
First Report of a Paucibranchia (Polychaeta, Eunicidae) Species without Lateral Palps in Korean Subtidal Waters, with Genetic Evidence for Its Taxonomic Position †


Abstract
1. Introduction
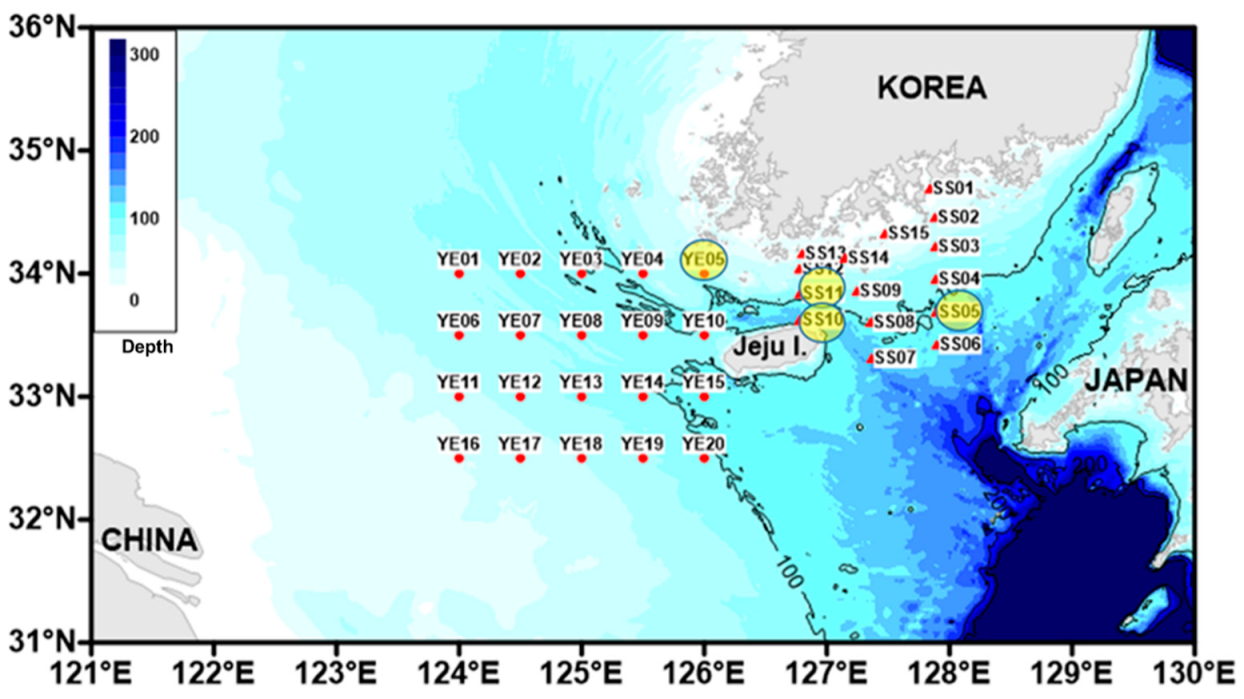
2. Materials and Methods
3. Results
3.1. Systematics
