
Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
- Subadult snails reared with shrimps: Each experimental tank was stocked with 40 P. canaliculate subadults (shell width of ~2 cm) along with 8 M. rosenbergii; to mitigate against damage caused by aggressive interactions, only female shrimps were selected;
- Subadult snails reared without shrimps: Each experimental tank was stocked with 40 P. canaliculate subadults (shell width of ~2 cm) and without the addition of M. rosenbergii;
- Recently hatched juvenile snails reared with shrimps: 10 egg masses (~20 g each) of P. canaliculate were evenly distributed on a wire mesh (2 mm aperture, which allowed newly hatched juveniles to pass through but prevented the passage of subadults) positioned at the top of each tank, and tank was stocked with 8 M. rosenbergii;
- Recently hatched juvenile snails reared without shrimp: Similar to the above treatment, 10 egg masses (~20 g each) of P. canaliculate were evenly distributed on the wire mesh on the top of each tank but without the presence of M. rosenbergii;
- Snail-absent control with only shrimps: The experimental tank contained only 8 M. rosenbergii.
2.3. Data Collection and Analysis
3. Results
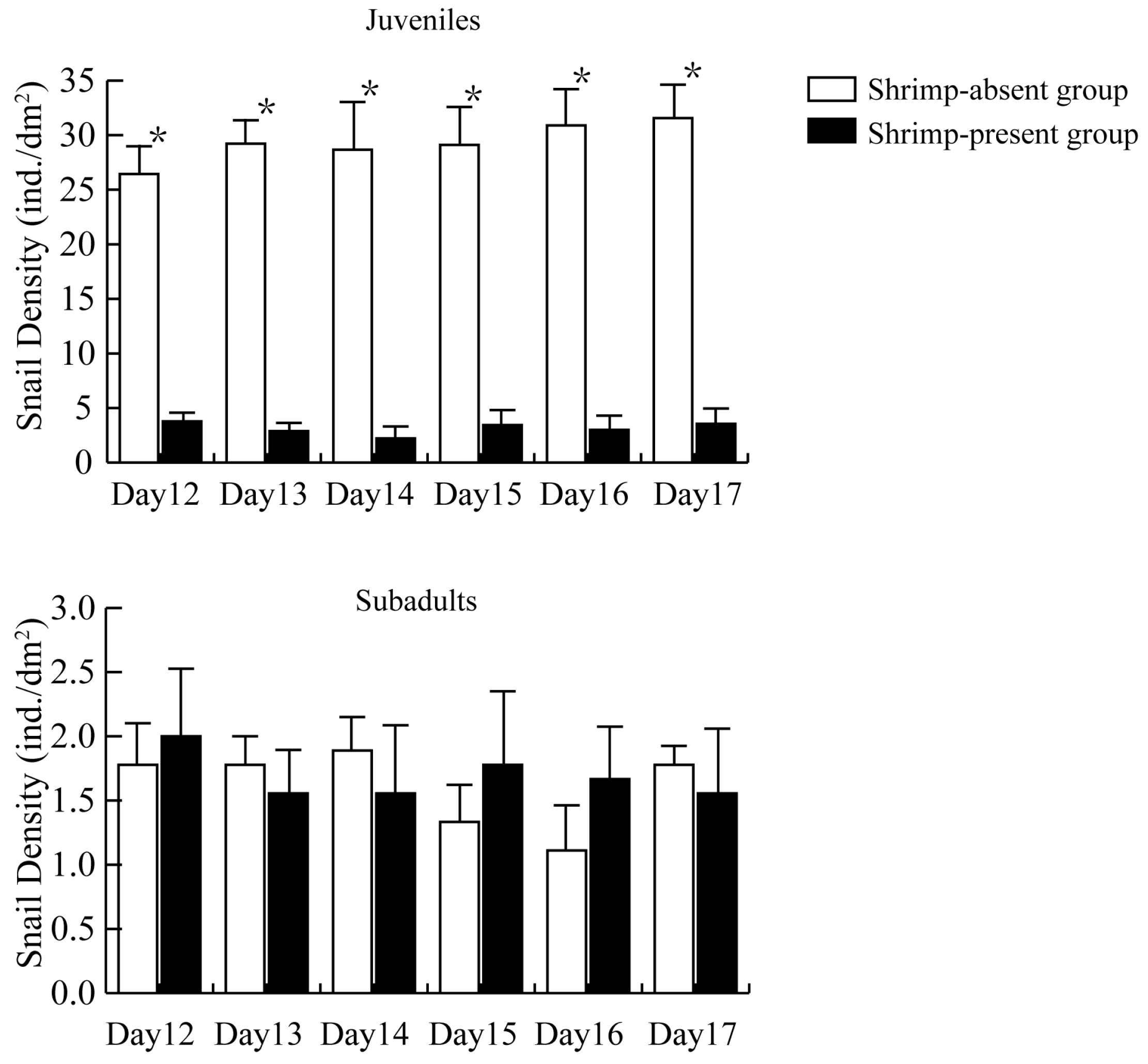
3.1. Impact of M. rosenbergii on the Quantity of P. canaliculata
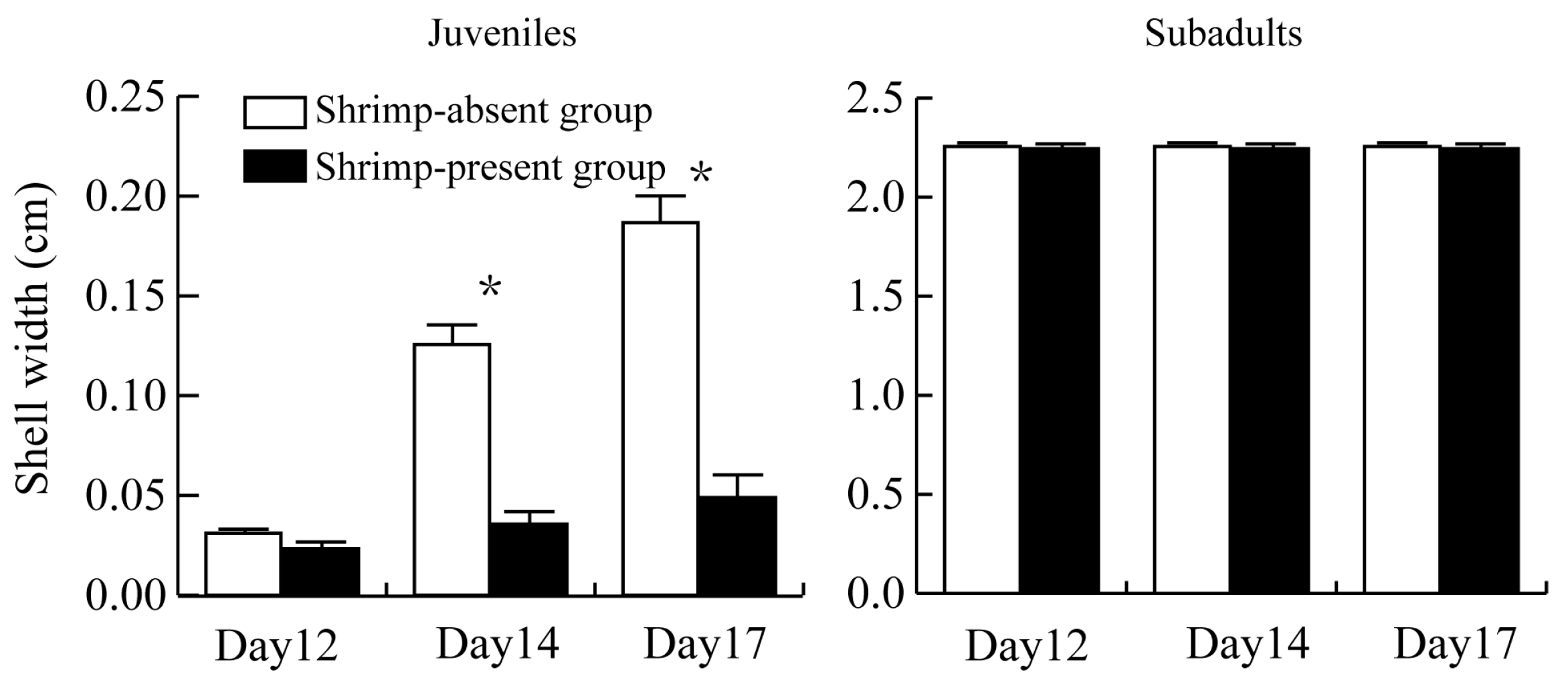
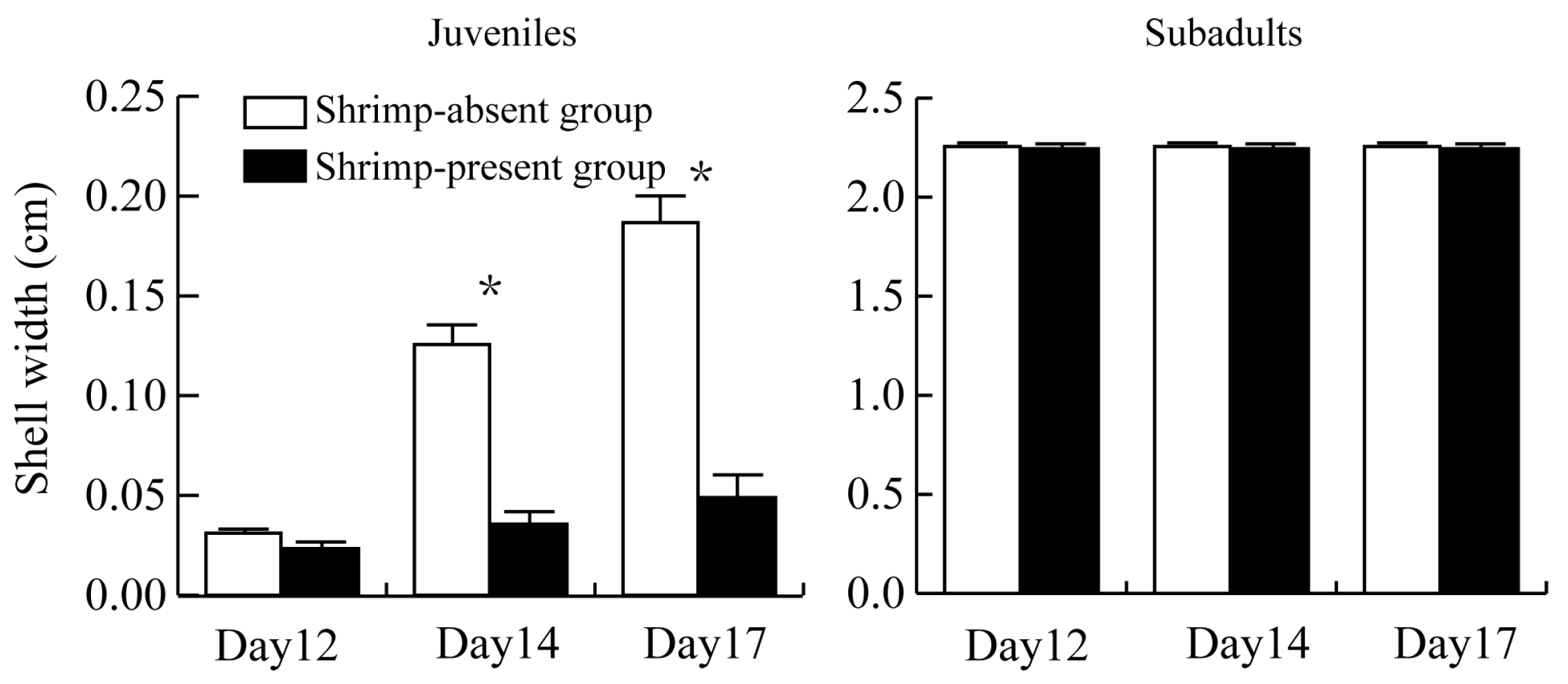
3.2. Size of P. canaliculata Surviving in the Presence of M. rosenbergii
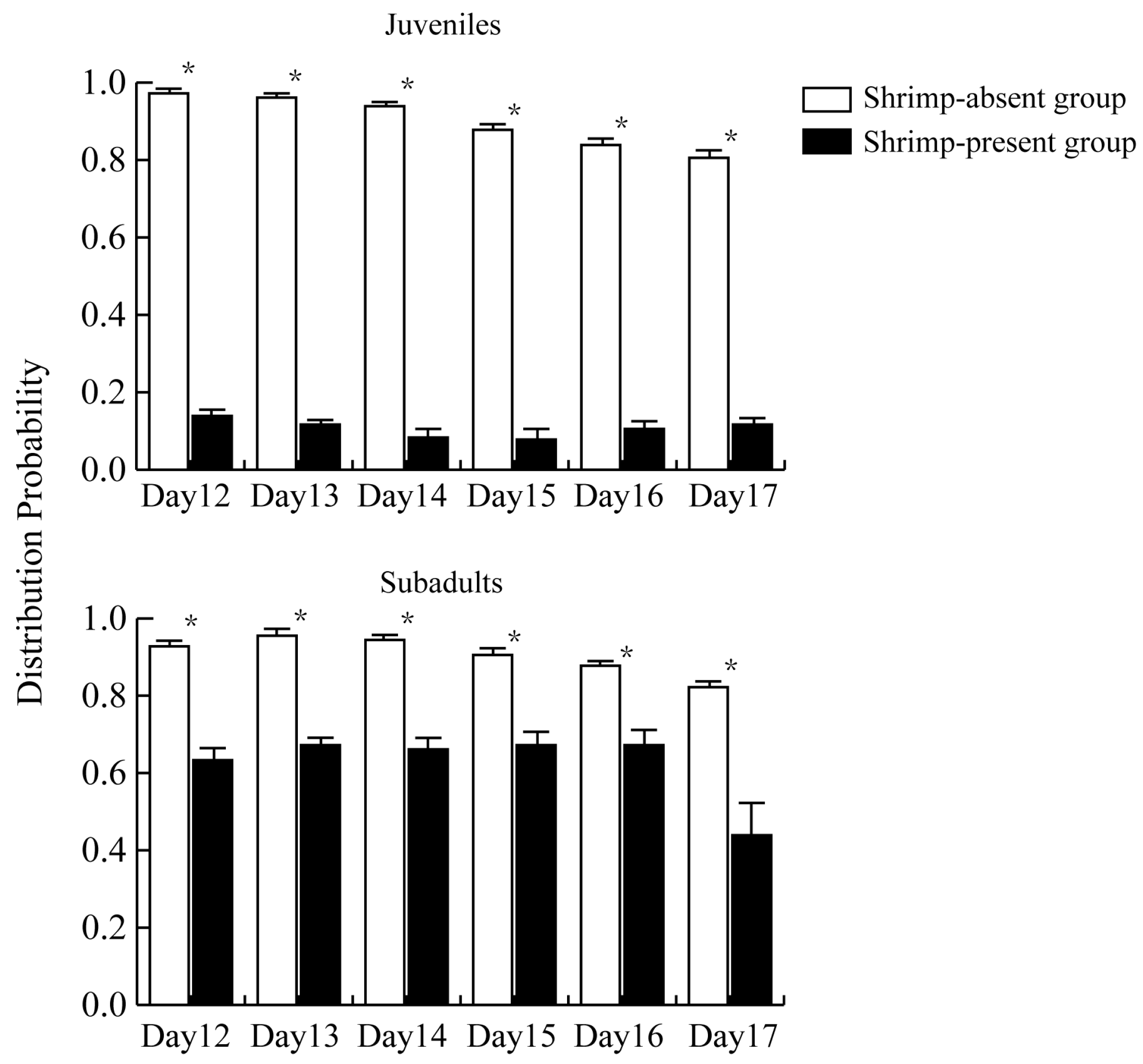
3.3. Influence of M. rosenbergii on the Activity of P. canaliculata
3.4. M. rosenbergii Survival and Size Changes and Estimation of their Control Efficiency on P. canaliculata
4. Discussion
4.1. Effectiveness of M. rosenbergii in Controlling P. canaliculata
4.2. Potential Application of M. rosenbergii for Controlling P. canaliculata Invasion
4.3. Potential Risks of Using M. rosenbergii for Biocontrol of P. canaliculata
- Consider its deployment in isolated water bodies, artificial wetlands, or rice paddies, avoiding introduction into natural water habitats;
