The Role of Dactylis Glomerata and Diesel Oil in the Formation of Microbiome and Soil Enzyme Activity




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Object
2.2. Study Design
2.3. Methodology of Microbiological Analyses
2.4. Physical, Chemical and Biological Properties of Soil
2.5. Statistical Computations and Analysis
3. Results
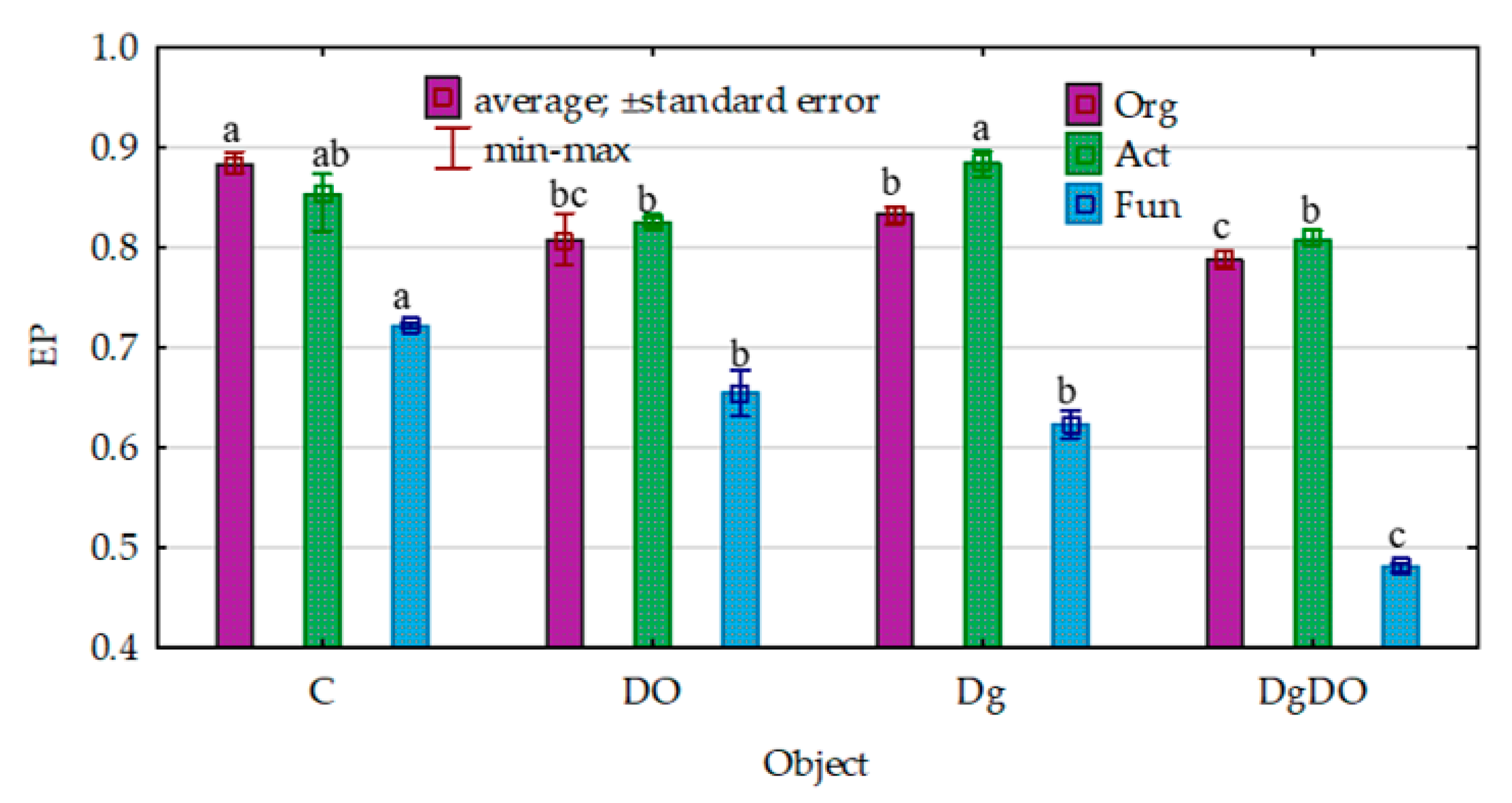
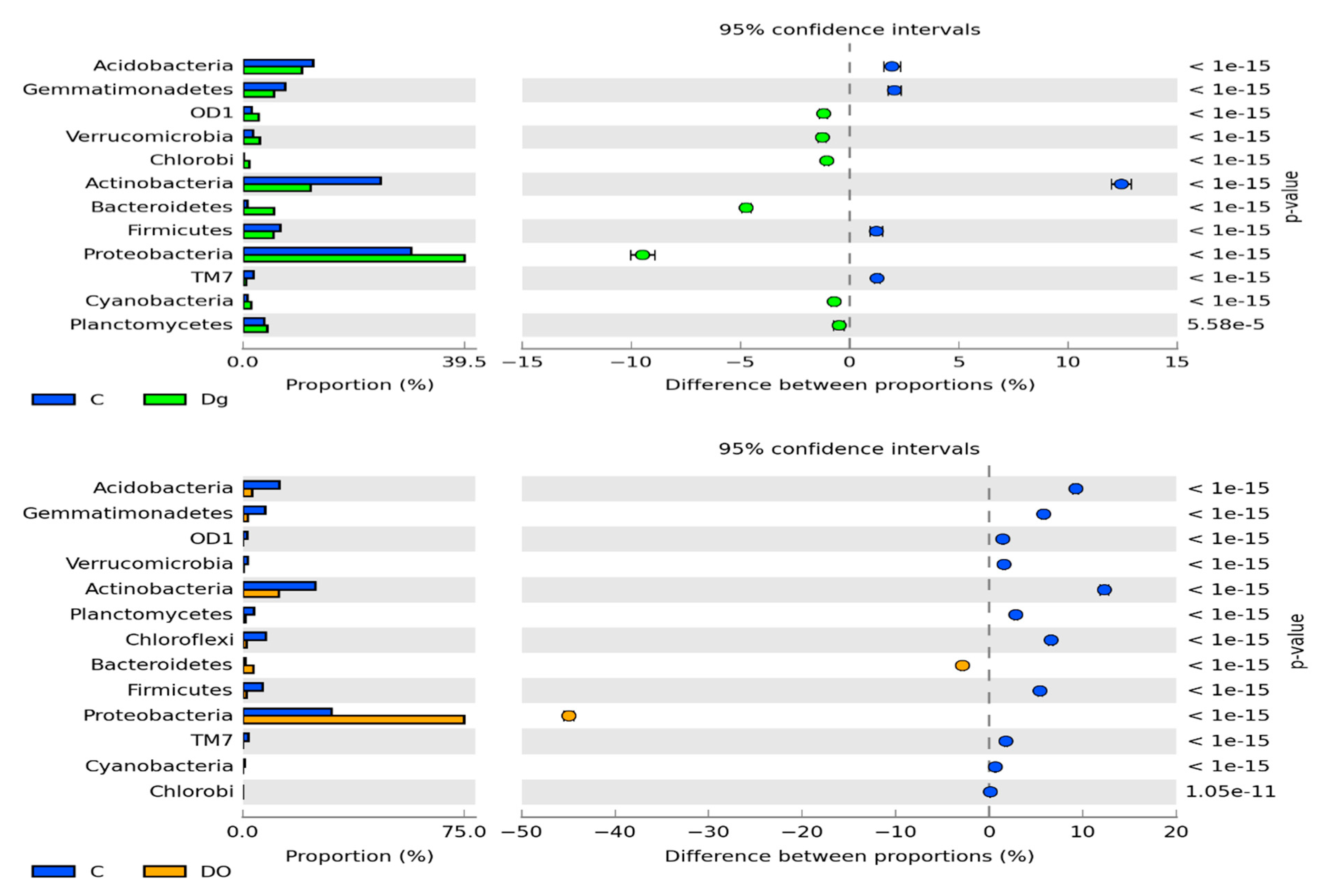
3.1. Counts and Diversity of Microorganisms in the Soil
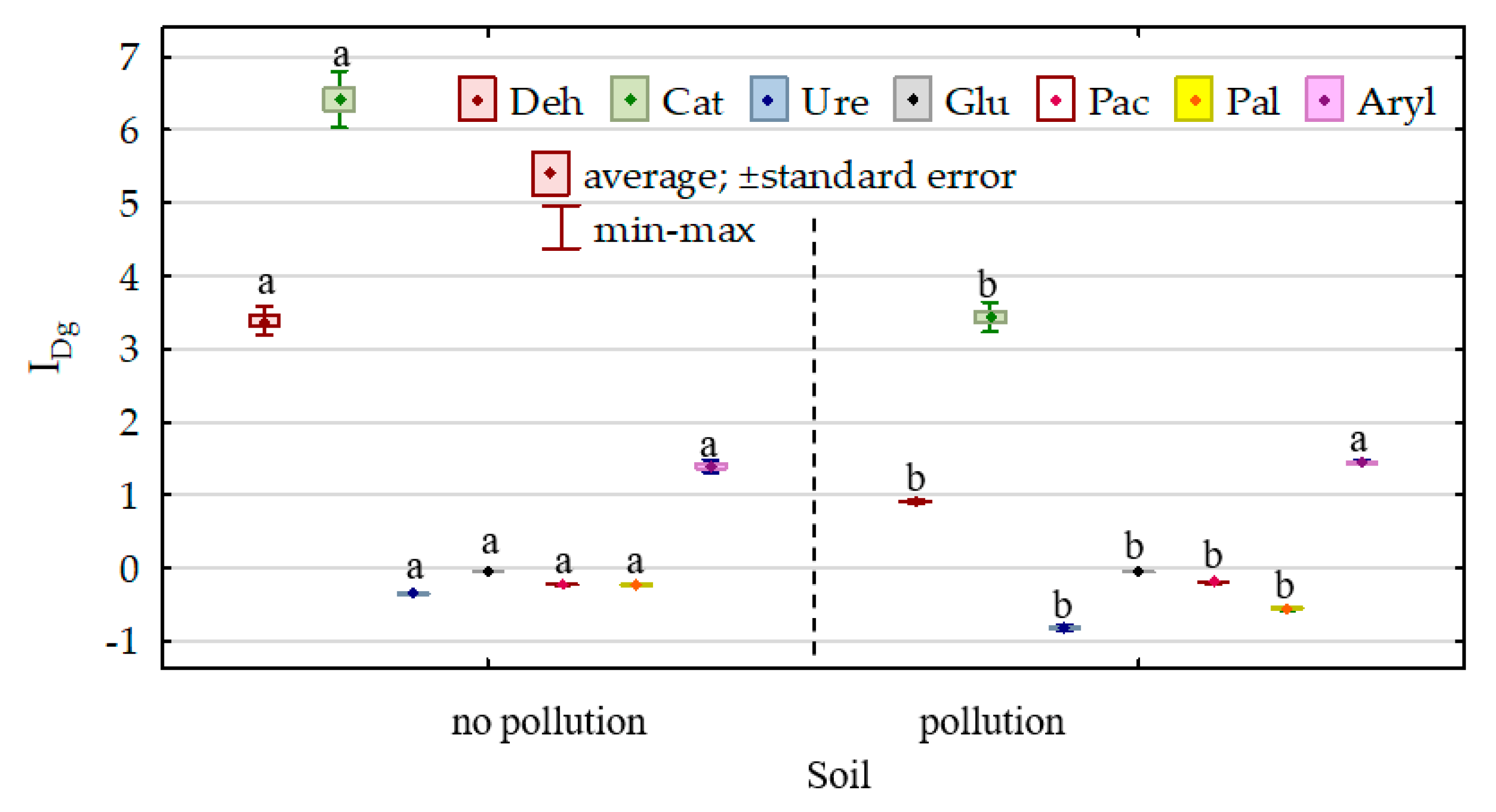
3.2. Activity of Soil Enzymes
3.3. Degradation of Hydrocarbons
3.4. Response of Dactylis Glomerata to Diesel Oil
4. Discussion
4.1. Counts and Diversity of Microorganisms in the Soil
4.2. Activity of Soil Enzymes
4.3. Degradation of Hydrocarbons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yang, Y.; Xu, M.; He, Z.; Guo, J.; Sun, G.; Zhou, J. Microbial electricity generation enhances decarbomodiphenyl ether (BDE-209) degradation. PLoS ONE 2013, 8, e70686. [Google Scholar] [CrossRef]
- Yang, S.; Wen, X.; Zhao, L.; Shi, Y.; Jin, H. Crude oil treatment leads to shift of bacterial communities in soils from the deep active layer and upper permafrost along the China-Russia crude oil pipeline route. PLoS ONE 2014, 9, e96552. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, E.; Dumolin, C.; Arends, J.B.A.; Kerckhof, F.M.; Hu, X.; Vandamme, P.; Boon, N. Enriched hydrogen-oxidizing microbiomes show a high diversity of co-existing hydrogen-oxidizing bacteria. Appl. Microbiol. 2019, 103, 8241–8253. [Google Scholar] [CrossRef] [PubMed]
- Akwukwaegbu, R.N.; Okerentugba, P.O.; Okpokwasili, G.C.; Stanley, H.O.; Ugboma, C.J. Hydrocarbon degradation potential of heterotrophic bacteria isolated from oil polluted sites in Sakpenwa community in rivers state. SAJRM 2019, 4, 1–12. [Google Scholar] [CrossRef]
- Smułek, W.; Sydow, M.; Zabielska-Matejuk, J.; Kaczorek, E. Bacteria involved in biodegradation of creosote PAH – A case study of long-term contaminated industrial area. Ecotoxicol. Environ. 2020, 187, 109843. [Google Scholar] [CrossRef] [PubMed]
- Purkamo, L.; Kietäväinen, R.; Nuppunen-Puputti, M.; Bomberg, M.; Cousins, C. Ultradeep microbial communities at 4.4 km within crystalline bedrock: Implications for habitability in a planetary context. Life 2020, 10, 2. [Google Scholar] [CrossRef]
