
Gastrodin Alleviates Angiotensin II-Induced Hypertension and Myocardial Apoptosis via Inhibition of the PRDX2/p53 Pathway In Vivo and In Vitro

Abstract
1. Introduction
2. Results
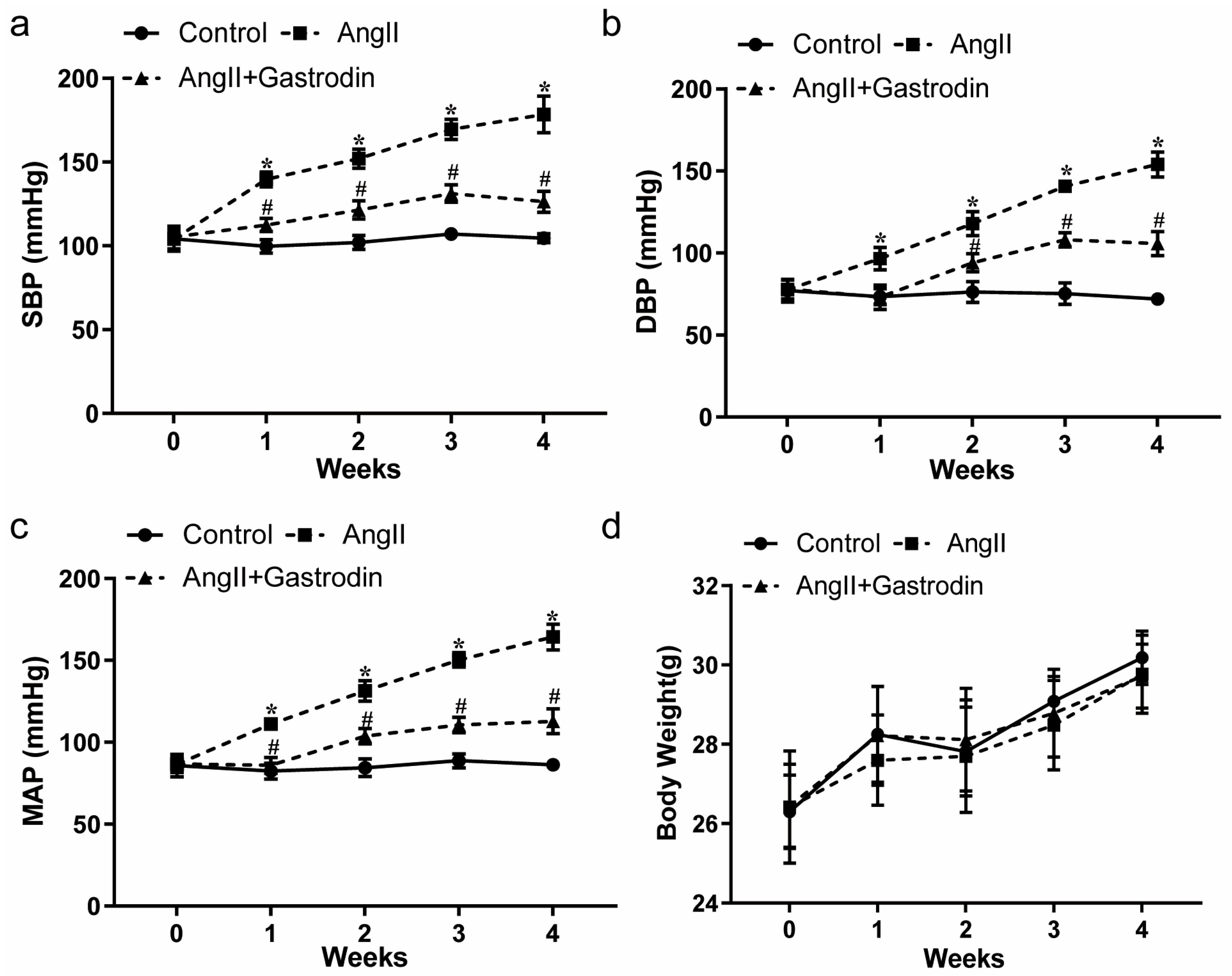
2.1. Gastrodin Reduces the Blood Pressure in Ang II-Induced Hypertensive Mice
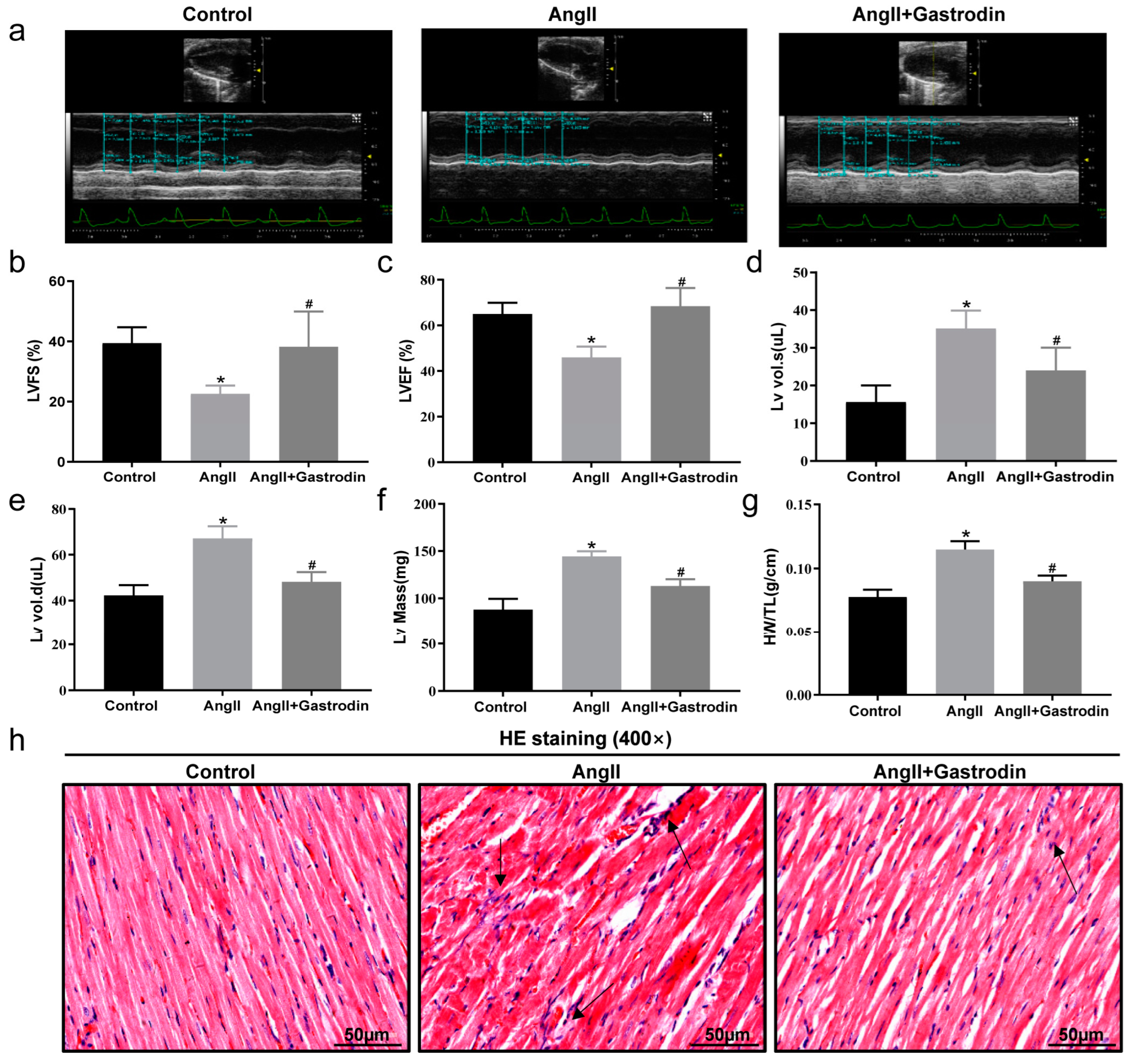
2.2. Gastrodin Attenuates Cardiac Dysfunction and Pathological Damage in Ang II-Induced Hypertensive Mice
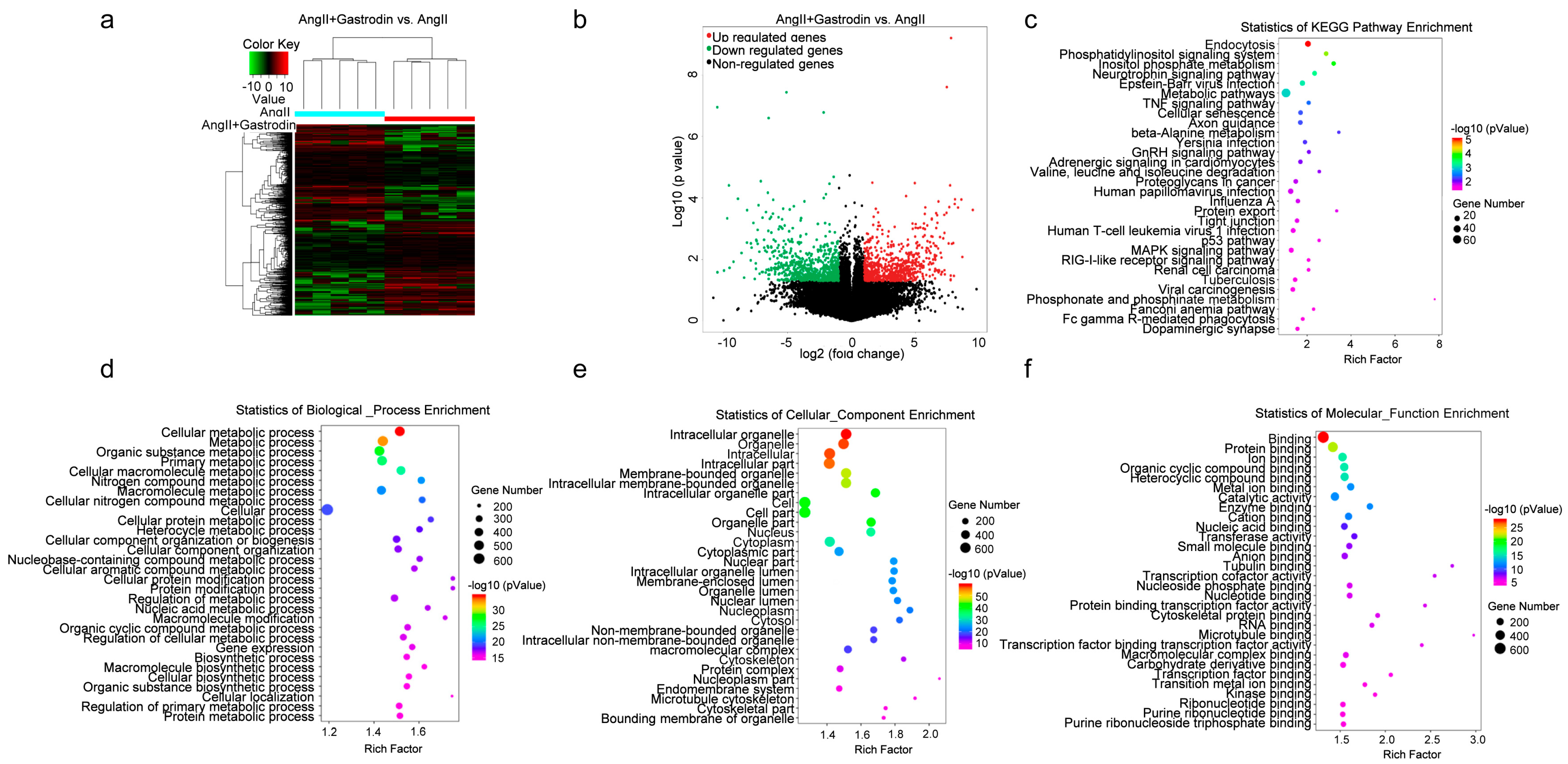
2.3. The Differentially Expressed Transcripts (DETs) and Pathway Enrichment in the Cardiac Tissues of Ang II-Induced Hypertensive Mice Following Gastrodin Treatment
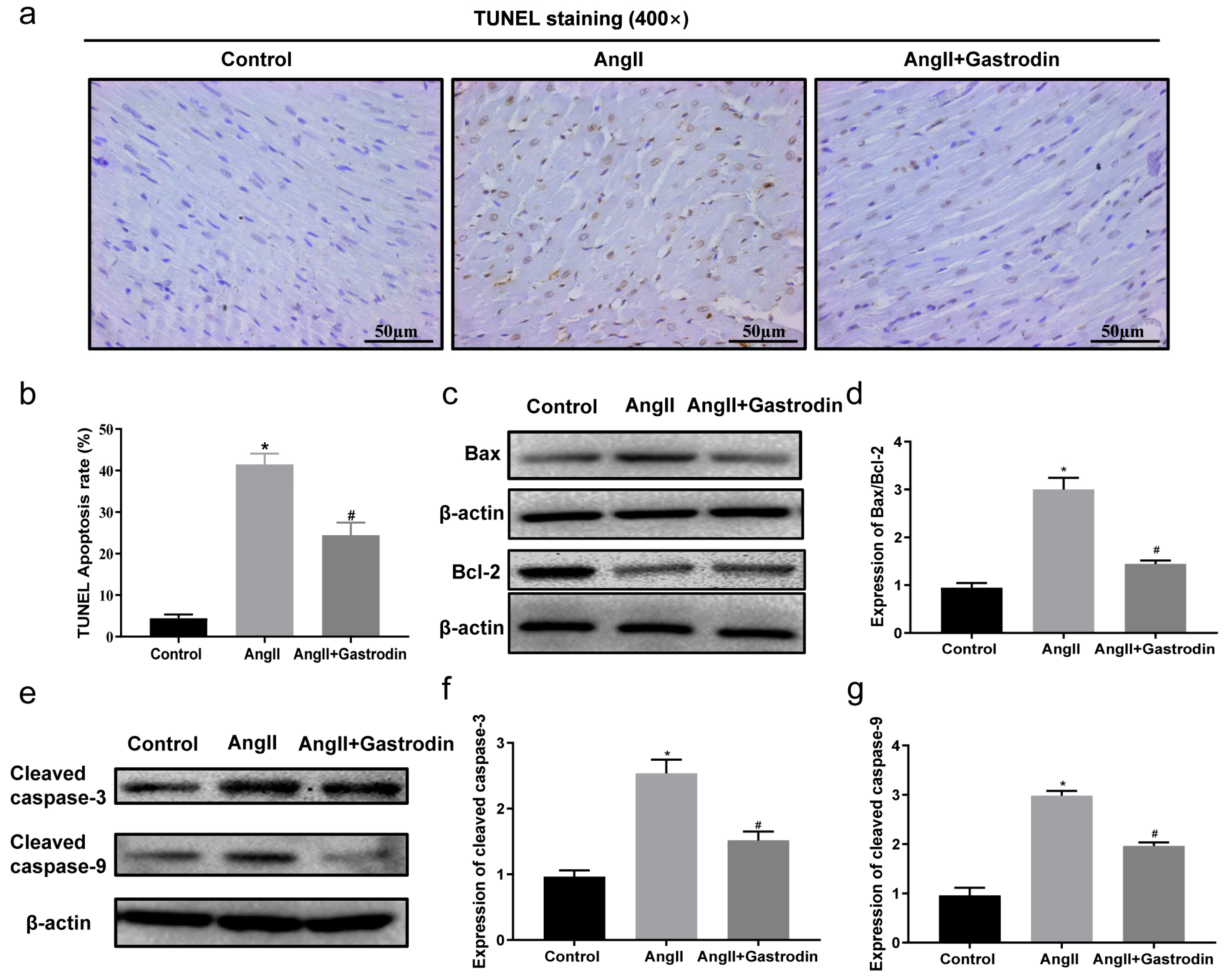
2.4. Gastrodin Reduces Ang II-Induced Myocardial Apoptosis In Vivo
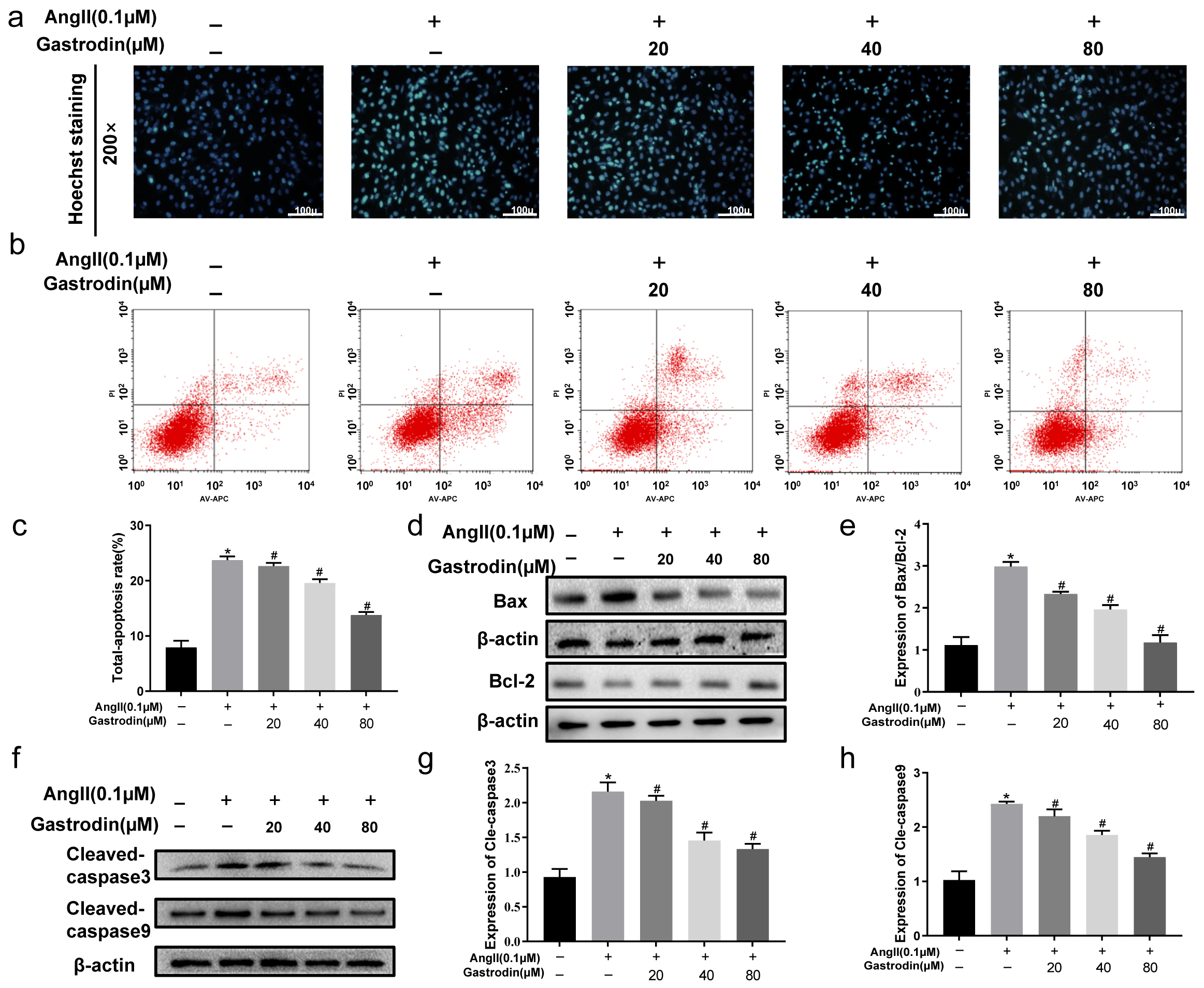
2.5. Gastrodin Attenuates Ang II-Induced Apoptosis in H9c2 Cells
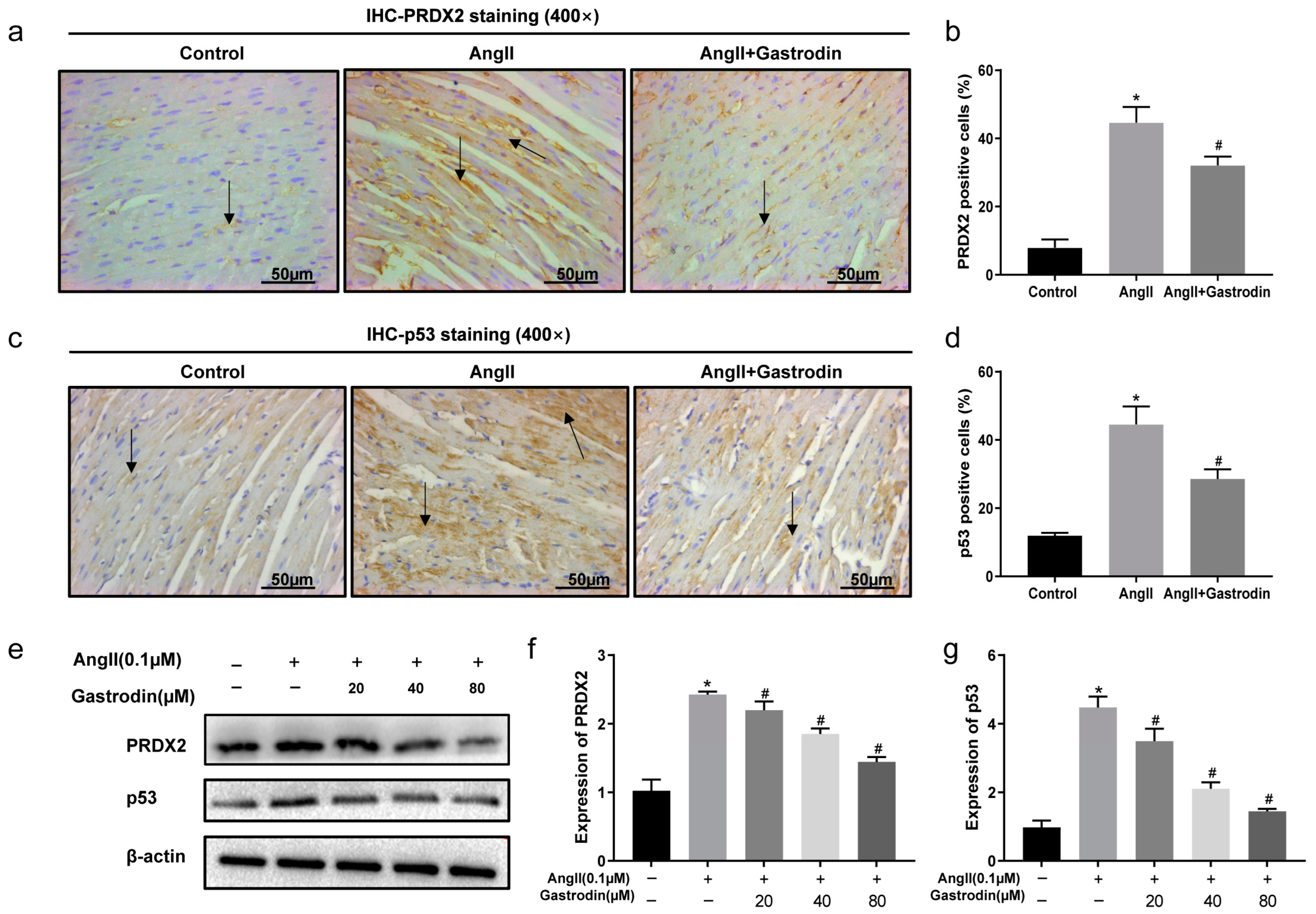
2.6. Gastrodin Inhibits PRDX2/p53 Pathway Activation in Ang II-Induced Hypertension In Vivo and In Vitro
3. Discussion
4. Materials and Methods
4.1. Reagents and Consumables
4.2. Animals and Experimental Protocols
4.3. Blood Pressure Measurement and Echocardiography
4.4. Determination of Heart Weight/Tibia Length Ratio and Hematoxylin and Eosin (H&E) Staining
4.5. Immunohistochemical (IHC) Analysis
4.6. TUNEL Staining
4.7. RNA Sequencing (RNA-Seq)
4.8. Cell Culture and Treatments
4.9. Hoechst Staining
4.10. Apoptosis Assay by Flow Cytometry
4.11. Western-Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Jarraya, F. Treatment of Hypertension: Which Goal for Which Patient? Adv. Exp. Med. Biol. 2017, 956, 117–127. [Google Scholar] [PubMed]
- Riaz, S.; Zeidan, A.; Mraiche, F. Myocardial proteases and cardiac remodeling. J. Cell Physiol. 2017, 232, 3244–3250. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pu, Z.; Xu, G.; Yang, Y.; Cui, Y.; Zhou, X.; Wang, C.; Zhong, Z.; Zhou, S.; Yin, J.; et al. Hypoxia-Induced Myocardial Hypertrophy Companies with Apoptosis Enhancement and p38-MAPK Pathway Activation. High Alt. Med. Biol. 2024, 25, 186–196. [Google Scholar] [CrossRef]
- Cheng, Y.; Yan, M.; He, S.; Xie, Y.; Wei, L.; Xuan, B.; Shang, Z.; Wu, M.; Zheng, H.; Chen, Y.; et al. Baicalin alleviates angiotensin II-induced cardiomyocyte apoptosis and autophagy and modulates the AMPK/mTOR pathway. J. Cell. Mol. Med. 2024, 28, e18321. [Google Scholar] [CrossRef]
- Wang, X.; Simpson, E.R.; Brown, K.A. p53: Protection against Tumor Growth beyond Effects on Cell Cycle and Apoptosis. Cancer Res. 2015, 75, 5001–5007. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Minamino, T.; Toko, H.; Miyauchi, H.; Orimo, M.; Qin, Y.; Akazawa, H.; Tateno, K.; Kayama, Y.; Harada, M. P53-induced Inhibition of Hif-1 Causes Cardiac Dysfunction during Pressure Overload. Nature 2007, 446, 444–448. [Google Scholar] [CrossRef]
