
Comparison of Gut Microbiota between Gentoo and Adélie Penguins Breeding Sympatrically on Antarctic Ardley Island as Revealed by Fecal DNA Sequencing



Abstract
1. Introduction
2. Materials and Methods
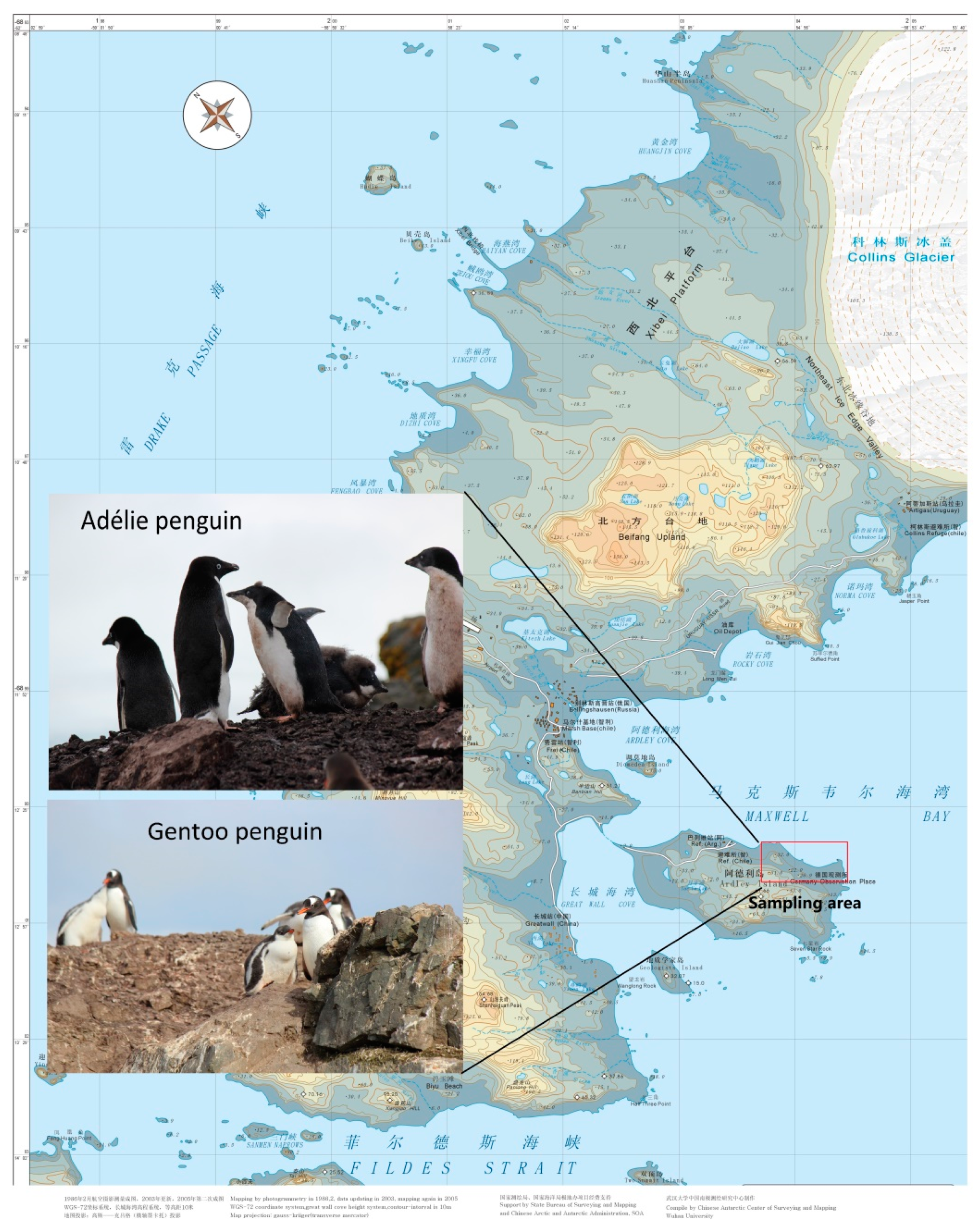
2.1. Sample Collection
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Bioinformatic Analysis
3. Results
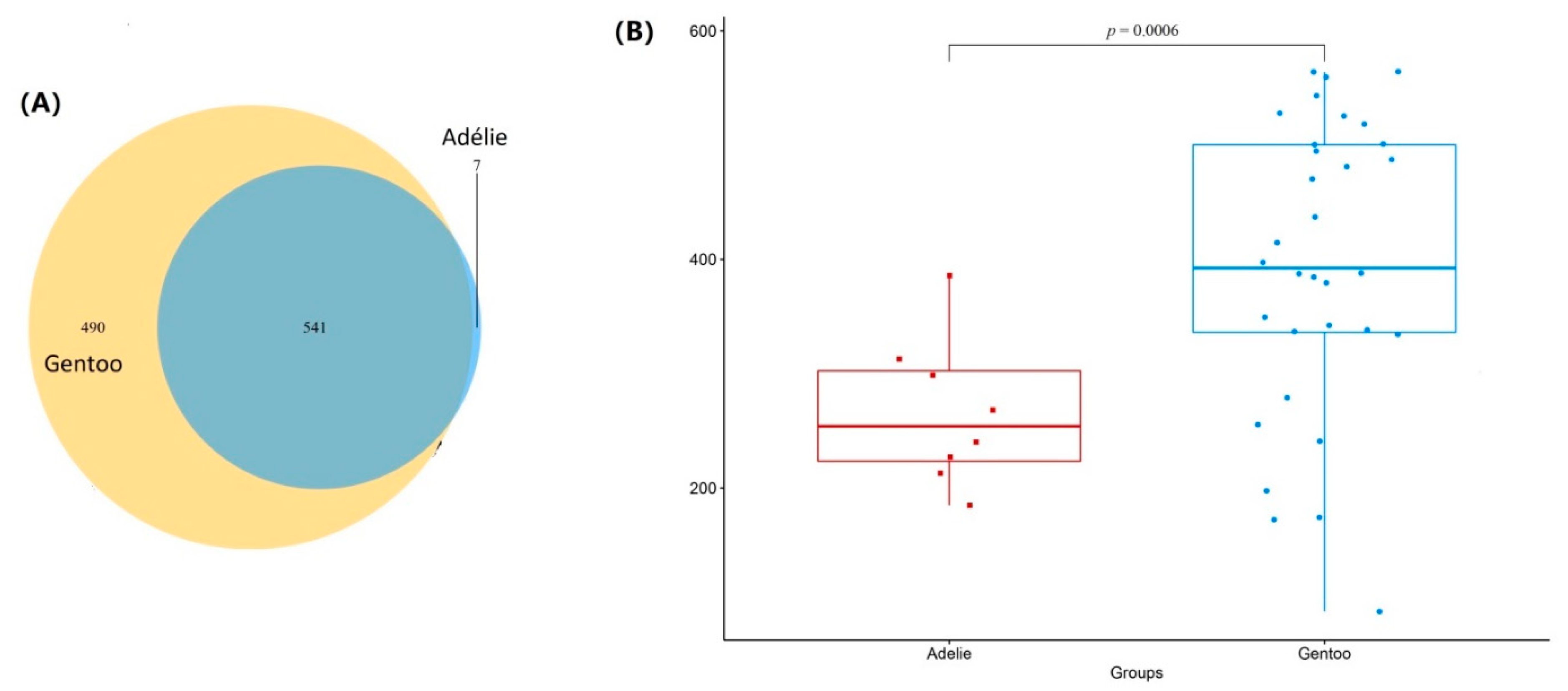
3.1. Sequence Analysis
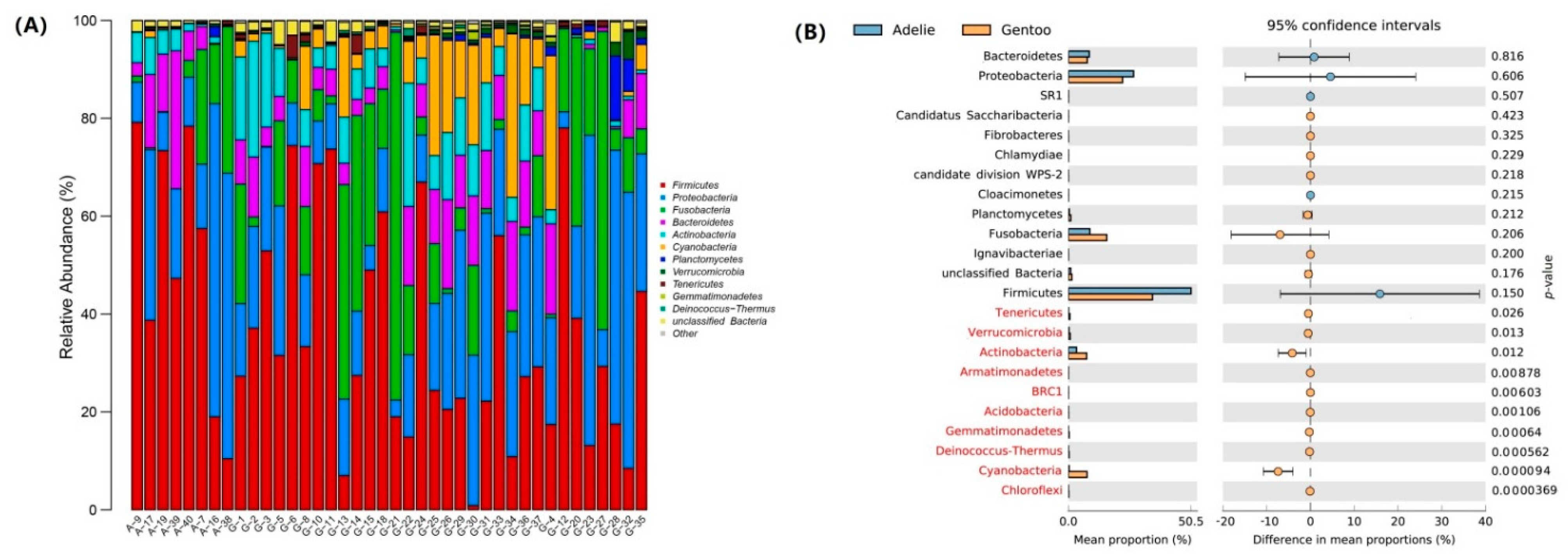
3.2. Bacterial Community Composition in Fecal Micobiota
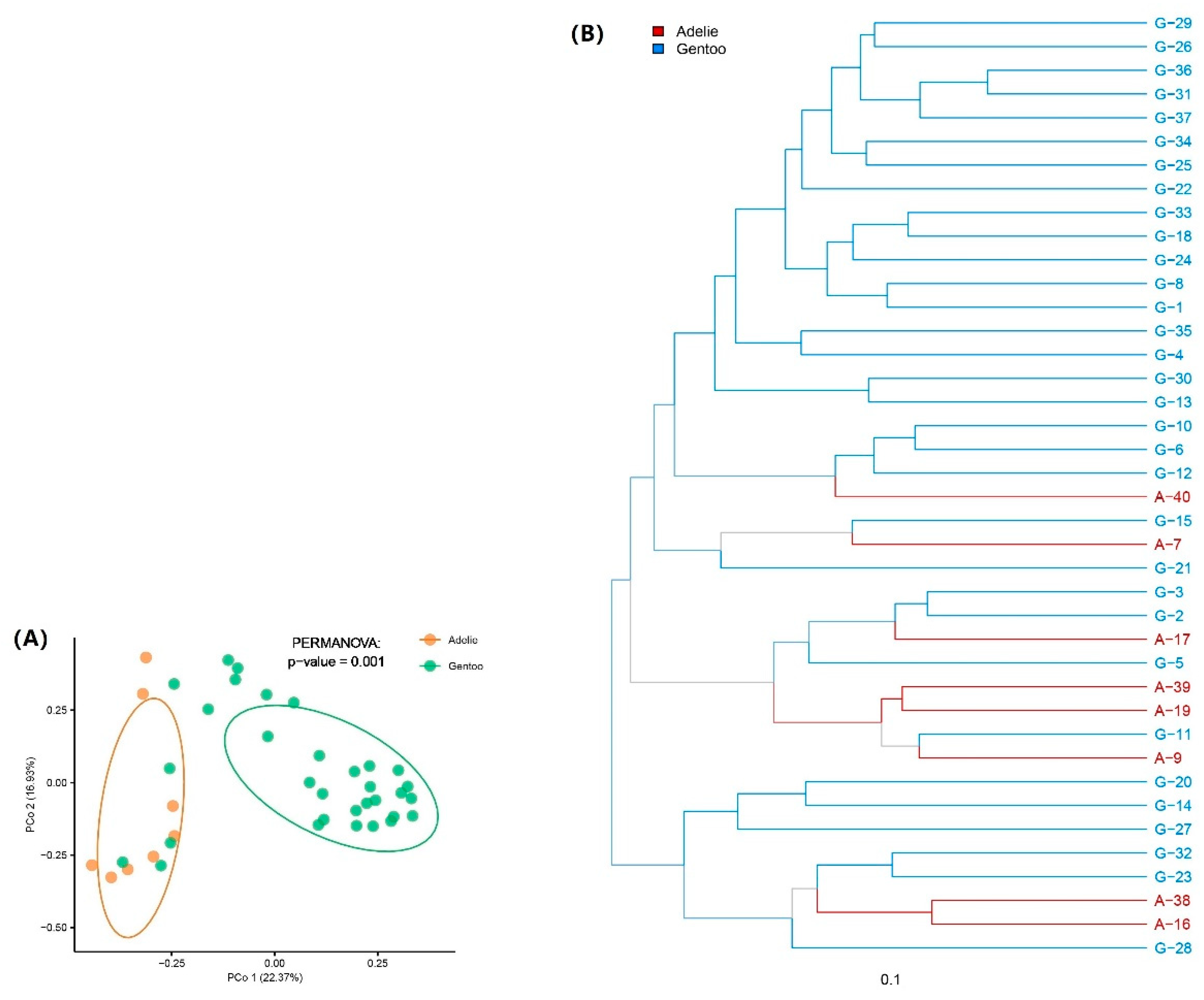
3.3. Differences in Gut Microbiota between Adélie and Gentoo Penguin Groups
4. Discussion
4.1. Firmicutes and Fusobacteria
4.2. Proteobacteria
4.3. Bacteroidetes and Actinobacteria
4.4. Cyanobacteria and Planctomycetes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lynch, H.J.; Naveen, R.; Trathan, P.N.; Fagan, W.F. Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 2012, 93, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Convey, P. Antarctic ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: San Diego, CA, USA, 2013; pp. 179–188. [Google Scholar]
- Peter, H.U.; Braun, C.; Janowski, S.; Nordt, A.; Nordt, A.; Stelter, M. The Current Environmental Situation and Proposals for the Management of the Fildes Peninsula Region. Available online: http://www.uba.de/uba-info-medien-e/4424.html (accessed on 15 February 2013).
- Dewar, M.L.; Arnould, J.P.Y.; Krause, L.; Trathan, P.; Dann, P.; Smith, S.C. Influence of fasting during moult on the faecal microbiota of penguins. PLoS ONE 2014, 9, e99996. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Taylor, M.W. Exploring the avian gut microbiota: Current trends and future directions. Front. Microbiol. 2015, 6, 673. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.; Balagué, V.; Valera, F.; Martínez, A.; Benzal, J.; Motas, M.; Diaz, J.I.; Mira, A.; Pedrós-Alió, C. Age-related differences in the gastrointestinal microbiota of chinstrap penguins (Pygoscelis antarctica). PLoS ONE 2016, 11, e0153215. [Google Scholar] [CrossRef]