- Ite, A.E.; Ibok, U.J. Role of plants and microbes in bioremediation of petroleum hydrocarbons contaminated soils. IJEBB 2019, 7, 1–19. [Google Scholar]
- Jiao, S.; Chen, W.; Wang, E.; Wang, J.; Liu, Z.; Li, Y.; Wei, G. Microbial succession in response to pollutants in batch-enrichment culture. Sci. Rep. 2016, 6, 21791. [Google Scholar] [CrossRef]
- Kucharski, J.; Tomkiel, M.; Boros, E.; Wyszkowska, J. The effect of soil contamination with diesel oil and petrol on the nitrification process. J. Elem. 2010, 15, 111–118. [Google Scholar] [CrossRef]
- Abena, M.T.B.; Chen, G.; Chen, Z.; Zheng, X.; Li, S.; Li, T.; Zhong, W. Microbial diversity changes and enrichment of potential petroleum hydrocarbon degraders in crude oil-, diesel-, and gasoline-contaminated soil. 3 Biotech. 2020, 10, 42. [Google Scholar] [CrossRef]
- Lipińska, A.; Kucharski, J.; Wyszkowska, J. The effect of polycyclic aromatic hydrocarbons on the structure of organotrophic bacteria and dehydrogenase activity in soil. Polycycl. Aromat. Comp. 2014, 34, 35–53. [Google Scholar] [CrossRef]
- Kucharski, J.; Wieczorek, K.; Wyszkowska, J. Changes in the enzymatic activity in sandy loam soil exposed to zinc pressure. J. Elem. 2011, 16, 577–589. [Google Scholar] [CrossRef]
- Boros-Lajszner, E.; Wyszkowska, J.; Kucharski, J. Application of white mustard and oats in the phytostabilisation of soil contaminated with cadmium with the addition of cellulose and urea. J. Soil Sediment. 2019, 20, 931–942. [Google Scholar] [CrossRef]
- Zaborowska, M.; Kucharski, J.; Wyszkowska, J. Biological activity of soil contaminated with cobalt, tin and molybdenum. Environ. Monit. Assess. 2016, 188, 398. [Google Scholar] [CrossRef] [PubMed]
- Zaborowska, M.; Kucharski, J.; Wyszkowska, J. Brown algae and basalt meal in maintaining the activity of arylsulfatase of soil polluted with cadmium. Water Air Soil Pollut. 2017, 228, 267. [Google Scholar] [CrossRef] [PubMed]
- Addorisio, V.; Pirozzi, D.; Esposito, S.; Sannino, F. Decontamination of waters polluted with simazine by sorption on mesoporous metal oxides. J. Hazard. Mater. 2011, 196, 242–247. [Google Scholar] [CrossRef]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Prieto, M.J.; Acevedo, S.O.A.; Prieto, G.F.; González, N.T. Phytoremediation of soils contaminated with heavy metals. Biodiversity Int. J. 2018, 2, 362–376. [Google Scholar] [CrossRef]
