Abstract
Mangroves and seagrasses present with high marine macroinvertebrate biodiversity that contributes to their structure and functioning. Macroinvertebrates possess a broad range of functional traits, making them excellent models for biodiversity and available-trait-based studies. This study aimed to characterize the biodiversity of marine macroinvertebrates as two different ecosystems situated along the coastline of Maputo Bay by compiling dispersed data from online databases. Specifically, this study addressed species richness, taxonomic and functional diversity based on two traits (habitat occupation and trophic guild), and the community structure of these traits. Mangroves presented with a higher species richness and taxonomic diversity than seagrasses. The functional diversity of mangroves was mostly explained by the trophic guild trait. In the case of seagrasses, functional diversity was mostly due to differences in habitat occupation in the 20th century, but the trophic guild accounted for this functional diversity from 2000 onwards. The comparison of community compositions between the two ecosystems showed low or no similarity. The use of digital databases revealed some limitations, mostly regarding the sampling methods and individual counts. The trends and data gaps presented in this study can be further used to inform subsequent systematic data acquisition and support the development of future research. A further step that may be taken to improve the use of digital data in future biodiversity studies is to fully incorporate functional traits, abundance and sampling methods into online databases.
1. Introduction
Mangrove forests and seagrass meadows play significant roles in marine and coastal ecosystem integrity, providing a vast number of goods and services which are highly valued by local populations [1,2,3,4]. Mangroves and seagrasses have different structural characteristics [5,6], and they may be ecologically linked in some geographical areas, contributing to the exchange of species and ecosystem processes [6,7]. Despite the importance of mangroves and seagrasses, they are decreasing due to habitat occupation, degradation and fragmentation. This may result in a significant decrease in their associated biodiversity and ecosystem functioning [1,2,8]. The loss of mangroves and seagrasses changes coastlines in terms of the sediment characteristics and stability, as well as benthic communities, threatening, for example, costal protection, nursery areas, fisheries, nutrient cycling, water flow and carbon sequestration [8,9].
Marine macroinvertebrate biodiversity, e.g., crustaceans, echinoderms, molluscs, polychaete worms, nematodes, coelenterates, tunicates and sponges [3,4], contributes to the structure and stability of ecosystem processes such as productivity, ecological networks, nutrient and biogeochemical cycling and stability, as shown in [10,11]. Macroinvertebrates possess a broad range of functional traits, making them excellent models for biodiversity and functional-trait-based studies [12]. Nevertheless, there are few studies addressing the macrobenthos functional diversity and community composition of seagrasses [13,14] and faunal diversity of mangroves [15].
Taxonomic diversity studies examining the functional traits of species can shed light on the consequences of habitat loss or changes regarding ecosystem functioning. An increasing number of studies have applied a combination of taxonomic-diversity- and trait-based measures to characterize marine communities [16]. The functional traits of species influence ecosystem properties and, as such, provide an essential link between taxonomic diversity and ecosystem functioning [17]. Functional diversity characterises the roles of species within communities based on their morphological, behavioural and life history traits [17], and these traits are representative of the functional structure of a community [18].
Maputo Bay is the largest coastal embayment in Mozambique [19], with an area of 1280 km2 [20]. The size of the Bay, the diversity of its ecosystems and the services they provide are fundamental to supporting the subsistence of a large and growing human population [19]. In Maputo Bay, mangrove environments occupy 14% of the bay [20,21]. They are used as nursery areas by numerous invertebrate species, with crabs accounting for most of the macrobenthic faunal composition, followed by gastropod molluscs [4]. Seagrass meadows occupy 3% of Maputo Bay [20,21], and their marine macroinvertebrate faunal composition is dominated by echinoderms, molluscs and crustaceans [3]. Many species also use this ecosystem as a nursery area.
Digital data available via online databases, such as the Global Biodiversity Information Facility (GBIF, http://www.gbif.org (accessed on 27 October 2022)), mobilize biodiversity data from natural history collections (NHC), surveys and other sources [22], gathering relevant information on the world’s biodiversity, both historical and current [23]. Online biodiversity databases cover extensive geographic and temporal data that can be readily used as occurrence data. These data can be supplemented with other information, such as habitat and species traits, obtained from georeferenced information and previously described functional traits. Dependable datasets on global mangrove tree diversity are available, but no such information exists for the associated fauna species composition, functional diversity and functional redundancy [24]. Although many studies have analysed biodiversity using digital data [25,26,27,28,29], as far as we know, there are no studies addressing marine macroinvertebrate biodiversity and functional diversity using online digital data.
