1. Introduction
The limestone hill at Zhoukoudian (Longgushan, “dragon bone hill”, now a UNESCO World Heritage Site) in the Fangshan district of the Beijing Municipality, about 50 km SW of the city of Beijing, became famous in the 1920s because of the palaeontological and palaeoanthropological discoveries that were made in the Pleistocene sediments which filled its many karstic cavities [
1]. What most attracted the attention of both scientists and the general public were Middle Pleistocene archaic humans, the so-called “Peking Man”, or
Sinanthropus (now considered as a representative of
Homo erectus). However, one of the localities at Zhoukoudian, the so-called “Upper Cave”, discovered in 1930 [
2], yielded a significantly younger faunal assemblage, together with various remains of anatomically modern
Homo sapiens and their stone and bone implements [
1,
2]. According to the latest radiocarbon dates available, an age of 35.1–33.5 ka is likely for the Upper Cave deposits [
3]. In the course of excavations carried out in the Upper Cave in 1933–1934, a large number of vertebrate fossil remains were unearthed, including some bird material (for lists of the avian species, see [
1,
4,
5]). The bird remains were first mentioned by Pei [
2], who noted the presence of two femora, ribs, and a vertebra of
Struthio. The avian specimens from all Zhoukoudian localities were described in a preliminary paper by Tsen-Hwang Shaw in 1935 [
4]. In it, he reported two ostrich femora from the Upper Cave. In 1937, Shaw [
6] published an additional short paper on these femora in which he compared them with those of recent ostriches (
Struthio camelus) and provided measurements but no illustrations. While one of the femora was incomplete, lacking the proximal half, the other one was complete. Shaw [
3,
4] referred them to the Late Pleistocene giant ostrich
Struthio anderssoni, described by Lowe [
7] on the basis of eggs from the Chinese loess [
8].
Subsequently, the complete femur was lost under unknown circumstances. There is no reason to believe that it was lost with all the human remains from Zhoukoudian during the ill-fated attempt to send the palaeoanthropological specimens to the United States at the end of 1941, as it is not in the list of the items that were boxed for shipping [
9]. According to Jia and Huang [
9], at the end of spring 1937, a number of fossils from the Cenozoic Research Laboratory, consisting mainly of mammal bones from Zhoukoudian but also including specimens from other sites, were packed to be shipped off for safety. The complete femur may have been part of them, although it is not listed in the published list [
9]. The specimens were eventually left in a storeroom at the Peking Union Medical College and were thrown away or destroyed by Japanese occupation forces in 1942 [
9]. Some looted specimens from the Upper Cave were found in Tokyo in 1945 and returned to China, but the complete ostrich femur is not in the published list of this material [
9]. Its whereabouts remain unknown. When Hou [
5] revised the avian fauna from Zhoukoudian, he could only give an illustrated description of the incomplete specimen. Recently, however, an excellent cast of the missing complete femur was identified in the palaeontology collection of the Natural History Museum, London. As discussed below, this cast provides significant new evidence about the large ostrich from the Upper Cave and highlights the scientific importance of old casts of fossils and the need to curate them properly.
Institutional abbreviations: IVPP: Institute of Vertebrate Paleontology and Paleoanthropology, Beijing; MNHN: Muséum National d’Histoire Naturelle, Paris; NHMUK: Natural History Museum, London.