- Gałązka, A.; Gałązka, R. Phytoremediation of polycyclic aromatic hydrocarbons in soils artificially polluted using plant-associated-endophytic bacteria and Dactylis glomerata as the bioremediation plant. Pol. J. Microbiol. 2015, 64, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Water Air Soil Pollut. 2018, 9, 2885. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, Q.; Qu, L.; Li, C. Characterization of oil-degrading bacteria from oil-contaminated soil and activity of their enzymes. Biotechnol. Biotechnol. Equip. 2013, 27, 3932–3938. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J. Bioaugmentation of soil contaminated with diesel oil. J. Elem. 2018, 23, 1161–1178. [Google Scholar] [CrossRef]
- Wolińska, A.; Gałązka, A.; Kuźniar, A.; Goraj, W.; Jastrzębska, N.; Grządziel, J.; Stępniewska, Z. Catabolic fingerprinting and diversity of bacteria in mollic gleysol contaminated with petroleum substances. Appl. Sci. 2018, 8, 1970. [Google Scholar] [CrossRef]
- Yang, R.; Zhang, G.; Li, S.; Moazeni, F.; Li, Y.; Wu, Y.; Zhang, W.; Chen, T.; Liu, G.; Zhang, B.; et al. Degradation of crude oil by mixed cultures of bacteria isolated from the Qinghai-Tibet plateau and comparative analysis of metabolic mechanisms. Environ. Sci Pollut Res. 2018, 26, 1834–1847. [Google Scholar] [CrossRef] [PubMed]
- Patowary, K.; Patowary, R.; Kalita, M.C.; Deka, S. Development of an efficient bacterial consortium for the potential remediation of hydrocarbons from contaminated sites. Front. Microbiol. 2016, 7, 1092. [Google Scholar] [CrossRef]
- Obi, L.U.; Atagana, H.I.; Adeleke, R.A. Isolation and characterisation of crude oil sludge degrading bacteria. Springer Plus 2016, 5, 1946. [Google Scholar] [CrossRef]
- Ibrahim, M.L.; Ijah, U.J.J.; Manga, S.B.; Bilbis, L.S.; Umar, S. Production and partial characterization of biosurfactant produced by crude oil degrading bacteria. Int. Biodeter Biodegr. 2013, 81, 28–34. [Google Scholar] [CrossRef]