This study aimed to characterize the biodiversity of marine macroinvertebrates (Annelida, Arthropoda, Cnidaria, Echinodermata and Mollusca) in two different ecosystems (mangroves and seagrass meadows) along the coastline of Maputo Bay in Mozambique by compiling dispersed data from online databases. We hypothesized that digital databases can be used as tools to provide primary data for biodiversity research. Specifically, this study addressed (i) different diversity metrics, namely species richness and taxonomic and functional diversity, based on two traits (habitat occupation and trophic guild) and the community composition regarding the diversity of these traits, and (ii) data limitations affecting the application of diversity indexes. This work provides integrated baseline information on the general patterns of taxonomic and functional diversity and community composition of marine macroinvertebrates in mangroves and seagrasses, which can be used as a starting point for future studies.
2. Materials and Methods
2.1. Study Area
Maputo Bay is situated between the latitudes 25°72′10″–26°28′30″ S and longitudes 32°49′10″–32°85′40″ E (Figure 1), with an area of 1280 km2 (approximately 90 km in coastal length by 32 km in width [20,30]). The ecosystems considered for this study were mangroves (including the associated supratidal bare flats, the area between the landward margin of the mangrove forests and the beginning of the terrestrial vegetation) and seagrass meadows [21].
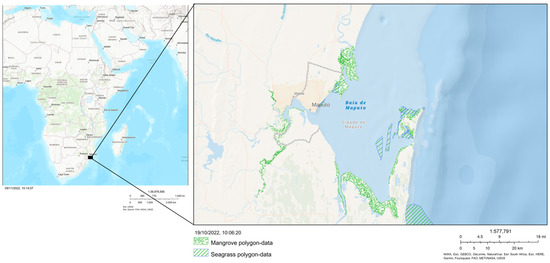
Figure 1.
Location of mangrove forests and seagrass meadows in Maputo Bay, Mozambique (maps created with ArcGis WebBuilder, https://ulisboa.maps.arcgis.com/, accessed on 19 October 2022).
The mangroves cover an area of about 176 km2 around Maputo Bay, and the seagrass meadows cover about 38 km2 [21].
2.2. Data Collection
The data were obtained through the database previously compiled [31] within the scope of the COBIONET project and included genus and species occurrence (defined here as the register of a taxon in a particular place on a specific date) data, the ecosystem of occurrence and species traits. The complete dataset can be assessed on the GBIF platform (https://doi.org/10.15468/w4s7cc (accessed on 10 November 2022)). The occurrence data were compiled from data downloaded from the GBIF and literature. The resulting data were subsequently subjected to taxonomic review and validation and georeferenced using the World Register of Marine Species (WoRMS, http://www.marinespecies.org, accessed on 18 March 2019), GEOLocate Collaborative Georeferencing tool (CoGe, https://coge.geo-locate.org, accessed on 18 March 2019) and Google Maps (https://www.google.com/maps, accessed on 18 March 2019), respectively. In this study, we only considered occurrence data within the coordinate boundaries of Maputo Bay and from the mangrove and seagrass meadow ecosystems, resulting in a derived dataset with 1095 occurrences, which is also accessible through the GBIF platform (https://doi.org/10.15468/dd.2qg6gj (accessed on 21 January 2023)) [32]. The data cover the period from 1950 to 2018. In many occurrences, there was no information regarding the sampling methods or individual count [33]. Therefore, in this study, we used only the species presence/absence and/or number of occurrences over a period of time and/or the ecosystem type, in accordance with [34,35,36]. Two functional traits were considered, i.e., habitat occupation (attributes: benthos, nekton and zooplankton) and trophic guild (attributes: deposit feeder, detritivore, filter feeder, generalist feeder, grazer, parasite, planktivore, predator, scavenger, suspension feeder and symbiotic). For each trait, most attributes were obtained from WoRMS, but some were also obtained from the Biological Traits Information Catalogue (BIOTIC, http://www.marlin.ac.uk/biotic/, accessed on 27 June 2021). Some species had no available information. In those instances, attribute data of similar species within the same genus or family were used. All attributes were categorical (i.e., qualitative, sensu [37]).
2.3. Biodiversity Estimates and Statistical Analyses
Species richness (S) [38], taxonomic diversity (Simpson’s diversity index—D [35,39]) and functional diversity (Rao index [35,40]), calculated per decade, were used as biodiversity estimates. To calculate D, both S and the number of occurrences of each species in each ecosystem were considered [35,41].
The community structure was assessed by estimating the diversity of each functional trait using the Shannon–Wiener diversity index (H’) [42] and the average taxonomic distinctness index (Δ+) [43], as described by Clarke and Warwick, as the path length between two randomly chosen species [44]. The community composition, based on the diversity of each functional trait, was analysed separately for each ecosystem to identify differences between them using multidimensional scaling (MDS). As the data had many zero values, a log(x+1) transformation was used [45], and resemblance matrices were created using the Bray–Curtis similarity index. These matrices were used to produce MDS cluster graphs.