2. The Cast and Its History
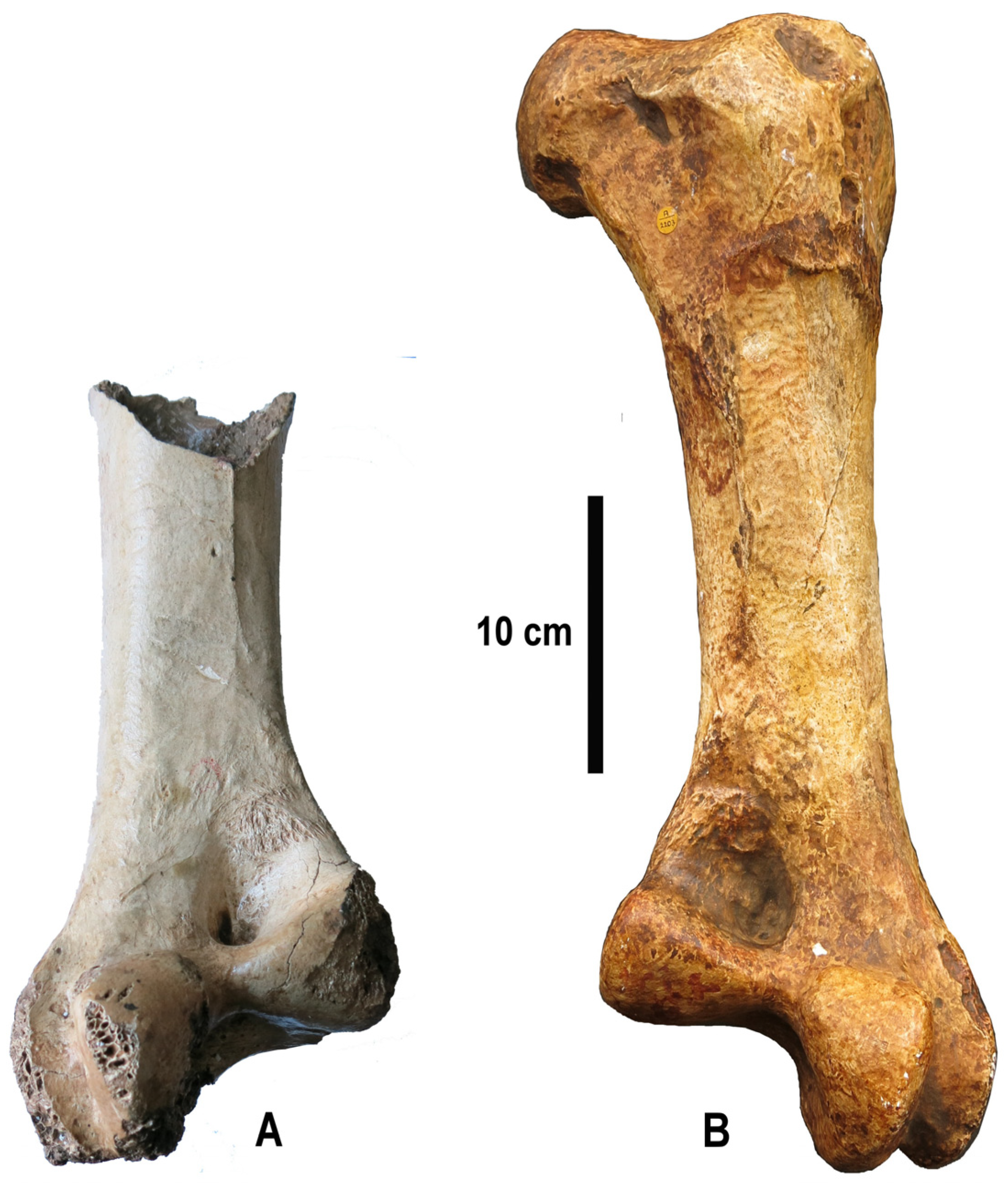
The cast of the missing complete ostrich femur from Zhoukoudian Upper Cave was identified as such by the author during a visit to the Natural History Museum (London) in March, 2022. It belongs to the collection of fossil birds and bears number PV A 2203. Its existence seems to have been largely forgotten, since it is not mentioned in any of the publications about the Zhoukoudian ostriches. It is a high-quality coloured plaster cast, showing only minimal traces of abrasion in a few places. Information about the cast is provided by both its label and an entry in the catalogue of the palaeontology collection. The label (
Figure 1), which contains various typing mistakes, describes the specimen (A 2203) as a right femur of
Struthio sp. Its origin is given as “Chen ‘Ken’ Tien cave near Peiping”. While Peiping is the name under which Beijing was known in the 1930s, “Chen ‘Ken’ Tien” is obviously a misspelling of Chou Kou Tien, the spelling that was used at that time, and was later replaced by the official
pinyin spelling Zhoukoudian. The label further mentions “cast made in museum, original in Peiping”. An entry in the catalogue of the palaeontology collection for the year 1937, in which the locality is correctly given as Chou Kou Tien, confirms that the cast was made “in the museum”.