- Employ physiologically stable M. rosenbergii juveniles for release to minimize the introduction of individuals with high variability. The utilization of sex-biased seedings of M. rosenbergii might to ensure biological security. Simultaneously, the potential for hybridization between M. rosenbergii and indigenous freshwater prawn species should also be investigated to prevent hybridization-induced population dispersion [57];
- Capture M. rosenbergii from the release areas during autumn and winter, subjecting them to dry ponds or sun-drying treatments.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hayes, K.A.; Joshi, R.C.; Thiengo, S.C.; Cowie, R.H. Out of South America: Multiple Origins of Non-native Apple Snails in Asia. Divers. Distrib. 2008, 14, 701–712. [Google Scholar] [CrossRef]
- Joshi, R.C.; Parera, X.V. The Rice Apple Snail in Spain: A Review. Int. Pest Control 2017, 59, 106. [Google Scholar]
- Horgan, F.G.; Stuart, A.M.; Kudavidanage, E.P. Impact of Invasive Apple Snails on the Functioning and Services of Natural and Managed Wetlands. Acta Oecologica 2014, 54, 90–100. [Google Scholar] [CrossRef]
- De Brito, F.C.; Joshi, R.C. The Golden Apple Snail Pomacea Canaliculata: A Review on Invasion, Dispersion and Control. Outlooks Pest Manage. 2016, 27, 157–163. [Google Scholar] [CrossRef]
- Byers, J.E.; Mcdowell, W.G.; Dodd, S.R.; Haynie, R.S.; Pintor, L.M.; Wilde, S.B. Climate and pH Predict the Potential Range of the Invasive Apple Snail (Pomacea insularum) in the Southeastern United States. PLoS ONE 2013, 8, e56812. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yang, Q.; Xu, Y. Golden Apple Snails. In Biological Invasions and Its Management in China: Volume 2; Wan, F., Jiang, M., Zhan, A., Eds.; Springer: Singapore, Singapore, 2017; pp. 33–47. ISBN 978-981-10-3427-5. [Google Scholar]
- Nghiem, L.T.P.; Soliman, T.; Yeo, D.C.J.; Tan, H.T.W.; Evans, T.A.; Mumford, J.D.; Keller, R.P.; Baker, R.H.A.; Corlett, R.T.; Carrasco, L.R. Economic and Environmental Impacts of Harmful Non-Indigenous Species in Southeast Asia. PLoS ONE 2013, 8, e71255. [Google Scholar] [CrossRef]
- Dumidae, A.; Janthu, P.; Subkrasae, C.; Polseela, R.; Mangkit, B.; Thanwisai, A.; Vitta, A. Population Genetics Analysis of a Pomacea Snail (Gastropoda: Ampullariidae) in Thailand and its Low Infection by Angiostrongylus cantonensis. Zool. Stud. 2021, 60, e31. [Google Scholar] [CrossRef]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the World’s Worst Invasive Alien Species. Biol. Invasions. 2014, 16, 981–985. [Google Scholar] [CrossRef]
