Abstract
Fifteen riparian populations of Lithuanian Lythrum salicaria were assessed for leaf macronutrient, micronutrient and non-essential element concentrations and compared to the former obtained molecular data at amplified fragment length polymorphism (PLP.AFLP) loci. Inductively coupled plasma mass spectrometry was used to profile the contents of 12 elements in the leaves. The leaf nutrient concentrations were within normal ranges for growth and development and heavy metal concentrations did not reach toxic levels. The concentrations of macroelements such as nitrogen, potassium, calcium and magnesium were in the range of 23,790–38,183; 7327–11,732; 7018–12,306; and 1377–3183 µg/g dry mass (d. m.), respectively; the concentrations of micronutrients such as sodium, iron, zinc and copper varied in the ranges of 536–6328; 24.7–167.1; 10.88–26.24; and 3.72–5.30 µg/g d. m., respectively, and the concentrations of non-essential elements such as lead, nickel, chromium, and cadmium were in the intervals of 0.136–0.940; 0.353–0.783; 0.207–0.467; and 0.012–0.028 µg/g d. m., respectively. When comparing the maximum and minimum values for site elements of L. salicaria, the concentration of N varied by 1.6, K—1.6, Ca—1.8, Mg—2.3, Na—6.1, Fe—6.8, Zn—2.4, Cu—1.5, Pb—6.9, Ni—2.2, Cr—2.2, and Cd—2.3 times. The coefficient of variation (CV) of element concentrations in sites was moderate to large: N—15.4%, K—14.3%, Ca—18.6%, Mg—24.8%, Na—50.7%, Fe—47.0%, Zn—24.9%, Cu—14.5%, Pb—57.1%, Ni—30.11%, Cr—26.0%, and Cd—38.6%. Lythrum salicaria populations growing near regulated riverbeds were characterized by significantly (p < 0.05) lower concentrations of Ca and Mg, and significantly (p < 0.05) higher concentrations of N, K, Fe, Na, Ni, Cr and Cd. The PLP.AFLP was negatively correlated with concentrations of N, Na, Fe, Ni, Cr, and Cd. The L. salicaria population with the lowest leaf N and Na concentration showed the highest genetic polymorphism (PLP.AFLP = 65.4%), while the least polymorphic population (PLP.AFLP = 35.0%) did not show extreme concentrations of either element. In conclusion, our elemental analysis of L. salicaria populations showed that ionomic parameters are related to genomic parameters, and some habitat differences are reflected in the ionomes of the populations.
1. Introduction
Wetlands are very valuable habitats with the greatest diversity of plant and animal species. These areas are very important because they reduce the impact of extreme climate conditions. Wetlands are permanently disturbed by anthropogenic activities; as a result, these ecosystems are becoming increasingly sensitive [1].
Alterations of physical and chemical nature by humans make wetlands highly susceptible to plant invasions. Because of the growing damage caused by the spread of non-native species, biologists are increasingly focusing on understanding aquatic invasions [2].
Purple loosestrife (Lythrum salicaria L.), a member of the order Myrtales, family Lythraceae, is a tall perennial herb with tristylous reddish-purple flowers. Due to its amphibian lifestyle, it grows either submerged under water or above water. It is found along riverbanks and ditches, on the edges of lakes, in marshes and in wet meadows. Lythrum salicaria is naturally growing in Europe and temperate Asia. It belongs to an invasive species with long lag phases from introduction to eventual spread [3]. In the early 1800s, it arrived in North America and, since the 1930s, has become invasive [4]. To date, L. salicaria is attributed to the most aggressive species, forming monospecific communities in the wetlands of the United States and southern Canada [5].
Intensive use of fertilizers, urban and industrial activities are causing leakages of an excess of nutrients, which drain into aquatic ecosystems. As a result, lakes and rivers gradually become eutrophic [6,7,8]. Regarding the condition of rivers, parameters of the abiotic environment are usually recorded [9] together with macrophyte composition and coverage data [10,11]. The requirement of species for soil nutrients is estimated either by examining the concentration of soil elements or by employing Ellenberg indicator values (EIV) [12]. As indirect evidence of the abundance of nutrients in the soil, there are various plant growth data, for example, the mass of the aboveground part of all growing species per unit area or the mass of the aboveground part of a single individual [13]. The most accurate information about the condition of a plant could be obtained by measuring the concentration of nutritional elements in the leaves [13,14]. For a long time, amounts of macro- and microelements have been well examined for important forest tree species and cultivated food plants [15].
In the second half of the last century, many efforts were made to collect knowledge about the morphology, especially the flower structure, of L. salicaria [16], pollination, reproduction [17], cytology [18], DNA polymorphism [19,20,21,22], secondary metabolites [23] and interaction with other plant species [24,25,26], and various measures to control and eradicate the species have been examined [27,28,29]. Finding the best solutions to improve the management of invasive wetland plants requires filling gaps in knowledge about exotic species [30].
For many alien species, the situation is inadequate when comparing data collected within invasive areas and in the area of the natural distribution range. This also applies to L. salicaria. The majority of investigations of this species have been carried out only in North America [4,16,20,24,31,32,33,34], with a much smaller proportion of data covering both invasive and natural areas [19,21] or only natural areas [22,35]. Ionome investigations are gaining increasing attention [36]. Nutrients are responsible for maintaining the functions of organisms. They affect physiological processes and lead to morphological and quantitative changes in plant growth and development. So far, nutritional topics related to invasive species have not received enough attention [37,38]. Soil nutrient richness was the most important factor, accounting for more than 20% of the variance in coverage of invasive herbaceous plants [37]. Despite more than half a century of extensive research on L. salicaria as an invasive species, to our knowledge, more extensive ionomic data on L. salicaria populations from invasive or pristine areas are still lacking. As nutrient loads (such as N or P) are reduced in large parts of wetlands, the wise use of these habitats will remain an important strategy for the future [39].
Ionomic data on L. salicaria have been obtained from several studies in experimental wetlands with artificially simulated pollution [40,41,42,43,44]; under such circumstances, the knowledge acquired cannot be transferred to naturally growing plants. In the Baltic countries, L. salicaria is a common riparian species along the edges of water bodies, growing near lakes, ponds, rivers and ditches. In contrast to the dense coverage of this plant within the invasive areas of North America, the coverage by L. salicaria in permanently over-moistened habitats of Poland is not high (5%) [45]; a similar situation in terms of its coverage is also found on the banks of riparian sites of Lithuania [10]. Our recent investigations of the genetic diversity of L. salicaria populations in Lithuania revealed significant population differentiation in polymorphism loci of amplified fragments of DNA (AFLP) [22]. There is a growing worldwide interest in linking the molecular properties of organisms to physiology, including the ionome [46,47,48,49,50,51,52,53].
The aim of the study was to evaluate the concentrations of macroelements (N, K, Ca, Mg), microelements (Fe, Na, Zn, Cu) and non-essential elements (Pb, Ni, Cr, Cd) in the leaves of L. salicaria populations in 15 sites in Lithuania. Special tasks were to relate elemental data of the species with the extent of polymorphism at AFLP loci; and to evaluate the influence of distinct habitats with respect to (1) river basin (Nemunas, Lielupė and Seaside Rivers basins), (2) land cover and use type (agricultural areas, artificial areas and forest), and (3) riverbed origin (natural and regulated).
2. Materials and Methods
2.1. Study Area
Purple loosestrife (Lythrum salicaria L.) was collected from 15 sites in Lithuania. In addition to elemental analysis, molecular diversity at amplified fragment length polymorphism (AFLP) loci was also examined at those sites, as previously described [22]. The selected sites belong to three river basins—Nemunas, Seaside Rivers and Lielupė basin (Figure 1).
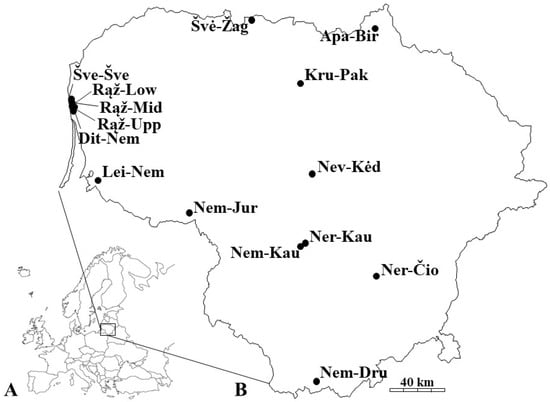
Figure 1.
Location of Lithuania (A), and 15 sites with purple loosestrife (Lythrum salicaria) growing in the country (B).
The sampling area extended between 54°01′12.9″ and 56°21′36.6″ North and between 21°03′21.8″ and 24°45′57.8″ East (Table 1).

Table 1.
The names and geographical locations of 15 populations of Lythrum salicaria in Lithuania (Figure 1) selected for element (macro-, microelement and non-essential) and AFLP [22] analysis.
2.2. Sampling and Element Analyses
Samples of L. salicaria leaf material were taken during the period of intensive flowering, 1–10 August 2015. In order to examine the concentration of macroelements, microelements and non-essential elements, healthy mature leaves, undamaged by fungi or insects, were collected from the middle part of the main shoot. Three batches of leaves were collected at each site for elemental analysis. The dried plant material was ground with a tungsten carbide grinder Retsch MM400 (Haan, Germany). The Kjeldahl method [54,55] was used to determine nitrogen concentration. For analyses, 0.2 g of L. salicaria leaves was digested in a Kjeldahl digestion unit DK-20S and subsequently analyzed with an automatic analyzer UDK 159 (VELP Scientifica, Usmate Velate, Italy). Quality assurance was achieved using standard reference materials SRM1515, SRM1575 and CRM125045.
