1. Introduction
One of the largest biotic evolutionary events in the Phanerozoic is the end-Permian mass extinction followed by the Triassic biotic recovery. This was associated with the collapse of the Paleozoic ecosystem and the establishment of a more “modern-type” ecosystem, and involving one of the most drastic changes in global paleoenvironment and paleoclimate [
1,
2,
3,
4,
5]. It represents a major transition where the air-breathing tetrapods played a key role. They invaded the seas and grew to be the largest animals in marine communities; some became the top predators in their ecosystems [
6], while the fishes and invertebrates also showed marked changes.
The late Ladinian (late Middle Triassic) Xingyi Fauna marks a key period in the major transition towards the “Modern-type” ecosystem. It is well defined by the appearance of large marine tetrapods and the earliest teleost fishes. The first marine reptile found in this fauna was the small pachypleurosaurid sauropterygian,
Keichousaurus hui Young, 1958, from the Zhuganpo Member of the Falang Formation at Dingxiao Town, Xingyi City, Guizhou Province, in southwest China [
7] (
Figure 1), and the first reported fishes were
Habroichthys orientalis (Su, 1959),
Asialepidotus shingyiensis Su, 1959, and
Sinoeugnathus kueichowensis Su, 1959 [
8]. Following their discoveries, abundant and complete skeletons of
Keichousaurus and
Asialepidotus were excavated by local farmers, but very few were collected and deposited in public institutes and museums, and there were almost no detailed studies of these taxa after their initial announcement and brief descriptions. Consequently, the diversity of the Xingyi Fauna was poorly understood; even the exact geographic and geological data concerning the fossil sites were not well constrained until the beginning of the 21st century [
9]. Since 2001, besides
Keichousaurus and
Asialepidotus, more late Ladinian marine reptiles and fishes have been found, not only at Dingxiao of Xingyi, the original site of the Xingyi Fauna, but also in the broad region around Xingyi City, especially at Wusha Town in the western part of Xingyi City, Banqiao Town in east Luoping County, and Shibalianshan Town in east Fuyuan County of Yunnan Province (
Figure 1). Although the geographic distribution of the Xingyi Fauna became better known, the stratigraphic distribution of individual taxa, the general diversity of the assemblage, and the absolute age of the fauna still remained poorly understood.
In 2007, an international collaborative research team composed of the Geological Museum of Peking University, the Department of Earth and Planetary Sciences of the University of California (Davis), the Department of Earth Sciences ‘A. Desio’, University of Milan, and the Field Museum (Chicago) began field investigations of the Xingyi Fauna, and a large and continuous excavation was carried out between 2008 and 2013. In total, thousands of skeletons of marine reptiles and fishes, as well as several invertebrate fossils, were collected, and their occurrences were recorded bed by bed. These fossils were prepared in a facility established locally, where new taxa and fossil assemblages were uncovered. The excavation site has since been protected, and an on-site field museum has been constructed to protect the stratigraphic sequence. The UNESCO Geopark Museum has also been built to exhibit and house the fossils collected during the excavations. The purpose of the present paper is to record and summarize the vertebrate diversity of the Xingyi Fauna, based both on the field investigations since 2007 as well as tprevious publications.
Institutional Abbreviations—GMPKU, Geological Museum, Peking University, Beijing; IVPP, Institute of Vertebrate Palaeontology and Palaeoanthropology, Chinese Academy of Science, Beijing; NIGP, Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing; NMNS, National Museum of Nature and Sciences, Taiwan; XNGM WS (=XNGM XY), Xingyi National Geopark Museum at Wusha Town, Xingyi City, Guizhou Province; ZMNH, Zhejiang Museum of Natural History, Hangzhou City, Zhejiang Province.
3. Diversity of the Marine Reptiles of the Xingyi Fauna
To date, 20 species of marine reptiles have been named or mentioned as present in the Xingyi Fauna, including nine sauropterygians, two ichthyopterygians, four thalattosauriforms, three archosauronmorphs, and two archosaurs (
Figure 4). The index taxon of the marine reptiles in the Xingyi Fauna is the pachypleurosaurid sauropterygian
Keichousaurus hui Young, 1958, which was the first reported marine reptile found and described in China [
7,
18,
19], while the more striking taxa are large ichthyosaurs and pistosaur sauropterygians. The list and short notes follow:
| |
| Sauropterygians: |
| Superorder Sauropterygia Owen, 1860 [20], |
| Order Placodontia Cope, 1871 [21], |
| Suborder Cyamodontoidea Nopcsa, 1923 [22], |
| Family Placochelyidae Romer, 1956 [23], |
| Genus Glyphoderma Zhao et al., 2008 [24], |
| Species Glyphoderma kangi Zhao et al., 2008 [24]. |
Glyphoderma kangi represents the oldest fossil record of an armored placodont reported in South China (
Figure 4h). The body size of the published specimens lies between 87 and 95 cm in length, suggesting a small- to medium-sized sauropterygian with a dorsal carapace and crushing teeth. The holotype was collected from Jiyangshan, Shibalianshan Town, eastern Fuyuan County of Yunnan Province, in addition to two recently described specimens [
24,
25]. The exact information of the fossil site and stratigraphy remain unclear, but probably they were excavated from the stratigraphic level correlated to the Lower Assemblage of the Xingyi Fauna.
| |
| Order Eosauropterygia Rieppel, 1994 [26], |
| Suborder Pachypleurosauria Nopcsa, 1928 [27], |
| Family Pachypleurosauridae Nopcsa, 1928 [28], |
| Genus Keichousaurus Young, 1958 [7], |
| Species Keichousaurus hui Young, 1958 [7]. |
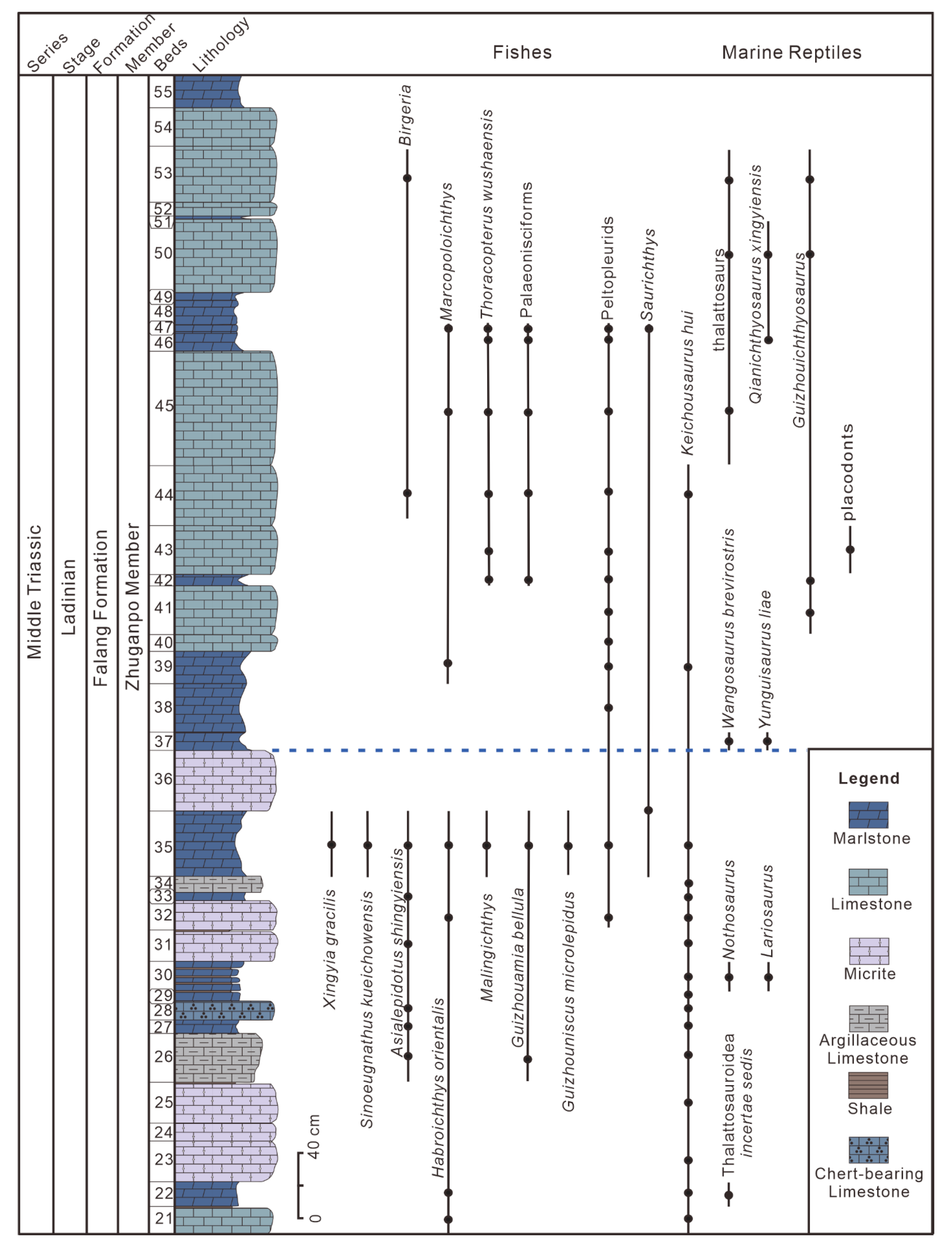
This was a small marine reptile, usually 10 to 30 cm long as an adult (
Figure 4f), with a broad geographic distribution, from Dingxiao to Wusha Town in Xingyi and further westward to eastern Fuyuan and Luoping of Yunnan Province. More than a thousand complete and well-articulated skeletons of
Keichousaurus hui have been discovered, typically from Bed 35 of the Lower Assemblage in the fossiliferous beds, which is a sedimentary bed 40–50 cm thick and comprising dark grey to black laminated marls and shales. This taxon ranges from Bed 21 to 44, but it is concentrated in the lower part of the sequence, from Bed 21 to 35 (
Figure 3). In Dingxiao, the type locality,
Keichousaurus is present throughout the sequence exposed near to the village on the top of the hill (Beds 181–193; the bed numbers are not continuous with the Wusha numbers), and sometimes mass mortality layers have been recorded (Bed 188) with up to four specimens per square meter. Oddly enough, both Bed 188 in Dingxiao and Bed 25 in Nimaugu yield a very high number of specimens, of very different sizes, but no other fossils are recorded from the beds, apart from a few specimens of
Schimperella, making the assemblage of these two beds unique.