- Chandankere, R.; Yao, J.; Cai, M.; Masakorala, K.; Jain, A.K.; Choi, M.M.F. Properties and characterization of biosurfactant in crude oil biodegradation by bacterium Bacillus methylotrophicus USTBa. Fuel 2014, 122, 140–148. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Kucharski, J. The resistance of Lolium perenne L. × hybridum, Poa pratensis, Festuca rubra, F. arundinacea, Phleum pratense and Dactylis glomerata to soil pollution by diesel oil and petroleum. Plant Soil Environ. 2019, 65, 307–312. [Google Scholar] [CrossRef]
- Goncharov, A.A.; Tiunov, A.V. Trophic chains in the soil. Trophic chains in the soil. Biol. Bull. Rev. 2014, 4, 393–403. [Google Scholar] [CrossRef]
- Kucharski, J.; Jastrzębska, E. Effects of heating oil on the count of microorganisms and physico-chemical properties of soil. Polish J. Environ. Stud. 2005, 14, 195–204. [Google Scholar]
- International Energy Agency (IEA). Oil Market Report; International Energy Agency: Paris, France, 2019; p. 55. [Google Scholar]
- Kiamarsi, Z.; Soleimani, M.; Nezami, A.; Kafi, M. Biodegradation of n-alkanes and polycyclic aromatic hydrocarbons using novel indigenous bacteria isolated from contaminated soils. Int. J. Environ. Sci. Technol. 2019, 16, 6805–6816. [Google Scholar] [CrossRef]
- Nedaa, A.; Dashti, N.; Khanafer, M.; Husain, A.A.; Samir, R. Bioremediation of soils saturated with spilled crude oil. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J. Remediation of soil contaminated with diesel oil. J. Elem. 2018, 23, 767–788. [Google Scholar] [CrossRef]
- Fatima, K.; Imran, A.; Amin, I.; Khan, Q.M.; Afzal, M. Successful phytoremediation of crude-oil contaminated soil at an oil exploration and production company by plants-bacterial synergism. Int. J Phytoremediat. 2018, 20, 675–681. [Google Scholar] [CrossRef]
- Iqbal, A.; Mukherjee, M.; Rashid, J.; Khan, S.A.; Ali, M.A.; Arshad, A. Development of plant-microbe phytoremediation system for petroleum hydrocarbon degradation: An insight from alkb gene expression and phytotoxicity analysis. Sci. Total Environ. 2019, 671, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Agnello, A.C.; Bagard, M.; van Hullebusch, E.D.; Esposito, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563–564, 693–703. [Google Scholar] [CrossRef]
- Estendorfer, J.; Stempfhuber, B.; Haury, P.; Vestergaard, G.; Rillig, M.C.; Joshi, J.; Schröder, P.; Schloter, M. The influence of land use intensity on the plant-associated microbiome of Dactylis glomerata L. Front. Plant Sci. 2017, 8, 930. [Google Scholar] [CrossRef]