For metal analyses with ICPS-MS, the mineralization of the ground material was performed in a high performance An Anton Paar Multiwave 3000 microwave digestion system using HNO3 and H2O2 solutions. Eleven essential and non-essential elements were analyzed with ICPS-MS (Thermo Scientific Element 2) according to the ISO 17294-2:2016 standard as it was done before [56,57,58]. The MERC ICP Multi-element solution IV standard was used for ICP-MS calibration.
2.3. Classification of Environment
Revealing the possible influence of the river and its environment on the nutritional status of the plant (evaluated by the concentration of leaf elements), the populations were grouped in several ways, as was done in the case of molecular analysis [22]: (1) three groups according to the river basin: Nemunas basin (Nem-Dru, Ner-Čio, Ner-Kau, Nem-Kau, Nev-Kėd, Nem-Jur, Lei-Nem; Figure 1, Table 1), Seaside Rivers basin (Rąž-Upp, Rąž-Mid, Rąž-Low, Dit-Nmr, Šve-Šve) and Lielupė basin (Švė-Žag, Apa-Bir, Kru-Pak); (2) three groups according to the land adjacent to the riverbank: artificial surfaces (ART; Nem-Kau, Rąž-Upp, Rąž-Mid, Rąž-Low, Dit-Nmr, Švė-Žag, Apa-Bir, Kru-Pak), agricultural areas (AGR; Nem-Dru, Ner-Čio, Ner-Kau, Nev-Kėd, Nem-Jur, Šve-Šve), forest and semi-natural areas (further in the text, named under forest areas, abbr. FOR; Lei-Nem), employing the CORINE Land Cover database (classification level (1) available for 2000 and 2006 [59]); (3) two groups according to riverbed origin: natural (Nem-Dru, Ner-Čio, Ner-Kau, Nem-Kau, Nev-Kėd, Nem-Jur, Šve-Šve, Švė-Žag, Apa-Bir, Kru-Pak) and regulated (Lei-Nem, Rąž-Upp, Rąž-Mid, Rąž-Low, Dit-Nmr) [60].
2.4. Data Analysis
Descriptive statistics were performed using the R software package (version 4.2.2) and the R package DescTools (version 0.99.42) [61,62]. The Kruskal–Wallis test was used to compare element concentrations between populations and between groups of populations. The R package compareGroups was used for this (version 4.5.1) [63]. The coefficient of variance (CV, in %) was calculated to characterize the heterogeneity of element concentrations at L. salicaria sites.
To relate physiological (element concentrations) and genetic data (percentage of polymorphic AFLP loci (PLP.AFLP) of L. salicaria, Spearman rank correlations were calculated by plotting a correlogram using the R package corrplot (version 0.90) [64]. Principal component analysis was performed for 12 physiological variables (concentrations of macroelements N, K, Ca, Mg, microelements Na, Fe, Zn and Cu, and non-essential elements Pb, Ni, Cr and Cd) and the molecular variable PLP.AFLP [22], using the statistical software R.
3. Results
3.1. Comparison of Populations Based on Elemental Concentrations
The median concentrations of macroelements in the leaves of Lithuanian populations of L. salicaria were within the following ranges (Figure 2): (1) N—23,790–38,183 μg/g d. m. (populations with extreme N concentrations, Kru-Pak and Nem-Kau, differed by a factor of 1.605); (2) K—7327–11,732 μg/g d. m. (populations with extreme K concentrations, Nem-Kau and Nem-Jur, differed by a factor of 1.601; (3) Ca—7018–12,306 μg/g d. m. (populations with extreme Ca concentrations, Kru-Pak and Nem-Jur, differed by a factor of 1.753); (4) Mg—1377–3183 μg/g d. m. (populations with extreme Mg concentrations, Kru-Pak and Nem-Jur, differed by a factor of 2.311). Multiple comparisons using the Kruskal–Wallis test did not reveal significant differences between populations in leaf concentrations of N, K, Ca and Mg.
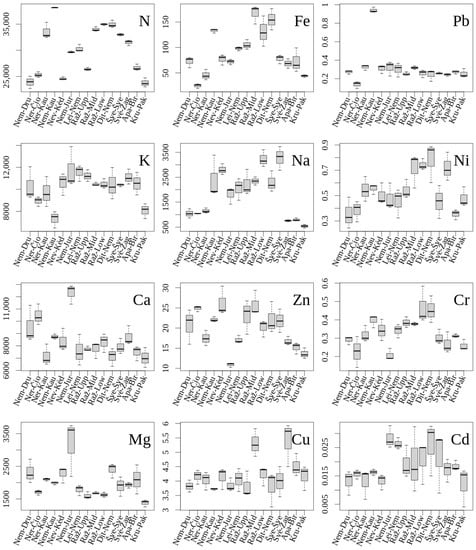
Figure 2.
Median values (box–whisker plots) of the leaf element concentrations (μg/g d. m. in y axes) in L. salicaria populations in Lithuania. The central line of each box indicates the median value; the boxes, the lower (25%) and upper (75%) quartiles; and the whiskers are from the 10th to 90th percentiles (typical range). The median (central horizontal line) with 25/75 percentiles (boxes), 10/90 percentiles (bars); the number of replicates per population n = 3.
The median concentrations of Na and microelements in the leaves of Lithuanian populations of L. salicaria were within the following ranges: (1) Na—536–6328 μg/g d. m. (populations with extreme Na concentrations, Kru-Pak and Šve-Šve, differed by a factor of 6.115); (2) Fe—24.7–167.1 μg/g d. m. (populations with extreme Fe concentrations, Ner-Čio and Raž-Mid, differed by a factor of 6.763); (3) Zn—10.88–26.24 μg/g d. m. (populations with extreme Zn concentrations, Nem-Jur and Nev-Kėd, differed by a factor of 2.413); (4) Cu—3.72–5.30 μg/g d. m. (populations with extreme Cu concentrations, Nem-Kau and Raž-Mid, differed by a factor of 1.448). Multiple comparisons using the Kruskal–Wallis test did not reveal significant differences between populations in leaf concentrations of Na, Fe, Zn and Cu.
The median concentrations of non-essential elements in the leaves of Lithuanian populations of L. salicaria were within the following ranges: (1) Pb—0.136–0.940 μg/g d. m. (populations with extreme Pb concentrations, Ner-Čio and Nem-Kau, differed by a factor of 6.884); (2) Ni—0.353–0.783 μg/g d. m. (populations with extreme Ni concentrations, Apa-Bir and Raž-Low, differed by a factor of 2.218); (3) Cr—0.207–0.467 μg/g d. m. (populations with extreme Cr concentrations, Nem-Jur and Raž-Low, differed by a factor of 2.218); (4) Cd—0.012–0.028 μg/g d. m. (populations with extreme Cd concentrations, Kru-Pak and Nem-Jur, differed by a factor of 2.349). Multiple comparisons using the Kruskal-–Wallis test did not reveal significant differences between populations in leaf concentrations of Pb, Ni, Cr and Cd.
When comparing the maximum and minimum site values for elements of L. salicaria, the concentration of N varied by 1.6 times (p < 0.05), K—1.6 (p < 0.05), Ca—1.8 (p < 0.05), Mg—2.3 (p < 0.05), Na—6.1 (p < 0.05), Fe—6.8 (p < 0.05), Zn—2.4 (p < 0.05), Cu—1.5 (p > 0.05), Pb—6.9 (p < 0.05), Ni—2.2 (p < 0.05), Cr—2.2 (p < 0.05), and Cd—2.3 (p < 0.05). The coefficient of variation (CV) of element concentrations in sites was moderate to large: N—15.4%, K—14.3%, Ca—18.6%, Mg—24.8%, Na—50.7%, Fe—47.0%, Zn—24.9%, Cu—14.5%, Pb—57.1%, Ni—30.11%, Cr—26.0%, and Cd—38.6%.
3.2. Comparison of Groups of Populations
Among the investigated populations of L. salicaria, 47% of the populations belonged to the Nemunas basin, 33% to the Seaside Rivers basin and 20% to the Lielupė basin. The leaves of the L. salicaria populations in the Nemunas basin were characterized by the following element concentrations (median value for the group of populations in μg/g d. m.) (Figure 3): N—29,620, K—9563, Ca—8822, Mg—2018, Na—1573, Fe—76.6, Zn—21.6, Cu—3.89, Pb—0.32, Ni—0.46, Cr—0.30, and Cd—0.02.

Figure 3.
Median values (box–whisker plots) of the leaf element concentrations (μg/g d. m. in y axes) of L. salicaria populations, growing in the Nemunas, Seaside Rivers and Lielupė basins. The central line of each box indicates the median value; the boxes, the lower (25%) and upper (75%) quartiles; the whiskers are from the 10th to the 90th percentile (typical range); the points are outliers. Population groups marked with different letters differ significantly (p < 0.05).
The leaves of L. salicaria populations collected in the Seaside Rivers basin were characterized by the following concentrations of elements (median value for the group of populations in μg/g d. m.) (Figure 3): N—33,830, K—10,452, Ca—7736, Mg—1705, Na—2760, Fe—129, Zn—21.7, Cu—4.20, Pb—0.27, Ni—0.63, Cr—0.39, and Cd—0.02. The leaves of L. salicaria populations collected in the Lielupė basin were characterized by the following element concentrations (median value for the group of populations in μg/g d. m.): N—26,450, K—10,482, Ca—7859, Mg—1834, Na—752, Fe—59.1, Zn—15.7, Cu—4.48, Pb—0.25, Ni—0.44, Cr—0.29, and Cd—0.02. Significant (p < 0.05) river basin-related differences were documented for the N, Ca, Mg, Fe, Na, Zn, Cu, Ni and Cr elements. The river basin did not have a significant effect on the concentrations of K, Mg, Pb and Cd. The leaves of L. salicaria populations collected in the Nemunas basin were characterized by significantly (p < 0.05) higher concentrations of Ca (compared to the populations of the Lielupė and Seaside Rivers basins) and Zn (compared to the populations of the Lielupė basin), and significantly lower concentrations of N, Fe, Na, Cu, Ni and Cr, (compared to the populations of the Seaside Rivers basin) (Figure 3). The populations of the Seaside Rivers basin were characterized by significantly (p < 0.05) higher concentrations of N, Fe, Na, Ni and Cr (compared to the populations of the Nemunas and Lielupė basins), and significantly (p < 0.05) lower concentrations of Ca (compared to the populations of the Nemunas basin) (Figure 3). Populations of the Lielupė basin were characterized by significantly higher concentrations of Cu (compared to the populations of the Nemunas basin), and significantly (p < 0.05) lower concentrations of N, Fe and Cr (compared to the populations of the Seaside Rivers basin), Ca (compared to the populations of the Nemunas basin) Na, and Zn (compared to the populations of the Nemunas and Seaside Rivers basins) (Figure 3).
