Regioselective Hydroxylation of Phloretin, a Bioactive Compound from Apples, by Human Cytochrome P450 Enzymes

 , , and
, , and
Abstract
1. Introduction
2. Results
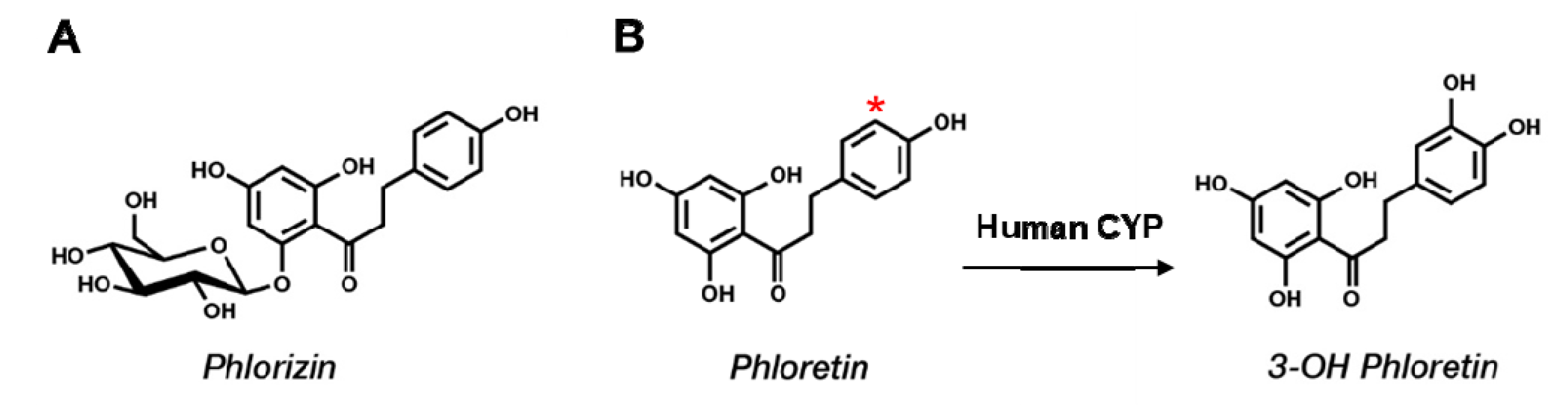
2.1. Phloretin Metabolism by Human Liver Microsomes and Identification of the Major Metabolite
2.2. Kinetics Parameters and Total Turnover Numbers (TTNs) of Phloretin Hydroxylation via HLMs and Recombinant Human CYPs
2.3. Inhibition of Phloretin Hydroxylation Activity by Antibodies in HLMs
2.4. Inhibitory Effect of Phloretin on Testosterone 6β-Hydroxylation Catalyzed by CYP3A4
2.5. Whole-Cell Biotransformation of Phloretin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Oxidation of Phloretin Catalyzed by Human Liver Microsomes
4.3. Phloretin’s Inhibitory Effect on Testosterone 6β-Hydroxylation Catalyzed via HLMs and CYP3A4
4.4. Immunoinhibition of Antibodies on Phloretin 3-Hydroxylation
4.5. LC-Mass Spectrometric Analysis of a Phloretin Metabolite
4.6. Optimal Expression of CYP3A4 in E. coli
4.7. Whole-Cell Biotransformations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Takeno, A.; Kanazawa, I.; Tanaka, K.; Notsu, M.; Sugimoto, T. Phloretin Suppresses Bone Morphogenetic Protein-2-Induced Osteoblastogenesis and Mineralization via Inhibition of Phosphatidylinositol 3-kinases/Akt Pathway. Int. J. Mol. Sci. 2019, 20, 2481. [Google Scholar] [CrossRef] [PubMed]
- Gosch, C.; Halbwirth, H.; Kuhn, J.; Miosic, S.; Stich, K. Biosynthesis of phloridzin in apple (Malus domestica Borkh.). Plant Sci. 2009, 176, 223–231. [Google Scholar] [CrossRef]
- Valenta, C.; Cladera, J.; O’Shea, P.; Hadgraft, J. Effect of phloretin on the percutaneous absorption of lignocaine across human skin. J. Pharm. Sci. 2001, 90, 485–492. [Google Scholar] [CrossRef]
- Nguyen, N.A.; Jang, J.; Le, T.-K.; Nguyen, T.H.H.; Woo, S.-M.; Yoo, S.-K.; Lee, Y.J.; Park, K.D.; Yeom, S.-J.; Kim, G.-J.; et al. Biocatalytic Production of a Potent Inhibitor of Adipocyte Differentiation from Phloretin Using Engineered CYP102A1. J. Agric. Food Chem. 2020, 68, 6683–6691. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef]
- Wilkinson, A.P.; Gee, J.M.; Dupont, M.S.; Needs, P.W.; Mellon, F.A.; Williamson, G.; Johnson, I.T. Hydrolysis by lactase phlorizin hydrolase is the first step in the uptake of daidzein glucosides by rat small intestine in vitro. Xenobiotica 2003, 33, 255–264. [Google Scholar] [CrossRef]
- Crespy, V.; Aprikian, O.; Morand, C.; Besson, C.; Manach, C.; Demigné, C.; Rémésy, C. Bioavailability of phloretin and phloridzin in rats. J. Nutr. 2001, 131, 3227–3230. [Google Scholar] [CrossRef]
- David-Silva, A.; Esteves, J.V.; Morais, M.R.P.T.; Freitas, H.S.; Zorn, T.M.; Correa-Giannella, M.L.; Machado, U.F. Dual SGLT1/SGLT2 Inhibitor Phlorizin Ameliorates Non-Alcoholic Fatty Liver Disease and Hepatic Glucose Production in Type 2 Diabetic Mice. Diabetes Metab. Syndr. Obes. 2020, 13, 739–751. [Google Scholar] [CrossRef]