A series of studies on its morphology, anatomy, ontogeny, phylogeny, viviparity, sexual dimorphism, and ecology have been conducted (e.g., [
28,
29,
30,
31,
32,
33]), although a detailed investigation on the bed-by-bed collected specimens is still underway.
| |
| Genus Qianxisaurus Cheng et al., 2012 [34], |
| Species Qianxisaurus chajiangensis Cheng et al., 2012 [34]. |
Qianxisaurus chajiangensis is a small- to medium-sized pachypleurosaurid eosauropterygian, and the holotype (NMNS-KIKO-F044630) is about 80 cm long in length (
Figure 4k). It appears morphologically similar to the European pachypleurosaurs
Serpianosaurus and
Neusticosaurus [
34]. The specimens of this taxon are rare, and the exact locality and horizon of the holotype is unknown, but according to our communications with the local fossil collectors, it probably was collected from the Lower Assemblage around Wusha Town.
| |
| Suborder Eusauropterygia Tschanz, 1989 [35], |
| Family Nothosauridae Baur, 1889 [36], |
| Genus Nothosaurus Münster, 1834 [37], |
| Species Nothosaurus youngi Li and Rieppel, 2004 [38]. |
Nothosaurus youngi is a small- to medium-sized nothosaurid sauropterygian, generally larger than, or at least of a similar body size to the coeval sister taxon
Lariosaurus xingyiensis (
Figure 4l). It may reach 2 m or even larger in total length, and its skull may reach 16–17 cm in length with large fangs and large upper temporal fenestrae. Abundant and complete skeletons of this species were collected and exhibited as one of the index fossils of the Xingyi Fauna. The specimen XNGM WS-30-R24 (
Figure 4l) was discovered from Bed 30 in the Lower Assemblage of the Xingyi Fauna at Wusha Town [
39] probably along with many other documented
N. youngi specimens, including the holotype (IVPP V 13590).
| |
| Genus Lariosaurus Curioni, 1847 [40], |
| Species Lariosaurus xingyiensis Li et al., 2002 [41]. |
Lariosaurus xingyiensis is a small- to medium-sized nothosaurid, with the holotype (IVPP V11866) being about 1.1 m in length and morphologically similar to the holotype of
Nothosaurus youngi. This species is smaller in size than
N. youngi and has a more flattened humerus and broader ulna. The upper temporal fenestra is longer than the orbit, but not as large as that in
N. youngi. The referred specimen (XNGM WS-30-R19) was also excavated from Bed 30 at Wusha Town (
Figure 4o). The preserved length of the skeleton is 70.5 cm, smaller than the holotype but more similar to the European
Lariosaurus [
42].
L. xingyiensis appears to be much less common than
N. youngi.
| |
| Genus Brevicaudosaurus Shang et al., 2020 [43], |
| Species Brevicaudosaurus jiyangshanensis Shang et al., 2020 [43]. |
Brevicaudosaurus jiyangshanensis is a small nothosaurid. The holotype (IVPP V 18625) is about 58.8 cm in length, and a referred specimen (IVPP V 26010) is even smaller (
Figure 4m). They were reported from the Zhuganpo Member (Ladinian) of the Falang Formation at Jiyangshan and Daqingtou in Shibalianshan Town, Fuyuan County of Yunnan Province. This species is the sister taxon of the
Nothosaurus–
Lariosaurus clade within Nothosauroidea, characterized by its short trunk with 14 dorsal vertebrae and short tail which is shorter than the skull–neck region [
43]. No precise stratigraphic information is known for the described specimens, although it likely belongs to the Lower Assemblage.
| Infraorder Pistosauroidea Baur, 1987–90 [44], |
| Family incertae sedis, |
| Genus Wangosaurus Ma et al., 2015 [45], |
| Species Wangosaurus brevirostris Ma et al., 2015 [45]. |
Wangosaurus brevirostris is a medium-sized long-necked pistosauroid sauropterygian, with a body length of about 2.2 m (
Figure 4d). The phylogenetic analysis [
45] indicates it is a basal member of the clade Pistosauroidea, although it also shares some similarities with the nothosaurid
Nothosaurus. The holotype (GMPKU-P-1529) was excavated from Bed 37 in the Upper Assemblage of the Xingyi Fauna at Wusha Town (
Figure 3). This species fills the “morphological gap” between the Triassic nothosaurids and the Jurassic–Cretaceous plesiosaurs.
| |
| Genus Yunguisaurus Cheng et al., 2006 [46], |
| Species Yunguisaurus liae Cheng et al., 2006 [46], |
| Genus Dingxiaosaurus Liu et al., 2002 [47], |
| Species Dingxiaosaurus luyinensis Liu et al., 2002 [47]. |
Yunguisaurus liae is a large pistosauroid sauropterygian (3–5 m length as an adult) with a long neck and long flippers (
Figure 4e). It is interpreted to be a derived Triassic pistosauroid most closely related to the Jurassic–Cretaceous plesiosaurs [
46,
48,
49]. According to local fossil collectors, the holotype was probably also collected from the stratigraphic interval above Bed 37 in the Upper Assemblage of the Xingyi Fauna at Wusha Town.
