CrossRef]
- Alimena, S.; Davis, J.; Fichorova, R.N.; Feldman, S. The Vaginal Microbiome: A Complex Milieu Affecting Risk of Human Papillomavirus Persistence and Cervical Cancer. Curr. Probl. Cancer 2022, 46, 100877. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Comprehensive Cervical Cancer Control: A Guide to Essential Practice, 2nd ed.; World Health Organization: Geneva, Switzerland, 2014; pp. 366–378. [Google Scholar]
- Eun, T.J.; Perkins, R.B. Screening for Cervical Cancer. Med. Clin. North Am. 2020, 104, 1063. [Google Scholar] [CrossRef]
- PDQ Screening; Prevention Editorial Board. Cervical Cancer Screening (PDQ®); National Cancer Institute: Bethesda, MD, USA, 2024; pp. 1–26. [Google Scholar]
- Ronco, G.; Dillner, J.; Elfström, K.M.; Tunesi, S.; Snijders, P.J.F.; Arbyn, M.; Kitchener, H.; Segnan, N.; Gilham, C.; Giorgi-Rossi, P.; et al. Efficacy of HPV-Based Screening for Prevention of Invasive Cervical Cancer: Follow-up of Four European Randomised Controlled Trials. Lancet 2014, 383, 524–532. [Google Scholar] [CrossRef]
- Gonçalves-Nobre, J.G.; Matos, A.; Carreira, M.; Santos, A.C.; Veiga, L.C.; Ginete, C.; Brito, M.; Pires, M.; Pereira, H.; Cardoso, C.; et al. The Interplay between HPV, Other Sexually Transmissible Infections and Genital Microbiome on Cervical Microenvironment (MicroCervixHPV Study). Front. Cell. Infect. Microbiol. 2024, 13, 1251913. [Google Scholar] [CrossRef]
- Wang, H.; Ma, Y.; Li, R.; Chen, X.; Wan, L.; Zhao, W. Associations of Cervicovaginal Lactobacilli With High-Risk Human Papillomavirus Infection, Cervical Intraepithelial Neoplasia, and Cancer: A Systematic Review and Meta-Analysis. J. Infect. Dis. 2019, 220, 1243–1254. [Google Scholar] [CrossRef]
- Morsli, M.; Gimenez, E.; Magnan, C.; Salipante, F.; Huberlant, S.; Letouzey, V.; Lavigne, J.P. The Association between Lifestyle Factors and the Composition of the Vaginal Microbiota: A Review. Eur. J. Clin. Microbiol. Infect. Dis. 2024, 43, 1869–1881. [Google Scholar] [CrossRef]
- Głowienka-Stodolak, M.; Bagińska-Drabiuk, K.; Szubert, S.; Hennig, E.E.; Horala, A.; Dąbrowska, M.; Micek, M.; Ciebiera, M.; Zeber-Lubecka, N. Human Papillomavirus Infections and the Role Played by Cervical and Cervico-Vaginal Microbiota—Evidence from Next-Generation Sequencing Studies. Cancers 2024, 16, 399. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Mageswaran, U.M.; Nisaa, A.A.; Balasubramaniam, S.D.; Rajendran, D.; Ismail, E.H.B.E.; Kadir, M.N.; Oon, C.E.; Tan, C.S.; Sany, S.B.; et al. Roles of Probiotics against HPV through the Gut-Vaginal Axis. Int. J. Gynecol. Obstet. 2024, 169, 1–8. [Google Scholar] [CrossRef]
- Mirzaei, R.; Kavyani, B.; Nabizadeh, E.; Kadkhoda, H.; Asghari Ozma, M.; Abdi, M. Microbiota Metabolites in the Female Reproductive System: Focused on the Short-Chain Fatty Acids. Heliyon 2023, 9, e14562. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Han, D.; Fan, X.; Zeng, Y.; Zhao, L. Analysis of the Correlation between Cervical HPV Infection, Cervical Lesions and Vaginal Microecology. Front. Cell. Infect. Microbiol. 2024, 14, 1405789. [Google Scholar] [CrossRef]
- Norenhag, J.; Edfeldt, G.; Stålberg, K.; Garcia, F.; Hugerth, L.W.; Engstrand, L.; Fransson, E.; Du, J.; Schuppe-Koistinen, I.; Olovsson, M. Compositional and Functional Differences of the Vaginal Microbiota of Women with and without Cervical Dysplasia. Sci. Rep. 2024, 14, 11183. [Google Scholar] [CrossRef]