3.2. Generic Diagnosis
3.3. Material Examined
3.4. Species Diagnosis
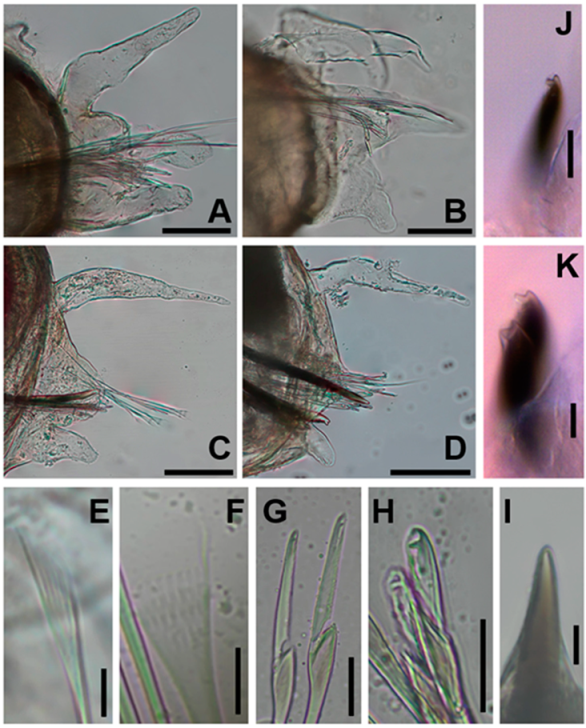
3.5. Description
3.6. Morphological Variation
3.7. Etymology
3.8. Distribution and Ecology
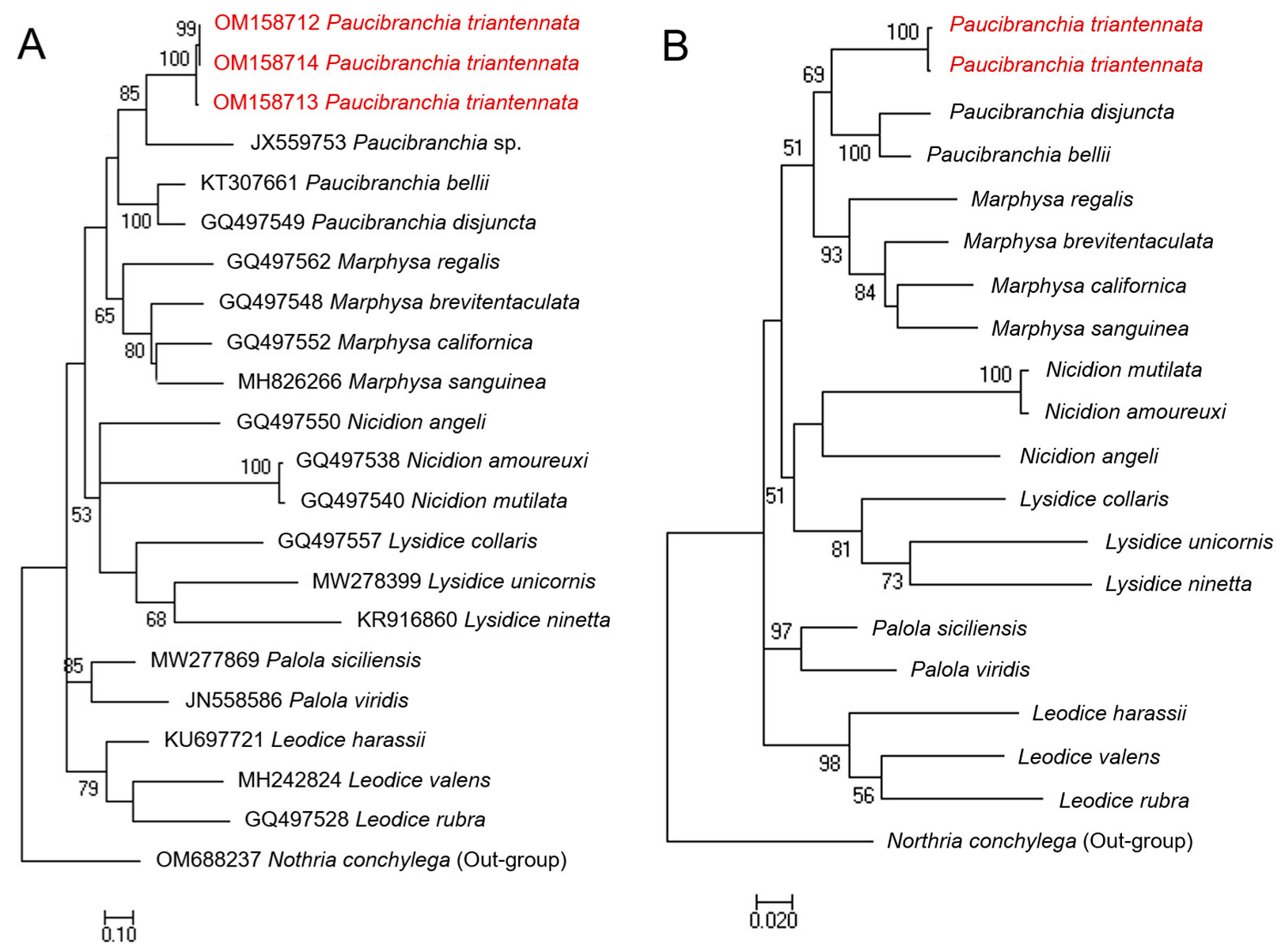
3.9. Molecular Comparison
3.10. Remarks
4. Discussion
- 1.