The holotype of
Dingxiaosaurus luyinensis is a fragmentary specimen that consists of only a pair of hind limbs, a few disarticulated vertebrae and ribs [
47] (
Figure 4j). It was collected by local farmers and donated to the Museum of
Keichousaurus at Dingxiao of Xingyi. The specimen was purported to be from the Yangliujing Formation, which is below the fossiliferous beds of the Xingyi Fauna in the Zhuganpo Member of the Falang Formation but this cannot be confirmed. The morphology of the limb bones, vertebrae, and ribs closely resembles that of
Yunguisaurus liae, which suggests that
D. luyinensis is a pistosauroid, very similar to
Y. liae. It is possible that
D. luyinensis is a senior synonym of
Y. liae or a
nomen dubium [
50], but additional material and further studies are required to clarify the situation.
| |
| Ichthyopterygians: |
| Superorder Ichthyopterygia Owen, 1840 [51], |
| Order Ichthyosauria Blainville, 1835 [52], |
| Suborder Merriamosauriformes McGowan and Motani, 2003 [53], |
| Family Shastasauridae Merriam, 1902 [54], |
| Genus Guizhouichthyosaurus Yin in Yin et al., 2000 [55], |
| Species Guizhouichthyosaurus sp. (the species is still unnamed). |
A few large long-snouted ichthyosaur specimens from beds 42, 46, 50, and 53 in the Upper Assemblage of the Xingyi Fauna at Wusha Town were excavated, with estimated total lengths of 5–9 m or even larger in some adults (
Figure 3 and
Figure 4a). They probably represent a new species of
Guizhouichthyosaurus (Shastasauridae, Ichthyosauria, Ichthyopterygia, Reptilia), but they are still under study. This taxon shows morphological similarities to the Late Triassic
Guizhouichthyosaurus tangae and is phylogenetically close to the North American Late Triassic
Shastasaurus. The body size of this species is the largest in the Ladinian Xingyi Fauna. Together with the cymbospondylids
Thalattoarchon and
Cymbospondylus from Nevada (USA) [
56,
57], it is one of the largest species in the Middle Triassic marine realm. Specimen XNGM-WS-50-R4, excavated from Bed 50, is 5 m in length and presents the oldest direct evidence for predation on megafauna. In its stomach, the trunk of a 4 m long thalattosaur,
Xinpusaurus xingyiensis, was discovered. This is also the largest prey found inside any Mesozoic marine reptile [
58], which demonstrates that this large ichthyosaur species had become the top predator in the Middle Triassic marine ecosystem.
| |
| Family Toretocnemidae Maisch and Matzke, 2000 [59], |
| Genus Qianichthyosaurus Li, 1999 [60], |
| Species Qianichthyosaurus xingyiensis Yang et al., 2013 [61]. |
Qianichthyosaurus xingyiensis is a medium-sized ichthyosaur, with a length between 1.5 m and 2 m as an adult. It has a typical ichthyosaurian tail bend, large orbit, and narrow postorbital region (
Figure 4c). The holotype XNGM WS2011-46-R1 was excavated from Bed 46 in the Upper Assemblage of the Xingyi Fauna at Wusha Town and has been reposited in the Xingyi UNESCO Geopark Museum, while the other referred specimen XNGM WS2011-50-R7 is still preserved and protected on site at Wusha Town. This species is the sister taxon of
Qianichthyosaurus zhoui from the Upper Triassic Wayao Member of the Falang Formation at Guanling [
60], and the genus is the sister taxon of the North American Late Triassic
Toretocnemus [
62].
Q. xingyiensis is the oldest member of the family Toretocnemidae and shows a strong phylogenetic relationship to the North American taxon and biogeographic affinity to east Panthalassa.
| |
| Thalattosauriforms: |
| Superorder Thalattosauriformes Nicholls, 1999 [63], |
| Order Thalattosauria Merriam, 1904 [64], |
| Superfamily Askeptosauroidea Kuhn-Schnyder, 1971 [65], |
| Family incertae sedis, |
| Genus Anshunsaurus Liu, 1999 [66], |
| Species Anshunsaurus wushaensis Rieppel et al., 2006 [67], |
| Species Anshunsaurus huangnihensis Cheng et al., 2007 [68]. |
Anshunsaurus wushaensis and
A. huangnihensis are medium-sized askeptosauroid thalattosauriforms. Their skeletons are 2–3 m in length as adults, with relatively long necks and tails (
Figure 4i,p). Both species were reported from the Ladinian Zhuganpo Member of the Falang Formation at Wusha Town, although the exact locality and stratigraphic level remains unclear. They were possibly collected from the same Lower Assemblage of the Xingyi Fauna as the nothosaurid species from the same area, but their stratigraphic position and locality remain largely uncertain. We did not recover these particular species from our extensive excavation at Wusha Town. The original description of
A. huangnihensis was very short and brief, and the holotype (YIGM V 30017) needs further preparation. In addition, a detailed comparison to
A. wushaensis is required to clarify whether this species is a synonym of
A. wushaensis. The occurrence of this species of the genus
Anshunsaurus in the Ladinian of the Middle Triassic indicates a connection of the Xingyi Fauna to the Late Triassic Guanling Biota but also demonstrates the paleobiogeographic affinity of the South China Block to western Tethys and phylogenetic affinities with the European
Askeptosaurus [
69].
| |
| Superfamily Thalattosauroidea Nopcsa, 1928 [27], |
| Family incertae sedis, |
| Genus Xinpusaurus Yin in Yin et al., 2000 [55], |
| Species Xinpusaurus xingyiensis Li et al., 2016 [70]. |
Xinpusaurus xingyiensis is a medium to large thalattosauroid thalattosauriform with a relatively short neck, compared to the askeptosauroid thalattosauriform
Anshunsaurus. The skeleton of the holotype (XNGM WS-53-R3) is 2.1 m in preserved length with half the tail missing (
Figure 4q), which was excavated from Bed 53 in the Upper Assemblage of the Xingyi Fauna at Wusha Town. An additional specimen preserved on the top surface of Bed 50 (XNGM WS2011-50-R6) is a 2 m long tail of
X. xingyiensis, probably derived from a large individual of about 4 m in total length. As already mentioned, in the stomach of the large ichthyosaur
Guizhouichthyosaurus specimen from the same bed, some bones of the trunk and limbs from an individual of
X. xingyiensis were identified [
58]. The presence of
Xinpusaurus also indicates a connection of the Ladinian Xingyi Fauna with the somewhat younger Lower Carnian Guanling Biota.
| |
| Species Thalattosauroidea incertae sedis |
A fragmentary specimen (XNGM WS-22-R5) was excavated from Bed 22 in the Lower Assemblage of the Xingyi Fauna at Wusha Town (
Figure 4g). It is only a partial rostrum, but with its anterior half vertically downturned and bearing low-crowned and bulbous teeth. This specimen is different from all other thalattosaurs previously reported from China, resembling
Hescheleria ruebel among Thalattosauroidea, although currently considered as
incertae sedis because of the incompleteness of the single specimen [
71].
| |
| Archosauromorphs: |
| Infraclass Archosauromorpha Huene, 1946 [72], |
| Family Tanystropheidae Camp, 1945 [73], |
| Genus Tanystropheus, |
| Species Tanystropheus hydroides [74]. |
| Genus Macrocnemus Nopcsa, 1930 [75], |
| Species Macronemus fuyuanensis Li et al., 2007 [76]. |
| Genus Fuyuansaurus Fraser et al., 2013 [77], |
| Species Fuyuansaurus acutirostris Fraser et al., 2013 [77]. |
Three tanystropheid species, belonging to three genera, have been reported from the Ladinian Zhuganpo Member of Falang Formation. The specimen GMPKU-P-1527, originally identified as
Tanystropheus cf.
T. longobardicus, was collected around Wusha Town of Xingyi [
78] (
Figure 4n). It is a large tanystropheid with its extraordinarily long neck and cervical ribs being remarkably similar to the larger specimens of
Tanystropheus from the Anisian/Ladinian Besano Formation of Monte San Giorgio in southern Switzerland and northern Italy. The recent recognition of two species of
Tanystropheus in western Tethys, with the larger specimens generally referred to as
T. hydroides, suggests that this specimen might also be referred to
T. hydroides [
74]. The occurrence of this large tanystropheid from the late Ladinian in the Middle Triassic of southwestern China is possibly slightly younger than records from alpine Europe where it was close to the boundary between the Anisian and Ladinian. Its occurrence demonstrates that this strange animal was potentially distributed broadly along the coastline of the western and eastern Tethys during the Middle Triassic [
78].
Both
Macrocnemus fuyuanensis and
Fuyuansaurus acutirostris were small- to medium-sized archosauromorphs (
Figure 4r,s), reported to be found at Jiyangshan, Huabi, Shibalianshan Town, Fuyuan County in Yunnan Province, which is another important fossil site for the Xingyi Fauna. The exact stratigraphic level remains unclear, but the fossil horizons are possibly correlated with the Lower Assemblage of the Xingyi Fauna.