- Yano, T.; Abe, K.; Tanno, M.; Miki, T.; Kuno, A.; Miura, T.; Steenbergen, C. Does p53 Inhibition Suppress Myocardial Ischemia-Reperfusion Injury? J. Cardiovasc. Pharm. Ther. 2018, 23, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.Y.; Hsu, C.Y.; He, J.R.; Liu, C.L.; Lo, S.J.; Chen, Y.C.; Huang, H.Y. Identifying apoptosis-evasion proteins/pathways in human hepatoma cells via induction of cellular hormesis by UV irradiation. J. Proteome Res. 2009, 8, 3977–3986. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Yan, S.; Li, Y.; Wu, L.; Zheng, Y.; Wang, Y.; Fang, Z. The effects of gastrodin injection on hypertension: A systematic review and meta-analysis. Medicine 2020, 99, e20936. [Google Scholar] [CrossRef]
- Alshalalfeh, M.; Sun, N.; Moraes, A.H.; Utani, A.P.A.; Xu, Y. Conformational Distributions of Phenyl β-D-Glucopyranoside and Gastrodin in Solution by Vibrational Optical Activity and Theoretical Calculations. Molecules 2023, 28, 4013. [Google Scholar] [CrossRef]
- Huang, B.; Lin, Z.; Chen, Z.; Chen, J.; Shi, B.; Jia, J.; Li, Y.; Pan, Y.; Liang, Y.; Cai, Z. Strain differences in the drug transport capacity of intestinal glucose transporters in Sprague-Dawley versus Wistar rats, C57BL/6J versus Kunming mice. Int. J. Pharm. 2023, 640, 123000. [Google Scholar] [CrossRef]
- Zhao, Y.; Gong, X.J.; Zhou, X.; Kang, Z.J. Relative bioavailability of gastrodin and parishin from extract and powder of Gastrodiae rhizoma in rat. J. Pharm. Biomed. Anal. 2014, 100, 309–315. [Google Scholar] [CrossRef]
- Lv, Y.; Cao, H.; Chu, L.; Peng, H.; Shen, X.; Yang, H. Effects of Gastrodin on BV2 cells under oxygen-glucose deprivation and its mechanism. Gene 2021, 766, 145152. [Google Scholar] [CrossRef]
- Hu, Y.; Li, C.; Shen, W. Gastrodin alleviates memory deficits and reduces neuropathology in a mouse model of Alzheimer’s disease. Neuropathology 2014, 34, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Shi, H.; Liu, K.; Zhong, L.; Wang, F.; You, Q. Protective effect of gastrodin on myocardial ischemia-reperfusion injury and the expression of Bax and Bcl-2. Exp. Ther. Med. 2019, 17, 4389–4394. [Google Scholar] [CrossRef]
- Fu, S.; Chen, L.; Wu, Y.; Tang, Y.; Tang, L.; Zhong, Y.; Wang, S.; Liu, H.; Wang, X.; Chen, A. Gastrodin pretreatment alleviates myocardial ischemia/reperfusion injury through promoting autophagic flux. Biochem. Biophys. Res. Commun. 2018, 503, 2421–2428. [Google Scholar] [CrossRef]
- Zhang, M.; Tan, Y.; Song, Y.; Zhu, M.; Zhang, B.; Chen, C.; Wang, S.; Liu, H.; Wang, X.; Chen, A. GLUT4 mediates the protective function of gastrodin against pressure overload-induced cardiac hypertrophy. Biomed. Pharmacother. 2023, 161, 114324. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Lo, C.Y.; Meng, Z.; Li, Z.; Zhong, M.; Zhang, P.; Lu, J.; Yang, Z.; Yan, F.; Zhang, Y.; et al. Gastrodin Inhibits Store-Operated Ca2+ Entry and Alleviates Cardiac Hypertrophy. Front. Pharmacol. 2017, 8, 222. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Y.; Hu, X.; Bian, X.; Nian, S. Gastrodin prevents homocysteine-induced human umbilical vein endothelial cells injury via PI3K/Akt/eNOS and Nrf2/ARE pathway. J. Cell. Mol. Med. 2021, 25, 345–357. [Google Scholar] [CrossRef]