- Only compound spinigers...............................................................................2
- -
- Only compound falcigers.................................................................................5
- -
- Both compound spinigers and falcigers............................................................12
- 2(1).
- Postchaetal lobes bluntly conical or (only) conical in pre-branchial chaetigers, compound spinigers with blades of different sizes in the same chaetiger........................3
- -
- Postchaetal lobes tongue-shaped in pre-branchial chaetigers, compound spinigers with all blades of similar size in the same chaetiger...................P. kinbergi (McIntosh, 1910)
- 3(2).
- Compound spinigers with blades of two sizes, pectinate chaetae with oblique distal edge in anterior chaetigers..............................................................................4
- -
- Compound spinigers with blades of three sizes, pectinate chaetae with transverse distal edge in anterior chaetigers...................................... P. gilberti Molina-Acevedo, 2018
- 4(3).
- Eyes present, chaetal lobes rectangular in pre-branchial region, dorsal cirri in postbranchial region almost three times longer than anterior ones............................................................................................................. P. disjuncta (Hartman, 1961)
- -
- Eyes absent, chaetal lobes rounded in pre-branchial region, dorsal cirri in post-branchial region two times longer than anterior ones......................... P. cinari (Kurt-Sahin, 2014)
- 5(1).
- Subacicular hooks with reddish basal end, distally amber or translucent................6
- -
- Subacicular hooks translucent...........................................................................9
- 6(5).
- Postchaetal lobes digitiform in pre-branchial chaetigers, eyes absent, falcigers with all blades of similar size in anterior chaetigers............ P. miroi Molina-Acevedo, 2018
- -
- Postchaetal lobes bluntly conical in pre-branchial chaetigers, eyes present, falcigers with blades of two or three sizes in anterior chaetigers..................................................7
- 7(6).
- Dorsal cirri in post-branchial region 1.4 times longer than pre-branchial region, falcigers with blades of three sizes in anterior region, pectinate with transverse distal edge in anterior chaetigers......................................................P. purcellana (Willey, 1904)
- -
- Dorsal cirri in pre- and post-branchial chaetigers of similar size, falcigers with blades of two sizes in anterior chaetigers, pectinate with oblique distal edge in anterior chaetigers............................................................................................................ 8
- 8(7).
- Falcigers with blades of two sizes in posterior chaetigers, one subacicular hook per chaetiger, prostomium with lateral palps....................... P. gathofi Molina-Acevedo, 2018
- -
- Falcigers with blade of one size in posterior chaetigers, one or two subacicular hooks per chaetiger, prostomium without lateral palps...........................P. triantennata sp. nov.
- 9(5).
- Aciculae with reddish basal end, distally translucent........... P. conferta (Moore, 1911)
- -
- Aciculae translucent.................................................................................... 10
- 10(9).
- Eyes absent, dorsal cirri in postbranchial region two times longer than prebranchial region, ventral cirri with swollen base start from chaetiger 14........................................................................................................... P. gemmata (Mohammad, 1973)
- -
- Eyes present, dorsal cirri in pre- and postbranchial regions of similar size, ventral cirri with a swollen base from between chaetigers 1 and 7.............................................. 11
- 11(10).
- Postchaetal lobes basally oval, digitiform distal end, tilted dorsally; falcigers with blades of similar sizes in anterior region; pectinate with oblique distal edge in anterior chaetiger........................................................ P. patriciae Molina-Acevedo, 2018
- -
- Postchaetal lobes bluntly conical, digitiform distal end or bluntly conical; falcigers with blades of two sizes in anterior region; pectinate with transverse distal edge in anterior chaetigers; branchiae from chaetigers 14–17 with up to 10–11 filaments...........................................................................................................P. adenensis (Gravier, 1900)
- 12(1).