Among the L. salicaria populations studied, according to land cover and use, 53% of populations grew adjacent to artificial areas, and 40% of the populations were located adjacent to agricultural areas. Only one population (7%) of L. salicaria grew near the forests. The leaves of L. salicaria populations growing next to artificial areas were characterized by the following concentrations of leaf elements (median value for the grouped populations (μg/g d. m.)) (Figure 4): N—33,000; K—10,388; Ca—7905; Mg—1894; Na—3281; Fe—77.80; Zn—21.80; Cu—3993; Pb—0.259; Ni—0.453; Cr—0.302; Cd—0.022.
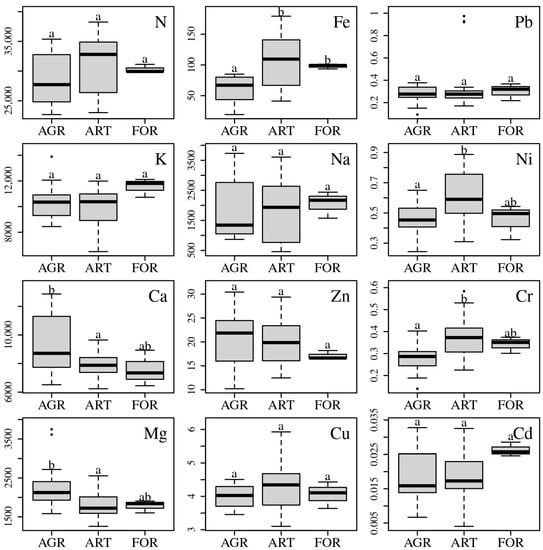
Figure 4.
Median values (box–whisker plots) of leaf element concentrations (μg/g d. m. in y axes) of L. salicaria populations growing near different type of land (according to cover and use): agricultural areas (AGR), artificial areas (ART) and forest (FOR). The central line of each box indicates the median value; the boxes, the lower (25%) and upper (75%) quartiles; the whiskers are from the 10th to the 90th percentiles (typical range); the points are outliers. Population groups marked with distinct letters differ significantly (p < 0.05).
The leaves of populations growing near agricultural areas were characterized by the following concentrations of elements (median values of grouped populations (μg/g d. m.): N—31,308; K—9958; Ca—7860; Mg—1837; Na—1822.5; Fe—110; Zn—19.85; Cu—4.368; Pb—0.348; Ni—0.611; Cr—0.365; Cd—0.019. Populations near agricultural land were characterized by significantly (p < 0.05) higher concentrations of such elements as Ca and Mg (compared to artificial areas) and significantly (p < 0.05) lower concentrations of Fe (compared to artificial and forest areas) and Ni and Cr (compared to artificial areas), while land cover and use type did not affect the concentrations of N, K, Na, Zn, Cu, Pb and Cd (Figure 4).
According to the nature of riverbeds, 67% of L. salicaria populations grew alongside natural riverbeds (N) and 33% alongside regulated riverbeds (R). The leaves of natural riverbed L. salicaria populations contained the following concentrations of elements (median values, μg/g d. m.) (Figure 5): N—28,440; K—9904; Ca—8444; Mg—2007; Na—1093; Fe—67.0; Zn—18.4; Cu—4.12; Pb—0.28; Ni—0.46; Cr—0.30; Cd—0.02. The leaves of L. salicaria populations of regulated riverbeds contained the following concentrations of elements (median values, (μg/g d. m.): N—33,830; K—10,722; Ca—7653; Mg—1705; Na—2345; Fe—129; Zn—21.1; Cu—4.20; Pb—0.28; Ni—0.63; Cr—0.39; Cd—0.02. Lythrum salicaria populations growing near natural riverbeds were characterized by significantly (p < 0.05) higher concentrations of Ca and Mg and significantly (p < 0.05) lower concentrations of N, K, Fe, Na, Ni, Cr and Cd, and the origin of riverbeds had no significant influence on Zn, Cu and Pb concentrations (Figure 5).
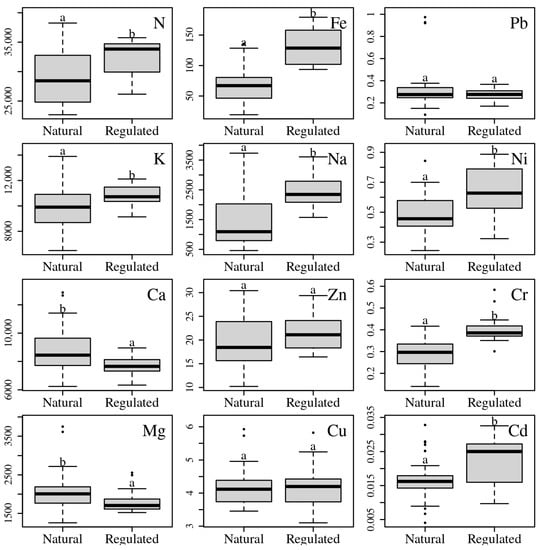
Figure 5.
Median values (box–whisker plots) of leaf element concentrations (μg/g d. m. in y axes) of L. salicaria populations growing near natural and regulated riverbeds. The central line of each box indicates the median value; the boxes, the lower (25%) and upper (75%) quartiles; the whiskers are from the 10th to 90th percentiles (typical range); the points are outliers. Population groups marked with distinct letters differ significantly (p < 0.05).
3.3. Relations between Element and Molecular Data
Looking for relationships between physiological and genetic parameters of L. salicaria populations, Spearman rank correlations between element concentrations and PLP.AFLP (percentage of amplified fragment length polymorphic loci) were calculated (Figure 6).

Figure 6.
The correlogram of Spearman rank correlation coefficients (Rs) (from −1 to 1) for all pairs of variables of Lythrum salicaria populations. Blue color represents positive and red color, negative, correlations. Color intensity indicates the strength of correlation, so the stronger the correlation, the darker the figures. Correlation numbers inside colored figures indicate level of significance; thin ellipsoid figures show significance (p < 0.05) while figures towards a spherical shape show insignificant correlations. PLP.AFLP—percentage of amplified fragment length polymorphic loci. N, K, Ca, and Mg—macroelements. Na—beneficial element. Fe, Zn, Cu—micronutrients. Pb, Ni, Cr, and Cd—non-essential elements.
Concentration of N was positively correlated with concentrations of Na (Rs = 0.536, p < 0.0396), Fe (Rs = 0.643, p < 0.0097), Ni (Rs = 0.679, p < 0.0054), and Cr (Rs = 0.631, p < 0.0117). The concentration of K was positively correlated with the concentration of Cd (Rs = 0.591, p < 0.0204). The concentration of Na was positively correlated with the concentrations of Fe (Rs = 0.732, p < 0.0019), Zn (Rs = 0.557, p < 0.031), and Cr (Rs = 0.66, p < 0.0075). The concentration of Fe was positively correlated with the concentrations of N (Rs = 0.643, p < 0.0097), Na (Rs = 0.732, p < 0.0019), and Cr (Rs = 0.854, p < 0.001). The concentration of Zn was positively correlated with the concentration of Na (Rs = 0.557, p < 0.031). The concentration of Ni was positively correlated with the concentration of N (Rs = 0.621, p < 0.0135). The concentration of Cr was positively correlated with the concentrations of N (Rs = 0.631, p < 0.0117), Na (Rs = 0.66, p < 0.0075), and Ni (Rs = 0.621, p < 0.0135). The concentration of Cd was positively correlated with the concentration of K (Rs = 0.591, p < 0.0204). The concentrations of Ca, Mg, Zn, Cu, and Pb did not correlate with any other element concentration.
The PLP.AFLP was negatively correlated with concentrations of N (Rs = −0.657, p < 0.0078), Na (Rs = −0.793, p < 0.0004), Fe (Rs = −0.721, p < 0.0024), Ni (Rs = −0.538, p < 0.0386), Cr (Rs = −0.584, p < 0.0221), and Cd (Rs = −0.781, p < 0.0006).
In order to clarify which parameters were most important for the variability of L. salicaria populations, a principal component (PC) analysis was performed (Figure 7) using concentrations of elements and PLP.AFLP variables.
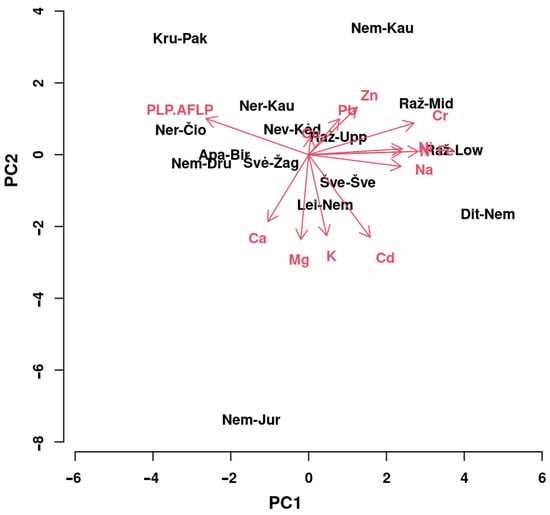
Figure 7.