The indication that the cast was made “in the museum” in 1937 is of special importance since it implies that the original fossil, which belonged to the collections of the Cenozoic Research Laboratory in Beijing, apparently under number K10 [
4], was brought to London where a plaster replica was made. Although the label and catalogue entry say nothing about this, the person who brought the specimen to London must have been the zoologist Tsen-Hwang Shaw (Shou Zhenhuang, 1899–1964) who, at that time, was working on the fossil ostrich material from the Upper Cave. In his short paper on the ostrich femora published on 13 December 1937, Shaw [
6] mentioned that he had compared the Zhoukoudian specimens with various
Struthio camelus femora from Africa and the Middle East (
Struthio camelus syriacus) kept at the British Museum (Natural History) and provided measurements of these bones. Despite the fact that there is no mention of a visit by Shaw in the visitors books of the zoology and geology departments of the BM(NH) for 1937 (S. Maidment, pers. com.), it seems clear that at some point during that year he was in London and compared the Chinese specimens with femora of recent ostriches and that he had brought the complete femur with him, as a result of which a cast was made at the museum. The only other option is that Shaw obtained measurements of ostrich bones from someone in London and did not visit the museum himself, but that does not explain how the original femur was sent to London for casting (and was subsequently returned to Beijing—since it did not remain in London). In this connection, it should be mentioned that in his 1937 paper, Shaw did not thank anybody in the BM(NH) for providing the ostrich bone measurements, while in other papers, he readily acknowledged the help of scientists from institutions in London and Berlin who sent him information [
10]; this suggests that he did study the ostrich femora in London himself. There is no mention of a visit to London in the recent short biography of Shaw by Wei and Wang [
11], but a search in Chinese archives may provide information about that trip. In the 1920s and 1930s, Shaw was working at the Fan Memorial Institute of Biology, one of the major scientific research institutions in Beijing at that time.
Beyond the information provided by the label and catalogue, the measurements of the cast perfectly fit those given by Shaw [
6] for the complete femur from the Upper Cave, which provides additional confirmation about the identity of the specimen.
3. A Brief Description of the Cast
Cast PV A 2203 corresponds to a complete, well preserved, right femur (
Figure 2). Shaw [
4] gave only a very brief description of it, focusing on a comparison of a few characters with those of the femur of
Struthio camelus and concentrating on measurements (which indicate a species significantly larger than
S. camelus). A more detailed description is given here. The anatomical terminology is that of Baumel and Witmer [
12].
PV A 2203 shows a massive trochanter femoris (
Figure 2, tf); the cranial face of which is slightly concave, while the caudal face is convex. The cranial margin of the facies articularis antitrochanterica forms a ridge which overhangs the cranial face of the trochanter. On the cranial face, the crista trochanteri (ct) is prominent. The caput femoris (cf) is large, well rounded, and bears a deep, groove-like fovea ligamenti capitis (flc). The collum femoris (cof) is relatively short. A pneumatic foramen (pf) is located on the caudal surface of the bone, close to its proximal margin. The diaphysis is robust and straight. A well- marked linea intermuscularis is present both on the cranial and on the caudal face. The linea intermuscularis cranialis (licr) runs in a slightly curved line, close to the lateral margin of the shaft, from the distal end of the crista trochanteri to the inception of the condylus lateralis (in this area. the surface of the bone is damaged, showing the cancellous tissue inside). The linea intermuscularis caudalis (lic) is rugose and close to the medial margin of the bone. It extends from the base of the collum to the area just proximal to the fossa poplitea, where it curves medialward. The fossa poplitea (fp) is rounded, not very deep, but wide, bordered proximomedially by a thickening of the bone which forms a protrusion along the medial margin of the femur. It is bounded distally by the pulley-like caudal part of the condylus medialis. The condylus medialis (cm) is smaller than the condylus lateralis (cl), which extends much farther distally than its medial counterpart. The condyles are separated by a deep sulcus intercondylaris (si). The condylus medialis projects more or less equally cranially and caudally and bears a depression on its medial surface. The condylus lateralis projects more strongly caudally than cranially and is subdivided into two semicondyles (to use Elzanowski’s terminology [
13]), which are separated by a deep and relatively wide cleft, the sulcus fibularis. The medial semicondylus tibiofibularis (st) is bulbous and broader than the lateral semicondylus fibularis (sf); the latter is relatively narrow and extends cranially into the epicondylus lateralis.