- Chen, J.; Gu, Y.T.; Xie, J.J.; Wu, C.C.; Xuan, J.; Guo, W.J.; Yan, Y.Z.; Chen, L.; Wu, Y.S.; Zhang, X.L.; et al. Gastrodin reduces IL-1β-induced apoptosis, inflammation, and matrix catabolism in osteoarthritis chondrocytes and attenuates rat cartilage degeneration in vivo. Biomed. Pharmacother. 2018, 97, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.H.; Huang, C.M.; Gao, X.; Wang, J.W.; Hao, L.L.; Ji, Q. Gastrodin inhibits high glucose-induced human retinal endothelial cell apoptosis by regulating the SIRT1/TLR4/NF-κBp65 signaling pathway. Mol. Med. Rep. 2018, 17, 7774–7780. [Google Scholar] [CrossRef]
- Cheng, Q.-Q.; Wan, Y.-W.; Yang, W.-M.; Tian, M.-H.; Wang, Y.-C.; He, H.-Y.; Zhang, W.D.; Liu, X. Gastrodin protects H9c2 cardiomyocytes against oxidative injury by ameliorating imbalanced mitochondrial dynamics and mitochondrial dysfunction. Acta Pharmacol. Sin. 2020, 41, 1314–1327. [Google Scholar] [CrossRef]
- Wang, W.; Wei, J.; Zhang, H.; Zheng, X.; Zhou, H.; Luo, Y.; Yang, J.; Deng, Q.; Huang, S.; Fu, Z. PRDX2 promotes the proliferation of colorectal cancer cells by increasing the ubiquitinated degradation of p53. Cell Death Dis. 2021, 12, 605. [Google Scholar] [CrossRef]
- Wu, F.; Tian, F.; Zeng, W.; Liu, X.; Fan, J.; Lin, Y.; Zhang, Y. Role of peroxiredoxin2 downregulation in recurrent miscarriage through regulation of trophoblast proliferation and apoptosis. Cell Death Dis. 2017, 8, e2908. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.H.; Fu, Q.W.; Li, L.X.; Zhou, R.; Liu, N.; Peng, J.H.; Chen, Y. Prx II reduces oxidative stress and cell senescence in chondrocytes by activating the p16-CDK4/6-pRb-E2F signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3448–3458. [Google Scholar] [PubMed]
- Huang, Z.; Chen, Z.; Wang, X.; Ding, X.; Cai, Z.; Li, W.; Cai, Z.; Lan, Y.; Chen, G.; Fang, W.; et al. Association of Cardiovascular Health Score Trajectory With Incident Myocardial Infarction in Hypertensive Patients. Hypertension 2022, 79, 2622–2630. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Tang, R.; Yang, N.; Chen, Y. Review on pharmacological effects of gastrodin. Arch. Pharm. Res. 2023, 46, 744–770. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Lu, B.; Qi, W.; Jiao, F.; Zhang, H.; Yuan, D. Metabolite profiling and quantification of phytochemicals of Tianma-Gouteng granule in human and rat urine using ultra high performance liquid chromatography coupled with tandem mass spectrometry. J. Sep. Sci. 2019, 42, 2762–2770. [Google Scholar] [CrossRef]
- Zhang, H.; Duan, S.; Wang, L.; Liu, J.; Qi, W.; Yuan, D. Identification of the absorbed components and their metabolites of Tianma-Gouteng granule in rat plasma and bile using ultra-high-performance liquid chromatography combined with quadrupole time-of-flight mass spectrometry. Biomed. Chromatogr. 2019, 33, e4480. [Google Scholar] [CrossRef]