Biplot of the principal component analysis for the first two principal components of a model testing variation of Lythrum salicaria populations depending on PLP at AFLP loci and concentrations of macronutrients (N, K, Ca, and Mg), Na, micronutrients (Fe, Zn, and Cu), and non-essential elements (Pb, Ni, Cr, Cd). Titles of populations are in black letters (Figure 1; Table 1), and red arrows with letters denote variables.
Our study showed that the first four PCs were quite informative, accounting for 79.76% of the overall variance for the entire set of variables. For separate principal components PC1, PC2, PC3, and PC4, the variance (with eigenvalues) was as follows: 36.38% (4.730), 19.50% (2.535), 14.11% (1.834), and 9.77% (1.270), respectively. PC1 variability was mainly caused by the concentrations of Fe, Cr, PLP.AFLP, and concentrations of Ni, N, Na, and Cd; the contribution of concentrations of most elements was positive, with the exception of Ca and Mg. For variability of PC2, concentrations of Mg, Cd, K, Ca and Zn, and PLP.AFLP were the most important; contributions of Zn, PLP.AFLP, Pb, Cr, Cu, Na, Ni, Fe and N were positive; and contributions of Mg, Cd, K, Ca and Na were negative. According to the variability of the parameters, displayed in the biplot of two principal components, the importance of the variables (in descending order) in PC1 was as follows: Fe > Cr > PLP.AFLP > Ni > N > Na > Cd > Zn > Ca > Pb > K > Mg > Cu, and the order of importance of the variables in PC2 was different: Mg > Cd > K > Ca > Zn > PLP.AFLP > Pb > Cr > Cu > Na > Ni > Fe > N.
The most extreme locations in the PC biplot were characteristic for populations of the Seaside Rivers basin (Dit-Nem, and Raž-Mid), Nem-Kau, Nem-Jur, and Kru-Pak. Compared to the remaining populations, (1) PLP.AFLP was the highest (65.4%,) and the concentrations of N and Ca, Mg and Na were the lowest, for population Kru-Pak; (2) the least-polymorphic population (PLP.AFLP = 35.0%), Dit-Nem, did not show extreme concentrations of either element; (3) the concentrations of N and Pb were the highest and the concentrations of K and Cu the lowest for population Nem-Kau; (4) the concentrations of K, Ca, Mg and Cd were the highest, and the concentrations of Zn and Cr were the lowest, for population Nem-Jur; (5) Rąž-Mid had the highest concentrations of Fe and Cu.
4. Discussion
Despite the fact that L. salicaria is widely distributed in the world as a naturally growing or invasive species, element data for this species were not found in the most extensive European and Asian studies of ionomics [13,14,65,66,67]. The concentrations of elements in the leaves, more often in the aboveground or in undetermined parts of L. salicaria growing in the natural environment, have been documented only in rare cases [68,69,70,71,72,73]. Recently, ionome analyses of L. salicaria have mostly been performed in hydroponic experiments with nutrient and/or heavy metal additions [74,75], so the comparison of our data with such cases is very relative.
4.1. Macroelements
Nitrogen. Nitrogen concentrations in Lithuanian L. salicaria (23,790–38,183 μg/g d. m., leaves, Figure 2) were similar to the nitrogen concentration in this species in the flood model (15,000–21,000 μg/g d. m., aboveground part) [75]. When L. salicaria was grown in a medium with nitrogen, phosphorus and potassium for 70 days, the nitrogen concentration was higher (42,000 µg/g d. m.) [76]. Compared to the other riparian species of Lithuania (Phalaris arundinacea, Bidens frondosa, Phragmites australis, Echinocystis lobata), Lythrum salicaria demonstrated the lowest leaf N concentration [77]. Very often, riparian community data are discussed in relation to the nutritional state of the species [78]. Compared to the abundance of other Lithuanian macrophytes [10], the abundance of L. salicaria individuals is not high. In contrast to our sampling sites, within the invasive range (wetlands of North America), Lythrum salicaria is forming dense monolithic stands [3,4,5]. Nitrogen is the most important element for biomass production. Lythrum salicaria may have encountered excess nitrogen in the invaded areas. Compared to the other species studied by us, L. salicaria showed the largest differences in nitrogen concentration between populations. Such a fact indicates the potential of the species to use higher amounts of nitrogen. Our nitrogen data are supported by the indicator value of soil nutrient richness (nitrophobic–nitrophil interval 1–9) [12], according to which L. salicaria was defined as a species growing in various soils. A soil richness study of L. salicaria in the Czech Republic showed an average indicatory value (5.6) among other species, with soil richness ranging from 2.4 to 7.1 [79]. The worldwide invasion success of European species is likely to have been promoted by the global increase in nutrient-enriched sites. It is postulated that alien plants are successful invaders because they have a broader capability for nutrient consumption than native plants [80,81]. In addition, it has been shown that species from more productive habitats are more invasive [79]. Greenhouse experiments on water and nitrogen additions to invasive Bidens frondosa and native B. tripartita revealed higher phenotypic plasticity for the alien species [68]. It has been shown that in eutrophic environments, Lythrum salicaria may invest nitrogen in building root biomass; thus, cultivation of the plants in constructed wetlands can successfully reduce the nitrogen concentration from wastewater [82].
The potential of the species to use elevated amounts of nitrogen, revealed in our study, corresponds to numerous phytoremediation studies. Cultivation of L. salicaria in a eutrophic environment (constructed wetland) successfully reduced the nitrogen concentration in wastewater [82]. In constructed wetland conditions, L. salicaria was a valuable nitrogen absorber which could be used to remove excess nitrogen [44]. This plant effectively removed nitrogen from antibiotic-contaminated wetlands [83]. In artificial wetlands with L. salicaria and Canna indica, the plants removed more than 90% of nitrate, ammonia or total of nitrogen [42]. In the studies with supplied contaminants, most often, only the nitrogen effect is examined. Wide-scope analyses of experimental data revealed that an excess of N might aggravate the uptake of phosphorus, and the consumption of other elements might be disturbed [84]; thus, a bigger set of elements should be included in phytoremediation assessments of L. salicaria.
Potassium. Potassium, along with calcium, is the most abundant element in leaves after nitrogen, highlighting its major contribution to plant functioning [85]. The potassium concentrations (7327–11,732 μg/g d. m., leaves) of Lithuanian L. salicaria were similar to the data on L. salicaria (5000–18,000 μg/g d. m., aboveground parts) in the flood simulation experiment [75]; a higher potassium concentration (16,500 μg/mg d. m., aboveground parts) was determined in L. salicaria growing in the flood plains of the Tisza River [68,69]. Lythrum salicaria growing in the flood plains of the Tisza River contained less potassium and much more sodium than meadow-hays [68]. After 70 days of incubation of L. salicaria in a medium containing nitrogen, phosphorus and potassium, very high concentrations of potassium (80,000 µg/g d. m., leaves) [76] were documented, in parallel with changes in plant architecture, dry mass of the aboveground part and leaf area.
Calcium. The calcium concentrations in Lithuanian L. salicaria (7018–12,306 μg/g d. m., leaves) were very similar to concentrations of calcium of L. salicaria from Rhodes Island in Greece (892–1247 µg/g d. m. leaves) [72] and L. salicaria in the flood simulation experiment (5000–18,000 μg/g d. m. aboveground parts) [75] and lower than the concentration of calcium of L. salicaria in the flood plains of the Tisza River (15,800 μg/mg d. m., aboveground part) [68,69]. After 70 days of incubation of L. salicaria in a medium containing nitrogen, phosphorus and potassium, higher concentrations of calcium (80,000 µg/g d. m., leaves) [76] were determined, similar to what was obtained in the case of potassium.
Magnesium. In addition to the well-known role of magnesium in the vitality of plants [86], it was shown that Mg inhibits the absorption of heavy metals and therefore reduces their toxicity in plants [87]. The magnesium concentrations in Lithuanian L. salicaria (1377–3183 μg/g d. m., leaves) encompassed the magnesium concentration of L. salicaria in the flood plains of the Tisza River (2640 μg/g d. m.) [69] and were lower than magnesium concentrations in L. salicaria on Rhodes Island, Greece (3469–4741 µg/g d. m., leaves) [72].
4.2. Microelements
Iron. It was estimated that a concentration of Fe > 500 μg/g d. m. is phytotoxic [87]. The iron concentrations in Lithuanian L. salicaria (24.7–167.1 μg/g d. m., leaves, Figure 3) were lower compared to L. salicaria in the flood plains of the Tisza River (254 μg/g d. m., aboveground parts) [69] and nearly twice the concentration in L. salicaria (328 μg/g d. m., aboveground parts) found in the tidal wetlands of Belgium and The Netherlands [88].
Sodium. Sodium is beneficial to many species at lower levels of supply and essential in some species (e.g., C4) [89]. According to the salt indicator value of 1 (on a scale of 0–9 for salt), [12] L. salicaria is salt tolerant; it usually grows in low-salt to salt-free soils, while most of the riparian species of Lithuania have a salt indicator value of 0 (glycophytes, which do not tolerate salts). Some of our selected populations grew close to the sea, where the atmosphere is enriched by sodium-containing aerosols. The sodium concentrations in L. salicaria were lower in the flood plains of the Tisza River (520 μg/g d. m., aboveground parts) [69].