M. fuyuanensis is closely comparable to the European
M. bassanii but can be ascribed to a separate species.
F. acutirostris is a small archosauromorph with a pronounced elongate rostrum, being different from all other tanystropheids. Their occurrences reinforce the close faunal affinity of the eastern and western Tethyan realms during the Middle Triassic, which hints at a common littoral, or at least coastal fauna distributed along the northern coastline of Paleotethys with a diversified archosauromorph fauna [
76,
77,
79].
| |
| Subclass Archosauriformes Gauthier et al., 1988 [80], |
| Order Phytosauria Jaeger 1828 [81], |
| Genus Diandongosuchus Li et al., 2012 [82], |
| Species Diandongosuchus fuyuanensis Li et al., 2012 [82]. |
| Subclass Archosauriformes Gauthier et al. 1988 [80], |
| Genus Litorosuchus Li et al., 2016 [83], |
| Species Litorosuchus somnii Li et al., 2016 [83]. |
Both archosaur taxa were reported to have been found from the Ladinian at Jiyangshan, Huabi, Shibalianshan Town, Fuyuan County in Yunnan Province.
Diandongosuchus fuyuanensis is a small- to medium-sized phytosaur, with a total skeleton length of 1.5 m (
Figure 4b). It contributes to an increasing amount of evidence that early archosauriforms and their close relatives possessed a wide range of ecologies [
81].
D. fuyuanensis preyed on fishes, as inferred from some bones found in the stomach contents [
82].
Litorosuchus somnii is a stem archosaur closely related to the aberrant
Vancleavea (
Figure 4t). It is probably a semiaquatic archosaur [
83].
Figure 4.
Marine reptiles of the Xingyi Fauna. (
a)
Guizhouichthyosaurus sp., XNGM WS-50-R4, scale bar = 40 cm; (
b)
Diandongsuchus fuyuanensis, ZMNH M8770, scale bar = 10 cm [
82]; (
c)
Qianichthyosaurus xingyiensis, (top) holotype, WS2011-46-R1; (bottom) referred specimen, WS2011-50-R7, scale bar = 15 cm [
60]; (
d)
Wangosaurus brevirostris, GMPKU-P-1529, scale bar = 20 cm [
46]; (
e)
Yunguisaurus liae, XNGMXY-2013-R1, scale bar = 25 cm [
56]; (
f)
Keichousaurus hui, XNGM WS-30-R41, scale bar = 10 cm; (
g) Thalattosauroidea indet., XNGM WS-22-R5, scale bar = 2 cm [
76]; (
h)
Glyphoderma kangi, XNGM FY-R01, scale bar = 10 cm [
28]; (
i)
Anshunsaurus huangnihensis, YIGM V 30017, scale bar = 5 cm [
70]; (
j)
Dingxiaosaurus luyinensis, DMK 8, scale bar = 5 cm [
56]; (
k)
Qianxisaurus chajiangensis, NMNS-KIKO-F044630, scale bar = 10 cm [
38]; (
l)
Nothosaurus youngi, XNGM WS-30-R24, scale bar = 20 cm [
41]; (
m)
Brevicaudosaurus jiyangshanensis, IVPP V 18625, scale bar = 5 cm [
45]; (
n)
Tanystropheus hydroides, GMPKU-P-1527, scale bar = 10 cm; (
o)
Lariosaurus xingyiensis, XNGM WS-30-R19, scale bar = 10 cm [
43]; (
p)
Anshunsaurus wushaensis, IVPP V 13782, scale bar = 20 cm [
68]; (
q)
Xinpusaurus xingyiensis, XNGM WS-53-R3, scale bar = 20 cm [
71]; (
r)
Macrocnemus fuyuanensis, GMPKU-P-3001, scale bar = 10 cm [
79]; (
s)
Fuyuansaurus acutirostris, IVPP V 17983, scale bar = 20 mm [
77]; (
t)
Litorosuchus somnii, IVPP V 16978, scale bar = 10 cm [
83].
Figure 4.
Marine reptiles of the Xingyi Fauna. (
a)
Guizhouichthyosaurus sp., XNGM WS-50-R4, scale bar = 40 cm; (
b)
Diandongsuchus fuyuanensis, ZMNH M8770, scale bar = 10 cm [
82]; (
c)
Qianichthyosaurus xingyiensis, (top) holotype, WS2011-46-R1; (bottom) referred specimen, WS2011-50-R7, scale bar = 15 cm [
60]; (
d)
Wangosaurus brevirostris, GMPKU-P-1529, scale bar = 20 cm [
46]; (
e)
Yunguisaurus liae, XNGMXY-2013-R1, scale bar = 25 cm [
56]; (
f)
Keichousaurus hui, XNGM WS-30-R41, scale bar = 10 cm; (
g) Thalattosauroidea indet., XNGM WS-22-R5, scale bar = 2 cm [
76]; (
h)
Glyphoderma kangi, XNGM FY-R01, scale bar = 10 cm [
28]; (
i)
Anshunsaurus huangnihensis, YIGM V 30017, scale bar = 5 cm [
70]; (
j)
Dingxiaosaurus luyinensis, DMK 8, scale bar = 5 cm [
56]; (
k)
Qianxisaurus chajiangensis, NMNS-KIKO-F044630, scale bar = 10 cm [
38]; (
l)
Nothosaurus youngi, XNGM WS-30-R24, scale bar = 20 cm [
41]; (
m)
Brevicaudosaurus jiyangshanensis, IVPP V 18625, scale bar = 5 cm [
45]; (
n)
Tanystropheus hydroides, GMPKU-P-1527, scale bar = 10 cm; (
o)
Lariosaurus xingyiensis, XNGM WS-30-R19, scale bar = 10 cm [
43]; (
p)
Anshunsaurus wushaensis, IVPP V 13782, scale bar = 20 cm [
68]; (
q)
Xinpusaurus xingyiensis, XNGM WS-53-R3, scale bar = 20 cm [
71]; (
r)
Macrocnemus fuyuanensis, GMPKU-P-3001, scale bar = 10 cm [
79]; (
s)
Fuyuansaurus acutirostris, IVPP V 17983, scale bar = 20 mm [
77]; (
t)
Litorosuchus somnii, IVPP V 16978, scale bar = 10 cm [
83].
![Diversity 17 00453 g004]()
4. Fish Diversity of the Xingyi Fauna
Fishes are also highly diverse in the Ladinian Xingyi Fauna; to date, 17 species have been reported (
Figure 5), and a few more are under description. This ichthyofauna is a typical Middle Triassic Fish Fauna (TMFF) [
84] with an abundance of small taxa and with a few new appearances such as flying fishes [
85] and basal teleosts [
86].
Guizhouiscus microlepidus Liu et al., 2003 [
87] is a fairly large paleonisciform fish, usually around 100 cm in length (
Figure 5c). This taxon is of interest, as its body is covered by rather thin ganoid scales, which is quite uncommon for paleonisciforms, where the scales are usually thick. The fusiform shape of this fish indicates an open water dweller, possibly one of the first actinopterygian fishes to achieve such an ecological niche, maybe alongside the larger
Birgeria [
88]. It is worth reiterating that marine reptiles in the Late Ladinian moved from a coastal environment to a much more open sea [
14].
Birgeria liui was first reported from the Ladinian Zhuganpo Member at Changdi Town, Luoping County in Yunnan Province [
84] (
Figure 5i), while at Wusha Town of Xingyi,
Birgeria is recorded in beds 45 and 53 in the Upper Assemblage of the Xingyi Fauna (
Figure 3). It is a large-sized carnivorous fish, considered to be a top predator among Triassic marine fishes, along with
Saurichthys. Ni et al. [
89] described the specimen XNGM WS-45-F21 from Bed 45, which is well-preserved, 76 cm in standard length (SL) and 94 cm in total length (TL), and fairly complete except for the most posterodorsal part of the skull. Contrary to the records for
Saurichthys, the anatomy of
Birgeria is quite uniform over its long-lasting duration, extending across most of the Triassic. Elsewhere
Birgeria is usually much less common than
Saurichthys, and so far, it has not been recorded in any of the other Late Ladinian marine vertebrate assemblages from the Alps, but in the Xingyi Fauna it appears more common than
Saurichthys and especially much larger, so that
Birgeria could be considered the top predator among fishes. As
Birgeria is most likely a suction or filter feeder owing to its very mobile skull [
84], large specimens from the Xingyi Fauna indicate the possibility of swimming in more open waters.
