

Liverwort Diversity in Cambodia: New Records Show There Is Still Much to Learn




Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Species List
3.2. Statistics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ingerpuu, N.; Vellak, K.; Hagborg, A.; Söderström, L. The Angkor Wat Kingdom—Liverworts from Cambodia. Nova Hedwig. Beih. 2020, 150, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Bakalin, V.; Choi, S.S.; Park, S.J. Review of Heteroscyphus Schiffn. (Lophocoleaceae, Marchantiophyta) in Cambodia. J. Bryol. 2021, 43, 52–61. [Google Scholar] [CrossRef]
- Ellis, L.T.; Ah-Peng, C.; Aslan, G.; Bakalin, V.A.; Bergamini, A.; Callaghan, D.A.; Campisi, P.; Raimondo, F.M.; Choi, S.S.; Csiky, J.; et al. New national and regional bryophyte records, 65. J. Bryol. 2021, 43, 67–91. [Google Scholar] [CrossRef]
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; Costa, D.P.; Crandall-Stotler, B.J.; et al. World Checklist of Hornworts and Liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [PubMed]
- Climate-Data: Phnom Penh. Available online: https://en.climate-data.org/asia/cambodia/phnom-penh/phnom-penh-4857/ (accessed on 6 February 2023).
- Climate-Data: Sen Monorom. Available online: https://en.climate-data.org/asia/cambodia/mondulkiri/sen-monorom-3147/ (accessed on 6 February 2023).
- Váňa, J.; Inoue, H. Studies in Taiwan Hepaticae, V. Jungermanniaceae. Bull. Natl. Sci. Mus. Tokyo Ser. B 1983, 9, 125–142. [Google Scholar]
- Shaw, B.; Crandall-Stotler, B.; Váňa, J.; Stotler, R.E.; von Konrat, M.; Engel, J.J.; Davis, E.C.; Long, D.G.; Sova, P.; Shaw, A.J. Phylogenetic relationships and morphological evolution in a major clade of leafy liverworts (phylum Marchantiophyta, order Jungermanniales): Suborder Jungermanniineae. Syst. Bot. 2015, 40, 27–45. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A.; Choi, S.S.; Nguyen, V.S. Blepharostoma trichophyllum s.l. (Marchantiophyta): The Complex of Sibling Species and Hybrids. Plants 2020, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Bakalin, V.A.; Vilnet, A.; Klimova, K.G.; Ma, W.Z.; Choi, S.S.; Hentschel, J. Hidden in plain view: An example from Ptilidium (Ptilidiaceae, Marchantiophyta). Phytotaxa 2021, 510, 29–42. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A.; Maltseva, Y.D.; Klimova, K.G.; Bakalin, D.A.; Choi, S.S. Hidden Diversity within Tetralophozia filiformis (Marchantiophyta, Anastrophyllaceae) in East Asia. Plants 2022, 11, 3121. [Google Scholar] [CrossRef] [PubMed]