- Yang, Q.; Yu, X. A New Species of Apple Snail in the Genus Pomacea (Gastropoda: Caenogastropoda: Ampullariidae). Zool. Stud. 2019, 58, e13. [Google Scholar] [CrossRef]
- Yahaya, H.; Badrulhadza, A.; Sivapragasam, A.; Nordin, M.; Muhamad Hisham, M.N.; Mirudin, H. Invasive apple snails in Malaysia. In Biology and Management of Invasive Apple Snails, 2nd ed.; Joshi, R.C., Cowie, R.H., Sebastian, L.S., Eds.; Philippine Rice Research Institute (PhilRice): Nueva Ecija, Philippine, 2017; pp. 169–195. [Google Scholar]
- Xu, Y.; Li, A.J.; Li, K.; Qin, J.; Li, H. Effects of Glyphosate-based Herbicides on Survival, Development and Growth of Invasive Snail (Pomacea canaliculata). Aquat. Toxicol. 2017, 193, 136–143. [Google Scholar] [CrossRef]
- Duong, H.V.; Nguyen, T.C.; Nguyen, X.T.; Nguyen, M.Q.; Nguyen, P.H.; Vo, T.T. Evaluating the Presence of Pesticide Residues in Organic Rice Production in An Giang Province, Vietnam. J. Sustain. Develop. 2022, 15, 49. [Google Scholar] [CrossRef]
- Plan, M.R.R.; Saska, I.; Cagauan, A.G.; Craik, D.J. Backbone Cyclised Peptides from Plants Show Molluscicidal Activity against the Rice Pest Pomacea canaliculata (Golden Apple Snail). J. Agric. Food. Chem. 2008, 56, 5237–5241. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Wang, W.; Dong, X.; Hu, R.; Nan, X. Molluscicidal Activity of Cardiac Glycosides from Nerium indicum Against Pomacea canaliculata and Its Implications for the Mechanisms of Toxicity. Environ. Toxicol. Pharmacol. 2011, 32, 226–232. [Google Scholar] [CrossRef]
- Ding, W.; Huang, R.; Zhou, Z.; He, H.; Li, Y. Ambrosia artemisiifolia as a Potential Resource for Management of Golden Apple Snails, Pomacea canaliculata (Lamarck). Pest Manag. Sci. 2018, 74, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Brito, F.C.D.; Gosmann, G.; Oliveira, G.T. Extracts of the Unripe Fruit of Ilex paraguariensis as a Potential Chemical Control against the Golden Apple Snail Pomacea canaliculata (Gastropoda, Ampullariidae). Nat. Prod. Res. 2019, 33, 2379–2382. [Google Scholar] [CrossRef] [PubMed]