As noted above,
Saurichthys is very rare in the Xingyi Fauna, and the recorded specimens are not large, mostly less than 50 cm in length. A single, almost complete, specimen has been collected from Bed 35, and some other remains were found in Bed 47 (
Figure 2). This yet to be described species appears to be a very slender one, with very small median fins. In common with
Birgeria,
Saurichthys is very rare in other Late Ladinian marine vertebrate assemblages, with but a few remains found from the Kalkschieferzone (Uppermost Meride Limestone, Late Ladinian) of the Monte San Giorgio area [
90] or from the new Pelsa/Vazzoler site in the Dolomites (AT pers. obs. 2016, 2025). The late Ladinian is the time of lowest diversity for
Saurichthys after the acme around the Anisian/Ladinian boundary (Besano Formation in Monte San Giorgio and Luoping Biota), followed by a slight recovery in the Carnian, with a few species in the Alps (
S. striolatus and
S. calcaratus [
91]) and one species from China,
S. taotie [
92], before a late flourish in the Norian of the Alps, with several very large species such as
S.
deperditus attaining at least 170 cm in length (AT pers. obs.).
Thoracopterus wushaensis Tintori et al., 2012 [
85] is the oldest record of the Triassic thoracopterid flying fishes. They span a period of over 20 my until at least the late Norian [
93] (
Figure 5e).
T. wushaensis is known from a few specimens found in situ. The holotype was found in Bed 43 of the Upper Assemblage of the Xingyi Fauna, and other referred specimens originated within beds 42 to 46 and possibly also 47.
Potanichthys xingyiensis Xu et al., 2013 [
94] is a junior synonym of
T.
wushaensis, and its exact locality and stratigraphic level are unknown. Recently, a specimen of
T. wushaensis was recorded from a new Late Ladinian fossiliferous site in the Italian Dolomites [
95], which suggests a wide distribution of this species, clearly able to cross all of Tethys. It also supports the stratigraphic correlation between the Xingyi Fauna and the Pelsa/Vazzoler Fossil Lagerstätte in the Dolomites, where, together with several fish taxa [
96,
97], large amounts of benthonic invertebrates have been collected [
98].
Wushaichthys exquisitus Xu et al., 2015 [
99] (
Figure 5h) is a peltopleurid, very close to
Peltopleurus and
Peripeltopleurus [
100], while Xu et al. [
90] and Shen and Arratia [
101] suggested it was a thoracopterid. The exact locality and stratigraphic level of the type material remain unknown. Indeed, there are many questions regarding this taxon, as underlined by Tintori [
100]. Furthermore, the high variability of peltopleurids must be taken into account as, based on bed-by-bed excavations at Nimaigu in Wusha Town of Xingyi, peltopleurids are recorded from several beds from both the Lower and Upper Assemblages. and this group must be subjected to a more detailed assessment. The presence of very small peltopleurids is common in Middle Triassic fish assemblages, especially in the Ladinian. Small peltopleurids are quite common in Bed 35 but are very rare in the other beds of the Lower Assemblage, while they are common in the Upper Assemblage where the preservation is unfortunately not so nice, and most material from the Nimaigu excavation needs to be properly prepared.
Peltopleurus tyrannos Xu, Ma, and Zhao, 2018 is a larger peltopleurid species [
102] (
Figure 5d). Its precise stratigraphic and geographic origins have not been provided, although it has been considered as belonging to the Xingyi Fauna. This species shows some characteristics that are not strictly typical of peltopleurids, including the preopercular shape, the very strong dentition, the anal fin not strongly dimorphic, and the pectinate posterior margin of the scales, making this species quite close to Perleididae. A revision is needed to assess its actual systematic position.
Habroichthys orientalis (Su, 1959) [
8] (
Figure 5f) is a very common miniature fish, most specimens being around 20 mm in standard length, mainly found in Bed 35 in the Nimaigu excavation, where it can appear as mass mortality surfaces with more than 20 specimens per square meter.
H.
orientalis is more common in several beds at Dingxiao of Xingyi, and this may explain why such a small fish was already described 70 years ago. It was originally described as
Peltopleurus orientalis by Su [
8], while Tintori et al. [
103] considered it to be a species of
Habroichthys. This was based on the presence of the characteristic large semicircular scale at the end of the very high flank scale row that represents one of the autopomorphic characteristics of
Habroichthys as recognized since the erection of this genus by Brough [
104]. A modern revision of this taxon is underway. Until recently,
H. orientalis was the latest species of the genus to be described, but now two additional species have been recorded from the coeval site of Pelsa/Vazzoler in the Dolomites [
97].
Habroichthys spp. cover most of the Middle Triassic on both ends of the Tethys, and it can be considered as one of the smallest fossil fishes, as most species are well under 30 mm as adults [
97].
Luganoia fortuna Xu, 2020 [
105] is a small fish, known only from the holotype (
Figure 5m). Since the detailed origin of the specimen is not clear, it is impossible to state whether this taxon belongs to the Lower or Upper Assemblage of the Xingyi Fauna, although the beautiful preservation of the holotype is more suggestive of the Lower Assemblage. As in the type of area of the genus (Monte San Giorgio), this taxon is quite rare.
Colobodus wushaensis Li et al., 2019 [
106] (
Figure 5g) was recognized based on a specimen which was supposed to be from the ‘Carnian’ at Wusha Town of Xingyi. The original description is very brief, and there is no real comparison with other
Colobodus species. We did not recover any
Colobodus specimens from the Late Ladinian Xingyi Fauna during the bed-to-bed excavations at Wusha Town, but specimens from the Lower Carnian of the Guanling area are known (AT pers. obs.). Therefore, at present, we do not consider
Colobodus to be a component of the Xingyi Fauna. Liu et al. [
107] described a large fish, up to 100 cm long, from the Wayao Member of the Falang Formation (Carnian) at Xiaowa of Xinpu near Guanling, referring it to the new species
Guizhoueugnatus largus Liu et al., 2006, but we suggest that it can be better ascribed to
Colobodus due to the scale ornamentation. Moreover, this ornamentation is also very similar to that shown by Li et al. [
106] for
C. wushaensis, so that the two species could be synonymous, with
C. wushaensis becoming a junior synonym of
Colobodus largus [
107].
Among neopterygians,
Asialepidotus shingyiensis Su, 1959 [
8] (
Figure 5o) is the most abundant and well-preserved taxon from the Lower Assemblage of the Xingyi Fauna. It was restudied at the beginning of this century when new discoveries across southern China boosted research on the Triassic marine vertebrates [
108]. Liu et al. [
87] described the new genus
Guizhouella, which was then changed to
Guizhoueugnathus by Liu [
109], while Tintori et al. [
84] considered it as a junior synonym of
Asialepidotus (see also [
110]).
Asialepidotus is mostly concentrated in a single bed at the Nimaigu site: Bed 35 in the Lower Assemblage of the Xingyi Fauna, where ontogenetic series occur. It is much rarer in the beds below Bed 35, as are all fishes, where the faunal composition is dominated by marine reptiles, most notably
Keichousaurus. The remains of
Asialepidotus, mainly scales, are also very common in the large 3D coprolites recorded from Bed 35, but they are absent in similar coprolites from the lower beds, where there are plenty of bones of
Keichousaurus (AT, pers. obs.).
Asialepidotus can therefore be considered an index taxon for the Late Ladinian Lower Assemblage of the Xingyi Fauna. Despite its name,
Asialepidotus does not belong to Ginglymodi but to Halecomorphi, as first proposed by Tintori et al. [
84], although Liu et al. [
87] considered
Guizhouella (then changed to
Guizhoueugnatus [
109]) to belong within the Amiiformes, not realizing the close resemblance to
Asialepidotus.