- Lee, W.Y.; Cho, H.; Kim, M.; Tripathi, B.M.; Jung, J.W.; Chung, H.; Kim, J.H. Faecal microbiota changes associated with the moult fast in chinstrap and gentoo penguins. PLoS ONE 2019, 14, e0216565. [Google Scholar] [CrossRef]
- Zielińska, S.; Kidawa, D.; Stempniewicz, L.; Łoś, M.; Łoś, J.M. Environmental DNA as a valuable and unique source of information about ecological networks in Arctic terrestrial ecosystems. Environ. Rev. 2017, 25, 282–291. [Google Scholar] [CrossRef]
- Banks, J.C.; Cary, S.C.; Hogg, I.D. The phylogeography of Adelie penguin faecal flora. Environ. Microbiol. 2009, 11, 577–588. [Google Scholar] [CrossRef]
- Dewar, M.L.; Arnould, J.P.; Dann, P.; Trathan, P.; Groscolas, R.; Smith, S. Interspecific variations in the gastrointestinal microbiota in penguins. MicrobiologyOpen 2013, 2, 195–204. [Google Scholar] [CrossRef]
- Grzesiak, J.; Kaczyńska, A.; Gawor, J.; Żuchniewicz, K.; Aleksandrzak-Piekarczyk, T.; Gromadka, R.; Zdanowski, M.K. A smelly business: Microbiology of Adélie penguin guano (Point Thomas rookery, Antarctica). Sci. Total Environ. 2020, 714, 136714. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2015, 18, 1403–1414. [Google Scholar] [CrossRef]
- Du, L.; Chen, Q.; Liu, P.; Zhang, X.; Wang, H.; Zhou, Q.; Xu, D.; Wu, Z. Phosphorus removal performance and biological dephosphorization process in treating reclaimed water by Integrated Vertical-flow Constructed Wetlands (IVCWs). Bioresour. Technol. 2017, 243, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef]