Zinc. Zinc enters the environment through sewage sludge and municipal waste [90]. The concentration of zinc in plants ranges from 15 to 100 μg/g d. m. [54]. At low concentrations, zinc acts as a micronutrient but becomes toxic when its concentration in the plant reaches 100 μg/g d. m. [87], and at a concentration of 300 μg/g d. m., visible symptoms of leaf damage appear. There is evidence that zinc is able to alleviate cadmium-induced toxicity [91]. The zinc concentrations in Lithuanian L. salicaria (10.88–26.24 μg/g d. m., leaves) were comparable to the zinc concentrations in L. salicaria from Sevan Lake in Armenia (the number was taken from figure, ~20 μg/g d. m., leaves) [71]. Two or more times’ higher concentrations of zinc were found in L. salicaria from the tidal wetlands of Belgium and The Netherlands (59.6 μg/g d. m., aboveground parts) [88], in L. salicaria near Köprüören Pond in Turkey (110 µg/g d. m., aboveground parts) [92], in L. salicaria from Rhodes Island in Greece (87.7–178.4 µg/g d. m. leaves) [72], and in L. salicaria from the flood plains of the Tisza River (172.8 μg/g d. m., aboveground parts) [69]. Comparative analyses showed that L. salicaria accumulates more Zn than other hydrophytes [69,71]. After seven days of growth of L. salicaria seedlings in the solutions of zinc (up to 100 mg/L zinc), the concentration of zinc accumulated in the leaves reached 1084.7 μg/g d. m. Considering the high capacity of L. salicaria to accumulate zinc and tolerate high zinc concentrations, this species might be used for phytoremediation [93].
Copper. At low concentrations, Cu, like Zn, normally acts as a micronutrient but becomes toxic when its concentration in the plant increases [87]. Copper concentrations in Lithuanian L. salicaria (3.72–5.30 μg/g d. m., leaves) were the same as in L. salicaria from Lake Sevan in Armenia (the number was taken from the figure, ~5 μg/g d. m., aboveground parts) [71] or from the tidal wetlands of Belgium and The Netherlands (6.81 μg/g d. m., aboveground parts) [88]. Almost twice the copper concentrations were found in L. salicaria from the flood plains of the Tisza River (9.8 μg/g d. m., aboveground parts) [69] and Rhodes Island in Greece (10.0–14.4 μg/g d. m., leaves) [72].
4.3. Heavy Metals
The toxicity of heavy metals is an important factor limiting plant growth and development [94,95,96]. Metal stress responses have been extensively examined among food species [97], with inconclusive information on naturally growing macrophytes [91] receiving unused fertilizers and pesticides. In the Baltic region, much attention has been paid to the study of heavy metals in conifers and mosses [98,99,100,101], while the data on elements of herbaceous angiosperms are still very scarce [102].
Lead. The toxic concentrations of lead for plants are >27 μg/g d. m. [87]. As in aboveground parts of vegetation collected from the buffer zones of wetlands in Lithuania [103], the lead concentrations in Lithuanian L. salicaria were within the background values (0.136–0.940 μg/g d. m. leaves, Figure 3) and similar to the lead concentrations in L. salicaria in Lake Sevan, Armenia (the number was taken from the figure, ~0.1 μg/g d. m., aboveground parts) [71] or to tidal wetlands of Belgium and The Netherlands (1.02 μg/g d. m., aboveground parts) [92] as well as to Rhode Island, Greece (0.1–1.5 μg/g d. m., leaves) [72]. Growth of L. salicaria in lead-containing soil reduced the aboveground mass, but the belowground parts of the plant remained intact due to the ability to replace damaged shoots with new ones developing from healthy roots [40,41]. Evaluating the effects of lead concentrations (up to 2000 mg/L) on the growth of L. salicaria revealed tolerance to lead contamination [40].
Nickel. Nickel concentrations in Lithuanian populations of L. salicaria were low (0.353–0.783 μg/g d. m., leaves) and were very similar to L. salicaria from the tidal wetlands of Belgium and The Netherlands (0.489 μg/g d. m., aboveground parts) [92] or lower than in Rhodes Island in Greece (0.5–1.5 μg/g d. m., leaves) [72] or Lake Sevan in Armenia (number was taken from figure, ~1.2 μg/g d. m., aboveground parts) [71]. After 15 days of hydroponic cultivation of L. salicaria seedlings in a medium enriched with nickel up to 100 mg/L, the plants accumulated significantly more nickel (418.4 μg/g d. m., leaves) [104] than in the natural environment (described above).
Chromium. The adverse effect of chromium on the growth of plants is related to the disturbed metabolism of nutritional elements [105]. Concentrations of chromium in Lithuanian populations of L. salicaria were low (0.207–0.467 μg/g d. m., leaves), compared to several times’ higher concentration of chromium in L. salicaria from the tidal wetlands of Belgium and The Netherlands (2.20 μg/g d. m., aboveground parts) [88] or to Rhodes Island in Greece (0922–4160 μg/g d. m., leaves) [72].
Cadmium. Cadmium is toxic to plants due to its ability to displace certain nutrients as components of enzymes. In addition, cadmium may impair nutrient uptake, causing a deficiency of calcium [106]. The cadmium concentrations in Lithuanian L. salicaria (0.012–0.028 μg/g d. m., leaves) were several times lower than those in Rhode Island (0.109–0.327 μg/g d. m., leaves) [72], in the tidal wetlands of Belgium and The Netherlands (0.173 μg/g d. m., leaves) [88], and in Lake Sevan in Armenia (number taken from figure, ~0.2 μg/g d. m., aboveground parts) [71]. According to the growth rate and the accumulation of Cd and other heavy metals (Zn, Cu, Pb), in a comparison of nine other Chinese plant species [43], Lythrum salicaria appeared to be the most resistant to pollutants and very good at absorbing and accumulating metals in stems and leaves.
4.4. Ionome-Environment Relationships
River basins. Biologists and agronomists are increasingly interested in the dependence of plant properties on environmental parameters [48,107]. Recent studies have shown that the elemental concentration profile of Arabidopsis halleri (L.) allowed discrimination depending on geographical location [108]. The classification of Lythrum salicaria populations studied by us into three groups according to the Nemunas, Seaside Rivers and Lielupė basins (Table 1) revealed statistically significant differences in element concentrations (Figure 3), which could be caused by both the natural environment and human influence. Higher sodium concentrations in L. salicaria populations located in the Seaside Rivers basin were probably related to the presence of sodium-containing aerosols near the Baltic Sea. Higher concentrations of N, Ni and Cr in L. salicaria populations from the Seaside Rivers basin may have resulted from very close residences, very intensive tourism and ornamental horticulture.
Land cover and use. Land cover and land use did not affect the concentrations of N, K, Na, Zn, Cu, Pb and Cd (Figure 4) for the groups of L. salicaria populations we studied (mostly belonging to agricultural and artificial areas) (Figure 4). Differences in land cover and use type were not reflected in leaf N concentrations of neighboring macrophyte species populations, either [77]. Such results could be caused by the reduced intensification of Lithuanian agriculture, transitioning to environmentally friendly farming, and the simultaneous increase in the load of the aforementioned elements in artificial territories. Growth of Lythrum salicaria in flooded or non-flooded soil cores from different land-use areas (agricultural and semi-natural grassland in the floodplain of the Beerze River in The Netherlands) also showed no significant differences in aboveground N concentration, but L. salicaria was more productive after winter flooding in agricultural areas [75]. An assessment of potential invasive effects on phosphorus metabolism in a waterlogged area in Minnesota with Lythrum salicaria displacing Typha sp. was inconclusive [70].
River regulation. After the Second World War, the intensification of Lithuanian agriculture was extremely important. More than 80% of the country’s riverbeds have been restructured [60]. There is evidence of various damage to the landscape (habitat fragmentation, reduced macrophyte diversity) caused by river regulation [109]. Based on the data of present study, populations of L. salicaria near regulated riverbeds were characterized by significantly lower concentrations of Ca and Mg and significantly higher concentrations of N, K, Fe, Na, Ni, Cr and Cd (Figure 5). Fragments of regulated Lithuanian riverbeds can occur in soils that were previously used for agriculture and were enriched with N and heavy metals due to the use of fertilizers or pesticides. In parallel with L. salicaria ionomic transformations, significant differences in molecular diversity were recorded at AFLP loci [22] between the same population groups from natural and regulated riverbeds. Studies of other aquatic species (Phalaris arundinacea, Batrachium spp. and Bidens spp.) have also revealed significant changes in genetic diversity associated with river canalization [110,111,112].
4.5. Ionome–Genome Relationships
A assessment of AFLP loci in the same populations of Lythrum salicaria has shown that molecular diversity of L. salicaria populations in Lithuania varied significantly depending on the river basin, land cover and use type, and river regulation [22]. This was supported by the results of a significant 7% molecular variation of microsatellite loci of Nuphar lutea populations from the Nemunas Basin [113].
Until now, there have been few attempts to integrate plant ionomics and genomics [51,96,114,115]. Due to heavy metal exposure, isoenzymatic and cytological changes have been recorded in Pinus sylvestris populations in subsequent generations [115]. Analyses of fragmented amplified polymorphic sequences and microsatellite loci markers in Brassicaceae have led to the discovery of genes associated with heavy metal tolerance and/or hyperaccumulation [51]. Genome-wide association studies revealed polymorphisms (SNPs) affecting genomic variation in Manihot esculenta [116].
Present study examined the interrelationships between genetic and physiological parameters of Lythrum salicaria populations. The results showed that populations with a higher percentage of polymorphic loci had lower leaf concentrations of N, Na, Fe, Ni, Cr and Cd (Figure 6). Furthermore, the percentage of polymorphic AFLP loci and Fe and Cr concentrations were the most important factors contributing to population variability (Figure 7).