- Mika, V.; Kohoutek, A.; Odstrcilova, V. Characteristics of important diploid and tetraploid subspecies of Dactylis from poInt. of view of the forage crop production. Rostl. Vyroba 2002, 48, 243–248. [Google Scholar] [CrossRef]
- Paliwa, B.P. Available online: https://www.bp.com/pl_pl/poland/home/produkty_uslugi/paliwa.html (accessed on 30 March 2020).
- Statistical Report 2019. FuelsEurope 2019, 1–68. Available online: https://www.fuelseurope.eu/wp-content/uploads/FuelsEurope-Statistical-Report-2019-2.pdf (accessed on 23 March 2020).
- Sanderson, M.A.; Skinner, R.H.; Elwinger, G.F. Seedling development and field performance of prairiegrass, grazing bromegrass, and orchard grass. Crop Sci. 2002, 42, 224–230. [Google Scholar] [CrossRef]
- Kaimi, E.; Mukaidani, T.; Miyoshi, S.; Tamaki, M. Ryegrass enhancement of biodegradation in diesel-contaminated soil. Environ. Exp. Bot. 2006, 55, 110–119. [Google Scholar] [CrossRef]
- Ruley, J.A.; Tumuhairwe, J.B.; Amoding, A.; Opolot, E.; Oryem-Origa, H.; Basamba, T. Assessment of plants for phytoremediation of hydrocarbon-contaminated soils in the Sudd Wetland of South Sudan. Plant Soil Environ. 2019, 65, 463–469. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Wyszkowski, M. Resistance of aerobic microorganisms and soil enzyme response to soil contamination with Ekodiesel Ultra fuel. Environ. Sci. Pollut. Res. 2017, 24, 24346–24363. [Google Scholar] [CrossRef] [PubMed]
- Öhlinger, R. Dehydrogenase activity with the substrate TTC. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandler, E., Margesin, R., Eds.; Springer: Berlin, Germany, 1996; pp. 241–243. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: Cambridge, MA, USA, 1998; pp. 316–365. [Google Scholar]
- Johnson, J.I.; Temple, K.L. Some variables affecting the measurement of catalase activity in soil. Soil Sci. Soc. Am. Pro. 1964, 28, 207–216. [Google Scholar] [CrossRef]
- ISO 18287. In Soil Quality—Determination of Polycyclic Aromatic Hydrocarbons (PAH)—Gas Chromatographic Method with Mass Spectrometric Detection (GC-MS); International Organization for Standardization: Geneva, Switzerland, 2006.