S was calculated in an Excel spreadsheet (Microsoft Corporation, 2018, Microsoft Excel, available at: https://office.microsoft.com/excel (accessed on 16 October 2022)). D and the Rao index of functional diversity were calculated using the ‘FunctDiv.exl’ macro of Lepš et al. [35]. H’, Δ+ and MDS statistical analyses were conducted using PRIMER 6 statistical software [46]. Although the abovementioned diversity indexes preferably use abundance data, they can also be applied to presence/absence and occurrence data [25,26,27,28,29,34,35,36]. Therefore, they were suitable for application in this study.
3. Results
Mangroves presented a higher marine macroinvertebrate species richness than seagrass meadows, except between 2000 and 2019 (Figure 2). From 1950 to 2019, 416 species were identified, including 282 in mangroves and 167 in seagrass meadows (33 species overlapped both ecosystems). Overall, the highest species richness was registered in 2010–2019 (124 species).
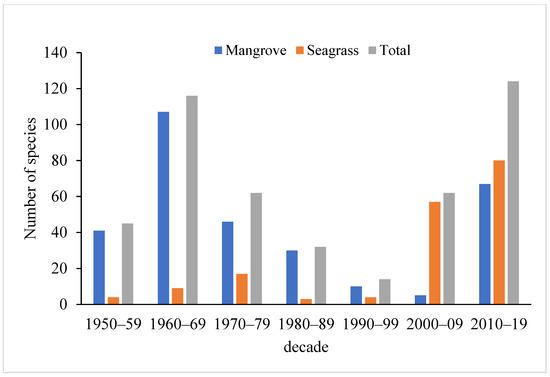
Figure 2.
Marine invertebrate species richness (S) in Maputo Bay(mangrove forests, seagrass meadows and total) per decade.
Mirroring the results of the marine invertebrate species richness, the taxonomic diversity (D) of Maputo Bay’s mangroves was higher than that of the seagrass meadows, except for the period between 2000 and 2019 (Figure 3).
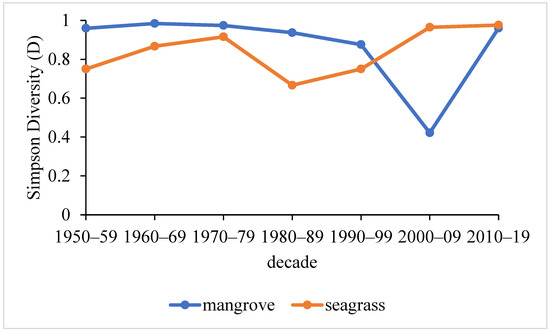
Figure 3.
Marine invertebrate taxonomic diversity (Simpson’s Diversity Index—D) of mangroves and seagrasses in Maputo Bay per decade.
The functional diversity (Rao Index) of mangroves was explained mainly by the trophic guild trait (Figure 4a). In the case of seagrasses, between 1950 and 1970, the functional diversity was mostly due to differences in habitat occupation. This trend has shifted in more recent decades (the 1980s onward), with the functional diversity being explained mainly by the trophic guild (Figure 4b).
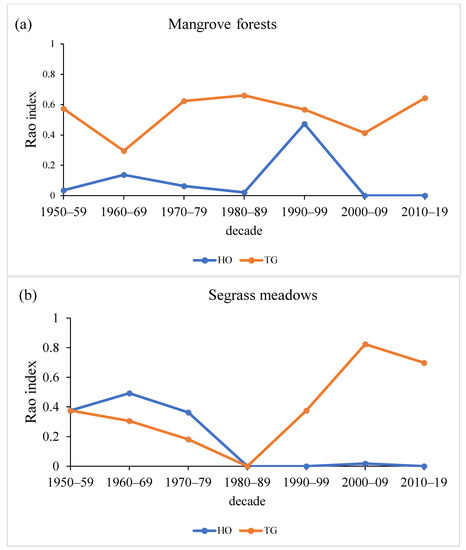
Figure 4.
Marine invertebrate functional diversity (Rao Index) based on habitat occupation (HO) and trophic guild (TG) traits per decade in Maputo Bay. (a) Mangroves; (b) seagrasses.
The assessment of the community composition of the mangroves (Figure 5) in regard to habitat occupation showed that the benthonic attribute had the highest Shannon–Wiener diversity index (H’ = 5.23) and average taxonomic distinctness index (Δ+ = 87.36), followed by the zooplanktonic (H’ = 2.00; Δ+ = 65.56) and nektonic (H’ = 1.28; Δ+ = 47.22) attributes (Figure 5a,c). As for the trophic guild, the “predator” attribute exhibited the highest diversity index (H’ = 4.38), closely followed by the “planktivore” (H’ = 3.94) attribute. The “parasite” and “detrivore” attributes presented with low diversity (H’ = 0.69 and H’ = 0, respectively) (Figure 5b). In this instance, Δ+ did not follow the same trend as H’. The “parasite” attribute had the lowest H’ value but the highest taxonomic distinctness (Δ+ = 83.33), being the same as the “symbiotic” attribute (Figure 5d).