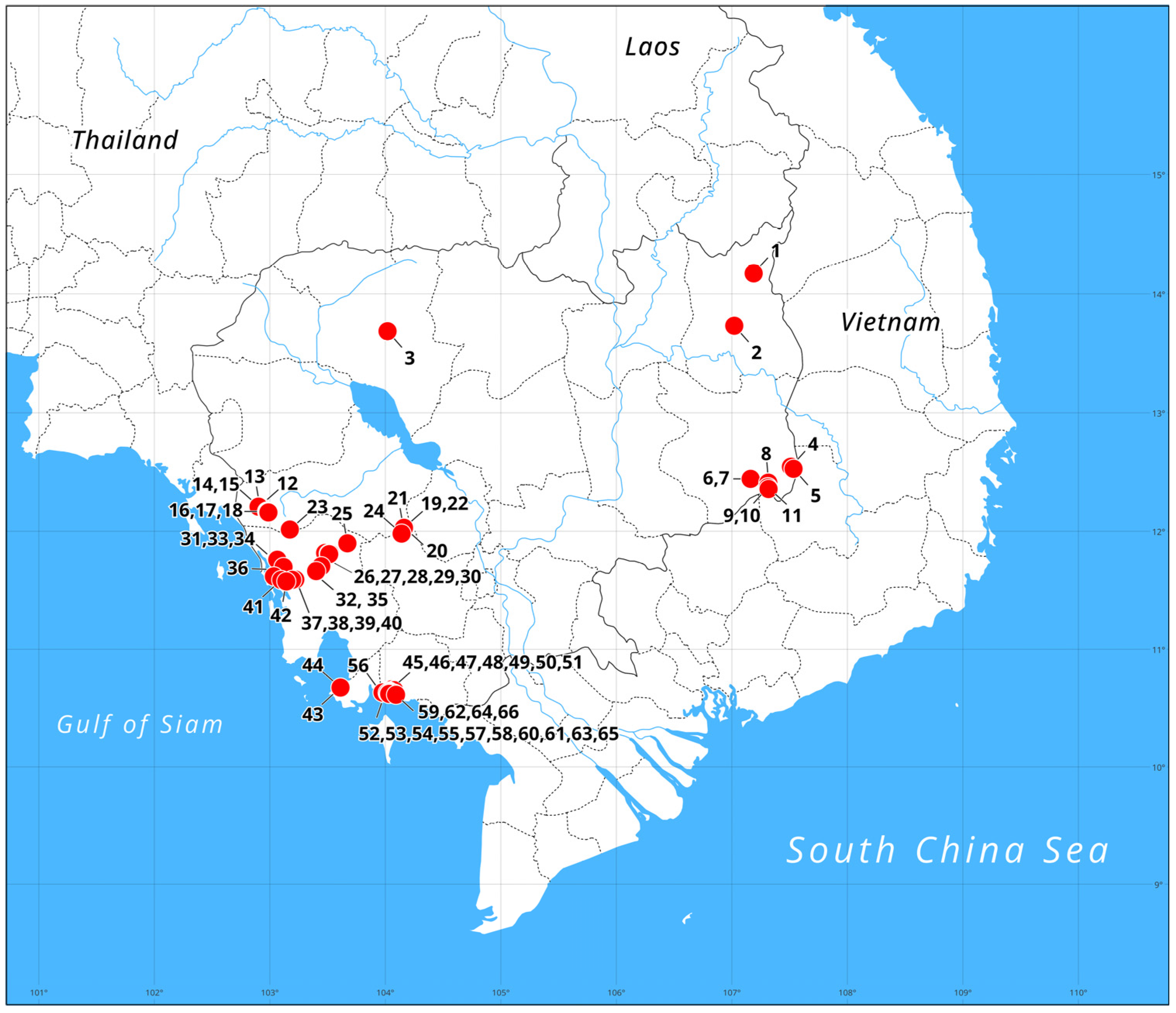




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Dates | Province Name | Total Number of Specimens |
|---|---|---|
| 5–8 December 2009 | Koh Kong | 160 |
| 10–11 December 2009 | Kampot | 182 |
| 13 December 2009 | Pursat | 9 |
| 4–7 May 2010 | Kampot | 246 |
| 8–9 May 2010 | Koh Kong | 56 |
| 15 December 2010 | Stung Treng | 17 |
| 16–18 December 2010 | Ratankiri | 60 |
| 20 December 2010 | Siem Reap | 2 |
| 17–20 December 2011 | Modulkiri | 441 |
| 22–26 December 2011 | Koh Kong | 551 |
| 27 December 2011 | Preah Sihanouk | 7 |
| 28–30 December 2011 | Kampot | 489 |
| 1–3 January 2012 | Pursat | 19 |
| 10–12 January 2013 | Kampong Speu | 142 |
| 15–18 January 2013 | Pursat | 148 |
| 22 January 2013 | Kampot | 77 |
| Number in the Map | Northern Latitude | Eastern Longitude | Province Name |
|---|---|---|---|
| 1 | 14.17100000 | 107.1861944 | Ratankiri |
| 2 | 13.73147222 | 107.0182222 | Ratankiri |
| 3 | 13.68538889 | 104.0163611 | Siem Reap |
| 4 | 12.54444444 | 107.5086111 | Modulkiri |
| 5 | 12.52686111 | 107.5312222 | Modulkiri |
| 6 | 12.44194444 | 107.1602778 | Modulkiri |
| 7 | 12.44191667 | 107.1598333 | Modulkiri |
| 8 | 12.40972222 | 107.3127778 | Modulkiri |
| 9 | 12.37194444 | 107.3091667 | Modulkiri |
| 10 | 12.37091667 | 107.3093611 | Modulkiri |
| 11 | 12.35611111 | 107.3150000 | Modulkiri |
| 12 | 12.21166667 | 102.9100000 | Pursat |
| 13 | 12.21161111 | 102.9103889 | Pursat |
| 14 | 12.21044444 | 102.8998333 | Pursat |
| 15 | 12.20697222 | 102.8995833 | Pursat |
| 16 | 12.16647222 | 102.9731389 | Pursat |
| 17 | 12.16313889 | 102.9796111 | Pursat |
| 18 | 12.15813889 | 102.9850278 | Pursat |
| 19 | 12.03241667 | 104.1710278 | Kampong Speu |
| 20 | 12.02988889 | 104.1629722 | Kampong Speu |
| 21 | 12.02852778 | 104.1535000 | Kampong Speu |
| 22 | 12.02833333 | 104.1588333 | Kampong Speu |
| 23 | 12.01375000 | 103.1704167 | Pursat |
| 24 | 11.97930556 | 104.1378611 | Kampong Speu |
| 25 | 11.89722222 | 103.6686944 | Koh Kong |
| 26 | 11.81555556 | 103.4761111 | Koh Kong |
| 27 | 11.81141667 | 103.5187222 | Koh Kong |
| 28 | 11.80880556 | 103.5067222 | Koh Kong |
| 29 | 11.80527778 | 103.5108333 | Koh Kong |
| 30 | 11.80500000 | 103.5110000 | Koh Kong |
| 31 | 11.75836111 | 103.0619444 | Koh Kong |