- Tremaroli, V.; Kovatcheva-Datchary, P.; Bäckhed, F. A role for the gut microbiota in energy harvesting? Gut 2010, 59, 1589–1590. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, J.; White, R.L.; Bearne, S.L. Proteomic investigation of amino acid catabolism in the indigenous gut anaerobe Fusobacterium varium. Proteomics 2008, 8, 2691–2703. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]
- Panda, A.K.; Rama Rao, S.V.; Raju, M.V.L.N.; Sunder, G.S. Effect of butyric acid in performance, gastrointestinal tract health and carcass characteristics in broiler chickens. Asian-Australas. J. Anim. Sci. 2009, 22, 1026–1031. [Google Scholar] [CrossRef]
- Slobodkin, A. The family Peptostreptococcaceae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 291–302. [Google Scholar]
- Tegtmeier, D.; Riese, C.; Geissinger, O.; Radek, R.; Brune, A. Breznakia blatticola gen. nov. sp. nov. and Breznakia pachnodae sp. nov., two fermenting bacteria isolated from insect guts, and emended description of the family Erysipelotrichaceae. Syst. Appl. Microbiol. 2016, 39, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Adachi, K.; Sugiyama, T.; Shimozato, A.; Ebi, M.; Ogasawara, N.; Funaki, Y.; Goto, C.; Sasaki, M.; Kasugai, K. Association of intestinal microbiota with metabolic markers and dietary habits patients with type 2 diabetes. Digestion 2016, 94, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Chang, C.C.; Mau, W.J.; Yen, L.S. Evaluation of N-acetylchitooligosaccharides as the main carbon sources for the growth of intestinal bacteria. FEMS Microbiol. Lett. 2002, 209, 53–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yew, W.C.; Pearce, D.A.; Dunn, M.J.; Samah, A.A.; Convey, P. Bacterial community composition in Adélie (Pygoscelis adeliae) and Chinstrap (Pygoscelis antarctica) Penguin stomach contents from Signy Island, South Orkney Islands. Polar Biol. 2017, 40, 2517–2530. [Google Scholar] [CrossRef]