- Norenhag, J.; Du, J.; Olovsson, M.; Verstraelen, H.; Engstrand, L.; Brusselaers, N. The Vaginal Microbiota, Human Papillomavirus and Cervical Dysplasia: A Systematic Review and Network Meta-Analysis. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.A.; Andralojc, K.M.; Huynen, M.A.; Leenders, W.P.J.; Melchers, W.J.G. In-Depth Insights into Cervicovaginal Microbial Communities and HrHPV Infections Using High-Resolution Microbiome Profiling. NPJ Biofilms Microbiomes 2022, 8, 1–10. [Google Scholar] [CrossRef]
- France, M.T.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A Nearest Centroid Classification Method for Vaginal Microbial Communities Based on Composition. Microbiome 2020, 8, 166. [Google Scholar] [CrossRef]
- Dong, W.; Wang, S.; Wang, X.; Xu, G.; Liu, Q.; Li, Z.; Lv, N.; Pan, Y.; Xiong, Q.; Liu, D.; et al. Characteristics of Vaginal Microbiota of Women of Reproductive Age with Infections. Microorganisms 2024, 12, 1030. [Google Scholar] [CrossRef]
- Audirac-Chalifour, A.; Torres-Poveda, K.; Bahena-Román, M.; Téllez-Sosa, J.; Martínez-Barnetche, J.; Cortina-Ceballos, B.; López-Estrada, G.; Delgado-Romero, K.; Burguete-García, A.I.; Cantú, D.; et al. Cervical Microbiome and Cytokine Profile at Various Stages of Cervical Cancer: A Pilot Study. PLoS ONE 2016, 11, e0153274. [Google Scholar] [CrossRef]
- Roachford, O.S.E.; Alleyne, A.T.; Nelson, K.E. Insights into the Vaginal Microbiome in a Diverse Group of Women of African, Asian and European Ancestries. PeerJ 2022, 10, e14449. [Google Scholar] [CrossRef]
- Wei, X.; Tsai, M.S.; Liang, L.; Jiang, L.; Hung, C.J.; Jelliffe-Pawlowski, L.; Rand, L.; Snyder, M.; Jiang, C. Vaginal Microbiomes Show Ethnic Evolutionary Dynamics and Positive Selection of Lactobacillus Adhesins Driven by a Long-Term Niche-Specific Process. Cell Rep. 2024, 43, 114078. [Google Scholar] [CrossRef]
- Javadi, K.; Ferdosi-Shahandashti, E.; Rajabnia, M.; Khaledi, M. Vaginal Microbiota and Gynecological Cancers: A Complex and Evolving Relationship. Infect. Agents Cancer 2024, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Y.; Yu, L.; Shi, X.; Min, M.; Xiong, L.; Pan, J.; Liu, P.; Wu, G.; Gao, G. A Cross-Sectional Analysis about Bacterial Vaginosis, High-Risk Human Papillomavirus Infection, and Cervical Intraepithelial Neoplasia in Chinese Women. Sci. Rep. 2022, 12, 6609. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, S.; Moll, W.M.; Swidsinski, A. Bacterial Vaginosis—Vaginal Polymicrobial Biofilms and Dysbiosis. Dtsch. Ärzteblatt Int. 2023, 120, 347. [Google Scholar] [CrossRef] [PubMed]
- So, K.A.; Hur, S.Y.; Cho, C.H.; Lee, J.K.; Seong, S.J.; Jeong, D.H.; Ki, M.; Kim, T.J. Menopausal Status Induces Vaginal Dysbiosis in Women with Human Papillomavirus Infection. Sci. Rep. 2024, 14, 7092. [Google Scholar] [CrossRef]
- Łaniewski, P.; Barnes, D.; Goulder, A.; Cui, H.; Roe, D.J.; Chase, D.M.; Herbst-Kralovetz, M.M. Linking Cervicovaginal Immune Signatures, HPV and Microbiota Composition in Cervical Carcinogenesis in Non-Hispanic and Hispanic Women. Sci. Rep. 2018, 8, 7593. [Google Scholar] [CrossRef]
- So, K.A.; Yang, E.J.; Kim, N.R.; Hong, S.R.; Lee, J.H.; Hwang, C.S.; Shim, S.H.; Lee, S.J.; Kim, T.J. Changes of Vaginal Microbiota during Cervical Carcinogenesis in Women with Human Papillomavirus Infection. PLoS ONE 2020, 15, e0238705. [Google Scholar] [CrossRef]