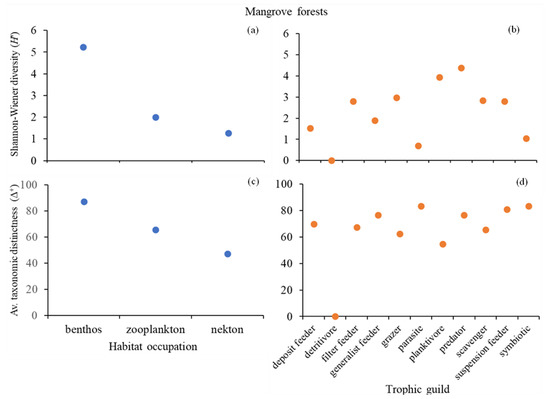
Figure 5.
Community composition of mangrove forests in Maputo Bay. Shannon–Wiener diversity index (H’) regarding (a) habitat occupation (HO) and (b) trophic guild (TG). Average taxonomic distinctness index (Δ+) regarding (c) HO and (d) TG. (Note: all values indicated in the graphs as 0 are due to the low number of occurrences).
Regarding the community composition of the seagrasses (Figure 6) in regard to habitat occupation, the benthonic attribute presented the highest Shannon–Wiener diversity index (H’ = 4.75), followed by the zooplanktonic (H’ = 2.13) and nektonic (H’ = 1.61) attributes (Figure 6a). The benthonic attribute also presented the highest taxonomic distinctness (Δ+ = 80.33), followed by the nektonic (Δ+ = 43.33) and zooplanktonic (Δ+ = 36.97) attributes (Figure 6c). Concerning the trophic guild (Figure 6), the “predator” attribute exhibited the highest H’ (4.11), followed by the “suspension feeder” (H’ = 3.12) attribute. The “parasite” (H’ = 0.69), “detrivore”, “generalist feeder” and “planktivore” attributes (all with H’ = 0) presented the lowest diversity indices (Figure 6b). As in the case of mangroves, the “parasite” attribute of seagrasses had one of the lowest H’ values but the highest average taxonomic distinctness (Δ+ = 100), followed by the “deposit feeder” (Δ+ = 90.74) attribute (Figure 6d).
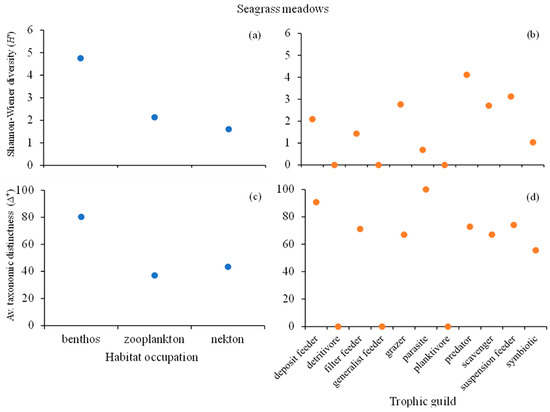
Figure 6.
Community composition of seagrass meadows in Maputo Bay. Shannon–Wiener diversity index (H’) regarding (a) habitat occupation (HO) and (b) trophic guild (TG). Average taxonomic distinctness index (Δ+) regarding (c) HO and (d) TG. (Note: all values indicated in the graphs as 0 are due to the low number of occurrences).
Comparing the community compositions of the mangroves and seagrasses (Figure 7a) in regard to habitat occupation, it can be noted that the benthic and zooplanktonic attributes had a level of similarity of 5%. However, the nektonic attribute was dissimilar between the two ecosystems. Groupings of species belonging to certain trophic attributes (e.g., scavengers, deposit feeders, predators, grazers and suspension feeders) showed a similarity of 5% between the two ecosystems (Figure 7b). The other trophic guild attributes presented dissimilarities between the two ecosystems.
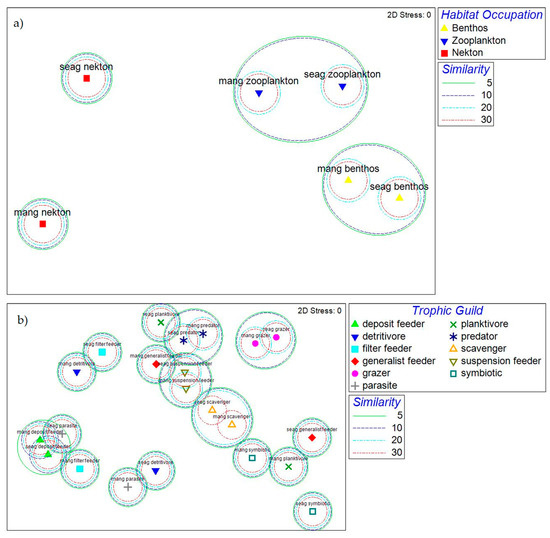
Figure 7.