Asialepidotus was considered in the family Panxianichthyidae (Panxianichthyiformes) [
110], a clade comprising several Triassic halecomorphs more advanced than Parasemionotiformes, which was the neopterigyan dominant group in Triassic Early Fish Fauna (TEFF in [
84]). Xu and Ma [
108] placed
Asialepidotus in Ionoscopiformes. However, the extension of this Late Jurassic/Early Cretaceous order down to the Middle Triassic, with a gap of about 100 my, has been shown to be untenable [
110]. Ionoscopiformes exhibit solid vertebral centra and elasmoid scales [
111], while
Asialepidotus has ganoid scales and a notochord that is still totally unconstricted.
Sinoeugnathus kueichowensis Su, 1959 [
8] (
Figure 5j) is also an halecomorph recently redescribed by Feng et al. [
112] based on specimens without any detailed stratigraphic information. As mentioned for the previous taxon, the referral of this genus to Ionoscopiformes is questionable. This taxon is smaller than
Asialepidotus in body size but is less well known. Based on the published material, its actual ratio against the other taxa cannot be established, and it was not collected from scientific excavations.
The basal teleosts
Malingichthys nimaiguensis Tintori et al., 2015 [
113] and
Malingichthys wanfenglinenisi Tintori et al. 2015 [
113] (
Figure 5a,b) are the oldest representatives of Pholidophoriformes. They indicate that the origin of the largest modern vertebrate group, the Teleostei, is in the Late Ladinian, earlier than previously considered based on the Early Carnian
Knerichthys [
114] from Raibl/Cave del Predil in northern Italy. At Raibl/Cave del Predil, the fish assemblage also yields
Thoracopterus niederristi Bronn, 1858 [
115], which is the first described flying fish and was geologically the oldest flying fish known prior to the discovery of
T. wushaensis.
Malingichthys is very common in Bed 35 of the Lower Assemblage of the Xingyi Fauna, but it could be also present in the Upper Assemblage, although there, the preservation of these small fishes is rather poor, and most material is still waiting to be prepared. Records of Late Ladinian basal teleosts are not known in the Kalkschieferzone at Monte San Giorgio [
116] and are very rare in the new Pelsa/Vazzoler Lagerstätte in the Dolomites (AT & DC pers. obs.). There is also a relative scarcity of Pholidophoridae in the Early Carnian Raibl/Cave del Predil and Polzberg assemblages ([
114], AT pers. obs.).
Marcopoloichthys sp. is apparently restricted to the Upper Assemblage of the Xingyi Fauna, since its first appearance is in Bed 39 and it is more common in Bed 47. This genus is a very specialized taxon that shows a long time span from the Anisian to the Norian ([
117,
118,
119], AT pers. obs), as well as a wide geographic distribution across Tethys, as with many other Triassic fish genera. In the Late Ladinian,
Marcopoloichthys is very common in the Pelsa/Vazzoler Lagerstätte in the Dolomites [
96,
97], but is missing in the Kalkschieferzone at Monte San Giorgio, where a similarly advanced neopterygian,
Prohalecites porroi (Bellotti, 1857) [
120], is the most common taxon [
116]. Such differences are probably related to paleoenvironmental conditions. The Xingyi Fauna lived in a quite large basin, while the Pelsa/Vazzoler fauna was from a small intraplatform basin/lagoon inside a large but isolated carbonate platform [
98], and the Monte San Giorgio fauna occurred in a complex coastal lagoon with periodic freshwater influxes [
116].
Brachysomus minor Liu et al., 2003 [
87] (
Figure 5p) was described as a new genus and species based on a single poorly preserved specimen. Liu [
109] changed the generic name to
Guizhoubrachysomus, because the original one was already occupied.
Guizhoubrachysomus minor (Liu et al., 2003) [
87,
109] is quite peculiar and is characterized by a deep body shape, an anteriorly placed dorsal fin, strong fringing fulcra on all fins, and an abbreviated heterocercal tail. This suggests it is a neopterygian not related to ‘subholosteans’ such as Luganoiformes or Perleidiformes, although the body shape is similar to some other Triassic fishes such as
Dipteronotus [
121] from the Ladinian of Monte San Giorgio.
Guizhouamia bellula Liu et al., 2002 [
122] (
Figure 5p) and
Xingyia gracilis Liu et al., 2003 [
87] (
Figure 5k) are both small-sized fishes with elongate dorsal fins.
Guizhouamia is naked, but
Xingyia is completely covered by ganoid scales. Liu et al. [
87] referred both taxa to Amiiformes, but a detailed description and discussion of the systematics is needed. They are not very common:
G.
bellula has been recorded in Bed 35 in the Lower Assemblage of the Xingyi Fauna, and this is probably also true for
X.
gracilis.
Figure 5.
Fishes of the Xingyi Fauna. (
a)
Malingichthys nimaiguensis, GMPKU-P-3162, 3163, scale bar = 5 mm [
113]; (
b)
Malingichthys wanfenglinensis, GMPKU-P-3181, scale bar = 5 mm [
113]; (
c)
Guizhouiscus microlepidus, Fish No. 1 in Guizhou Stone Culture Palace, scale bar = 120 mm [
87]; (
d)
Peltopleurus tyrannos, IVPP V 19986, scale bar = 16 mm [
102]; (
e)
Thoracopterus wushaensis, GMPKU-P-3061, scale bar = 10 mm [
85]; (
f)
Habroichthys orientalis, V. 2432, scale bar = 6 mm [
8]; (
g)
Colobodus wushaensis, GZ12203845, scale bar = 50 mm [
106]; (
h)
Wushaichthys exquisitus, IVPP V19959, scale bar = 10 mm [
99]; (
i)
Birgeria liui, IVPP V 12569, scale bar = 50 mm [
88,
89]; (
j)
Sinoeugnathus kueichowensis, V. 2433, scale bar = 20 mm [
8]; (
k)
Xingyia gracilis, Fish No. 1 in
Keichousaurus Museum, Dingxiao, scale bar = 20 mm [
87]; (
l)
Guizhoueugnathus analilepida, NIGP136040, scale bar = 60 mm [
87,
109]; (
m)
Luganoia fortuna, IVPP V 25693, scale bar = 10 mm [
105]; (
n)
Guizhouamia bellula, NIGP134837, scale bar = 20 mm [
122]; (
o)
Asialepidotus shingyiensis, V. 2434, scale bar = 40 mm [
8]; (
p)
Guizhoubrachysomus minor, NIGP136039, scale bar = 8 mm [
87,
109].
Figure 5.
Fishes of the Xingyi Fauna. (
a)
Malingichthys nimaiguensis, GMPKU-P-3162, 3163, scale bar = 5 mm [
113]; (
b)
Malingichthys wanfenglinensis, GMPKU-P-3181, scale bar = 5 mm [
113]; (
c)
Guizhouiscus microlepidus, Fish No. 1 in Guizhou Stone Culture Palace, scale bar = 120 mm [
87]; (
d)
Peltopleurus tyrannos, IVPP V 19986, scale bar = 16 mm [
102]; (
e)
Thoracopterus wushaensis, GMPKU-P-3061, scale bar = 10 mm [
85]; (
f)
Habroichthys orientalis, V. 2432, scale bar = 6 mm [
8]; (
g)
Colobodus wushaensis, GZ12203845, scale bar = 50 mm [
106]; (
h)
Wushaichthys exquisitus, IVPP V19959, scale bar = 10 mm [
99]; (
i)
Birgeria liui, IVPP V 12569, scale bar = 50 mm [
88,
89]; (
j)
Sinoeugnathus kueichowensis, V. 2433, scale bar = 20 mm [
8]; (
k)
Xingyia gracilis, Fish No. 1 in
Keichousaurus Museum, Dingxiao, scale bar = 20 mm [
87]; (
l)
Guizhoueugnathus analilepida, NIGP136040, scale bar = 60 mm [
87,
109]; (
m)
Luganoia fortuna, IVPP V 25693, scale bar = 10 mm [
105]; (
n)
Guizhouamia bellula, NIGP134837, scale bar = 20 mm [
122]; (
o)
Asialepidotus shingyiensis, V. 2434, scale bar = 40 mm [
8]; (
p)
Guizhoubrachysomus minor, NIGP136039, scale bar = 8 mm [
87,
109].