- Benković, G.; Bojić, M.; Maleš, Ž.; Tomić, S. Screening of flavonoid aglycons’ metabolism mediated by the human liver cytochromes P450. Acta Pharm. 2019, 69, 541–562. [Google Scholar] [CrossRef]
- Gao, S.S.; Chen, X.Y.; Zhu, R.Z.; Choi, B.-M.; Kim, S.J.; Kim, B.-R. Dual effects of phloretin on aflatoxin B1 metabolism: Activation and detoxification of aflatoxin B1. Biofactors 2012, 38, 34–43. [Google Scholar] [CrossRef]
- Pohl, C.; Will, F.; Dietrich, H.; Schrenk, D. Cytochrome P450 1A1 Expression and Activity in Caco-2 Cells: Modulation by Apple Juice Extract and Certain Apple Polyphenols. J. Agric. Food Chem. 2006, 54, 10262–10268. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.D. The roles of cytochrome b5 in cytochrome P450 reactions. J. Biochem. Mol. Toxicol. 2002, 16, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Shimada, T. Comparative studies of in vitro inhibition of cytochrome P450 3A4-dependent testosterone 6beta-hydroxylation by roxithromycin and its metabolites, troleandomycin, and erythromycin. Drug Metab. Dispos. 1998, 26, 1053–1057. [Google Scholar] [PubMed]
- Yamazaki, H.; Shimada, T. Progesterone and Testosterone Hydroxylation by Cytochromes P450 2C19, 2C9, and 3A4 in Human Liver Microsomes. Arch. Biochem. Biophys. 1997, 346, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.H.; Woo, S.-M.; Nguyen, N.A.; Cha, G.-S.; Yeom, S.-J.; Kang, H.-S.; Yun, C.-H. Regioselective Hydroxylation of Naringin Dihydrochalcone to Produce Neoeriocitrin Dihydrochalcone by CYP102A1 (BM3) Mutants. Catalysts 2020, 10, 823. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Bondonno, C.P.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. The cardiovascular health benefits of apples: Whole fruit vs. isolated compounds. Trends Food Sci. Technol. 2017, 69, 243–256. [Google Scholar] [CrossRef]
- Waldbauer, K.; McKinnon, R.; Kopp, B. Apple Pomace as Potential Source of Natural Active Compounds. Planta Med. 2017, 83, 994–1010. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Kim, D.-O.; Lee, H.J.; Lee, C.Y. Major phenolics in apple and their contribution to the total antioxidant capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, D.; Laparra-Llopis, J.M.; Zielinski, H.; Szawara-Nowak, D.; Giménez-Bastida, J.A. Role of Apple Phytochemicals, Phloretin and Phloridzin, in Modulating Processes Related to Intestinal Inflammation. Nutrients 2019, 11, 1175. [Google Scholar] [CrossRef]
- Takeno, A.; Kanazawa, I.; Notsu, M.; Tanaka, K.; Sugimoto, T. Phloretin Promotes Adipogenesis via Mitogen-Activated Protein Kinase Pathways in Mouse Marrow Stromal ST2 Cells. Int. J. Mol. Sci. 2018, 19, 1772. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-T.; Tu, S.-H.; Yang, P.-S.; Hsu, S.-P.; Lee, W.-H.; Ho, C.-T.; Wu, C.-H.; Lai, Y.-H.; Chen, M.-Y.; Chen, L.-C. Apple Polyphenol Phloretin Inhibits Colorectal Cancer Cell Growth via Inhibition of the Type 2 Glucose Transporter and Activation of p53-Mediated Signaling. J. Agric. Food Chem. 2016, 64, 6826–6837. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Lu, N.-S.; Zhu, X.-T.; Xu, Y.; Du, M.-Q.; Xie, Q.-P.; Zhu, C.