- Qingqing, B.; Jie, Z.; Songben, Q.; Juan, C.; Lei, Z.; Mu, X. Cervicovaginal Microbiota Dysbiosis Correlates with HPV Persistent Infection. Microb. Pathog. 2021, 152, 104617. [Google Scholar] [CrossRef]
- Kyrgiou, M.; Mitra, A.; Moscicki, A.B. Does the Vaginal Microbiota Play a Role in the Development of Cervical Cancer? Transl. Res. J. Lab. Clin. Med. 2017, 179, 168–182. [Google Scholar] [CrossRef]
- Wiik, J.; Sengpiel, V.; Kyrgiou, M.; Nilsson, S.; Mitra, A.; Tanbo, T.; Monceyron Jonassen, C.; Møller Tannæs, T.; Sjøborg, K. Cervical Microbiota in Women with Cervical Intra-Epithelial Neoplasia, Prior to and after Local Excisional Treatment, a Norwegian Cohort Study. BMC Women’s Health 2019, 19, 30. [Google Scholar] [CrossRef]
- Lavitola, G.; Della Corte, L.; De Rosa, N.; Nappi, C.; Bifulco, G. Effects on Vaginal Microbiota Restoration and Cervical Epithelialization in Positive HPV Patients Undergoing Vaginal Treatment with Carboxy-Methyl-Beta-Glucan. BioMed Res. Int. 2020, 2020, 5476389. [Google Scholar] [CrossRef]
- Chen, Y.; Hong, Z.; Wang, W.; Gu, L.; Gao, H.; Qiu, L.; Di, W. Association between the Vaginal Microbiome and High-Risk Human Papillomavirus Infection in Pregnant Chinese Women. BMC Infect. Dis. 2019, 19, 677. [Google Scholar] [CrossRef] [PubMed]
- Irina, P.; Alena, V.; Arsene, M.M.J.; Milana, D.; Alla, P.; Lyudmila, K.; Boris, E. Comparison of Vaginal Microbiota in HPV-Negative and HPV-Positive Pregnant Women Using a Culture-Based Approach. Diagn. Microbiol. Infect. Dis. 2024, 110, 116419. [Google Scholar] [CrossRef]
- Baud, A.; Hillion, K.H.; Plainvert, C.; Tessier, V.; Tazi, A.; Mandelbrot, L.; Poyart, C.; Kennedy, S.P. Microbial Diversity in the Vaginal Microbiota and Its Link to Pregnancy Outcomes. Sci. Rep. 2023, 13, 9061. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Hu, X.; Ying, C. Advances in Research on the Relationship between Vaginal Microbiota and Adverse Pregnancy Outcomes and Gynecological Diseases. Microorganisms 2023, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Gudnadottir, U.; Debelius, J.W.; Du, J.; Hugerth, L.W.; Danielsson, H.; Schuppe-Koistinen, I.; Fransson, E.; Brusselaers, N. The Vaginal Microbiome and the Risk of Preterm Birth: A Systematic Review and Network Meta-Analysis. Sci. Rep. 2022, 12, 7926. [Google Scholar] [CrossRef]
- Prodan-Barbulescu, C.; Bratosin, F.; Folescu, R.; Boeriu, E.; Popa, Z.L.; Citu, C.; Ratiu, A.; Rosca, O.; Ilie, A.C. Analysis of Vaginal Microbiota Variations in the Third Trimester of Pregnancy and Their Correlation with Preterm Birth: A Case-Control Study. Microorganisms 2024, 12, 417. [Google Scholar] [CrossRef]
- Liu, X.; Cao, Y.; Xie, X.; Qin, X.; He, X.; Shi, C.; Zeng, W.; Guo, Y.; Lin, Y. Association between Vaginal Microbiota and Risk of Early Pregnancy Miscarriage. Comp. Immunol. Microbiol. Infect. Dis. 2021, 77, 101669. [Google Scholar] [CrossRef]
- Niyibizi, J.; Niyibizi, J.; Zanré, N.; Mayrand, M.H.; Mayrand, M.H.; Mayrand, M.H.; Trottier, H.; Trottier, H. Association between Maternal Human Papillomavirus Infection and Adverse Pregnancy Outcomes: Systematic Review and Meta-Analysis. J. Infect. Dis. 2020, 221, 1925–1937. [Google Scholar] [CrossRef]
- MacIntyre, D.A.; Chandiramani, M.; Lee, Y.S.; Kindinger, L.; Smith, A.; Angelopoulos, N.; Lehne, B.; Arulkumaran, S.; Brown, R.; Teoh, T.G.; et al. The Vaginal Microbiome during Pregnancy and the Postpartum Period in a European Population. Sci. Rep. 2015, 5, 8988. [Google Scholar] [CrossRef]