Leaf nutrient concentrations were within normal limits for growth and development, and heavy metal concentrations did not reach toxic levels. Our ionomic data show that the growth of perennial L. salicaria in environments more polluted with nitrogen or heavy metals may have negative consequences on the genetic structure. This assessment extended the ionomic data on leaf elemental concentrations in L. salicaria populations growing in their natural range. To confirm the results of this study, future assessments of L. salicaria should include more populations that differ in environmental characteristics and represent both native and invasive ranges of the species.
5. Conclusions
A comparison of the ionomic data on Lythrum salicaria populations with the amounts of elements defined for the species grown in constructed wetlands with artificial contaminants show that nitrogen or heavy metals pollution levels are not high in Lithuanian riparian habitats. Concentration ranges of Lithuanian Lythrum salicaria leaf elements further demonstrated the species’ potential for indicatory and phytoremediation applications.
Author Contributions
Conceptualization, E.K. (Eugenija Kupčinskienė); data curation, E.K. (Edvina Krokaitė), L.J. and E.K. (Eugenija Kupčinskienė); software, T.R. and L.J.; funding acquisition, E.K. (Eugenija Kupčinskienė); investigation, L.J., E.K. (Edvina Krokaitė), D.S., D.V. and T.R.; methodology, E.K. (Eugenija Kupčinskienė), E.K. (Edvina Krokaitė), L.J. and D.V.; project administration, L.J.; visualization, L.J., E.K. (Edvina Krokaitė) and T.R.; writing—original draft, E.K. (Eugenija Kupčinskienė), E.K. (Edvina Krokaitė), L.J., D.S., T.R. and D.V.; writing—review and editing, E.K. (Eugenija Kupčinskienė), L.J., E.K. (Edvina Krokaitė) and T.R. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Research Council of Lithuania, Grant No. SIT-02/2015.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Jūratė Šikšnianienė for technical assistance in capillary electrophoresis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Janse, J.H.; Van Dam, A.A.; Hes, E.M.; de Klein, J.J.; Finlayson, C.M.; Janssen, A.B.; van Wijk, D.; Mooij, M.W.; Verhoeven, J.T. Towards a global model for wetlands ecosystem services. Curr. Opin. Environ. Sustain. 2019, 36, 11–19. [Google Scholar] [CrossRef]
- van Kleunen, M.; Bossdorf, O.; Dawson, W. The ecology and evolution of alien plants. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 25–47. [Google Scholar] [CrossRef]
- Anderson, N.O. Throwing out the bathwater but keeping the baby: Lessons learned from purple loosestrife and reed canarygrass. HortTechnology 2019, 29, 539–548. [Google Scholar] [CrossRef]
- Thompson, D.Q.; Stuckey, R.L.; Thompson, E.B. Spread, impact, and control of purple loosestrife (Lythrum salicaria) in North American wetlands. U.S. Fish Wildl. Serv. 1987, 1–55. [Google Scholar]
- Lavoie, C. Should we care about purple loosestrife? The history of an invasive plant in North America. Biol. Invasions 2010, 12, 1967–1999. [Google Scholar] [CrossRef]
- Vet, R.; Artz, R.S.; Carou, S.; Shaw, M.; Ro, C.U.; Aas, W.; Baker, A.; Van Bowersox, C.; Dentener, F. A global assessment of precipitation chemistry and deposition of sulfur, nitrogen, sea salt, base cations, organic acids, acidity and pH, and phosphorus. Atmos. Environ. 2014, 93, 3–100. [Google Scholar] [CrossRef]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Baumgarte, I.; Freeman, A.; Gunn, I.D.M.; Lázár, A.N.; Sinclair, R.; Wade, A.J.; Bowes, M.J. Responses of aquatic plants to eutrophication in rivers: A revised conceptual model. Front. Plant Sci. 2018, 9, 451. [Google Scholar] [CrossRef]
- Erős, T.; Kuehne, L.; Dolezsaib, A.; Sommerwerk, N.; Wolter, C. A systematic review of assessment and conservation management in large floodplain rivers–actions postponed. Ecol. Indic. 2019, 98, 453–461. [Google Scholar] [CrossRef]
- Fuller, M.R.; Doyle, M.W.; Strayer, D.L. Causes and consequences of habitat fragmentation in river networks. Ann. N. Y. Acad. Sci. 2015, 1355, 31–51. [Google Scholar] [CrossRef]
- Zviedre, E.; Vītola, I.; Vizule-Kahovska, L.; Upena, I. Evaluation of Phytobenthos and Macrophytes of the Inland Surface Waters and Ecological Status Defined by Macrophyte Reference Index; Report, Part II, Rivers; Latvian Environment, Geology and Meteorology Centre: Ryga, Latvia, 2015. [Google Scholar]
- Hille, S.; Larsen, S.E.; Rubæk, G.H.; Kronvang, B.; Baattrup-Pedersen, A. Does regular harvesting increase plant diversity in buffer strips separating agricultural land and surface waters? Front. Environ. Sci. 2018, 6, 58. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Indicator values of plants in Central Europe. Scr. Geobot. 1992, 18, 258. [Google Scholar]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2015, 168, 377–385. [Google Scholar] [CrossRef]
- Thompson, K.; Parkinson, J.A.; Band, S.R.; Spencer, R.E. A comparative study of leaf nutrient concentrations in a regional herbaceous flora. New Phytol. 1997, 136, 679–689. [Google Scholar] [CrossRef]
- Innes, J.L. Influence of air pollution on the foliar nutrition of conifers in Great Britain. Environ. Pollut. 1995, 88, 183–192. [Google Scholar] [CrossRef]
- Anderson, N.O.; Ascher, P.D. Style morph frequencies in Minnesota populations of Lythrum (Lythraceae) II. Tristylous L. salicaria L. Sex. Plant Reprod. 1995, 8, 105–112. [Google Scholar] [CrossRef]
- Caswell, W.D.; Davis, A.R. Pollen and ovule production, floral nectary structure, and nectar secretion dynamics in tristylous Lythrum salicaria L. Plant Syst. Evol. 2011, 294, 127–145. [Google Scholar] [CrossRef]
- Kubátová, B.; Trávníček, P.; Bastlová, D.; Čurn, V.; Jarolímová, V.; Suda, J. DNA ploidy-level variation in native and invasive populations of Lythrum salicaria at a large geographical scale. J. Biogeogr. 2008, 35, 167–176. [Google Scholar] [CrossRef]
- Chun, Y.J.; Nason, J.D.; Moloney, K.A. Comparison of quantitative and molecular genetic variation of native vs. invasive populations of purple loosestrife (Lythrum salicaria L., Lythraceae). Mol. Ecol. 2009, 18, 3020–3035. [Google Scholar] [CrossRef]
- Houghton-Thompson, J.; Prince, H.H.; Smith, J.J.; Hancock, J.F. Evidence of Hybridization between Lythrum salicaria (Purple Loosestrife) and L. alatum (Winged Loosestrife) in North America. Ann. Bot. 2005, 96, 877–885. [Google Scholar] [CrossRef]
- Middleton, B.A.; Travis, S.E.; Kubátová, B.; Johnson, D.; Edwards, K.R. Morphology and genetics of Lythrum salicaria from latitudinal gradients of the Northern Hemisphere grown in cold and hot common gardens. PLoS ONE 2019, 14, e0208300. [Google Scholar] [CrossRef]
- Jocienė, L.; Krokaitė, E.; Shakeneva, D.; Rekašius, T.; Stanys, V.; Šikšnianienė, J.B.; Žvingila, D.; Paulauskas, A.; Kupčinskienė, E. Relationship between genetic and environmental characteristics of Lithuanian populations of purple loosestrife (Lythrum salicaria). J. Environ. Eng. Landsc. Manag. 2022, 30, 81–93. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Chemical constituents and pharmacological effects of Lythrum salicaria—A review. IOSR J. Pharm. 2019, 9, 51–59. [Google Scholar]
- Brown, B.J.; Mitchell, R.J.; Graham, S.A. Competition for pollination between an invasive species (purple loosestrife) and a native congener. Ecology 2002, 83, 2328–2336. [Google Scholar] [CrossRef]
- Flanagan, R.J.; Mitchell, R.J.; Karron, J.D. Increased relative abundance of an invasive competitor for pollination, Lythrum salicaria, reduces seed number in Mimulus ringens. Oecologia 2010, 164, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Hovivk, S.M.; Bunker, D.E.; Peterson, C.J.; Carson, W.P. Purple loosestrife suppresses plant species colonization far more than broad-leaved cattail: Experimental evidence with plant community implications. J. Ecol. 2011, 99, 225–234. [Google Scholar] [CrossRef]