-J.; Xu, Q.; Wang, S.-B.; Wang, L.-N.; et al. Phloretin promotes adipocyte differentiation in vitro and improves glucose homeostasis in vivo. J. Nutr. Biochem. 2014, 25, 1296–1308. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Y.; Hou, S.; Huang, S.; Yang, W.; Lai, X.; Zeng, X. Identification of the bioactive components of orally administered Lithocarpus polystachyus Rehd and their metabolites in rats by liquid chromatography coupled to LTQ Orbitrap mass spectrometry. J. Chromatogr. B 2014, 962, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Pedretti, A.; Vistoli, G. Reactions and enzymes in the metabolism of drugs and other xenobiotics. Drug Discov. Today 2012, 17, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.F.; Ioannides, C.; Parke, D.V. Cytochromes P450 and species differences in xenobiotic metabolism and activation of carcinogen. Environ. Health Perspect. 1998, 106, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Lewis, D.F.V.; Eddershaw, P.J.; Goldfarb, P.S.; Tarbit, M.H. Molecular modelling of CYP3A4 from an alignment with CYP102: Identification of key interactions between putative active site residues and CYP3A-specific chemicals. Xenobiotica 1996, 26, 1067–1086. [Google Scholar] [CrossRef]
- Kapelyukh, Y.; Paine, M.J.I.; Maréchal, J.-D.; Sutcliffe, M.J.; Wolf, C.R.; Roberts, G.C.K. Multiple Substrate Binding by Cytochrome P450 3A4: Estimation of the Number of Bound Substrate Molecules. Drug Metab. Dispos. 2008, 36, 2136–2144. [Google Scholar] [CrossRef]
- Long, A.; Walker, J.D. Quantitative structure-activity relationships for predicting metabolism and modeling cytochrome P450 enzyme activities. Environ. Toxicol. Chem. 2003, 22, 1894–1899. [Google Scholar] [CrossRef]
- Mega, J.L.; Simon, T.; Collet, J.-P.; Anderson, J.L.; Antman, E.M.; Bliden, K.; Cannon, C.P.; Danchin, N.; Giusti, B.; Gurbel, P.; et al. Reduced-function CYP2C19 genotype and risk of adverse clinical outcomes among patients treated with clopidogrel predominantly for PCI: A meta-analysis. JAMA 2010, 304, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Schadt, S.; Bister, B.; Chowdhury, S.K.; Funk, C.; Hop, C.E.C.A.; Humphreys, W.G.; Igarashi, F.; James, A.D.; Kagan, M.; Khojasteh, S.C.; et al. A Decade in the MIST: Learnings from Investigations of Drug Metabolites in Drug Development under the “Metabolites in Safety Testing” Regulatory Guidance. Drug Metab. Dispos. 2018, 46, 865–878. [Google Scholar] [CrossRef]
- Lambert, J.D.; Sang, S.; Lu, A.Y.H.; Yang, C.S. Metabolism of dietary polyphenols and possible interactions with drugs. Curr. Drug Metab. 2007, 8, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.; Gillam, E.M.; Guengerich, F.P. Drug metabolism by Escherichia coli expressing human cytochromes P450. Nat. Biotechnol. 1997, 15, 784–788. [Google Scholar] [CrossRef]
- Yun, C.-H.; Yim, S.-K.; Kim, D.-H.; Ahn, T. Functional expression of human cytochrome P450 enzymes in Escherichia coli. Curr. Drug Metab. 2006, 7, 411–429. [Google Scholar] [CrossRef]