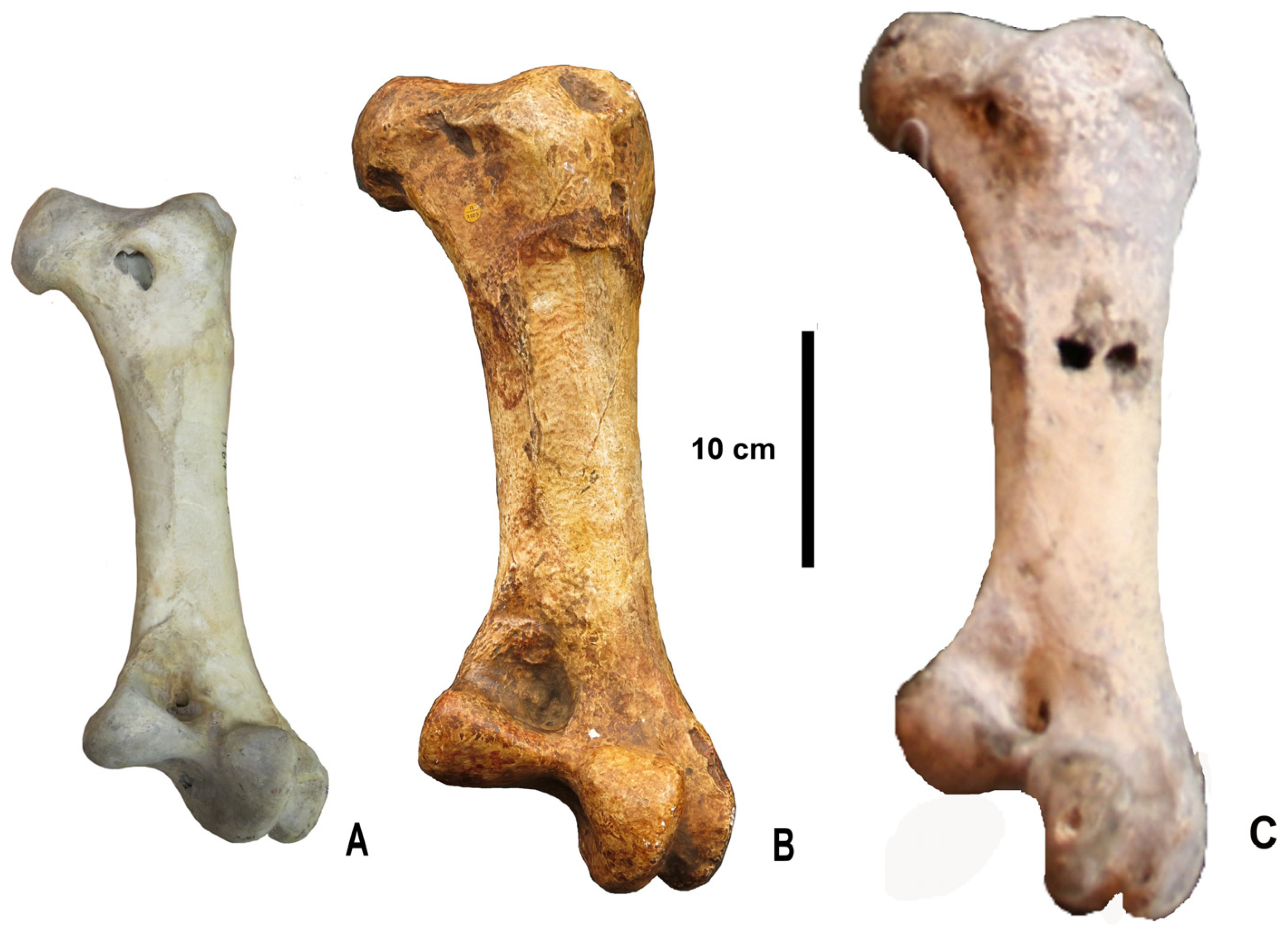
The complete femur from Zhoukoudian is very similar to the incomplete one (IVPP V6942) described and illustrated by Hou [
5]. Similarities include the strong distal projection of the lateral condyle, the broad but relatively shallow fossa poplitea, and the strongly marked lineae intermuscularis, as well as the general robustness (
Figure 3). There are slight differences in measurements; in particular, the minimum shaft circumference of IVPP V6942 is slightly larger than that of NHMUK PV A 2203. This may reflect individual variation or possibly sexual dimorphism, since a subtle sexual dimorphism has been reported in recent ostriches [
14], females having slightly more robust limb bones (although males are larger and heavier). However, it would be imprudent to draw far-reaching conclusions from only two bones.