- Tsoi, H.; Chu, E.S.H.; Zhang, X.; Sheng, J.; Nakatsu, G.; Ng, S.C.; Chan, A.W.H.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. Peptostreptococcus anaerobius induces intracellular cholesterol biosynthesis in colon cells to induce proliferation and causes dysplasia in mice. Gastroenterology 2017, 152, 1419–1433.e5. [Google Scholar] [CrossRef]
- Jarmakiewicz-Czaja, S.; Sokal, A.; Filip, R. What was first, obesity or inflammatory bowel disease? What does the gut microbiota have to do with it? Nutrients 2020, 12, 3073. [Google Scholar] [CrossRef]
- Chapagain, P.; Arivett, B.; Cleveland, B.M.; Walker, D.M.; Salem, M. Analysis of the fecal microbiota of fast- and slow-growing rainbow trout (Oncorhynchus mykiss). BMC Genom. 2019, 20, 788. [Google Scholar] [CrossRef]
- Williams, T.D. The Penguins: Spheniscidae; Oxford University Press: New York, NY, USA, 1995; pp. 1–295. [Google Scholar]
- Denger, K.; Stackebrandt, E.; Cook, A.M. Desulfonispora thiosulfatigenes gen. nov., sp. nov., a taurine-fermenting, thiosulfate-producing anaerobic bacterium. Int. J. Syst. Bacteriol. 1999, 49, 1599–1603. [Google Scholar] [PubMed]
- Opriessnig, T.; Forde, T.; Shimoji, Y. Erysipelothrix spp.: Past, present, and future directions in vaccine research. Front. Vet. Sci. 2020, 7, 174. [Google Scholar] [CrossRef]
- Boerner, L.; Nevis, K.R.; Hinckley, L.S.; Weber, E.S.; Frasca, S., Jr. Erysipelothrix septicemia in a little blue penguin (Eudyptula minor). J. Vet. Diagn. Investig. 2004, 16, 145–149. [Google Scholar] [CrossRef]
- Roggenbuck, M.; Bærholm Schnell, I.; Blom, N.; Bælum, J.; Bertelsen, M.F.; Sicheritz-Pontén, T.; Sørensen, S.J.; Gilbert, M.T.; Graves, G.R.; Hansen, L.H. The microbiome of New World vultures. Nat. Commun. 2014, 5, 5498. [Google Scholar] [CrossRef]