- Athanasopoulou, K.; Adamopoulos, P.G.; Scorilas, A. Unveiling the Human Gastrointestinal Tract Microbiome: The Past, Present, and Future of Metagenomics. Biomedicines 2023, 11, 827. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, Y.; Wei, X.; Zhu, J.; Wang, X.; Xie, X.; Lu, W. The Alterations of Vaginal Microbiome in HPV16 Infection as Identified by Shotgun Metagenomic Sequencing. Front. Cell. Infect. Microbiol. 2020, 10, 533812. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Yang, W.; Yan, R.; Chi, J.; Xia, Q.; Yang, Y.; Wang, Y.; Sun, L.; Li, P. Co-Evolution of Vaginal Microbiome and Cervical Cancer. J. Transl. Med. 2024, 22, 559. [Google Scholar] [CrossRef] [PubMed]
- Tango, C.N.; Seo, S.S.; Kwon, M.; Lee, D.O.; Chang, H.K.; Kim, M.K. Taxonomic and Functional Differences in Cervical Microbiome Associated with Cervical Cancer Development. Sci. Rep. 2020, 10, 9720. [Google Scholar] [CrossRef]
- Zhou, Z.; Feng, Y.; Xie, L.; Ma, S.; Cai, Z.; Ma, Y. Alterations in Gut and Genital Microbiota Associated with Gynecological Diseases: A Systematic Review and Meta-Analysis. Reprod. Biol. Endocrinol. RBE 2024, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Hu, N.; Liu, J.; Zhao, K.; Li, H.; Wang, J.; Zhang, M.; Zhang, L.; Song, L.; Lyu, Y.; et al. Cervicovaginal Microbiota Disorder Combined with the Change of Cytosine Phosphate Guanine Motif- Toll like Receptor 9 Axis Was Associated with Cervical Cancerization. J. Cancer Res. Clin. Oncol. 2023, 149, 17371–17381. [Google Scholar] [CrossRef]
- Kwon, M.; Seo, S.S.; Kim, M.K.; Lee, D.O.; Lim, M.C. Compositional and Functional Differences between Microbiota and Cervical Carcinogenesis as Identified by Shotgun Metagenomic Sequencing. Cancers 2019, 11, 309. [Google Scholar] [CrossRef]
- Fang, B.; Li, Q.; Wan, Z.; OuYang, Z.; Zhang, Q. Exploring the Association Between Cervical Microbiota and HR-HPV Infection Based on 16S RRNA Gene and Metagenomic Sequencing. Front. Cell. Infect. Microbiol. 2022, 12, 922554. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Y.; Liu, Y.; Cao, L.; Han, X.; Gao, S.; Zhang, C. Vaginal Microbiome Dysbiosis Is Associated with the Different Cervical Disease Status. J. Microbiol. 2023, 61, 423–432. [Google Scholar] [CrossRef]
- Teka, B.; Yoshida-Court, K.; Firdawoke, E.; Chanyalew, Z.; Gizaw, M.; Addissie, A.; Mihret, A.; Colbert, L.E.; Napravnik, T.C.; El Alam, M.B.; et al. Cervicovaginal Microbiota Profiles in Precancerous Lesions and Cervical Cancer among Ethiopian Women. Microorganisms 2023, 11, 833. [Google Scholar] [CrossRef]
- Wu, M.; Gao, J.; Wu, Y.; Li, Y.; Chen, Y.; Zhao, F.; Li, C.; Ying, C. Characterization of Vaginal Microbiota in Chinese Women with Cervical Squamous Intra-Epithelial Neoplasia. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2020, 30, 1500–1504. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Liu, J.; Su, M.; Diao, X.; Liang, X.; Zhang, J.; Wang, Q.; Zhan, Y. Characteristics of Vaginal Microbiota in Various Cervical Intraepithelial Neoplasia: A Cross-Sectional Study. J. Transl. Med. 2023, 21, 816. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, L.; Han, X.; Ma, Y.; Liu, Y.; Gao, S.; Zhang, C. Altered Vaginal Eukaryotic Virome Is Associated with Different Cervical Disease Status. Virol. Sin. 2023, 38, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Ding, X.; Kong, Y.; Acharya, S.; Wu, H.; Huang, C.; Liang, Y.; Nong, X.; Chen, H. The Feature of Cervical Microbiota Associated with the Progression of Cervical Cancer among Reproductive Females. Gynecol. Oncol. 2021, 163, 348–357. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, Q.; Chen, Y.; Dong, B.; Xue, H.; Lei, H.; Lu, Y.; Wei, X.; Sun, P. Changes of the Vaginal Microbiota in HPV Infection and Cervical Intraepithelial Neoplasia: A Cross-Sectional Analysis. Sci. Rep. 2022, 12, 2812. [Google Scholar] [CrossRef]