MDS configuration of marine macroinvertebrate community composition related to (a) habitat occupation and (b) trophic guild in Maputo Bay´s mangroves and seagrasses, with superimposed clustering based on Bray–Curtis similarities (similarity in percentage).
4. Discussion
Marine macroinvertebrates presented higher species richness and taxonomic diversity in mangrove forests than in seagrass meadows until the end of the 20th century. From the 2000s onwards, this trend was reversed. This can be explained by a greater collection effort in the case of seagrass meadows in recent years [47,48,49,50]. Similar to our findings, the authors of [5] discovered that seagrasses displayed greater species richness and taxonomic diversity than mangroves in the Matapoury Estuary (New Zealand) in 2006. On the other hand, mangroves cover a much larger area than seagrasses in Maputo Bay, mainly due to the sediment input from five rivers: Incomati, Maputo, Tembe, Umbeluzi and Matola [20]. This probably led to sampling bias in studies focusing primarily on mangroves in the 20th century [3,4].
The trophic guild trait mostly explained the functional diversity of mangroves, whereas habitat occupation mostly explained the functional diversity of seagrasses. This trend in seagrasses has shifted in recent years, with the functional diversity being explained mostly by the trophic guild. As trophic guild diversity is related to resource availability and food web interactions, it is often considered as a way of describing community functioning [18]. However, the data on a trait must be available for all species to be valid on the community level [51], as was achieved in our study.
Regarding habitat occupation, the community composition, in relation to biodiversity (H’) and taxonomic distinctiveness (Δ+), was identical in mangroves and seagrasses. Thus, benthos presented higher biodiversity and taxonomic distinctiveness in both ecosystems than the other attributes, while in seagrasses, nekton had a lower biodiversity than zooplankton but a higher taxonomic distinctiveness. This indicates that there are fewer nektonic than zooplanktonic species, but the former are more distinct on the taxonomic level. However, it must be noted that nektonic species presented a low n (seven occurrences in mangroves and five in seagrass meadows). As for the trophic guild, the community composition, in related to the highest biodiversity and taxonomic distinctiveness, i.e., the predator and parasite attributes, respectively, was identical in mangroves and seagrasses. Few trophic guild attributes did not display distinctiveness (probably due to a low n), and some were underrepresented or absent in the database, e.g., the “detrivore” attribute in seagrasses (n = 1) and mangroves (n = 0). This may be explained by data gaps in the original digital databases, which can partially be attributed to the limited number of studies, e.g., those conducted [15] in Kenya and [13,14] in the Republic of South Africa, directed towards functional diversity and community composition in Eastern Africa. The application of the MDS to compare the community composition between mangroves and seagrasses revealed a low degree of similarity (5%) or dissimilarity between the two ecosystems in terms of habitat occupation and trophic guild traits. Our results are consistent with the MDS findings from [5], which showed dissimilarity in the community structure and composition between mangroves and seagrasses.
The use of digital databases to compile information regarding different facets of marine macroinvertebrate diversity has some limitations. There was no or little information on the sampling methods and individual count (the lack of this information can invalidate the usefulness of these data for different quantitative studies), in addition to out-of-date taxa IDs [22,52,53], which, nevertheless, can be updated using specialised taxonomic platforms and literature. The decline in the number of occurrences from 1960 to 1999 can be explained in part by the political instability between 1964 and 1992 [54] but also by sampling bias (e.g., the available data being limited to studies of specific ecosystems or target species used as food sources, as exemplified by the data compiled for this study, which show that 66% of the species occurrences were from mangrove forests vs. 34% from seagrass meadows) and data gaps in the original digital databases [22,33,52,53]. The lack of work on mangroves and especially seagrasses can be attributed to the reduced number of marine biology education/research institutions in Mozambique. Hence, the first study on marine biology in Mozambique was carried out by South African researchers on Inhaca island. Mozambique did not have an institutional setting to carry out marine biology research until 1985, when the first marine biology course was implemented. Additionally, the Fisheries Research Institute was only established in 1980. Digital biodiversity repository managers need to state the need to register data as the objective of the study, together with the methods used and number of individuals, as accurately as possible. The increase in the number of accessible digital databases and the associated metadata should also increase the accuracy of functional diversity estimates. More accurate biodiversity data integrated into global digital databases will allow us to gain a better understanding of marine invertebrate communities worldwide. Standardized sampling protocols, such as those provided by the Indo-Pacific Seagrass Network [55], can also help to improve the accuracy of databases.