- Noorshilawati, A.A.; Suraya, A.N.; Rossiyah, S.S. Molluscicidal Activity of Ipomoea batatas Leaf Extracts against Pomacea canaliculata (Golden Apple Snail). Food Res. 2020, 4, 131–137. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, S.; Zeng, J.; Chen, Y.; Guo, Y.; Liu, J.; He, A. Molluscicidal Activity of Nicotiana tabacum Extracts on the Invasive Snail Pomacea canaliculata. Sci. Rep. 2023, 13, 11597. [Google Scholar] [CrossRef]
- Culliney, T.W. Benefits of Classical Biological Control for Managing Invasive Plants. Crit. Rev. Plant Sci. 2005, 24, 131–150. [Google Scholar] [CrossRef]
- Azmi, W.A.; Khoo, S.C.; Ng, L.C.; Baharuddin, N.; Aziz, A.A.; Ma, N.L. The Current Trend in Biological Control Approaches in the Mitigation of Golden Apple Snail Pomacea spp. Biol. Control. 2022, 175, 105060. [Google Scholar] [CrossRef]
- Dong, S.; Zheng, G.; Yu, X.; Fu, C. Biological Control of Golden Apple Snail, Pomacea canaliculata by Chinese Soft-shelled Turtle, Pelodiscus sinensis in the Wild Rice, Zizania latifolia Field. Sci. Agric. 2012, 69, 142–146. [Google Scholar] [CrossRef]
- Yam, R.; Fan, Y.; Wang, T. Importance of Macrophyte Quality in Determining Life-History Traits of the Apple Snails Pomacea canaliculata: Implications for Bottom-Up Management of an Invasive Herbivorous Pest in Constructed Wetlands. Int. J. Environ. Res. Public Health 2016, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Yusa, Y.; Sugiura, N.; Wada, T. Predatory Potential of Freshwater Animals on an Invasive Agricultural Pest, the Apple Snail Pomacea canaliculata (Gastropoda: Ampullariidae), in Southern Japan. Biol. Invasions 2006, 8, 137–147. [Google Scholar] [CrossRef]
- Horgan, F.G. Ecology and Management of Apple Snails in Rice. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 393–417. ISBN 978-3-319-47516-5. [Google Scholar]
- Ip, K.K.L.; Liang, Y.; Lin, L.; Wu, H.; Xue, J.; Qiu, J. Biological Control of Invasive Apple Snails by Two Species of Carp: Effects on Non-target Species Matter. Biol. Control 2014, 71, 16–22. [Google Scholar] [CrossRef]
- Gilioli, G.; Pasquali, S.; Martín, P.R.; Carlsson, N.; Mariani, L. A Temperature-dependent Physiologically Based Model for the Invasive Apple Snail Pomacea canaliculata. Int. J. Biometeorol. 2017, 61, 1899–1911. [Google Scholar] [CrossRef]