- Li, Y.; Li, F. Mechanism and Prospect of Gastrodin in Osteoporosis, Bone Regeneration, and Osseointegration. Pharmaceuticals 2022, 15, 1432. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Dong, Z.; Lu, X.Q.; Xia, Y.P.; Zhu, S.B. Analysis on 315 cases of clinical adverse drug reaction/event induced by gastrodin. Zhongguo Zhong Yao Za Zhi 2015, 40, 2037–2041. [Google Scholar] [PubMed]
- Guo, Z.; Yang, X.; Wu, M.; Shen, A.; Li, J.; Zhang, X.; Cheng, Y.; Xie, Q.; Peng, J. Gastrodin attenuates angiotensin II-induced vascular contraction and MLCK/p-MLC(2) pathway activation. Pharm. Biol. 2023, 61, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhang, X.; Wei, L.; Wu, M.; Cheng, Y.; Zheng, H.; Shen, A.; Fu, C.; Ali, F.; Long, L.; et al. Gastrodin attenuates renal injury and collagen deposition via suppression of the TGF-β1/Smad2/3 signaling pathway based on network pharmacology analysis. Front. Pharmacol. 2023, 14, 1082281. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Wu, M.; Ali, F.; Guo, Z.; Fang, Y.; Zhou, Y.; Zhang, S.; Zhang, W.; Wen, Y.; Yu, M.; et al. Based on network pharmacology, gastrodin attenuates hypertension-induced vascular smooth muscle cell proliferation and PI3K/AKT pathway activation. Sci. Rep. 2023, 13, 12140. [Google Scholar] [CrossRef]
- Liu, W.; Su, B.L.; Wang, Z.S.; Zhang, X.; Gao, Y.S.; Song, S.W. Gastrodin improved baroreflex sensitivity and increased gamma-amino butyric acid content in brains without decreasing blood pressure in spontaneously hypertensive rats. CNS Neurosci. Ther. 2012, 18, 873–875. [Google Scholar] [CrossRef]
- Zhu, M.; Deng, W.; Di, S.; Qin, M.; Liu, D.; Yi, B. Gastrodin Protects Cardiomyocytes from Anoxia/Reoxygenation Injury by 14-3-3η. Oxid. Med. Cell. Longev. 2018, 2018, 3685391. [Google Scholar] [CrossRef]
- Shu, C.; Chen, C.; Zhang, D.P.; Guo, H.; Zhou, H.; Zong, J.; Bian, Z.; Dong, X.; Dai, J.; Zhang, Y.; et al. Gastrodin protects against cardiac hypertrophy and fibrosis. Mol. Cell. Biochem. 2012, 359, 9–16. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Härtlein, I.; Smith, S.M.E.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Liu, S.J.; Liu, X.Y.; Li, J.H.; Guo, J.; Li, F.; Gui, Y.; Li, X.H.; Yang, L.; Wu, C.Y.; Yuan, Y.; et al. Gastrodin attenuates microglia activation through renin-angiotensin system and Sirtuin3 pathway. Neurochem. Int. 2018, 120, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Jalmi, S.K.; Sinha, A.K. ROS mediated MAPK signaling in abiotic and biotic stress- striking similarities and differences. Front. Plant Sci. 2015, 6, 769. [Google Scholar] [CrossRef]
- Xu, J.M.; Tan, R.; Hu, D.X. Effect of ischemic preconditioning on myocardial bcl-2, bax, p53 gene expression during ischemia/reperfusion period in rabbits. Hunan Yi Ke Da Xue Xue Bao 2003, 28, 111–113. [Google Scholar] [PubMed]