- Compound spinigers present in all chaetigers...P. totospinata (Lu and Fauchald, 1998)
- -
- Compound spinigers present only in anterior or median chaetigers; compound falcigers present in all the chaetigers............................................................................ 13
- 13(12).
- Subacicular hooks with reddish basal end, distally amber............................................................................................................P. andresi Molina-Acevedo, 2018
- -
- Subacicular hooks translucent or entirely amber.................................................. 14
- 14(13).
- More than four subacicular hooks per chaetiger........................................... 15
- -
- One or two subacicular hooks per chaetiger....................................................... 16
- 15(14).
- Postchaetal lobes digitiform in pre-branchial chaetigers, with 5–6 subacicular hooks per chaetiger in median region............................... P. stragulum (Grube, 1878)
- -
- Postchaetal lobes conical in pre-branchial chaetigers, with more than 10 subacicular hooks per chaetiger in median region..................... P. carrerai Molina-Acevedo, 2018
- 16(14).
- Compound falcigers present from chaetigers 14–22.... P. oculata (Treadwell, 1921)
- -
- Compound falciger present from first chaetiger..................................................17
- 17(16).
- Compound falcigers with blades of two sizes in anterior chaetigers................ 18
- -
- Compound falcigers with blades of similar size in anterior chaetigers; dorsal cirri in pre- and postbranchial region of similar length; postchaetal lobes bluntly conical in branchial region........................................................................P. fallax (Marion and Bobretzky, 1875)
- 18(17).
- Aciculae dark in anterior median region, postchaetal lobes tongue-shaped in pre-branchial chaetigers, pectinate chaetae with oblique distal edge in anterior chaetigers.................................................. P. bellii (Audouin and Milne-Edwards, 1833)
- -
- Aciculae amber in anterior median region, postchaetal lobes basally and with digitiform end in pre-branchial chaetigers, pectinate chaetae with transverse distal edge in anterior chaetigers.......................................................... P. sinensis (Monro, 1934)
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latreille, P.A.; Berthold, A.A. Latreille’s Naturliche Familien des Thierreichs; Mit Anmerkungen und Zusätzen; Landes-Industrie-Comptoirs: Weimar, Germany, 1827; p. 606. [Google Scholar] [CrossRef]
- Read, G.; Fauchald, K. (Eds.) World Polychaeta Database. 2021. Eunicidae Berthold, World Register of Marine Species. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=966 (accessed on 30 December 2021).
- Rouse, G.; Pleijel, F. Polychaetes; Oxford University Press: New York, NY, USA, 2001; p. 354. [Google Scholar]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of Worms Emended: An Update of Polychaete Feeding Guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [PubMed]
- Cole, V.J.; Chick, R.C.; Hutchings, P.A. A review of global fisheries for polychaete worms as a resource for recreational fishers: Diversity, sustainability and research needs. Rev. Fish Biol. Fish. 2018, 28, 543–565. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, B.K.; Kim, S.K.; Phoo, W.W.; Maran, B.A.V.; Kim, C.-H. Appropriate feeding for early juvenile stages of eunicid polychaete Marphysa sanguinea. Fish. Aquat. Sci. 2017, 20, 1–9. [Google Scholar] [CrossRef]
- Watson, G.J.; Murray, J.M.; Schaefer, M.; Bonner, A. Bait worms: A valuable and important fishery with implications for fisheries and conservation management. Fish Fish. 2016, 18, 374–388. [Google Scholar] [CrossRef]
- Pombo, A.; Baptista, T.; Granada, L.; Ferreira, S.; Gonçalves, S.; Anjos, C.; Sá, E.; Chainho, P.; Da Fonseca, L.C.; E Costa, P.F.; et al. Insight into aquaculture’s potential of marine annelid worms and ecological concerns: A review. Rev. Aquac. 2018, 12, 107–121. [Google Scholar] [CrossRef]
- Vijayan, K.; Raj, V.S.; Balasubramanian, C.; Alavandi, S.; Sekhar, V.T.; Santiago, T. Polychaete worms—A vector for white spot syndrome virus (WSSV). Dis. Aquat. Org. 2005, 63, 107–111. [Google Scholar] [CrossRef]
- Zanol, J.; Carrera-Parra, L.; Steiner, T.; Amaral, A.; Wiklund, H.; Ravara, A.; Budaeva, N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 2021, 13, 74. [Google Scholar] [CrossRef]
- De Quatrefages, A. Histoire Naturelle des Annelés Marins et D’eau Douce. Annélides et Géphyriens; Volume Librarie Encyclopédique de Roret: Paris, France, 1866; p. 588. Available online: http://www.biodiversitylibrary.org/page/52110858 (accessed on 10 November 2022).