![Diversity 17 00453 g005]()
There are abundant micro-remains of sharks in the sequence above the fossiliferous level of the Xingyi Fauna, but in the main beds, only very rare shark micro-teeth were recovered, although a few large and complete skeletons have been collected (AT, pers. obs. 2018), which is quite peculiar for the Xingyi Fauna. The rarity of shark micro-remains is shared in common with other Middle–Late Triassic marine vertebrate faunas such as the Panxian Fauna, the Luoping Fauna, and the Guanling Biota, as well as in the Alpine Middle Triassic sites. There are a few shark micro-remains reported from the Besano Formation at Monte San Giorgio [
123], and a few
Palaeobates teeth, together with a partial body and dorsal fin spine, have been collected in the new site of Pelsa/Vazoller.
6. Establishment of the Modern-Type Marine Ecosystem Indicated by the Xingyi Fauna
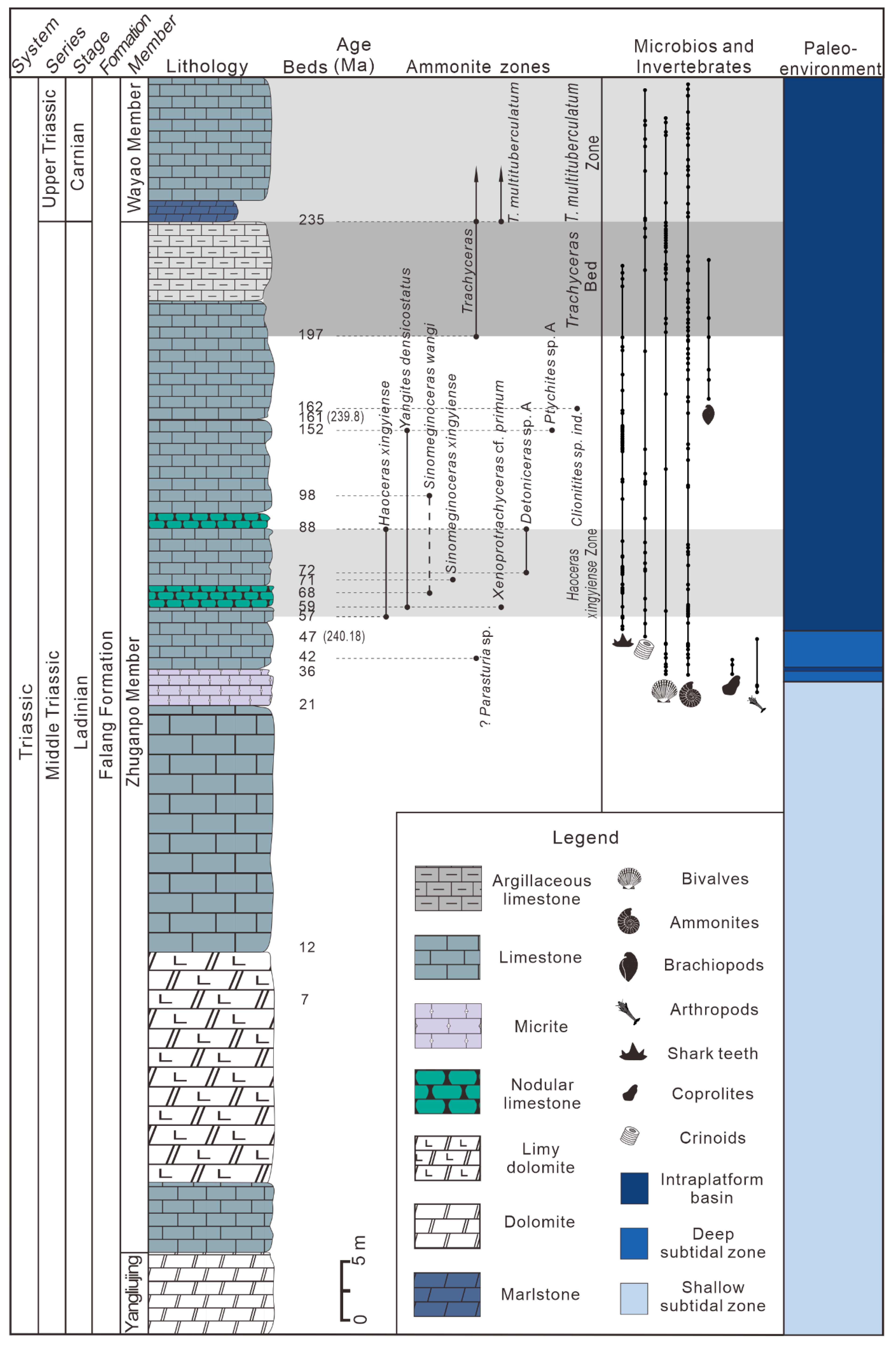
The 6.61 m thick fossiliferous beds of the Xingyi Fauna can be subdivided into two main parts containing two separate marine vertebrate assemblages, while the lithofacies analysis supports a paleoenvironmental change to a shallow or deep subtidal to intraplatform basin from a restricted to more open carbonate platform [
13] (
Figure 2 and
Figure 3). In the Lower Assemblage, small- to medium-sized sauropterygians were more common, including one species of cyamodontoid, two species of pachypleurosaurids, and three species of nothosaurids. Medium-sized thalattosauriforms, including two species of askeptosauroid and an unnamed thalattosauroid with a bent rostrum, also occur in this assemblage. Additionally, three species of tanystropheid and two species of archosaur were probably recovered from the Lower Assemblage, but no ichthyopterygian has been found in this subtidal assemblage. The Fusea assemblage (Tolmezzo, Friuli, Italy) at the Ladinian/Carnian boundary is probably the stratigraphically closest marine fossil assemblage in the Alps, in which most fossils comprise disarticulated remains including large fragments of the dorsal shield of placodonts, bones of nothosaurs, cervical vertebra of
Tanystropheus, a few small isolated teeth of archosauriforms, and lungfish remains [
130,
131,
132]. Sauropterygians (the placodont
Cyamodus sp. and the nothosaur
Nothosaurus cf.
N.
giganteus) are the most common remains, while ichthyosaurs are completely missing, as in the Lower Assemblage of the Xingyi Fauna. This vertebrate assemblage probably represents a coastal paleoenvironment, with freshwater influx. The composition of the marine reptiles in the Lower Assemblage of the Xingyi Fauna can be closely compared to that of the Late Anisian/Early Ladinian Besano Formation at Monte San Giorgio, with common genera including the nothosaurid sauropterygians
Nothosaurus and
Lariosaurus, the archosauromorphs
Tanystropheus and
Macronemus, and the askeptosauroid thalattosauriforms
Anshunsaurus and
Askeptosaurus. It shows a strong geographic affinity to western Tethys in the Anisian to Ladinian [
15].