- Kai, H.; Kudo, H.; Takayama, N.; Yasuoka, S.; Kajimoto, H.; Imaizumi, T. Large blood pressure variability and hypertensive cardiac remodeling—Role of cardiac inflammation. Circ. J. 2009, 73, 2198–2203. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.R.; Chung, A.C.; Yang, F.; Yue, W.; Deng, C.; Lau, C.P.; Tse, H.F.; Lan, H.Y. Smad3 mediates cardiac inflammation and fibrosis in angiotensin II-induced hypertensive cardiac remodeling. Hypertension 2010, 55, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Shen, A.; Wu, X.; Shen, Z.; Chen, X.; Li, J.; Liu, L.; Lin, X.; Wu, M.; Chen, Y.; et al. Qingda granule attenuates angiotensin II-induced cardiac hypertrophy and apoptosis and modulates the PI3K/AKT pathway. Biomed. Pharmacother. 2021, 133, 111022. [Google Scholar] [CrossRef]
- González, A.; Fortuño, M.A.; Querejeta, R.; Ravassa, S.; López, B.; López, N.; Díez, J. Cardiomyocyte apoptosis in hypertensive cardiomyopathy. Cardiovasc. Res. 2003, 59, 549–562. [Google Scholar] [CrossRef]
- Shao, F.; Zhou, L.; Zhang, Y.; Chen, H.; Zhang, Y.; Guan, Z. Gastrodin alleviates inflammatory injury of cardiomyocytes in septic shock mice via inhibiting NLRP3 expression. In Vitro Cell. Dev. Biol. Anim. 2021, 57, 571–581. [Google Scholar] [CrossRef]
- Chunhacha, P.; Pinkaew, D.; Sinthujaroen, P.; Bowles, D.E.; Fujise, K. Fortilin inhibits p53, halts cardiomyocyte apoptosis, and protects the heart against heart failure. Cell Death Discov. 2021, 7, 310. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, K.; Zheng, Q.; Zhang, C.; Tang, H.; Babicheva, A.; Jiang, Q.; Li, M.; Chen, Y.; Carr, S.; et al. Divergent changes of p53 in pulmonary arterial endothelial and smooth muscle cells involved in the development of pulmonary hypertension. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2019, 316, L216–L228. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Li, Y.; Li, R.; Zhang, A.A.; Shang, B.; Yu, J.; Xie, X. Tetrahydrobiopterin Protects against Radiation-induced Growth Inhibition in H9c2 Cardiomyocytes. Chin. Med. J. 2016, 129, 2733–2740. [Google Scholar] [CrossRef]
- Shan, R.; Zhang, Y.; Shi, Y.; Wang, X.; Wang, X.; Ma, G.; Li, Q. Activation of Cannabinoid Type 2 Receptor in Microglia Reduces Neuroinflammation through Inhibiting Aerobic Glycolysis to Relieve Hypertension. Biomolecules 2024, 14, 333. [Google Scholar] [CrossRef]
- Wu, F.; Zuo, H.J.; Ren, X.Q.; Wang, P.X.; Li, F.; Li, J.J. Gastrodin Regulates the Notch-1 Signal Pathway via Renin-Angiotensin System in Activated Microglia. NeuroMol. Med. 2023, 25, 40–52. [Google Scholar] [CrossRef]