- Cuvier, G.; Dall, W.H.; Goode, G.B.; Latreille, P.A.; Laurillard, C.L.; Louvet, G.P.; Pierron, J.A.; Schaus, W.; Wood, W.W.; Congress, L.O. Le Règne Animal Distribué D’après son Organisation, Tome 2 Contenant les Reptiles, les Poissons, les Mollusques, les Annelids; Deterville: Paris, France, 1817; p. 532. [Google Scholar] [CrossRef]
- Zanol, J.; Halanych, K.; Struck, T.H.; Fauchald, K. Phylogeny of the bristle worm family Eunicidae (Eunicida, Annelida) and the phylogenetic utility of noncongruent 16S, COI and 18S in combined analyses. Mol. Phylogenetics Evol. 2010, 55, 660–676. [Google Scholar] [CrossRef]
- Zanol, J.; Halanych, K.M.; Fauchald, K. Reconciling taxonomy and phylogeny in the bristleworm family Eunicidae (polychaete, Annelida). Zool. Scr. 2013, 43, 79–100. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C.; Carrera-Parra, L.F. Revision of Marphysa de Quatrefages, 1865 and some species of Nicidion Kinberg, 1865 with the erection of a new genus (Polychaeta: Eunicidae) from the Grand Caribbean. Zootaxa 2017, 4241, 1–62. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C. Morphological revision of the Subgroup 1 Fauchald, 1970 of Marphysa de Quatrefages, 1865 (Eunicidae: Polychaeta). Zootaxa 2018, 4480, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Montagu, G.I. Descriptions of several new or rare Animals, principally marine, discovered on the South Coast of Devonshire. Trans. Linn. Soc. Lond. 1813, 11, 1–26. [Google Scholar] [CrossRef]
- Hutchings, P.; Karageorgopoulos, P. Designation of a neotype of Marphysa sanguinea (Montagu, 1813) and a description of a new species of Marphysa from eastern Australia. Hydrobiologia 2003, 496, 87–94. [Google Scholar] [CrossRef]
- Lavesque, N.; Daffe, G.; Bonifácio, P.; Hutchings, P. A new species of the Marphysa sanguinea complex from French waters (Bay of Biscay, NE Atlantic) (Annelida, Eunicidae). Zookeys 2017, 716, 1–17. [Google Scholar] [CrossRef]
- Lavesque, N.; Daffe, G.; Grall, J.; Zanol, J.; Gouillieux, B.; Hutchings, P. Guess who? On the importance of using appropriate name: Case study of Marphysa sanguinea (Montagu, 1813). ZooKeys 2019, 859, 1–15. [Google Scholar] [CrossRef]
- Paik, E.I. Illustrated Encyclopedia of Fauna and Flora of Korea; Volume 31, Polychaeta; Ministry of Education: Seoul, Republic of Korea, 1989; p. 764.