- Dávalos, A.; Blossey, B. Matrix habitat and plant damage influence colonization of purple loosestrife patches by specialist leaf-beetles. Environ. Entomol. 2011, 40, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Boag, A.E.; Eckert, C.G. The effect of host abundance on the distribution and impact of biocontrol agents on purple loosestrife (Lythrum salicaria, Lythraceae). Écoscience 2013, 20, 90–99. [Google Scholar] [CrossRef]
- Rogers, J. Controlling purple loosestrife (Lythrum salicaria) along roadsides in St. Lawrence County: Monitoring and biological controls. Adirond. J. Environ. Stud. 2019, 23, 5. [Google Scholar]
- Adams, C.R.; Hovick, S.M.; Anderson, N.O.; Kettenring, K.M. We can better manage ecosystems by connecting solutions to constraints: Learning from wetland plant invasions. Front. Environ. Sci. 2021, 9, 715350. [Google Scholar] [CrossRef]
- Blossey, B.; Skinner, L.C.; Taylor, J. Impact and management of purple loosestrife (Lythrum salicaria) in North America. Biodivers. Conserv. 2001, 10, 1787–1807. [Google Scholar] [CrossRef]
- Russell-Mercierand, J.L.; Sargent, R.D. Indirect effects of herbivory on plant pollinator interactions in invasive Lythrum salicaria. Am. J. Bot. 2015, 102, 1–8. [Google Scholar] [CrossRef]
- Shi, J.; Macel, M.; Tielbörger, K.; Verhoeven, K.J. Effects of admixture in native and invasive populations of Lythrum salicaria. Biol. Invas. 2018, 20, 2381–2393. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, C. The Relative Roles of Herbivore- and Pollinator-Mediated Selection on the Evolution of Floral Display in the Invasive Plant, Lythrum salicaria. Ph.D. Thesis, Université d’Ottawa/University of Ottawa, Ottawa, ON, Canada, 2015. [Google Scholar]
- Olsson, K.; Gren, J.A. Latitudinal population differentiation in phenology, life history and flower morphology in the perennial herb Lythrum salicaria. J. Evol. Biol. 2002, 15, 983–996. [Google Scholar] [CrossRef]
- Campos, A.C.A.L.; Van Dijk, W.F.A.; Ramakrishna, P.; Giles, T.; Korte, P.; Douglas, A.; Smith, P.; Salt, D.E. 1135 ionomes reveal the global pattern of leaf and seed mineral nutrient and trace element diversity in Arabidopsis thaliana. Plant J. 2021, 106, 536–554. [Google Scholar] [CrossRef]
- Jarčuška, B.; Slezák, M.; Hrivnák, R.; Senko, D. Invasibility of alien Impatiens parviflora in temperate forest understories. Flora 2016, 224, 14–23. [Google Scholar] [CrossRef]
- Schmitz, D.; Girardi, J.; Jamin, J.; Bundschuh, M.; Geng, B.; Feldmann, R.; Rösch, V.; Riess, K.; Schirmel, J. Copper Uptake and Its Effects on Two Riparian Plant Species, the Native Urtica dioica, and the Invasive Fallopia japonica. Plants 2023, 12, 481. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.; Acreman, M.C. Wetland nutrient removal: A review of the evidence. Hydrol. Earth Syst. Sci. 2004, 8, 673–685. [Google Scholar] [CrossRef]
- Uveges, J.L.; Corbett, A.L.; Mal, T.K. Effects of lead contamination on the growth of Lythrum salicaria (purple loosestrife). Environ. Pollut. 2002, 120, 319–323. [Google Scholar] [CrossRef]
- Nicholls, A.M.; Mal, T.K. Effects of lead and copper exposure on growth of an invasive weed, Lythrum salicaria L. (purple loosestrife). Ohio J. Sci. 2003, 103, 129–133. [Google Scholar]
- Zhou, Q.; Zhu, H.; Bañuelos, G.; Yan, B.; Liang, Y.; Yu, J.; Li, H. Impacts of vegetation and temperature on the treatment of domestic sewage in constructed wetlands incorporated with Ferric-Carbon micro-electrolysis material. Int. J. Phytoremediation 2017, 19, 915–924. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, B.; Ma, J. Comparison of heavy metal accumulation ability in rainwater by 10 sponge city plant species. Environ. Sci. Pollut. Res. 2019, 26, 26733–26747. [Google Scholar] [CrossRef]
- Zuo, X.; Zhang, H.; Yu, J. Microbial diversity for the improvement of nitrogen removal in stormwater bioretention cells with three aquatic plants. Chemosphere 2020, 244, 125626. [Google Scholar] [CrossRef] [PubMed]
- Dudek, T.; Wolański, P.; Rogut, K. The content of macro-and micro minerals in the sward of different types of semi-natural meadows of temperate climate in SE Poland. Agronomy 2020, 10, 273. [Google Scholar] [CrossRef]
- Muller, L.A.H.; Lambaerts, M.; Vangronsveld, J.; Colpaert, J.V. AFLP-based assessment of the effects of environmental heavy metal pollution on the genetic structure of pioneer populations of Suillus luteus. New Phytol. 2004, 164, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.A.; Vangronsveld, J.; Colpaert, J.V. Genetic structure of Suillus luteus populations in heavy metal polluted and nonpolluted habitats. Mol. Ecol. 2007, 16, 4728–4737. [Google Scholar] [CrossRef]
- Houdegbe, A.C.; Achigan-Dako, E.G.; Sogbohossou, E.O.; Schranz, M.E.; Odindo, A.O.; Sibiya, J. Leaf elemental composition analysis in spider plant [Gynandropsis gynandra L. (Briq.)] differentiates three nutritional groups. Front. Plant Sci. 2022, 3091. [Google Scholar] [CrossRef]
- Cao, F.; Chen, F.; Sun, H.; Zhang, G.; Chen, Z.-H.; Wu, F. Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley. BMC Genom. 2014, 15, 611. [Google Scholar] [CrossRef]
- Ansari, M.K.A.; Ahmad, A.; Umar, S.; Zia, M.H.; Iqbal, M.; Owens, G. Genotypic variation in phytoremediation potential of Indian mustard exposed to nickel stress: A hydroponic study. Int. J. Phytoremediation 2015, 17, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Liu, C.; Cao, B.; Qin, M.; Long, D.; Xiang, Z.; Zhao, A. Genome-Wide Identification and characterization of four gene families putatively involved in cadmium uptake, translocation and sequestration in mulberry. Front. Plant Sci. 2018, 9, 879. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.E. Analysis of vegetation and other organic materials. In Chemical Analysis of Ecological Materials, 2nd ed.; Allen, S.E., Ed.; Blackwell Scientific Publications: Oxford/London, UK, 1989; pp. 46–61. [Google Scholar]
- Kjeldahl, C. A new method for the determination of nitrogen in organic matter. J. Anal. Chem. 1883, 22, 366–372. [Google Scholar]
- Lujanienė, G.; Valiulis, D.; Byčenkienė, S.; Šakalys, J.; Povinec, P.P. Plutonium isotopes and 241Am in the atmosphere of Lithuania: A comparison of different source terms. Atmos. Environ. 2012, 61, 419–427. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, J.S.; Lee, J.M.; Lee, K.B.; Park, T.S.; Lujaniene, G.; Valiulis, D.; Šakalys, J. Distribution characteristics of 137Cs, Pu isotopes and 241Am in soil in Korea. Appl. Radiat. Isot. 2013, 81, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Lujanienė, G.; Šemčuk, S.; Lečinskytė, A.; Kulakauskaitė, I.; Mažeika, K.S.; Valiulis, D.; Pakštas, V.; Skapas, M.; Tumėnas, S. Magnetic graphene oxide based nano-composites for removal of radionuclides and metals from contaminated solutions. J. Environ. Radioact. 2017, 166, 166–174. [Google Scholar] [CrossRef] [PubMed]
- CLC. CORINE Land Cover Nomenclature Conversion to Land Cover Classification System. Available online: https://land.copernicus.eu/eagle/files/eagle-related-projects/pt_clcconversion-to-fao-lccs3_dec2010 (accessed on 30 December 2022).
- Gailiušis, B.; Jablonskis, J.; Kovalenkovienė, M. Lithuanian Rivers: Hydrography and Runoff; Lithuanian Energy Institute: Kaunas, Lithuania, 2001. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Bolker, B. DescTools: Tools for Descriptive Statistics; R Package Version 0.99.42; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Subirana, I.; Sanz, H.; Vila, J. Building Bivariate Tables: The compareGroups Package for R. J. Stat. Soft. 2014, 57, 1–16. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.90). Available online: https://github.com/taiyun/corrplot (accessed on 30 December 2022).