- Wang, X.; Lin, L.; Lan, B.; Wang, Y.; Du, L.; Chen, X.; Li, Q.; Liu, K.; Hu, M.; Xue, Y.; et al. IGF2R-initiated proton rechanneling dictates an anti-inflammatory property in macrophages. Sci. Adv. 2020, 6, eabb7389. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Ma, X.; Gao, W.C.; Zhang, X.; Fu, Y.; Liu, Q.; Tian, L.; Qin, X.; Yang, W.; Zheng, H.; et al. Gastrodin Exerts Cardioprotective Action via Inhibition of Insulin-Like Growth Factor Type 2/Insulin-Like Growth Factor Type 2 Receptor Expression in Cardiac Hypertrophy. ACS Omega 2021, 6, 16763–16774. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Xing, G.H.; Hong, B.; Li, X.M.; Zou, Y.; Zhang, X.J.; Dong, M.X. Gastrodin prevents motor deficits and oxidative stress in the MPTP mouse model of Parkinson’s disease: Involvement of ERK1/2-Nrf2 signaling pathway. Life Sci. 2014, 114, 77–85. [Google Scholar] [CrossRef]
- Lin, J.; Shi, Y.; Miao, J.; Wu, Y.; Lin, H.; Wu, J.; Zeng, W.; Qi, F.; Liu, C.; Wang, X.; et al. Gastrodin Alleviates Oxidative Stress-Induced Apoptosis and Cellular Dysfunction in Human Umbilical Vein Endothelial Cells via the Nuclear Factor-Erythroid 2-Related Factor 2/Heme Oxygenase-1 Pathway and Accelerates Wound Healing In Vivo. Front. Pharmacol. 2019, 10, 1273. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.J.; Zhou, Y.J.; Xiong, X.M.; Li, N.S.; Zhang, J.J.; Luo, X.J.; Peng, J. Ubiquitin-specific protease 7 promotes ferroptosis via activation of the p53/TfR1 pathway in the rat hearts after ischemia/reperfusion. Free Radic. Biol. Med. 2021, 162, 339–352. [Google Scholar] [CrossRef]
- Xing, P.; Li, X.; Bai, Y.; Jiao, Z. Cypermethrin and/or sulfamethoxazole exposure effect on apoptosis and endoplasmic reticulum of grass carp cardiomyocyte. Ecotoxicol. Environ. Saf. 2023, 252, 114594. [Google Scholar] [CrossRef]
- Shen, Z.; Shen, A.; Chen, X.; Wu, X.; Chu, J.; Cheng, Y.; Peng, M.; Chen, Y.; Weygant, N.; Wu, M.; et al. Huoxin pill attenuates myocardial infarction-induced apoptosis and fibrosis via suppression of p53 and TGF-β1/Smad2/3 pathways. Biomed. Pharmacother. 2020, 130, 110618. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Z.; Jia, J.; Chen, A.; Gao, Y.; Qian, J.; Ge, J. Rosuvastatin Alleviates Coronary Microembolization-Induced Cardiac Injury by Suppressing Nox2-Induced ROS Overproduction and Myocardial Apoptosis. Cardiovasc. Toxicol. 2022, 22, 341–351. [Google Scholar] [CrossRef]
- Jiang, G.; Wu, H.; Hu, Y.; Li, J.; Li, Q. Gastrodin inhibits glutamate-induced apoptosis of PC12 cells via inhibition of CaMKII/ASK-1/p38 MAPK/p53 signaling cascade. Cell. Mol. Neurobiol. 2014, 34, 591–602. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Y.; Zhang, B.; Zheng, Y.; Ji, H.; Li, S. The improvement effect of gastrodin on LPS/GalN-induced fulminant hepatitis via inhibiting inflammation and apoptosis and restoring autophagy. Int. Immunopharmacol. 2020, 85, 106627. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Tandon, S.; Tandon, C. An update on vascular calcification and potential therapeutics. Mol. Biol. Rep. 2021, 48, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Huang, Y.; Wang, Q.; Wang, C.; Hu, H.; Zhang, H.; Lu, D.; Jiang, H.; Shen, R.; Zhang, W.; et al. Ainsliadimer A induces ROS-mediated apoptosis in colorectal cancer cells via directly targeting peroxiredoxin 1 and 2. Cell Chem. Biol. 2023, 30, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, C.; Li, D.; Su, Q.; Hang, Y.; Zhang, P.; Hu, W. PRDX2 in Myocyte Hypertrophy and Survival is Mediated by TLR4 in Acute Infarcted Myocardium. Sci. Rep. 2017, 7, 6970. [Google Scholar] [CrossRef]