- Kim, H.; Kim, K.Y.; Phoo, W.W.; Kim, C.H. The first record of the Marphysa victori (Polychaeta, Eunicida, Eunicidae) from Korea, with DNA Barcode Data. Anim. Syst. Evol. Divers. 2021, 37, 1–8. [Google Scholar] [CrossRef]
- Choi, H.K.; Kim, J.G.; Kang, D.W.; Yoon, S.M. A new species of Leodice from Korean waters (Annelida, Polychaeta, Eunicidae). ZooKeys 2017, 715, 53–67. [Google Scholar] [CrossRef][Green Version]
- Aylagas, E.; Borja, Á.; Irigoien, X.; Rodríguez-Ezpeleta, N. Benchmarking DNA metabarcoding for biodiversity-based mon-itoring and assessment. Front. Mar. Sci. 2016, 3, 96. [Google Scholar] [CrossRef]
- Struck, T.H.; Westheide, W.; Purschke, G. Progenesis in Eunicida (“Polychaeta,” Annelida)—Separate evolutionary events? Evidence from molecular data. Mol. Phylogenetics Evol. 2002, 25, 190–199. [Google Scholar] [CrossRef]
- Schulze, A.; Timm, L. Palolo and un: Distinct clades in the genus Palola (Eunicidae, Polychaeta). Mar. Biodivers. 2011, 42, 161–171. [Google Scholar] [CrossRef]
- Miralles, L.; Ardura, A.; Arias, A.; Borrell, Y.; Clusa, L.; Dopico, E.; de Rojas, A.H.; Lopez, B.; Muñoz-Colmenero, M.; Roca, A.; et al. Barcodes of marine invertebrates from north Iberian ports: Native diversity and resistance to biological invasions. Mar. Pollut. Bull. 2016, 112, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Purschke, G.; Halanych, K.M. A scaleless scale worm: Molecular evidence for the phylogenetic placement of Pisione remota (Pisionidae, Annelida) Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark. Mar. Biol. Res. 2005, 1, 243–253. [Google Scholar] [CrossRef]
- Lobo, J.; Teixeira, M.A.L.; Borges, L.M.S.; Ferreira, M.S.G.; Hollatz, C.; Gomes, P.T.; Sousa, R.; Ravara, A.; Costa, M.H.; Costa, F.O. Starting a DNA barcode reference library for shallow water polychaetes from the southern European Atlantic coast. Mol. Ecol. Resour. 2015, 16, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A Tri-Oceanic Perspective: DNA Barcoding Reveals Geographic Structure and Cryptic Diversity in Canadian Polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Audouin, J.V.; Milne Edwards, H. Classification des Annélides et description de celles qui habitent les côtes de la France. Ann. Sci. Nat. 1833, 30, 411–425. [Google Scholar] [CrossRef]
- Jeong, M.-K.; Wi, J.H.; Suh, H.-L. A reassessment of Capitella species (Polychaeta: Capitellidae) from Korean coastal waters, with morphological and molecular evidence. Mar. Biodivers. 2017, 48, 1969–1978. [Google Scholar] [CrossRef]
- Jeong, M.-K.; Soh, H.Y.; Suh, H.-L. Three new species of Heteromastus (Annelida, Capitellidae) from Korean waters, with genetic evidence based on two gene markers. ZooKeys 2019, 869, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Stair, J.B. An account of Palolo, a sea-worm eaten in the Navigator Islands, with a description by JE Gray. In Proceedings of the Zoological Society of London; Zoological Society of London: London, UK, 1847; Volume 15, pp. 17–18. [Google Scholar]
- Lamarck, J.B. Histoire Naturelle des Animaux Sans Vertèbres, Présentant les Caracteres Généraux et Particuliers de ces Animaux, Leur Distribution, Leurs Classes, Leurs Familles, Leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Précédées d´une Introduction Offrant la Détermination des Caractères Essentiels de l´Animal, sa Distinction du Végétal et des Autres Corps Naturels, en fin, l´Exposition des Principes Fondamentaux de la Zoologie; Deterville & Verdiere: Paris, France, 1815; Volume 5, p. 612. [Google Scholar] [CrossRef]
- Moore, J.P. The polychætous annelids dredged by the USS “Albatross” off the coast of Southern California in 1904: III. Euphrosynidæ to Goniadidæ. Proc. Acad. Nat. Sci. Phila. 1911, 63, 234–318. Available online: http://www.biodiversityli-brary.org/bibliography/6885 (accessed on 10 November 2022).
- Kinberg, J.G.H. Annulata Nova. Öfversigt af Königlich Vetenskaps-Akademiens Förhandlingar. Stockholm 1865, 21, 559–574. Available online: http://www.biodiversitylibrary.org/bibliography/15534 (accessed on 9 November 2022).