The complete femur is generally similar to that of
Struthio camelus, as already noted by Shaw [
6], who however mentioned some morphological differences. He noted that in
S. camelus, the trochanter femoris had sharper edges, which is correct, and that the neck was stouter. This remark may seem puzzling in view of the greater general robustness of the femur from Zhoukoudian, but in proximal view, the collum does appear more constricted in the latter than in
S. camelus because the caput femoris is relatively larger, especially as compared with the area located lateral to the collum. This large caput femoris may be related to the great weight of the bird, necessitating an especially strong hip articulation (Shaw [
6] already remarked on the bulk of the articular head, which made it impossible to fit it within the acetabulum of the type pelvis of
Struthio wimani, described by Lowe [
7] from red beds in Shanxi, which are now considered as Late Miocene in age [
15]). In addition, it should be noted that the femur from Zhoukoudian is generally more robust than that of living ostriches and that its muscular insertions, notably the lineae intermuscularis, are very strongly marked, more so than in
S. camelus. This indicates an especially strong thigh musculature in keeping with the robustness of the bone and the great mass of the bird. Other differences are the greater distal projection of the lateral condyle and the wider, but relatively shallow, fossa poplitea in the Zhoukoudian femora (
Figure 4).
Shaw [
6] considered the measurements of the Zhoukoudian femur as especially noteworthy because they indicated a bird significantly larger than the living
Struthio camelus, as shown in his table comparing various measurements of the femur from Zhoukoudian and femora of several
S. camelus specimens (belonging to several subspecies, including
S. molybdophanes, now considered as a distinct species, and the recently extinct
S. camelus syriacus) in the London collection, as well as one specimen of
S. camelus spatzi in the Berlin Natural History Museum. Cast PV A 2203 confirms Shaw’s measurements, including a length of 355 mm, noticeably exceeding that of the longest femur in the
S. camelus collection used for comparison (311 mm). A measurement, not provided by Shaw, was the minimum circumference of the shaft, which is 187 mm. This measurement is important because it can be used to estimate the mass of the bird. A previous estimate by Buffetaut and Angst [
16] using Campbell and Marcus’s equation [
17], based on the incomplete left femur from Zhoukoudian kept at the IVPP in Beijing (collection number V 6943) with a minimum shaft diameter of 190 mm, was 270 kg. Using the same method for a minimum shaft diameter of 187 mm, the estimated mass is 258 kg. Although this result is slightly lower than that obtained for the incomplete femur, it is well above the maximum mass recorded for the largest male
S. camelus (156 kg). These results confirm Shaw’s conclusion that the Zhoukoudian ostrich was at least 1.5 times larger than the largest
Struthio camelus.
4. Identification of the Zhoukoudian Ostrich and Comparison with Other Ostriches
Shaw [
4] identified the ostrich bones from the Upper Cave as
Struthio andersoni [sic] Lowe without explaining the reasons for that attribution. In 1937 [
6], he remarked that the specimens were too large to be attributed to
Struthio wimani, which in any case came from older (Miocene) deposits than those of the Upper Cave, and confirmed their attribution to
S. anderssoni. Apparently, although he did not elaborate on this, Shaw referred the Upper Cave specimens to
Struthio anderssoni largely for stratigraphic reasons: the large eggs on which Lowe [
7] had based the species
S. anderssoni came from the Late Pleistocene loess and were therefore roughly contemporaneous with the Upper Cave sediments. Moreover, Young [
18] had reported the occurrence of
S. anderssoni eggshell fragments at Zhoukoudian (albeit in one of the
Sinanthropus localities—they were later reported from the Upper Cave deposits, too [
1,
5]).
Although it is obviously difficult to unequivocally refer skeletal elements to a species originally described exclusively on eggshell material, there are good reasons to refer the femora from Zhoukoudian to
Struthio anderssoni. In addition to the above-mentioned stratigraphic congruence, the mass estimates obtained by Buffetaut and Angst [
16] for
Struthio anderssoni on the basis of eggshell measurements and those derived from the minimum shaft circumference of the Zhoukoudian femora are in good agreement, indicating ostriches about 150 % the mass of the living
S. camelus (approximately 250–270 kg). There are therefore solid grounds to assume that the Zhoukoudian femora do belong to
Struthio anderssoni.