This study demonstrates the utility of online databases as a proxy for in situ studies, limiting the environmental pressures that fieldwork can sometimes create or problems involving locations that are difficult to access (both logistically and political), as well as their limitations, such as incomplete metadata entries and reliance on the high-quality digitisation of NHC and fieldwork data. Specifically, this study contributes temporal baseline data on species richness, taxonomic and functional diversity, and overall community composition of marine macroinvertebrates in mangroves and seagrasses in Maputo Bay. The trends and data gaps shown in this study can be further used to inform subsequent systematic data acquisition and support the development of future management and conservation plans, especially for endangered, unique and endemic species. A further step for future studies is to incorporate functional traits into online biodiversity and NHC databases.
Author Contributions
Conceptualization, M.B. and A.M.C.; methodology, M.B.; software, M.B.; validation, M.B. and A.M.C.; formal analysis, M.B.; investigation, M.B.; data curation, M.B.; writing—original draft preparation, M.B. and A.M.C.; writing—review and editing, M.B., S.B., J.P. and A.M.C.; visualization, M.B., J.P. and A.M.C.; supervision, S.B., J.P. and A.M.C.; project administration, S.B., J.P. and A.M.C.; funding acquisition, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was part of the project COBIO-NET (Coastal biodiversity and food security in peri-urban sub-Saharan Africa: assessment, capacity building and regional networking in contrasting Indian and Atlantic Oceans), funded by the Aga Khan Development Network (AKDN) and Fundação para a Ciência e a Tecnologia, I.P. (FCT). MB acknowledges a grant funded by FCT (Ref. SFRH/BD/147875/2019).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available and accessible through the GBIF portal under the Creative Commons Attribution 4.0 International license at (https://doi.org/10.15468/dd.2qg6gj (accessed on 21 January 2023)) (Bento, 2023).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lefcheck, J.S.; Marion, S.R.; Lombana, A.V.; Orth, R.J. Faunal Communities Are Invariant to Fragmentation in Experimental Seagrass Landscapes. PLoS ONE 2016, 11, e0156550. [Google Scholar] [CrossRef] [PubMed]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of Mangrove Forests Degradation on Biodiversity and Ecosystem Functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, S.; Gullström, M.; Balidy, H.; Samussone, D.; Cossa, D. Seagrass Meadows in Maputo Bay. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 147–170. ISBN 9789987955930. [Google Scholar]
- Paula, J.; Macamo, C.; Bandeira, S. Mangroves of Maputo Bay. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 109–126. [Google Scholar]
- Alfaro, A.C. Benthic Macro-Invertebrate Community Composition within a Mangrove/Seagrass Estuary in Northern New Zealand. Estuar. Coast. Shelf Sci. 2006, 66, 97–110. [Google Scholar] [CrossRef]
- Phang, V.X.H.; Chou, L.M.; Friess, D.A. Ecosystem Carbon Stocks across a Tropical Intertidal Habitat Mosaic of Mangrove Forest, Seagrass Meadow, Mudflat and Sandbar. Earth Surf. Process. Landforms 2015, 40, 1387–1400. [Google Scholar] [CrossRef]
- Kruitwagen, G.; Nagelkerken, I.; Lugendo, B.R.; Mgaya, Y.D.; Bonga, S.E.W. Importance of Different Carbon Sources for Macroinvertebrates and Fishes of an Interlinked Mangrove-Mudflat Ecosystem (Tanzania). Estuar. Coast. Shelf Sci. 2010, 88, 464–472. [Google Scholar] [CrossRef]
- Corte, G.N.; Checon, H.H.; Shah Esmaeili, Y.; Lefcheck, J.S.; Amaral, A.C.Z. Mangrove Fragments as Key Coastal Reservoirs of Taxonomic and Functional Biodiversity. Biodivers. Conserv. 2021, 30, 1573–1593. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A Global Crisis for Seagrass Ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Bruno, J.F.; Duffy, J.E. Understanding the Effects of Marine Biodiversity on Communities and Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 739–766. [Google Scholar] [CrossRef]
- Coccia, C.; Almeida, B.A.; Green, A.J.; Gutiérrez, A.B.; Carbonell, J.A. Functional Diversity of Macroinvertebrates as a Tool to Evaluate Wetland Restoration. J. Appl. Ecol. 2021, 58, 2999–3011. [Google Scholar] [CrossRef]
- Barnes, R.S.K.; Hendy, I.W. Seagrass-Associated Macrobenthic Functional Diversity and Functional Structure along an Estuarine Gradient. Estuar. Coast. Shelf Sci. 2015, 164, 233–243. [Google Scholar] [CrossRef]