- Chapin, F.S. The mineral nutrition of wild plants. Ann. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Watanabe, T.; Broadley, M.R.; Jansen, S.; White, P.J.; Takada, J.; Satake, K.; Takamatsu, T.; Tuah, S.J.; Osaki, M. Evolutionary control of leaf element composition in plants. New Phytol. 2007, 174, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef]
- Kozma, A.; Tölgyesi, G. Plant associations of flood plains along the Middle Tisza and their agricultural utilization. Tiscia 1979, 14, 105–122. [Google Scholar]
- Tölgyesi, G.; Kozma, A. Taxonomic ans ecological comments relating macro- and micro-element concentrations in plant species of inundation area. Tiscia 1983, 18, 71–74. [Google Scholar]
- Emery, S.L.; Perry, J.A. Aboveground biomass and phosphorus concentrations of Lythrum salicaria (Purple loosestrife) and Typha spp. (Cattail) in 12 Minnesota wetlands. Am. Midl. Nat. 1995, 394–399. [Google Scholar] [CrossRef]
- Vardanyan, L.; Schmieder, K.; Sayadyan, H.; Heege, T.; Heblinski, J.; Agyemang, T.; De, J.; Breuer, J. Heavy Metal Accumulation by Certain Aquatic Macrophytes from Lake Sevan (Armenia). In Proceedings of the Taal: The 12th World Lake Conference, Jaipur, India, 28 October–2 November 2008; pp. 1028–1038. [Google Scholar]
- Hubbard, A.R. Assessment of Trace Metal Concentrations in Lythrum salicaria at Three Rhode Island Sites Using ICP-MS Analysis. Master’s Thesis, Bryant University, Smithfield, RI, USA, 2015. [Google Scholar]
- Yue, K.; Peng, Y.; Fornara, D.A.; Van Meerbeek, K.; Vesterdal, L.; Yang, W.; Peng, C.; Tan, B.; Zhou, W.; Xu, Z.; et al. Responses of nitrogen concentrations and pools to multiple environmental change drivers: A meta-analysis across terrestrial ecosystems. Glob. Ecol. Biogeogr. 2019, 28, 690–724. [Google Scholar] [CrossRef]
- Wei, C.; Tang, S.; Pan, Y.; Li, X. Plastic responses of invasive Bidens frondosa to water and nitrogen addition. Nord J. Bot. 2017, 35, 232–239. [Google Scholar] [CrossRef]
- Beumer, V.; Ohm, J.N.; van Wirdum, G.; Beltman, B.; Griffioen, J.; Verhoeven, J.T.A. Biogeochemical plant site conditions in stream valleys after winter flooding: A phytometer approach. Biogeosci. Discuss. 2008, 5, 5203–5232. [Google Scholar] [CrossRef]
- Shamsi, S.R.A.; Whitehead, F.H. Comparative eco-physiology of Epilobium hirsutum L. and Lythrum salicaria L. III. Mineral nutrition. J. Ecol. 1977, 65, 55–70. [Google Scholar] [CrossRef]
- Krokaitė, E.; Shakeneva, D.; Juškaitytė, E.; Rekašius, T.; Nemaniūtė-Gužienė, J.; Butkuvienė, J.; Patamsytė, J.; Rančelienė, V.; Vyšniauskienė, R.; Duchovskienė, L.; et al. Nitrogen concentration of the aquatic plant species in relation to land cover type and other variables of the environment. Zemdirbyste 2019, 106, 203–212. [Google Scholar] [CrossRef]
- Yu, H.; Wang, L.; Liu, C.; Fan, S. Coverage of native plants is key factor influencing the invasibility of freshwater ecosystems by exotic plants in China. Front. Plant Sci. 2018, 9, 250. [Google Scholar] [CrossRef]
- Dostal, P.; Dawson, W.; van Kleunen, M.; Keser, L.H.; Fischer, M. Central European plant species from more productive habitats are more invasive at a global scale: Habitat productivity at origin-plant invasiveness elsewhere. Global Ecol. Biogeogr. 2013, 22, 64–72. [Google Scholar] [CrossRef]
- Legault, R.; Zogg, G.P.; Travis, S.E. Competitive interactions between native Spartina alterniflora and non-native Phragmites australis depend on nutrient loading and temperature. PLoS ONE 2018, 13, e0192234. [Google Scholar] [CrossRef]
- Yang, B.; Cui, M.; Du, Y.; Ren, G.; Li, J.; Wang, C.; Li, G.; Dai, Z.; Rutherford, S.; Wan, J.S.H.; et al. Influence of multiple global change drivers on plant invasion: Additive effects are uncommon. Front. Plant Sci. 2022, 13, 1020621. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Liu, Y.; Zhao, S.; Dai, L.; Zhang, Z.; Zhang, M.; Zhang, Z. How Waterlogged Conditions Influence the Nitrogen Dynamics in a Soil–Water–Plant System: Implications for Wetland Restoration. Water 2021, 13, 2957. [Google Scholar] [CrossRef]
- Tong, X.; Wang, X.; He, X.; Sui, Y.; Shen, J.; Feng, J. Effects of antibiotics on nitrogen uptake of four wetland plant species grown under hydroponic culture. Environ. Sci. Pollut. Res. 2019, 26, 10621–10630. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, S.; Yu, G. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Global Change Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2011. [Google Scholar]
- Istvan, P.; Benton, J. Trace Elements; Lucie Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Teuchies, J.; Jacobs, A.; Oosterlee, L.; Bervoets, L.; Meire, P. Role of plants in metal cycling in a tidal wetlands: Implications for phytoremediation. Sci. Total Environ. 2013, 445–446, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef]
- Tsonev, T.; Lidon, F.J.C. Zinc in plants—An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Aravind, P.; Prasad, M.N.V. Zinc alleviates cadmium-induced oxidative stress in Ceratophyllum demersum L.: A free floating freshwater macrophyte. Plant Physiol. Biochem. 2003, 41, 391–397. [Google Scholar] [CrossRef]
- Betül, A.K.I.N.; Bingöl, N.A. Heavy metal accumulation in wetland plants and water-sediment relationship in Köprüören-Kütahya. J. Limnol. Freshw. Fish. Res. 2019, 5, 76–82. [Google Scholar] [CrossRef]
- Bingöl, N.A.; Akin, B.; Kocaçalişkan, İ.L.; Nalbantoğlu, B.; Meşeli, O. Effect of zinc on phytoremediation potential and carbonic anhydrase and polyphenol oxidase activities of Lythrum salicaria L. Turk. J. Bot. 2021, 45, 553–562. [Google Scholar] [CrossRef]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [PubMed]
- Riyazuddin, R.; Nisha, N.; Ejaz, B.; Khan, M.I.R.; Kumar, M.; Ramteke, P.W.; Gupta, R. A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 2021, 12, 43. [Google Scholar] [CrossRef]
- Sperdouli, I. Heavy metal toxicity effects on plants. Toxics 2022, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, D.; Krivokapic, S. The effect of Cu, Zn, Cd, and Pb accumulation on biochemical parameters (proline, chlorophyll) in the water caltrop (Trapa natans L.), Lake Skadar, Montenegro. Plants 2020, 9, 1287. [Google Scholar] [CrossRef] [PubMed]
- Čeburnis, D.; Steinnes, E. Conifer needles as biomonitors of atmospheric heavy metal deposition: Comparison with mosses and precipitation, role of the canopy. Atmos. Environ. 2000, 34, 4265–4271. [Google Scholar] [CrossRef]
- Kupcinskiene, E. Foliar elements of Scots pine across the transect from the ammonia emission source (1995–1998). Ekológia 2003, 22, 171–175. [Google Scholar]
- Kuznetsova, T.; Lukjanova, A.; Mandre, M.; Lõhmus, K. Aboveground biomass and nutrient accumulation dynamics in young black alder, silver birch and Scots pine plantations on reclaimed oil shale mining areas in Estonia. For. Ecol. Manag. 2011, 262, 56–64. [Google Scholar] [CrossRef]
- Jocienė, L.; Krokaitė, E.; Rekašius, T.; Vilčinskas, R.; Judžentienė, A.; Marozas, V.; Kupčinskienė, E. Ionomic parameters of populations of common juniper (Juniperus communis L.) depending on the habitat type. Plants 2023, 12, 961. [Google Scholar] [CrossRef] [PubMed]
- Abolina, L.; Osvalde, A.; Karlsons, A. Habitat characteristics and mineral nutrition status of Rubus chamaemorus L. in Latvia. Plants 2023, 12, 528. [Google Scholar] [CrossRef]
- Fastovetska, K.; Šlepetiene, A.; Vigricas, E.; Urbaitis, G.; Belova, O. Lead content in plant materials in the buffer zones of surface water bodies of Northwestern and Central regions of Lithuania. Zemdirbyste 2022, 109, 335–340. [Google Scholar] [CrossRef]
- Bingöl, N.A.; Akin, B.; Ozmal, V. Phytoremediation and Biosorption Potential of Lythrum salicaria L. for Nickel Removal from Aqueous Solutions. Pol. J. Environ. Stud. Vol. 2017, 26, 2479–2485. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Yan, D. Chromium bioaccumulation and its impacts on plants: An overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueño, M.C.; Del Río, L.A.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-Y.; Salt, D.E. Plant ionomics: From elemental profiling to environmental adaptation. Mol. Plant. 2016, 9, 787–797. [Google Scholar] [CrossRef]
- Stein, R.J.; Höreth, S.; De Melo, J.R.F.; Syllwasschy, L.; Lee, G.; Garbin, M.L.; Clemens, S.; Krämer, U. Relationships between soil and leaf mineral composition are element-specific, environment-dependent and geographically structured in the emerging model Arabidopsis halleri. New Phytol. 2017, 213, 1274–1286. [Google Scholar] [CrossRef]
- Brezinová, T.; Vymazal, J. Nitrogen standing stock in Phragmites australis growing in constructed wetlands—Do we evaluate it correctly? Ecol. Eng. 2015, 74, 286–289. [Google Scholar] [CrossRef]
- Anderson, N.O.; Jocienė, L.; Krokaitė, E.; Rekašius, T.; Paulauskas, A.; Kupčinskienė, E. Genetic diversity of Phalaris arundinacea populations in relation to river regulation in the Merkys basin, Lithuania. River Res. Appl. 2018, 34, 1–10. [Google Scholar] [CrossRef]
- Butkuvienė, J.; Sinkevičienė, Z.; Naugžemys, D.; Žvingila, D. Floristic structure of two Batrachium rich plant communities in relation to habitat conditions in rivers of Lithuania. Pol. J. Ecol. 2018, 66, 1–13. [Google Scholar] [CrossRef]
- Vyšniauskienė, R.; Rančelienė, V.; Naugžemys, D.; Patamsytė, J.; Sinkevičienė, Z.; Butkuvienė, J.; Žvingila, D. Genetic diversity of populations of Bidens genera invasive and native species in Lithuania. Zemdirbyste 2018, 105, 183–190. [Google Scholar] [CrossRef]
- Vyšniauskienė, R.; Rančelienė, V.; Naugžemys, D.; Rudaitytė-Lukošienė, E.; Patamsytė, J.; Butkauskas, D.; Kupčinskienė, E.; Žvingila, D. Genetic diversity of Nuphar lutea in Lithuanian river populations. Aquat. Bot. 2020, 161, 103173. [Google Scholar] [CrossRef]
- Basic, N.; Besnard, G. Gene polymorphisms for elucidating the genetic structure of the heavy-metal hyperaccumulating trait in Thlaspi caerulescens and their cross-genera amplification in Brassicaceae. J. Plant Res. 2006, 119, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Prus-Glowacki, W.; Chudzinska, E.; Wojnicka-Poltorak, A.; Fagiewicz, K. Effects of heavy metal pollution on genetic variation and cytological disturbances in the Pinus sylvestris L. population. J. Appl. Genet. 2006, 47, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yan, Y.; Hu, W.; Liu, G.; Zeng, H.; Wei, Y.; Shi, H. Genome-wide association studies reveal genetic basis of ionomic variation in cassava. Plant J. 2022, 112, 1212–1223. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).