- Portrait, V.; Cottenceau, G.; Pons, A.M. A Fusobacterium mortiferum strain produces a bacteriocin-like substance(s) inhibiting Salmonella enteritidis. Lett. Appl. Microbiol. 2000, 31, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, C.; Coronado, J.; Silva, A.; Romero, J. Cetobacterium is a major component of the microbiome of giant Amazonian fish (Arapaima gigas) in Ecuador. Animals 2018, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Sugita, H.; Miyajima, C.; Deguchi, Y. The vitamin B12-producing ability of the intestinal microflora of freshwater fish. Aquaculture 1991, 92, 267–276. [Google Scholar] [CrossRef]
- Michel, A.J.; Ward, L.M.; Goffredi, S.K.; Dawson, K.S.; Baldassarre, D.T.; Brenner, A.; Gotanda, K.M.; McCormack, J.E.; Mullin, S.W.; O’Neill, A.; et al. The gut of the finch: Uniqueness of the gut microbiome of the Galápagos vampire finch. Microbiome 2018, 6, 167. [Google Scholar] [CrossRef]
- Dewar, M.L.; Arnould, J.P.; Allnutt, T.R.; Crowley, T.; Krause, L.; Reynolds, J.; Dann, P.; Smith, S.C. Microbiota of little penguins and short-tailed shearwaters during development. PLoS ONE 2017, 12, e0183117. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Jerzak, L.; Wilharm, G.; Golke, J.; Busse, H.J.; Glaeser, S.P. Psychrobacter ciconiae sp. nov., isolated from white storks (Ciconia ciconia). Int. J. Syst. Evol. Microbiol. 2015, 65, 772–777. [Google Scholar] [CrossRef]
- Kämpfer, P.; Glaeser, S.P.; Irgang, R.; Fernández-Negrete, G.; Poblete-Morales, M.; Fuentes-Messina, D.; Cortez-San Martín, M.; Avendaño-Herrera, R. Psychrobacter pygoscelis sp. nov. isolated from the penguin Pygoscelis papua. Int. J. Syst. Evol. Microbiol. 2020, 70, 211–219. [Google Scholar] [CrossRef]