- Li, J.; Wang, C.; Wang, W.; Liu, L.; Zhang, Q.; Zhang, J.; Wang, B.; Wang, S.; Li, H.; Gao, C.; et al. PRDX2 Protects Against Atherosclerosis by Regulating the Phenotype and Function of the Vascular Smooth Muscle Cell. Front. Cardiovasc. Med. 2021, 8, 624796. [Google Scholar] [CrossRef]
- Li, Y.; Ji, Y.; Li, F. A review: Mechanism and prospect of gastrodin in prevention and treatment of T2DM and COVID-19. Heliyon 2023, 9, e21218. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, M.; Lv, T.; Huang, M.; Cheng, B.; Zhang, Y.; Zhu, J. Gastrodin Inhibits Virus Infection by Promoting the Production of Type I Interferon. Front. Pharmacol. 2020, 11, 608707. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, S.; Zhang, J.; Wang, Y.; Wang, Y.; Zhang, L.; Zhang, L.; Li, L.; Dong, J.; Wang, B. Gastrodin, a traditional Chinese medicine monomer compound, can be used as adjuvant to enhance the immunogenicity of melanoma vaccines. Int. Immunopharmacol. 2019, 74, 105699. [Google Scholar] [CrossRef]
- Shu, G.; Yang, T.; Wang, C.; Su, H.; Xiang, M. Gastrodin stimulates anticancer immune response and represses transplanted H22 hepatic ascitic tumor cell growth: Involvement of NF-κB signaling activation in CD4+ T cells. Toxicol. Appl. Pharmacol. 2013, 269, 270–279. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Gui, L.; Deng, Y.; Wang, L.; Jiao, J.; Hu, Y.; Lan, X.; Hou, J.; Li, Y.; et al. Sequential gastrodin release PU/n-HA composite scaffolds reprogram macrophages for improved osteogenesis and angiogenesis. Bioact. Mater. 2023, 19, 24–37. [Google Scholar] [CrossRef]
- Wei, L.; Wang, L.; Yu, J.; Fordjour, A.P.; Zhao, Y.Q. Gastrodin Reduces Blood Pressure by Intervening with RAAS and PPARγ in SHRs. Evid.-Based Complement. Altern. Med. 2015, 2015, 132–1030. [Google Scholar]
- Yang, H.; Li, Q.; Li, L.; Chen, S.; Zhao, Y.; Hu, Y.; Wang, L.; Lan, X.; Zhong, L.; Lu, D. Gastrodin modified polyurethane conduit promotes nerve repair via optimizing Schwann cells function. Bioact. Mater. 2022, 8, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Yongguang, L.; Xiaowei, W.; Huichao, Y.; Yanxiang, Z. Gastrodin promotes the regeneration of peripheral nerves by regulating miR-497/BDNF axis. BMC Complement. Med. 2022, 22, 45. [Google Scholar] [CrossRef]
- Liu, C.M.; Tian, Z.K.; Zhang, Y.J.; Ming, Q.L.; Ma, J.Q.; Ji, L.P. Effects of Gastrodin against Lead-Induced Brain Injury in Mice Associated with the Wnt/Nrf2 Pathway. Nutrients 2020, 12, 1805. [Google Scholar] [CrossRef]
- Ye, T.; Meng, X.; Wang, R.; Zhang, C.; He, S.; Sun, G.; Sun, X. Gastrodin Alleviates Cognitive Dysfunction and Depressive-Like Behaviors by Inhibiting ER Stress and NLRP3 Inflammasome Activation in db/db Mice. Int. J. Mol. Sci. 2018, 19, 3977. [Google Scholar] [CrossRef]