- EN ISO 16703. In Soil Quality—Determination of Content of Hydrocarbon in the Range C10 to C40 by Gas Chromatography; International Organization for Standardization: Geneva, Switzerland, 2004.
- EN ISO 22155. In Soil Quality—Gas Chromatographic Determination of Volatile Aromatic and Halogenated Hydrocarbons and Selected Ethers—Static Headspace Method; International Organization for Standardization: Geneva, Switzerland, 2016.
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Method of Soil Analysis; Sparks, D.L., Ed.; Chemical Methods, American Society of Agronomy: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar]
- ISO 11261. In Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method; International Organization for Standardization: Geneva, Switzerland, 1995.
- Egner, H.; Riehm, H.; Domingo, W.R. Untersuchungen über diechemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chemische Extractions methoden zur Phosporund Kaliumbestimmung. Ann. R. Agric. Coll. Swed. 1960, 26, 199–215. [Google Scholar]
- Schlichting, E.; Blume, H.P.; Stahr, K. Bodenkundliches Praktikum. In Pareys Studientexte; Auflage, 295 S.; Blackwell Wissenschafts-Verlag: Berlin, Germany, 1995; Volume 81, p. 2. [Google Scholar]
- PN-R-04032. In Soil and MINERAL materials—Sampling and Determination of Particle Size Distribution; Polish Committee for Standardization: Warsaw, Poland, 1998.
- ISO 11464. In Soil Quality—Pre-Treatment of Samples for Physico-Chemical Analysis; International Organization for Standardization: Geneva, Switzerland, 2006.
- ISO 10390. In Soil Quality—Determination of pH; International Organization for Standardization: Geneva, Switzerland, 2005.
- Klute, A. Methods of Soil Analysis; American Society of Agronomy, Agronomy Monograph 9: Madison, WI, USA, 1996. [Google Scholar]
- De Leij, F.A.A.M.; Whipps, J.M.; Lynch, J.M. The use of colony development for the characterization of bacterial communities in soil and on roots. Microb. Ecol. 1993, 27, 81–97. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 23 February 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 23 February 2020).
- Gregory, R.W.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. gplots: Various R Programming Tools for Plotting Data; R Package Version 2.17.0. 2020. Available online: https://CRAN.R-project.org/package=gplots (accessed on 23 February 2020).
- Orwin, K.H.; Wardle, D.A. New indices for quantifying the resistance and resilience of soil biota to exogenous disturbances. Soil Biol. Biochem. 2004, 36, 1907–1912. [Google Scholar] [CrossRef]
- Dell Inc. Dell Statistica (Data Analysis Software System), Version 13.1; Dell Inc.: Tulsa, OK, USA, 2016. [Google Scholar]
- Gu, Y.; Wang, Y.; Lu, S.; Xiang, Q.; Yu, X.; Zhao, K.; Zou, L.; Chen, Q.; Tu, S.; Zhang, X. Long-term fertilization structures bacterial and archaeal communities along soil depth gradient in a paddy soil. Front. Microbiol. 2017, 8, 1516. [Google Scholar] [CrossRef]
- Kumar, V.; AlMomin, S.; Al-Aqeel, H.; Al-Salameen, F.; Nair, S.; Shajan, A. Metagenomic analysis of rhizosphere microflora of oil-contaminated soil planted with barley and alfalfa. PLoS ONE 2018, 13, e0202127. [Google Scholar] [CrossRef]
- Gałązka, A.; Grządziel, J.; Gałązka, R.; Ukalska-Jaruga, A.; Strzelecka, J.; Smreczak, B. Genetic and functional diversity of bacterial microbiome in soils with long term impacts of petroleum hydrocarbons. Front. Microbiol. 2018, 9, 1923. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N.A. Global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Wemheuer, F.; Kaiser, K.; Karlovsky, P.; Daniel, R.; Vidal, S.; Wemheuer, B. Bacterial endophyte communities of three agricultural important grass species differ in their response towards management regimes. Sci Rep. 2017, 7, 40914. [Google Scholar] [CrossRef] [PubMed]
- Islam, Z.F.; Cordero, P.R.F.; Feng, J.; Chen, Y.-J.; Bay, S.K.; Jirapanjawat, T.; Gleadow, R.M.; Carere, C.R.; Stott, M.B.; Chiri, E.; et al. Two Chloroflexi classes independently evolved the ability to persist on atmospheric hydrogen and carbon monoxide. ISME J. 2019, 13, 1801–1813. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 70, 6295–6300. [Google Scholar] [CrossRef]