- Barnes, R.S.K. What Does Measuring Species Diversity in Estuarine Seagrass Systems Actually Assess? Mar. Environ. Res. 2021, 172, 105500. [Google Scholar] [CrossRef]
- Barbanera, A.; Markesteijn, L.; Kairo, J.; Juma, G.A.; Karythis, S.; Skov, M.W. Functional Responses of Mangrove Fauna to Forest Degradation. Mar. Freshw. Res. 2022, 73, 762–763. [Google Scholar] [CrossRef]
- Henseler, C.; Nordström, M.C.; Törnroos, A.; Snickars, M.; Pecuchet, L.; Lindegren, M.; Bonsdorff, E. Coastal Habitats and Their Importance for the Diversity of Benthic Communities: A Species- and Trait-Based Approach. Estuar. Coast. Shelf Sci. 2019, 226, 106272. [Google Scholar] [CrossRef]
- Wong, M.C.; Dowd, M. Patterns in Taxonomic and Functional Diversity of Macrobenthic Invertebrates Across Seagrass Habitats: A Case Study in Atlantic Canada. Estuaries Coasts 2015, 38, 2323–2336. [Google Scholar] [CrossRef]
- Paganelli, D.; Marchini, A.; Occhipinti-Ambrogi, A. Functional Structure of Marine Benthic Assemblages Using Bio-logical Traits Analysis (BTA): A Study along the Emilia-Romagna Coastline (Italy, North-West Adriatic Sea). Estuar. Coast. Shelf Sci. 2012, 96, 245–256. [Google Scholar] [CrossRef]
- Paula, J.; Bandeira, S. An Introduction to the Maputo Bay. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 3–10. ISBN 978-9987-9559-3-0. [Google Scholar]
- Machava-António, V.; Fernando, A.; Cravo, M.; Massingue, M.; Lima, H.; Macamo, C.; Bandeira, S.; Paula, J. A Comparison of Mangrove Forest Structure and Ecosystem Services in Maputo Bay (Eastern Africa) and Príncipe Island (Western Africa). Forests 2022, 13, 1466. [Google Scholar] [CrossRef]
- Ferreira, M.A.; Bandeira, S. Maputo Bay’s Coastal Habitats. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 21–30. [Google Scholar]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial Bias in the GBIF Database and Its Effect on Modeling Species’ Geographic Distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Smith, V.S.; Blagoderov, V. Bringing Collections out of the Dark. Zookeys 2012, 209, 1–6. [Google Scholar] [CrossRef]
- Cannicci, S.; Lee, S.Y.; Bravo, H.; Cantera-Kintz, J.R.; Dahdouh-Guebas, F.; Fratini, S.; Fusi, M.; Jimenez, P.J.; Nordhaus, I.; Porri, F.; et al. A Functional Analysis Reveals Extremely Low Redundancy in Global Mangrove Invertebrate Fauna. Proc. Natl. Acad. Sci. USA 2021, 118, e2016913118. [Google Scholar] [CrossRef]
- Bender, I.M.A.; Kissling, W.D.; Böhning-Gaese, K.; Hensen, I.; Kühn, I.; Nowak, L.; Töpfer, T.; Wiegand, T.; Dehling, D.M.; Schleuning, M. Projected Impacts of Climate Change on Functional Diversity of Frugivorous Birds along a Tropical Elevational Gradient. Sci. Rep. 2019, 9, 17708. [Google Scholar] [CrossRef] [PubMed]
- Freitas, C.; Brum, F.T.; Cássia-Silva, C.; Maracahipes, L.; Carlucci, M.B.; Collevatti, R.G.; Bacon, C.D. Incongruent Spatial Distribution of Taxonomic, Phylogenetic, and Functional Diversity in Neotropical Cocosoid Palms. Front. For. Glob. Chang. 2021, 4, 739468. [Google Scholar] [CrossRef]
- Granado-Lorencio, C.; Guisande, C.; Pelayo-Villamil, P.; Manjarrés-Hernández, A.; García-Roselló, E.; Heine, J.; Pérez-Costas, E.; González-Vilas, L.; González-Dacosta, J.; Lobo, J.M. Diversity Dimensions of Freshwater Fish Species around the World. J. Geogr. Inf. Syst. 2021, 13, 106371. [Google Scholar] [CrossRef]
- Lajeunesse, A.; Fourcade, Y. Temporal Analysis of GBIF Data Reveals the Restructuring of Communities Following Climate Change. J. Anim. Ecol. 2022, 1–12. [Google Scholar] [CrossRef]
- Toussaint, A.; Brosse, S.; Bueno, C.G.; Pärtel, M.; Tamme, R.; Carmona, C.P. Extinction of Threatened Vertebrates Will Lead to Idiosyncratic Changes in Functional Diversity across the World. Nat. Commun. 2021, 12, 5162. [Google Scholar] [CrossRef]
- da Silva, A.; Rafael, J. Geographical and Socio-Economic Setting of Maputo Bay. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 11–20. [Google Scholar]
- Bento, M.; Niza, H.; Cartaxana, A.; Bandeira, S.; Paula, J.; Marçal Correia, A. A Dataset of Marine Macroinvertebrate Diversity from Mozambique and São Tomé and Príncipe; Version 1.6; Departamento de Biologia Animal, Faculdade de Ciências, Universidade de Lisboa: Lisboa, Portugal, Occurrence Dataset. [CrossRef]
- Bento, M. A Dataset of Marine Macroinvertebrate Diversity from Maputo Bay, Mozambique. Available online: https://www.gbif.org/derivedDataset/10.15468/dd.2qg6gj (accessed on 22 January 2023).