- Constantine, K.L.; Makale, F.; Mugambi, I.; Chacha, D.; Rware, H.; Muvea, A.; Kipngetich, V.K.; Tambo, J.; Ogunmodede, A.; Djeddour, D.; et al. Assessment of the Socio-economic Impacts Associated with the Arrival of Apple Snail (Pomacea canaliculata) in Mwea Irrigation Scheme, Kenya. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef]
- Bertolero, A.; Navarro, J. A Native Bird as a Predator for the Invasive Apple Snail, a Novel Rice Field Invader in Europe. Aquatic Conserv: Mar Freshw Ecosyst. 2018, 28, 1099–1104. [Google Scholar] [CrossRef]
- Tan, K.; Wang, W. The Early Life Culture and Gonadal Development of Giant Freshwater Prawn, Macrobrachium rosenbergii: A Review. Aquaculture 2022, 559, 738357. [Google Scholar] [CrossRef]
- Pillai, B.R.; Ponzoni, R.W.; Das Mahapatra, K.; Panda, D. Genetic improvement of giant freshwater prawn Macrobrachium rosenbergii: A review of global status. Rev. Aquac. 2022, 14, 1285–1299. [Google Scholar] [CrossRef]
- Fu, H.; Jiang, S.; Xiong, Y. Current Status and Prospects of Farming the Giant River Prawn (Macrobrachium rosenbergii) and the Oriental River Prawn (Macrobrachium nipponense) in China. Aquac. Res. 2012, 43, 993–998. [Google Scholar] [CrossRef]
- Chen, X.; Fan, L.; Qiu, L.; Dong, X.; Wang, Q.; Hu, G.; Meng, S.; Li, D.; Chen, J. Metagenomics Analysis Reveals Compositional and Functional Differences in the Gut Microbiota of Red Swamp Crayfish, Procambarus clarkii, Grown on Two Different Culture Environments. Front. Microbiol. 2021, 12, 735190. [Google Scholar] [CrossRef]
- Dong, S.; Gao, Y.; Gao, Y.; He, M.; Liu, F.; Yan, F.; Wang, F. Evaluation of the Trophic Structure and Energy Flow of a Rice-Crayfish Integrated Farming Ecosystem Based on the Ecopath Model. Aquaculture 2021, 539, 736626. [Google Scholar] [CrossRef]
- Nair, C.M.; Salin, K.R.; Joseph, J.; Aneesh, B.; Geethalakshmi, V.; New, M.B. Organic Rice–prawn Farming Yields 20 % Higher Revenues. Agron. Sustain. Dev. 2014, 34, 569–581. [Google Scholar] [CrossRef]
- Li, Y.; Wu, T.; Wang, S.; Ku, X.; Zhong, Z.; Liu, H.; Li, J. Developing Integrated Rice-animal Farming Based on Climate and Farmers Choices. Agric. Syst. 2023, 204, 103554. [Google Scholar] [CrossRef]