- Shu, J.; Qiu, G.; Mohammad, I. A Semi-Automatic Image Analysis Tool for Biomarker Detection in Immunohistochemistry Analysis. In Proceedings of the 2013 Seventh International Conference on Image and Graphics, Qingdao, China, 26–28 July 2013; IEEE: Piscataway, NJ, USA, 2013. [Google Scholar]
- Long, L.; Zhang, X.; Wen, Y.; Li, J.; Wei, L.; Cheng, Y.; Liu, H.; Chu, J.; Fang, Y.; Xie, Q.; et al. Qingda Granule Attenuates Angiotensin II-Induced Renal Apoptosis and Activation of the p53 Pathway. Front. Pharmacol. 2021, 12, 770863. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; White, E. p53-dependent apoptosis pathways. Adv. Cancer Res. 2001, 82, 55–84. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mean ± SD | Median | IQRs 1 (25%) | IQRs (75%) |
|---|---|---|---|---|
| Control | 104.55 ± 2.69 | 104.83 | 103.79 | 99.91 |
| Ang II | 178.47 ± 10.96 | 174.54 | 172.60 | 169.09 |
| Ang II + gastrodin | 126.34 ± 6.31 | 125.44 | 123.28 | 117.89 |
| Group | Mean ± SD | Median | IQRs 1 (25%) | IQRs (75%) |
|---|---|---|---|---|
| Control | 71.93 ± 2.68 | 72.37 | 71.27 | 67.18 |
| Ang II | 154.04 ± 7.60 | 150.57 | 149.24 | 147.93 |
| Ang II + gastrodin | 105.72 ± 7.34 | 104.85 | 99.47 | 98.00 |
| Group | Mean ± SD | Median | IQRs 1 (25%) | IQRs (75%) |
|---|---|---|---|---|
| Control | 86.20 ± 1.74 | 86.68 | 85.46 | 83.27 |
| Ang II | 164.21 ± 7.87 | 161.24 | 158.85 | 158.13 |
| Ang II + gastrodin | 112.76 ± 7.59 | 111.90 | 106.89 | 104.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, N.; Xie, Q.; Chen, Y.; Li, J.; Zhang, X.; Zheng, H.; Cheng, Y.; Wu, M.; Shen, A.; Wei, L.; et al. Gastrodin Alleviates Angiotensin II-Induced Hypertension and Myocardial Apoptosis via Inhibition of the PRDX2/p53 Pathway In Vivo and In Vitro. Pharmaceuticals 2024, 17, 1200. https://doi.org/10.3390/ph17091200
Xu N, Xie Q, Chen Y, Li J, Zhang X, Zheng H, Cheng Y, Wu M, Shen A, Wei L, et al. Gastrodin Alleviates Angiotensin II-Induced Hypertension and Myocardial Apoptosis via Inhibition of the PRDX2/p53 Pathway In Vivo and In Vitro. Pharmaceuticals. 2024; 17(9):1200. https://doi.org/10.3390/ph17091200
Chicago/Turabian StyleXu, Nanhui, Qiurong Xie, Youqin Chen, Jiapeng Li, Xiuli Zhang, Huifang Zheng, Ying Cheng, Meizhu Wu, Aling Shen, Lihui Wei, and et al. 2024. "Gastrodin Alleviates Angiotensin II-Induced Hypertension and Myocardial Apoptosis via Inhibition of the PRDX2/p53 Pathway In Vivo and In Vitro" Pharmaceuticals 17, no. 9: 1200. https://doi.org/10.3390/ph17091200
APA StyleXu, N., Xie, Q., Chen, Y., Li, J., Zhang, X., Zheng, H., Cheng, Y., Wu, M., Shen, A., Wei, L., Yao, M., Yang, Y., Sferra, T. J., Jafri, A., Fang, Y., & Peng, J. (2024). Gastrodin Alleviates Angiotensin II-Induced Hypertension and Myocardial Apoptosis via Inhibition of the PRDX2/p53 Pathway In Vivo and In Vitro. Pharmaceuticals, 17(9), 1200. https://doi.org/10.3390/ph17091200