- Bento, M.; Niza, H.; Cartaxana, A.; Bandeira, S.; Paula, J.; Correia, A.M. Mind the Gaps: Taxonomic, Geographic and Temporal Data of Marine Invertebrate Databases from Mozambique and São Tomé and Príncipe. Diversity 2023, 15, 70. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional Diversity (FD), Species Richness and Community Composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Lepš, J.; de Bello, F.; Lavorel, S.; Berman, S. Quantifying and Interpreting Functional Diversity of Natural Communities: Practical Considerations Matter. Preslia 2006, 78, 481–501. [Google Scholar]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New Multidimensional Functional Diversity Indices for a Multifaceted Framework in Functional Ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional Diversity: Back to Basics and Looking Forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef]
- Moore, J.C. Diversity, Taxonomic versus Functional. Encycl. Biodivers. Second Ed. 2013, 2, 648–656. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Rao, R. Diversity and Dissimilarity. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Pélissier, R.; Couteron, P.; Dray, S.; Sabatier, D. Consistency between Ordination Techniques and Diversity Measurements: Two Strategies for Species Occurrence Data. Ecology 2003, 84, 242–251. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Theory of Mathematical Communication, 10th ed.; The University of Illinois Press: Chicago, IL, USA, 1964; pp. 1–131. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. A Further Biodiversity Index Applicable to Species Lists: Variation in Taxonomic Distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A Taxonomic Distinctness Index and Its Statistical Properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Paula, J.; Bandeira, S.; Msangameno, D.; Queiroga, H. Measures of Biological Diversity. In Exercises in Marine Biodiversity and Ecology—A Training Manual Using the Intertidal Habitats of the WIO Region; WIOMSA: Zanzibar Town, Tanzania, 2021; pp. 59–78. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual PRIMER-E; Primer-E Ldt: Plymouth, UK, 2006; pp. 1–91. [Google Scholar]
- Chitará-Nhandimo, S.; Chissico, A.; Mubai, M.E.; de Cabral, A.S.; Guissamulo, A.; Bandeira, S. Seagrass Invertebrate Fisheries, Their Value Chains and the Role of LMMAs in Sustainability of the Coastal Communities—Case of Southern Mozambique. Diversity 2022, 14, 170. [Google Scholar] [CrossRef]
- Tavares, A.I.; Assis, J.; Patrício, A.R.; Ferreira, R.; Cheikh, M.A.S.; Bandeira, S.; Regalla, A.; Santos, I.; Potouroglou, M.; Nicolau, S.; et al. Seagrass Connectivity on the West Coast of Africa Supports the Hypothesis of Grazer-Mediated Seed Dispersal. Front. Mar. Sci. 2022, 9, 852. [Google Scholar] [CrossRef]
- Cheikh, M.A.S.; Bandeira, S.O.; Traganos, D.; Poursanidis, D.; Vegh, T. Seagrasses of West Africa: New Discoveries, Distribution Limits, Ecosystem Services and Prospects for Management. Diversity 2022, 15, 5. [Google Scholar] [CrossRef]
- Amone-Mabuto, M.; Hollander, J.; Lugendo, B.; Adams, J.B.; Bandeira, S. A Field Experiment Exploring Disturbance-and-Recovery, and Restoration Methodology of Zostera Capensis to Support Its Role as a Coastal Protector. Nord. J. Bot. 2023, 2023, e03632. [Google Scholar] [CrossRef]
- de Oliveira, S.E.; Bozelli, R.L. How Planktonic Microcrustaceans Respond to Environment and Affect Ecosystem: A Functional Trait Perspective. Int. Aquat. Res. 2019, 11, 207–223. [Google Scholar] [CrossRef]
- Faith, D.; Collen, B.; Ariño, A.; Patricia Koleff, P.K.; Guinotte, J.; Kerr, J.; Chavan, V. Bridging the Biodiversity Data Gaps: Recommendations to Meet Users’ Data Needs. Biodivers. Inform. 2013, 8, 41–58. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating Species Diversity and Distribution in the Era of Big Data: To What Extent Can We Trust Public Databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Neves, I.Q.; da Mathias, M.L.; Bastos-Silveira, C. Mapping Knowledge Gaps of Mozambique’s Terrestrial Mammals. Sci. Rep. 2019, 9, 18184. [Google Scholar] [CrossRef]
- Indo-Pacific Seagrass Network (IPSN). Available online: https://indopacificseagrassnetwork.wordpress.com/ (accessed on 26 December 2022).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).