- Islam, M.S.; Shofiquzzoha, A.F.M.; Begum, N. Efficacy of Formulated Feed Preference and Stocking Density on Growth and Survival of Baby Pila globosa Reared in Laboratory Condition. Annu. Res. Rev. Biol. 2021, 36, 86–91. [Google Scholar] [CrossRef]
- Moss, A.S.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Dawood, M.A.O. Effects of Different Levels of Marine Snail Shells in the Diets of Juvenile Kuruma Shrimps Marsupenaeus japonicus as a Source of Calcium. N. Am. J. Aquac. 2019, 81, 55–66. [Google Scholar] [CrossRef]
- Hossain, M.M.; Chakraborty, S.C. Growth and Economic Analysis of Freshwater Prawn, Macrobrachium rosenbergii (de Man), Produced with Feeds Substituting Sunflower Cake for Fish Meal, Soya Bean Meal and Mustard Oil Cake. Aquac. Res. 2017, 48, 5418–5429. [Google Scholar] [CrossRef]
- Yang, R.; Cao, R.; Gong, X.; Feng, J. Large Shifts of Niche and Range in the Golden Apple Snail (Pomacea canaliculata), an Aquatic Invasive Species. Ecosphere 2023, 14, e4391. [Google Scholar] [CrossRef]
- Roberts, J.K.; Kuris, A.M. Predation and Control of Laboratory Populations of the Snail Biomphalaria glabrata by the Freshwater Prawn Macrobrachium rosenbergii. Ann. Trop. Med. Parasitol. 1990, 84, 401–412. [Google Scholar] [CrossRef]
- Savaya-Alkalay, A.; Ovadia, O.; Barki, A.; Sagi, A. Size-selective Predation by All-male prawns: Implications for Sustainable Biocontrol of Snail Invasions. Biol. Invasions 2018, 20, 137–149. [Google Scholar] [CrossRef]
- Sokolow, S.H.; Lafferty, K.D.; Kuris, A.M. Regulation of Laboratory Populations of Snails (Biomphalaria and Bulinus spp.) by River Prawns, Macrobrachium spp. (Decapoda, Palaemonidae): Implications for Control of Schistosomiasis. Acta Trop. 2014, 132, 64–74. [Google Scholar] [CrossRef]
- Monde, C.; Syampungani, S.; Rico, A.; van den Brink, P.J. The Potential for Using Red Claw Crayfish and Hybrid African Catfish as Biological Control Agents for Schistosoma Host Snails. Afr. J. Aquat. Sci. 2017, 42, 235–243. [Google Scholar] [CrossRef]