- Herpin, R. Recherches biologiques sur la reproduction et le développement de quelques Annélides polychètes. Bull. Société Sci. Nat. L’ouest Fr. 1925, 5, 1–250. [Google Scholar]
- Prevedelli, D.; Massamba N’Siala, G.; Ansaloni, I.; Simonini, R. Life cycle of Marphysa sanguinea (Polychaeta: Eunicidae) in the Venice lagoon (Italy). Mar. Ecol. 2007, 28, 384–393. [Google Scholar] [CrossRef]
- Grube, A.E. Actinien, Echinodermen und Wurmen des Adriatischen und Mittelmeers nach Eigenen Sammlungen Beschrieben; J. H. Bon: Königsberg, Russia, 1840; p. 92. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | GenBank Accession Number | Reference | |
|---|---|---|---|
| mtCOI (Locality) | 18S rRNA (Locality) | ||
| Paucibranchia triantennata sp. nov. (MABIKNA00157767) | OM158712 (Republic of Korea) | OM230034 (Republic of Korea) | This study |
| Paucibranchia triantennata sp. nov. (MABIKNA00157768) | OM158713 (Republic of Korea) | OM230035 (Republic of Korea) | This study |
| Paucibranchia triantennata sp. nov. (MABIKNA00157769) | OM158714 (Republic of Korea) | - | This study |
| Paucibranchia sp. | JX559753(Philippines) | - | [13] |
| Paucibranchia fallax | - | GQ497505 (Italy) | [13] |
| Paucibranchia bellii | KT307661 | AF412789 (France) | [24,25] |
| Paucibranchia disjuncta | GQ497549 (Spain) | GQ497504 (USA) | [13] |
| Marphysa californica | GQ497552 (USA) | GQ497507 (USA) | [13] |
| Marphysa brevitentaculata | GQ497548 (Mexico) | GQ497503 (Mexico) | [13] |
| Marphysa sanguinea | MH826266 (USA) | GQ497502 (France) | [13] |
| Marphysa regalis | GQ497562 (Brazil) | GQ497510 (Brazil) | [13] |
| Palola siciliensis | MW277869 (USA) | GQ497514 (Spain) | [13] |
| Palola viridis | JN558586 (Vanuatu) | GQ497513 (Micronesia) | [13,26] |
| Nicidion angeli | GQ497550 (Belize) | GQ497506 (Belize) | [13] |
| Nicidion amoureuxi | GQ497538 (Belize) | GQ497490 (Belize) | [13] |
| Nicidion mutilata | GQ497540 (Belize) | GQ497492 (Belize) | [13] |
| Leodice harassii | KU697721 (Spain) | AY525620 | [27,28] |
| Leodice valens | MH242824 (USA) | GQ497485 (USA) | [13] |
| Leodice rubra | GQ497528 (Brazil) | GQ497478 (Brazil) | [13]) |
| Lysidice collaris | GQ497557 (Spain) | GQ497516 (Spain) | [13] |
| Lysidice unicornis | MW278399 (USA) | AF412792 (Malaysia) | [25] |
| Lysidice ninetta | KR916860 (Portugal) | AF412793 (France) | [25,29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.H.; Soh, H.Y.; Jeong, M.-K. First Report of a Paucibranchia (Polychaeta, Eunicidae) Species without Lateral Palps in Korean Subtidal Waters, with Genetic Evidence for Its Taxonomic Position. Diversity 2022, 14, 1131. https://doi.org/10.3390/d14121131
Kim DH, Soh HY, Jeong M-K. First Report of a Paucibranchia (Polychaeta, Eunicidae) Species without Lateral Palps in Korean Subtidal Waters, with Genetic Evidence for Its Taxonomic Position. Diversity. 2022; 14(12):1131. https://doi.org/10.3390/d14121131
Chicago/Turabian StyleKim, Dae Hun, Ho Young Soh, and Man-Ki Jeong. 2022. "First Report of a Paucibranchia (Polychaeta, Eunicidae) Species without Lateral Palps in Korean Subtidal Waters, with Genetic Evidence for Its Taxonomic Position" Diversity 14, no. 12: 1131. https://doi.org/10.3390/d14121131
APA StyleKim, D. H., Soh, H. Y., & Jeong, M.-K. (2022). First Report of a Paucibranchia (Polychaeta, Eunicidae) Species without Lateral Palps in Korean Subtidal Waters, with Genetic Evidence for Its Taxonomic Position. Diversity, 14(12), 1131. https://doi.org/10.3390/d14121131