The taxonomic status of
Struthio anderssoni may also be reconsidered on the basis of morphological information provided by the cast of the complete Zhoukoudian femur. On the basis of eggshell evidence, Mikhailov and Zelenkov [
19] stated that “we should regard
Str. anderssoni as late Pleistocene Asian populations of the same biological ostrich species” as
Struthio camelus. This supposition seems to be hardly tenable in view of the significantly larger size and different morphological features indicated by the skeletal material from Zhoukoudian. As noted by Buffetaut and Angst [
15], for the complete
Struthio anderssoni femur from the Upper Cave, the stoutness index, as defined by Burchak-Abramovich and Vekua [
20] (minimum shaft width/total length ratio), is 19.44, whereas it does not exceed 16.4 in
S. camelus.
Struthio anderssoni clearly was a very large and robust species which can hardly be considered as a simple local variant of
Struthio camelus.
This raises the question of the possible referral of
Struthio anderssoni to an ostrich genus distinct from
Struthio. In the Early Pleistocene, remarkably large and robust ostriches lived in China, as exemplified by the femur from the Nihewan basin described by Buffetaut and Angst [
15]. They have been referred to the genus
Pachystruthio, first described as a distinct genus or a subgenus of
Struthio by Kretzoi [
21] on the basis of material from the Lower Pleistocene of Hungary including a phalanx, eggshell fragments, and “copro-uroliths”.
Pachystruthio was considered as a distinct genus, rather than a subgenus, by Zelenkov et al. [
22]. It had a very wide geographical distribution in the Early Pleistocene, being recorded in Hungary, Crimea, Georgia, and China [
15,
19]. Considering the large size and robustness of the Upper Cave ostrich, attribution to
Pachystruthio rather than
Struthio may be considered. Mass estimates for
Pachystruthio are higher than those obtained for
Struthio anderssoni; a mass of 300 kg was obtained on the basis of the minimum shaft circumference of the femur from Nihewan [
15], significantly more than the mass estimated on the basis of the complete Upper Cave femur. The mass estimates based on the
Pachystruthio femur from Taurida cave in Crimea are even higher, ranging from 453 to 462 kg, depending on the method used [
22]. These estimates suggest that
Struthio anderssoni was less bulky than Early Pleistocene
Pachystruthio. However, morphologically, the femora of
Struthio anderssoni and
Pachystruthio dmanisensis do not differ markedly (see
Figure 4 and figures of nearly complete
P. dmanisensis femora in Vekua [
23]). Notable common characters are the strong lineae intermuscularis, the wide but relatively shallow fossa poplitea, and the marked distal protrusion of the lateral condyle (
Figure 4). Their stoutness indices are similar (19 to 20), according to Buffetaut and Angst [
15], although the total length of the femur is greater in
P. dmanisensis (380 mm in a specimen from Dmanisi, Georgia) than in
S. anderssoni from Zhoukoudian. The main difference between
S. anderssoni and
Pachystruthio thus seems to lie in the greater size and mass of the latter rather than in morphological differences or proportions. The question may then be asked whether
Struthio anderssoni should in fact be considered as a Late Pleistocene species of
Pachystruthio, smaller than the Early Pleistocene
Pachystruthio dmanisensis but larger and more robustly built than
Struthio camelus.
Eggshell evidence may seem difficult to reconcile with this hypothesis; however, the only locality where eggshell fragments are clearly associated with skeletal remains of
Pachystruthio is Kisláng in Hungary [
19,
21]. The thick eggshell fragments (thickness 2.6–3.4 mm vs 2.1–2.4 mm in
S. anderssoni) were not illustrated or described in detail, but Mikhailov and Zelenkov [
19] assumed that they may belong to their “A” type (defined on the basis of the pore pattern). On the other hand, the eggshells of
Struthio anderssoni are relatively thin (although the eggs are larger than those of recent
Struthio) and belong to the “S” type according to Mikhailov and Zelenkov [
19]. These differences would seem to make an assignment of
S. anderssoni to the genus
Pachystruthio difficult. However, uncertainties remain as to the exact type of eggshell associated with
Pachystruthio, and as shown by Mikhailov and Zelenkov’s detailed study, ostrich eggshell evolution in Eurasia was a highly complex process, and intermediate types have been reported. The “S” type has been interpreted as an adaptation to arid conditions [
19], and the Late Pleistocene ostriches of Northern China lived in a cold environment, which was probably dryer (as evidenced by loess deposition) than the Early Pleistocene climate. This may explain why the Late Pleistocene ostrich eggs from northeastern Asia belong to type S. In addition, the smaller thickness of the eggs may simply be related to the relatively smaller size of
Pachystruthio anderssoni, since eggshell thickness is partly linked to the mass of the parent birds [
19].