- Chimbari, M.J.; Madsen, H.; Ndamba, J. Laboratory Experiments on Snail Predation by Sargochromis codringtoni, a Candidate for Biological Control of the Snails that Transmit Schistosomiasis. Ann. Trop. Med. Parasitol. 1997, 91, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Styles, D.; Cao, Y.; Ye, X. The Sustainability of Rice-crayfish Coculture Systems: A Mini Review of Evidence from Jianghan Plain in China. J. Sci. Food. Agric. 2021, 101, 3843–3853. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Liu, H.; Wang, H.; Wu, S.; Bashir, M.A.; Reis, S.; Sun, Q.; Xu, J.; Gu, B. Rice-Animal Co-Culture Systems Benefit Global Sustainable Intensification. Earth’s Future 2023, 11, e2022EF002984. [Google Scholar] [CrossRef]
- Mat Zaib, N.A.N.; Tay, C.; Hashim, S.N.; Mohd Hassan, W.R. Azadirachta indica as Natural Pesticide for Pomacea canaliculata Control: A Review. Sci. Lett. 2023, 17, 1–11. [Google Scholar] [CrossRef]
- Wagiman, F.X.; Ariani Bunga, J.; Hasoloan Purba Sidadolog, J. Sustainable Control of the Golden Snail (Pomacea canaliculata Lamarck) on Irrigated Rice Field in Malaka Regency, East Nusa Tenggara Province, Indonesia. Kne Life Sci. 2019, 4, 156–165. [Google Scholar] [CrossRef]
- Zhang, J.; Quan, G.; Huang, Z.; Luo, S.; Ouyang, Y. Evidence of Duck Activity Induced Anatomical Structure Change and Lodging Resistance of Rice Plant. Agroecol. Sustain. Food Syst. 2013, 37, 975–984. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Q.; Dong, L.; Zhang, J. Cleaner Agricultural Production in Drinking-Water Source Areas for the Control of Non-Point Source Pollution in China. J. Environ. Manag. 2021, 285, 112096. [Google Scholar] [CrossRef]
- Palm, H.W.; Knaus, U.; Appelbaum, S.; Goddek, S.; Strauch, S.M.; Vermeulen, T.; Haїssam Jijakli, M.; Kotzen, B. Towards Commercial Aquaponics: A Review of Systems, Designs, Scales and Nomenclature. Aquac. Int. 2018, 26, 813–842. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G. Apple Snails as Animal Feed. In Biology and Management of Invasive Apple Snails, 2nd ed.; Joshi, R.C., Cowie, R.H., Sebastian, L.S., Eds.; Philippine Rice Research Institute (PhilRice): Nueva Ecija, Philippine, 2017; pp. 369–385. [Google Scholar]
- Oficialdegui, F.J.; Sánchez, M.I.; Clavero, M. One Century Away from Home: How the Red Swamp Crayfish Took Over the World. Rev. Fish. Biol. Fish. 2020, 30, 121–135. [Google Scholar] [CrossRef]
- Farmanfarmaian, A.; Moore, R. Diseasonal Thermal Aquaculture— 1. Effect of Temperature and Dissolved Oxygen on Survival and Growth of Macrobrachium rosenbergii. Proc. Annu. Meet.-World Maric. Soc. 1978, 9, 55–66. [Google Scholar] [CrossRef]