- de León-Lorenzana, A.S.; Delgado-Balbuena, L.; Domínguez-Mendoza, C.; Navarro-Noya, Y.E.; Luna-Guido, M.; Dendooven, L. Reducing salinity by flooding an extremely alkaline and saline soil changes the bacterial community but its effect on the archaeal community is limited. Front. Microbiol. 2017, 8, 466. [Google Scholar] [CrossRef]
- Raju, M.N.; Leo, R.; Herminia, S.S.; Morán, R.E.; Venkateswarlu, K.; Laura, S. Biodegradation of diesel, crude oil and spent lubricating oil by soil isolates of Bacillus spp. Bull Environ Contam Toxicol. 2017, 98, 698–705. [Google Scholar] [CrossRef]
- Mukherjee, A.; Chettri, B.; Langpoklakpam, J.S.; Basak, P.; Prasad, A.; Mukherjee, A.K.; Bhattacharyya, M.; Singh, A.K.; Chattopadhyay, D. Bioinformatic approaches including predictive metagenomic profiling reveal characteristics of bacterial response to petroleum hydrocarbon contamination in diverse environments. Sci. Rep. 2017, 7, 1108. [Google Scholar] [CrossRef]
- Røberg, S.; Østerhus, J.I.; Landfald, B. Dynamics of bacterial community exposed to hydrocarbons and oleophilic fertilizer in high-arctic intertidal beach. Polar Biol. 2011, 34, 1455–1465. [Google Scholar] [CrossRef][Green Version]
- Hemkemeyer, M.; Dohrmann, A.B.; Christensen, B.T.; Tebbe, C.C. Bacterial preferences for specific soil particle size fractions revealed by community analyses. Front. Microbiol. 2018, 9, 149. [Google Scholar] [CrossRef]
- Telesiński, A.; Krzyśko-Łupicka, T.; Cybulska, K.; Pawłowska, B.; Biczak, R.; Śnieg, M.; Wróbel, J. Comparison of oxidoreductive enzyme activities in three coal tar creosote-contaminated soils. Soil Res. 2019, 57, 814–824. [Google Scholar] [CrossRef]
- Lipińska, A.; Kucharski, J.; Wyszkowska, J. Activity of arylsulphatase in soil contaminated with polycyclic aromatic hydrocarbons. Water Air Soil Pollut. 2014, 225, 2097. [Google Scholar] [CrossRef] [PubMed]
- Wolińska, A.; Kuźniar, A.; Szafranek-Nakonieczna, A.; Jastrzębska, N.; Roguska, E.; Stępniewska, Z. Biological activity of autochthonic bacterial community in oil-contaminated soil. Water Air Soil Pollut. 2016, 227, 130. [Google Scholar] [CrossRef] [PubMed]
- Lipińska, A.; Kucharski, J.; Wyszkowska, J. Activity of phosphatases in soil contaminated with PAHs. Water Air Soil Pollut. 2019, 230, 298. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Kucharski, M.; Kucharski, J. Application of the activity of soil enzymes in the evaluation of soil contamination by diesel oil. Polish J. Environ. Stud. 2006, 15, 501–506. Available online: http://www.pjoes.com/Application-of-the-Activity-of-Soil-Enzymes-in-the-Evaluation-of-Soil-Contamination,87896,0,2.html (accessed on 23 March 2020).
- Xiaoyan, Z.; Xianghua, W.; Yan, F. Influence of glucose feeding on the ligninolytic enzyme production of the white-rot fungus Phanerochaete chrysosporium. Front. Environ. Sci. Eng. China 2007, 1, 89–94. [Google Scholar] [CrossRef]
- Chen, R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Chang. Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Zhang, C.; Tan, X.; Yang, X.; Wan, C.; Lee, D.J. Enhancement of anaerobic degradation of petroleum hydrocarbons by electron intermediate: Performance and mechanism. Bioresour. Technol. 2020, 295, 122305. [Google Scholar] [CrossRef]
- Gianfreda, L. Enzymes of importance to rhizosphere processes. J. Soil Sci. Plant Nut. 2015, 15, 283–306. [Google Scholar] [CrossRef]
- Duly, O.; Nannipieri, P. Intracellular and extracellular enzyme activity in soil with reference to elemental cycling. Zeitschrift für Pflanzenernährung und Bodenkunde 1998, 161, 243–248. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Sarma, H.; Nava, A.R.; Prasad, M.N.V. Mechanistic understanding and future prospect of microbe-enhanced phytoremediation of polycyclic aromatic hydrocarbons in soil. Environ. Technol. Inno. 2018, 13, 318–330. [Google Scholar] [CrossRef]
- Hou, L.; Liu, R.; Li, N.; Dai, Y.; Yan, J. Study on the efficiency of phytoremediation of soils heavily polluted with PAHs in petroleum-contaminated sites by microorganism. Environ. Sci Pollut Res. 2019, 26, 31401–31413. [Google Scholar] [CrossRef] [PubMed]