While it cannot be denied that eggshell evidence can provide important information about ostrich evolution, especially because eggshell remains are much more abundant than bones, the evidence is not always easy to interpret, especially when there is no clear association between eggs and skeletal material. When skeletal remains are available, osteological evidence should probably be considered as more reliable than oological evidence for systematic and phylogenetic purposes for the simple reason that skeletal characters, rather than eggshell features, are the main evidence used in avian palaeontology and are probably less subject to environment-induced variation. The present case is made more complicated by the fact that Struthio anderssoni is an egg-based taxon. However, as noted above, there are various good reasons to refer the femora from the Upper Cave to that taxon.
As noted above, there is convincing osteological evidence to consider that “
Struthio”
anderssoni should in fact be placed in the genus
Pachystruthio. An evolution from the extremely large
Pachystruthio dmanisensis of the Early Pleistocene to the relatively smaller (but still very large)
P. anderssoni of the Late Pleistocene would imply a certain amount of size reduction and eggshell thinning (and possibly changes in pore pattern). Examples of such a size reduction from the Early to the Late Pleistocene are known in other groups of large tetrapods, such as the mammoths, with a decrease in size from the very large
Mammuthus meridionalis and
M. trogontherii to the smaller
M. primigenius [
24]. A discussion of the causes of this process is beyond the scope of the present paper, but it is possible that a similar reduction in size took place in the genus
Pachystruthio, resulting in the relatively smaller size of the Late Pleistocene species.
The genus Pachystruthio, as envisioned here, therefore includes the following species:
- -
Pachystruthio pannonensis Kretzoi, 1954, the type species from the Early Pleistocene of Hungary, known from a phalanx, referred eggshell fragments, and “copro-uroliths” [
21].
- -
Pachystruthio dmanisenis, Burchak-Abramovich and Vekua, 1990, from the Early Pleistocene of Georgia [
20,
23] and Crimea [
22], known from femora. It cannot be excluded that
Pachystruthio dmanisenis is in fact a junior synonym of
P. pannonensis [
20].
- -
Pachystruthio indet from the Early Pleistocene of Northern China, known from a femur [
15].
- -
Pachystruthio anderssoni (Lowe, 1931) from the Late Pleistocene of Northern China, Mongolia, and Siberia, known from femora (Zhoukoudian) and eggshell material, including complete eggs.
5. Conclusions
The identification of cast PV A 2203 as a replica of the lost complete ostrich femur from the Upper Cave at Zhoukoudian makes it possible to study in some detail a specimen that had never been figured and had only very briefly been described and complements, in important respects, the incomplete femur from the same locality still preserved in Beijing. This leads to a reappraisal of the femoral morphology of “Struthio” anderssoni, a species that appears to be clearly distinct from living ostriches and should be placed in the genus Pachystruthio.
More generally, cast PV A 2203 highlights the importance of casts of significant fossils for palaeontological research, especially in cases when the original specimen has been lost or destroyed. This was already emphasized by Buffetaut et al. [
25] about the type series of the giant bird species
Aepyornis maximus, the originals of which cannot currently be located, while casts of them are kept in various museums in France and England. Similarly, recently rediscovered casts of a historically important ichthyosaur specimen, destroyed in World War II, have provided significant information and testify to the usefulness of such old replicas [
26].
While the cast of the missing ostrich femur from Zhoukoudian was perfectly well curated at the Natural History Museum, old palaeontological casts tend to be neglected in some museums, often because their significance and potential scientific importance are unrecognised or underrated. This, in some instances, can lead to inadequate curation, loss of information about the originals, and eventually damage to the casts. Although they obviously cannot provide some types of information, notably about the internal structure or chemical composition, good quality casts are a precious source of morphological information, especially when the original specimens were not illustrated or were only summarily described (as in the case of the Zhoukoudian femur). They are of course of special importance when the originals have been lost or destroyed. Replicas of fossils are an important part of any palaeontological collection and should be properly taken care of. In addition, one of the main aims of producing casts has always been to disseminate them in order to make them available for display, study, and comparison outside the institution where the originals are kept. Both the proper curation of old replicas and the making (using new as well as traditional techniques) and diffusion of new ones should be encouraged.






{kind=link}
{kind=link}
{kind=link}
{kind=link}