- Reid, G.K.; Gurney-Smith, H.J.; Flaherty, M.; Garber, A.F.; Forster, I.; Brewer-Dalton, K.; Knowler, D.; Marcogliese, D.J.; Chopin, T.; Moccia, R.D.; et al. Climate Change and Aquaculture: Considering Adaptation Potential. Aquac. Environ. Interact. 2019, 11, 603–624. [Google Scholar] [CrossRef]
- Savaya-Alkalay, A.; Ndao, P.D.; Jouanard, N.; Diane, N.; Aflalo, E.D.; Barki, A.; Sagi, A. Exploitation of Reproductive Barriers between Macrobrachium Species for Responsible Aquaculture and Biocontrol of Schistosomiasis in West Africa. Aquac. Environ. Interact. 2018, 10, 487–499. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Mean Prawn Size | |||
|---|---|---|---|
| Prawns with Juvenile Snails | Prawns with Subadult Snails | Controls (Prawns without Snails) | |
| Number of M. rosenbergii at the start of the experiment | 8 | 8 | 8 |
| Number of M. rosenbergii at the end of the experiment | 8 | 8 | 8 |
| Average length of M. rosenbergii (cm) | 8.30 ± 0.01 | 8.29 ± 0.01 | 8.28 ± 0.01 |
| Length of M. rosenbergii on day 1 (cm) | 8.23 ± 0.01 | 8.23 ± 0.01 | 8.25 ± 0.01 |
| Length of M. rosenbergii on day 12 (cm) | 8.31 ± 0.05 | 8.30 ± 0.02 | 8.28 ± 0.02 |
| Length of M. rosenbergii on day 18 (cm) | 8.36 ± 0.03 | 8.35 ± 0.02 | 8.31 ± 0.02 |
| Number | Shell Width | Range of Activity | |
|---|---|---|---|
| Juvenile | 89.26% | 68.61% | 88.16% |
| Subadult | −4.60% | 0.49% | 30.98% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, Y.; Lv, H.; Sheng, Q. Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming. Diversity 2023, 15, 1001. https://doi.org/10.3390/d15091001
Wang Y, Zhang Y, Lv H, Sheng Q. Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming. Diversity. 2023; 15(9):1001. https://doi.org/10.3390/d15091001
Chicago/Turabian StyleWang, Yingying, Yixiang Zhang, He Lv, and Qiang Sheng. 2023. "Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming" Diversity 15, no. 9: 1001. https://doi.org/10.3390/d15091001
APA StyleWang, Y., Zhang, Y., Lv, H., & Sheng, Q. (2023). Efficacy of Giant River Prawn Macrobrachium rosenbergii in Controlling the Invasive Snail Pomacea canaliculata: Implications for Ecological Farming. Diversity, 15(9), 1001. https://doi.org/10.3390/d15091001