In the Lower Assemblage of the Xingyi Fauna, abundant and diversified fishes were found. There are more than 10 species, and most are small, including
Asialepidotus shingyiensis,
Sinoeugnathus kueichowensis,
Habroichthys orientalis,
Malingichthys nimaiguensis, and
Malingichthys wanfenglinensis, as well as
Guizhouiscus microlepidus,
Guizhoubrachysomus minor,
Guizhouamia bellula,
Xingyia gracilis, and
Guizhoueugnathus analilepida (
Figure 3 and
Figure 5). There are some ammonoids, bivalves, gastropods, and crustaceans associated in this assemblage and quite abundant coprolites yielding
Keichousaurus bones (
Figure 2). Comparing fishes across Tethys can be more informative, as the Late Ladinian sites in the Alps yield large numbers of fish taxa, but very rare marine reptiles, and usually limited to
Lariosaurus [
133,
134]. This is very different from the situation in the Lower Assemblage of the Xingyi Fauna. The Kalkschiferzone (Upper Member of Meride Limestone) at Monte San Giorgio yields a peculiar fish assemblage dominated by the miniature advanced neopterygian
Prohalecites porroi but also abundant peltopleurids [
135,
136] [AT pers obs], as in most Middle Triassic assemblages [
84]. It should be emphasized that, apart from
Peltopleurus and the rare
Saurichthys, there are no other genera in common between the fishes of the Xingyi Fauna and the Kalkschieferzone. In the latter, very few specimens of halecomorphs have been found, namely
Allolepidotus [
137], while at Xingyi there are a number of halecomorphs, including the very common
Asialepidotus,
Sinoeugnathus, and possibly
Guizhouamia and
Xingyia. By contrast, Gynglimodi and
Sangiorgioichthys spp. are present in the Kalkschieferzone [
116,
138], while Gynglimodi are totally missing in the Xingyi Fauna. The basal teleosts
Malingichthys spp. and
Marcopoloichthys are common in the Xingyi Fauna but missing in the coeval Monte San Giorgio Fauna, in which
Prohalecites is common. Such major differences are probably related to the different paleoenvironmental conditions. It was a strictly marine environment at Xingyi, while the Kalkschieferzone shows a high variability from ‘fresh water’ to brackish and to marine environments in uneven fluctuations [
116]. However, in the newly discovered Late Ladinian site in the Dolomites (NE Italy) [
96],
Habroichthys [
97],
Marcopolichthys,
Peltopleurus, halecomorphs, and possibly basal teleosts are present and appearing more similar to those at Xingyi. The flying fish
Thoracopterus wushaensis also appears in the Dolomites, which further supports the wide distribution of this marine vertebrate all over Tethys and beyond.
In the Upper Assemblage, the faunal composition exhibited a marked change. The small- to medium-sized cyamodontoid and nothosaurid sauropterygians, askeptosauroid thalattosauriforms, tanystropheids, and archosaurs disappeared, while medium- to large-sized ichthyopterygians, pistosauroid sauropterygians, and thalattosauroid thalattosauriforms appeared, associated with a few small pachypleurosaurid sauropterygians, as well as diverse fishes including the large-sized predator
Birgeria, the oldest species of the flying fish
Thoracopterus, abundant
Marcopoloichthys, and several small peltopleuriforms (
Figure 3) [
84]. Abundant ammonoids, bivalves such as
Halobia, crinoids, crustaceans, and also coprolites and woods have been found in this assemblage. This assemblage appears more similar to the Early Carnian Guanling Biota in Guizhou, with the same genera including the shastasaurid ichthyopterygian
Guizhouichthyosaurus and the thalattosauroid thalattosauriform
Xinpusaurus. In turn, this is similar to the North American California Fauna, as
Guizhouichthyosaurus and
Xinpusaurus are phylogenetically close to
Shastasaurus and
Thalattosaurus, respectively. It shows a paleobiogeographic affinity to east Panthalassa during the late Ladinian to the Carnian. This suggests a transition in paleobiogeographic affinity from west Tethys, which dominated most of the Middle Triassic, to east Panthalassa from the late Ladinian through to the Carnian. The South China Block probably was the paleobiogeological exchange center for Triassic biota migrating from western Tethys to east Pathalassa, with new clades also emerging [
15]. Further complicating the picture are the Carnian sites in the Alps such as Raibl/Cave del Predil in Italy and Polzberg/Lunz in Austria with marine vertebrate assemblages mostly rich in fishes [
139,
140] and only rare reptile remains [
132,
141].
The shastasaurid ichthyopterygian
Guizhouichthyosaurus, pistosauroid sauropterygian
Yunguisaurus, and thalattosauroid thalattosauriform
Xinpusaurus xingyiensis are particularly large marine reptiles (
Figure 4 and
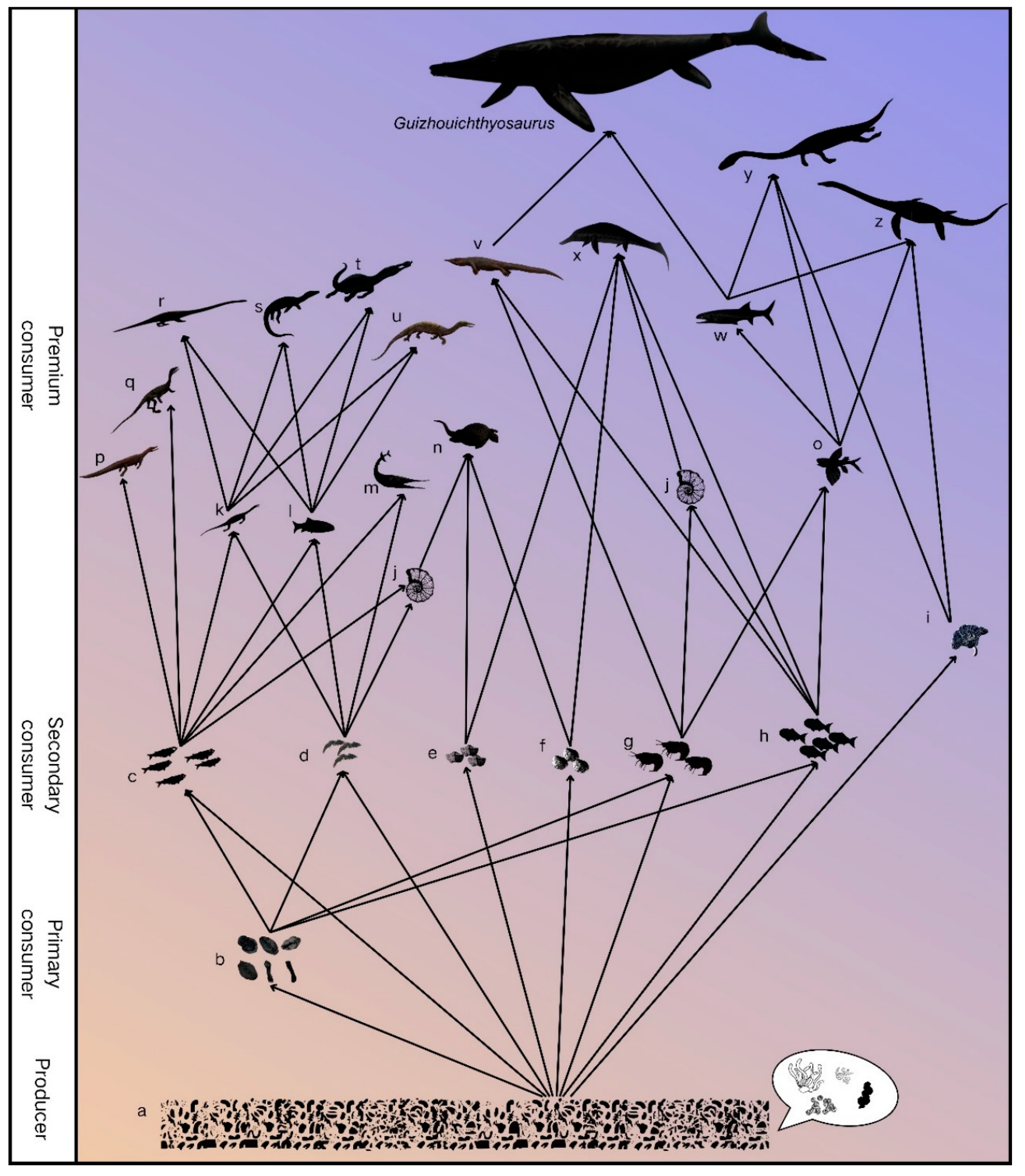
Figure 6). The largest individuals may reach 9 m or more in total length, which suggests that diverse large species of marine reptiles appeared in the late Ladinian marine ecosystem, after the invasion of the seas by Mesozoic reptiles in the Early Triassic.
Birgeria is also a large fish. These large species became apex predators that would have been capable of feeding upon other very large reptiles and fishes [
15,
61] (
Figure 6).
The late Ladinian Xingyi Fauna records the shift from a coastal assemblage to an oceanic assemblage. It also documents the appearance of very large species of marine reptiles and fishes together with flying fishes, which then migrated globally in open oceanic waters. This new marine ecosystem established in the late Ladinian, was dominated by the large air-breathing tetrapods (
Figure 6), and a “modern-type” marine ecosystem. Considering that several invertebrate groups also experienced significant diversification at the same time, the Late Ladinian can be regarded as one of the most important intervals following the end-Permian mass extinction.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}