- Nguyen, H.D.T.; Le, T.M.; Lee, E.; Lee, D.; Choi, Y.; Cho, J.; Park, N.J.Y.; Chong, G.O.; Seo, I.; Han, H.S. Relationship between Human Papillomavirus Status and the Cervicovaginal Microbiome in Cervical Cancer. Microorganisms 2023, 11, 1417. [Google Scholar] [CrossRef]
- Cascardi, E.; Cazzato, G.; Daniele, A.; Silvestris, E.; Cormio, G.; Di Vagno, G.; Malvasi, A.; Loizzi, V.; Scacco, S.; Pinto, V.; et al. Association between Cervical Microbiota and HPV: Could This Be the Key to Complete Cervical Cancer Eradication? Biology 2022, 11, 1114. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.S.; Morton, A.N.; Hocking, J.; Garland, S.M.; Morris, M.B.; Moss, L.M.; Horvath, L.B.; Kuzevska, I.; Fairley, C.K. High Recurrence Rates of Bacterial Vaginosis over the Course of 12 Months after Oral Metronidazole Therapy and Factors Associated with Recurrence. J. Infect. Dis. 2006, 193, 1478–1486. [Google Scholar] [CrossRef]
- Lagenaur, L.A.; Hemmerling, A.; Chiu, C.; Miller, S.; Lee, P.P.; Cohen, C.R.; Parks, T.P. Connecting the Dots: Translating the Vaginal Microbiome Into a Drug. J. Infect. Dis. 2021, 223, S296–S306. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, X.; Wu, F.; Chen, J.; Luo, J.; Wu, C.; Chen, T. Effectiveness of Vaginal Probiotics Lactobacillus Crispatus Chen-01 in Women with High-Risk HPV Infection: A Prospective Controlled Pilot Study. Aging 2024, 16, 11446–11459. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.C.; Fu, H.C.; Tseng, C.W.; Wu, C.H.; Tsai, C.C.; Lin, H. The Influence of Probiotics on Genital High-Risk Human Papilloma Virus Clearance and Quality of Cervical Smear: A Randomized Placebo-Controlled Trial. BMC Women’s Health 2019, 19, 103. [Google Scholar] [CrossRef] [PubMed]
- Dellino, M.; Cascardi, E.; Laganà, A.S.; Di Vagno, G.; Malvasi, A.; Zaccaro, R.; Maggipinto, K.; Cazzato, G.; Scacco, S.; Tinelli, R.; et al. Lactobacillus Crispatus M247 Oral Administration: Is It Really an Effective Strategy in the Management of Papillomavirus-Infected Women? Infect. Agents Cancer 2022, 17, 53. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, S.K.; Falkenthal, T.E.H.; Sundström, K.; Munk, C.; Sture, T.; Bautista, O.; Group, T.; Rawat, S.; Luxembourg, A. Long-Term Effectiveness of the Nine-Valent Human Papillomavirus Vaccine: Interim Results after 12 Years of Follow-up in Scandinavian Women. Hum. Vaccines Immunother. 2024, 20, 2377903. [Google Scholar] [CrossRef]
- Restrepo, J.; Herrera, T.; Samakoses, R.; Reina, J.C.; Pitisuttithum, P.; Ulied, A.; Bekker, L.G.; Moreira, E.D.; Olsson, S.E.; Block, S.L.; et al. Ten-Year Follow-up of 9-Valent Human Papillomavirus Vaccine: Immunogenicity, Effectiveness, and Safety. Pediatrics 2023, 152, e2022060993. [Google Scholar] [CrossRef]
- Ellingson, M.K.; Sheikha, H.; Nyhan, K.; Oliveira, C.R.; Niccolai, L.M. Human Papillomavirus Vaccine Effectiveness by Age at Vaccination: A Systematic Review. Hum. Vaccines Immunother. 2023, 19, 2239085. [Google Scholar] [CrossRef]