- Yun, C.H.; Ahn, T.; Guengerich, F.P.; Yamazaki, H.; Shimada, T. Phospholipase D activity of cytochrome P450 in human liver endoplasmic reticulum. Arch. Biochem. Biophys. 1999, 367, 81–88. [Google Scholar] [CrossRef]
- Kim, D.-H.; Ahn, T.; Jung, H.-C.; Pan, J.-G.; Yun, C.-H. Generation of the human metabolite piceatannol from the anticancer-preventive agent resveratrol by bacterial cytochrome P450 BM3. Drug Metab. Dispos. 2009, 37, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Le, T.-K.; Jang, H.-H.; Nguyen, H.T.H.; Doan, T.T.M.; Lee, G.-Y.; Park, K.D.; Ahn, T.; Joung, Y.H.; Kang, H.-S.; Yun, C.-H. Highly regioselective hydroxylation of polydatin, a resveratrol glucoside, for one-step synthesis of astringin, a piceatannol glucoside, by P450 BM3. Enzym. Microb. Technol. 2017, 97, 34–42. [Google Scholar] [CrossRef]
- Yamazaki, H.; Gillam, E.M.J.; Dong, M.-S.; Johnson, W.W.; Guengerich, F.P.; Shimada, T. Reconstitution of Recombinant Cytochrome P450 2C10(2C9) and Comparison with Cytochrome P450 3A4 and Other Forms: Effects of Cytochrome P450–P450 and Cytochrome P450–b5 Interactions. Arch. Biochem. Biophys. 1997, 342, 329–337. [Google Scholar] [CrossRef]
- Yun, C.H.; Shimada, T.; Guengerich, F.P. Roles of human liver cytochrome P4502C and 3A enzymes in the 3-hydroxylation of benzo(a)pyrene. Cancer Res. 1992, 52, 1868–1874. [Google Scholar] [PubMed]
- Omura, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. I. evidence for its hemopreotein nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | kcat (min−1 ) | Km (μM) | kcat/Km (min−1μM−1) |
|---|---|---|---|
| HLMs | 0.094 ± 0.006 | 120 ± 36 | 0.00083 ± 0.00025 |
| CYP3A4 | 3.1 ± 2.9 | 63 ± 11 | 0.049 ± 0.023 |
| CYP2C19 | 5.8 ± 0.6 | 208 ± 50 | 0.028 ± 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.A.; Cao, N.T.; Nguyen, T.H.H.; Le, T.-K.; Cha, G.S.; Choi, S.-K.; Pan, J.-G.; Yeom, S.-J.; Kang, H.-S.; Yun, C.-H. Regioselective Hydroxylation of Phloretin, a Bioactive Compound from Apples, by Human Cytochrome P450 Enzymes. Pharmaceuticals 2020, 13, 330. https://doi.org/10.3390/ph13110330
Nguyen NA, Cao NT, Nguyen THH, Le T-K, Cha GS, Choi S-K, Pan J-G, Yeom S-J, Kang H-S, Yun C-H. Regioselective Hydroxylation of Phloretin, a Bioactive Compound from Apples, by Human Cytochrome P450 Enzymes. Pharmaceuticals. 2020; 13(11):330. https://doi.org/10.3390/ph13110330
Chicago/Turabian StyleNguyen, Ngoc Anh, Ngoc Tan Cao, Thi Huong Ha Nguyen, Thien-Kim Le, Gun Su Cha, Soo-Keun Choi, Jae-Gu Pan, Soo-Jin Yeom, Hyung-Sik Kang, and Chul-Ho Yun. 2020. "Regioselective Hydroxylation of Phloretin, a Bioactive Compound from Apples, by Human Cytochrome P450 Enzymes" Pharmaceuticals 13, no. 11: 330. https://doi.org/10.3390/ph13110330
APA StyleNguyen, N. A., Cao, N. T., Nguyen, T. H. H., Le, T.-K., Cha, G. S., Choi, S.-K., Pan, J.-G., Yeom, S.-J., Kang, H.-S., & Yun, C.-H. (2020). Regioselective Hydroxylation of Phloretin, a Bioactive Compound from Apples, by Human Cytochrome P450 Enzymes. Pharmaceuticals, 13(11), 330. https://doi.org/10.3390/ph13110330