- Placek, A.; Grobelak, A.; Kacprzak, M. Improving the phytoremediation of heavy metals contaminated soil by use of sewage sludge. Int. J. Phytoremediat. 2015, 18, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tan, X.; Guo, J.; Liang, X.; Xie, Q.; Chen, S. Bioremediation of oil-contaminated soil by combination of soil conditioner and microorganism. J Soils Sediments 2020, 20, 2121–2129. [Google Scholar] [CrossRef]
- An, D.; Brown, D.; Chatterjee, I.; Dong, X.; Ramos-Padron, E.; Wilson, S.; Bordenave, S.; Caffrey, S.M.; Gieg, L.M.; Sensen, C.W. Microbial community and potential functional gene diversity involved in anaerobic hydrocarbon degradation and methanogenesis in an oil sands tailings pond. Genome 2013, 56, 612–618. Available online: https://www.nrcresearchpress.com/doi/full/10.1139/gen-2013-0083#.Xleb0dThDGg (accessed on 23 March 2020). [CrossRef]
- Chukwuma, C.; Ikewuchi, J.; Monanu, M. Removal of hydrocarbons from crude oil contaminated agricultural soil by phytoremediation using Mariscus alternifolius and Fimbristylis ferruginea. Eur. J. Biol. Res. 2019, 9, 34–44. [Google Scholar] [CrossRef]
- Cheng, L.; Zhou, Q.; Yu, B. Responses and roles of roots, microbes, and degrading genes in rhizosphere during phytoremediation of petroleum hydrocarbons contaminated soil. Int. J. Phytoremediat. 2019, 21, 1161–1169. [Google Scholar] [CrossRef]
- Hunt, L.J.; Duca, D.; Dan, T.; Knopper, L.D. Petroleum hydrocarbon (PHC) uptake in plants: A literature review. Environ. Pollution. 2019, 245, 472–484. [Google Scholar] [CrossRef]
- Sandermann, H. Higher plant metabolism of xenobiotics: The ‘green liver’ concept. Pharmacogenetics 1994, 4, 225–241. [Google Scholar] [CrossRef]
- Jing, W.; Xinying, Z.; Xiaoyan, L.; Xia, L.; Xueping, C. Influence of root components of celery on pyrene bioaccessibility, soil enzymes and microbial communities in pyrene and pyrene-diesel spiked soils. Sci. Total Environ. 2017, 599–600, 50–57. [Google Scholar] [CrossRef]
- Al-Mailem, D.M.; Kansour, M.K.; Radwan, S.S. Cross-bioaugmentation among four remote soil samples contaminated with oil exerted just inconsistent effects on oil-bioremediation. Front. Microbiol. 2019, 10, 2827. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Object 1 | Deh | Cat | Ure | Glu | Pac | Pal | Aryl |
|---|---|---|---|---|---|---|---|
| µmol TFF | Mol O2 | mmol N-NH4 | mmol PNP | ||||
| C | 0.666 c | 0.098 d | 0.364 b | 0.277 c | 1.201 b | 0.160 b | 0.102 d |
| DO | 3.137 b | 0.290 c | 2.134 a | 0.334 a | 1.298 a | 0.346 a | 0.270 b |
| Dg | 2.920 b | 0.727 b | 0.235 c | 0.265 c | 0.930 d | 0.122 c | 0.244 c |
| DgDO | 5.987 a | 1.286 a | 0.387 b | 0.316 b | 1.034 c | 0.154 b | 0.659 a |
| Object | C6–C12 | C12–C35 | Ben | EtB | Tol | Xyl | Nap |
| NS | 65.92 b | 52.26 b | 44.50 b | 98.01 a | 97.86 b | 98.61 a | 97.22 b |
| S | 82.79 a | 60.87 a | 75.00 a | 99.36 a | 99.31 a | 99.55 a | 98.86 a |
| Object | Ant | Chr | BaA | BaP | BbF | BkF | IP |
| NS | 80.52 b | 13.91 b | 18.75 b | 27.78 b | 18.18 b | 22.00 b | 10.00 b |
| S | 92.78 a | 52.17 a | 37.50 a | 44.44 a | 33.33 a | 40.00 a | 12.50 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borowik, A.; Wyszkowska, J.; Kucharski, M.; Kucharski, J. The Role of Dactylis Glomerata and Diesel Oil in the Formation of Microbiome and Soil Enzyme Activity. Sensors 2020, 20, 3362. https://doi.org/10.3390/s20123362
Borowik A, Wyszkowska J, Kucharski M, Kucharski J. The Role of Dactylis Glomerata and Diesel Oil in the Formation of Microbiome and Soil Enzyme Activity. Sensors. 2020; 20(12):3362. https://doi.org/10.3390/s20123362
Chicago/Turabian StyleBorowik, Agata, Jadwiga Wyszkowska, Mirosław Kucharski, and Jan Kucharski. 2020. "The Role of Dactylis Glomerata and Diesel Oil in the Formation of Microbiome and Soil Enzyme Activity" Sensors 20, no. 12: 3362. https://doi.org/10.3390/s20123362
APA StyleBorowik, A., Wyszkowska, J., Kucharski, M., & Kucharski, J. (2020). The Role of Dactylis Glomerata and Diesel Oil in the Formation of Microbiome and Soil Enzyme Activity. Sensors, 20(12), 3362. https://doi.org/10.3390/s20123362