| 32 | 11.70336111 | 103.4423889 | Koh Kong |
| 33 | 11.69644444 | 103.1160278 | Koh Kong |
| 34 | 11.69638889 | 103.1158333 | Koh Kong |
| 35 | 11.66200000 | 103.3984722 | Koh Kong |
| 36 | 11.61783000 | 103.0323100 | Koh Kong |
| 37 | 11.59694444 | 103.2255556 | Koh Kong |
| 38 | 11.59158333 | 103.2244167 | Koh Kong |
| 39 | 11.59058333 | 103.2177778 | Koh Kong |
| 40 | 11.58800000 | 103.1910000 | Koh Kong |
| 41 | 11.58700000 | 103.0960000 | Koh Kong |
| 42 | 11.57447222 | 103.1398333 | Koh Kong |
| 43 | 10.67447222 | 103.6089722 | Preah Sihanouk |
| 44 | 10.67400000 | 103.6090000 | Preah Sihanouk |
| 45 | 10.66236111 | 104.0484444 | Kampot |
| 46 | 10.65900000 | 104.0510000 | Kampot |
| 47 | 10.65892000 | 104.0512800 | Kampot |
| 48 | 10.65891667 | 104.0512778 | Kampot |
| 49 | 10.65888900 | 104.0513890 | Kampot |
| 50 | 10.65230556 | 104.0741667 | Kampot |
| 51 | 10.65208333 | 104.0731389 | Kampot |
| 52 | 10.64164000 | 104.0304700 | Kampot |
| 53 | 10.64163889 | 104.0304722 | Kampot |
| 54 | 10.63341667 | 104.0335278 | Kampot |
| 55 | 10.63297222 | 104.0348889 | Kampot |
| 56 | 10.63216667 | 103.9730833 | Kampot |
| 57 | 10.62953000 | 104.0178600 | Kampot |
| 58 | 10.62952778 | 104.0178611 | Kampot |
| 59 | 10.62633000 | 104.0498600 | Kampot |
| 60 | 10.62378000 | 104.0273900 | Kampot |
| 61 | 10.62377778 | 104.0273889 | Kampot |
| 62 | 10.62333333 | 104.0727778 | Kampot |
| 63 | 10.62119444 | 104.0315833 | Kampot |
| 64 | 10.62116667 | 104.0783611 | Kampot |
| 65 | 10.62038889 | 104.0303889 | Kampot |
| 66 | 10.61269444 | 104.0898056 | Kampot |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Choi, S.S.; Park, S.J. Liverwort Diversity in Cambodia: New Records Show There Is Still Much to Learn. Diversity 2023, 15, 241. https://doi.org/10.3390/d15020241
Bakalin VA, Choi SS, Park SJ. Liverwort Diversity in Cambodia: New Records Show There Is Still Much to Learn. Diversity. 2023; 15(2):241. https://doi.org/10.3390/d15020241
Chicago/Turabian StyleBakalin, Vadim A., Seung Se Choi, and Seung Jin Park. 2023. "Liverwort Diversity in Cambodia: New Records Show There Is Still Much to Learn" Diversity 15, no. 2: 241. https://doi.org/10.3390/d15020241
APA StyleBakalin, V. A., Choi, S. S., & Park, S. J. (2023). Liverwort Diversity in Cambodia: New Records Show There Is Still Much to Learn. Diversity, 15(2), 241. https://doi.org/10.3390/d15020241