- Meites, E.; Szilagyi, P.G.; Chesson, H.W.; Unger, E.R.; Romero, J.R.; Markowitz, L.E. Human Papillomavirus Vaccination for Adults: Updated Recommendations of the Advisory Committee on Immunization Practices. MMWR. Morb. Mortal. Wkly. Rep. 2019, 68, 698–702. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Human papillomavirus vaccines: WHO position paper, May 2017—Recommendations. Vaccine 2017, 35, 5753–5755. [Google Scholar] [CrossRef]
- Giuliano, A.R.; Joura, E.A.; Garland, S.M.; Huh, W.K.; Iversen, O.E.; Kjaer, S.K.; Ferenczy, A.; Kurman, R.J.; Ronnett, B.M.; Stoler, M.H.; et al. Nine-Valent HPV Vaccine Efficacy against Related Diseases and Definitive Therapy: Comparison with Historic Placebo Population. Gynecol. Oncol. 2019, 154, 110–117. [Google Scholar] [CrossRef]
- Whitworth, H.S.; Mounier-Jack, S.; Choi, E.M.; Gallagher, K.E.; Howard, N.; Kelly, H.; Mbwanji, G.; Kreimer, A.R.; Basu, P.; Barnabas, R.; et al. Efficacy and Immunogenicity of a Single Dose of Human Papillomavirus Vaccine Compared to Multidose Vaccination Regimens or No Vaccination: An Updated Systematic Review of Evidence from Clinical Trials. Vaccine X 2024, 19, 100486. [Google Scholar] [CrossRef]
- Giraldo, P.C.; Sanches, J.M.; Sparvolli, L.G.; Amaral, R.; Migliorini, I.; Gil, C.D.; Taddei, C.R.; Witkin, S.S.; Discacciati, M.G. Relationship between Papillomavirus Vaccine, Vaginal Microbiome, and Local Cytokine Response: An Exploratory Research. Braz. J. Microbiol. 2021, 52, 2363–2371. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Nakagawa, M.; Coleman, H.N.; Owens, S.M.; Greenfield, W.W.; Sasagawa, T.; Robeson, M.S. Evaluation of DNA Extraction Protocols from Liquid-Based Cytology Specimens for Studying Cervical Microbiota. PLoS ONE 2021, 16, e0237556. [Google Scholar] [CrossRef] [PubMed]
- Usyk, M.; Zolnik, C.P.; Castle, P.E.; Porras, C.; Herrero, R.; Gradissimo, A.; Gonzalez, P.; Safaeian, M.; Schiffman, M.; Burk, R.D. Cervicovaginal Microbiome and Natural History of HPV in a Longitudinal Study. PLoS Pathog. 2020, 16, e1008376. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Shui, Y.; Qian, Y. A Crosstalk Analysis of High-Risk Human Papillomavirus, Microbiota and Vaginal Metabolome in Cervicovaginal Microenvironment. Microb. Pathog. 2024, 194, 106826. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Published by MDPI on behalf of the Lithuanian University of Health Sciences. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazlauskaitė, J.; Žukienė, G.; Rudaitis, V.; Bartkevičienė, D. The Vaginal Microbiota, Human Papillomavirus, and Cervical Dysplasia—A Review. Medicina 2025, 61, 847. https://doi.org/10.3390/medicina61050847
Kazlauskaitė J, Žukienė G, Rudaitis V, Bartkevičienė D. The Vaginal Microbiota, Human Papillomavirus, and Cervical Dysplasia—A Review. Medicina. 2025; 61(5):847. https://doi.org/10.3390/medicina61050847
Chicago/Turabian StyleKazlauskaitė, Justė, Guoda Žukienė, Vilius Rudaitis, and Daiva Bartkevičienė. 2025. "The Vaginal Microbiota, Human Papillomavirus, and Cervical Dysplasia—A Review" Medicina 61, no. 5: 847. https://doi.org/10.3390/medicina61050847
APA StyleKazlauskaitė, J., Žukienė, G., Rudaitis, V., & Bartkevičienė, D. (2025). The Vaginal Microbiota, Human Papillomavirus, and Cervical Dysplasia—A Review. Medicina, 61(5), 847. https://doi.org/10.3390/medicina61050847