- Sun, Y.Z.; Yang, H.L.; Ma, R.L.; Zhang, C.X.; Lin, W.Y. Effect of dietary administration of Psychrobacter sp. on the growth, feed utilization, digestive enzymes and immune responses of grouper Epinephelus coioides. Aquac. Nutr. 2011, 17, e733–e740. [Google Scholar] [CrossRef]
- Gregersen, R.H.; Neubauer, C.; Christensen, H.; Korczak, B.; Bojesen, A.M.; Hess, M.; Bisgaard, M. Characterization of Pasteurellaceae-like bacteria isolated from clinically affected psittacine birds. J. Appl. Microbiol. 2010, 108, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Riester, C.J.; Skinner, B.M.; Newell, A.W.; Swingley, W.D.; Madigan, M.T.; Jung, D.O.; Asao, M.; Chen, M.; Loughlin, P.C.; et al. Genome sequence of Rhodoferax antarcticus ANT.BRT.; A psychrophilic purple nonsulfur bacterium from an Antarctic microbial mat. Microorganisms 2017, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.D.H.; Amoroso, G.; Ventura, T.; Minich, J.J.; Elizur, A. Atlantic Salmon (Salmo salar L., 1758) gut microbiota profile correlates with flesh pigmentation: Cause or Effect? Mar. Biotechnol. 2020, 22, 786–804. [Google Scholar] [CrossRef]
- Huang, Z.; Li, X.; Wang, L.; Shao, Z. Changes in the intestinal bacterial community during the growth of white shrimp, Litopenaeus vannamei. Aquac. Res. 2016, 47, 1737–1746. [Google Scholar] [CrossRef]
- Morley, S.A.; Barnes, D.A.; Dunn, M.J. Predicting which species succeed in climate-forced polar seas. Front. Mar. Sci. 2019, 5, 507. [Google Scholar] [CrossRef]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef]
- Wang, H.W.; Miao, C.Y.; Liu, J.; Zhang, Y.; Zhu, S.Q.; Zhou, B.H. Fluoride-induced rectal barrier damage and microflora disorder in mice. Environ. Sci. Pollut. Res. Int. 2020, 27, 7596–7607. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Newsham, K.K.; Pearce, D.A.; Kowalchuk, G.A. Patterns of bacterial diversity across a range of Antarctic terrestrial habitats. Environ. Microbiol. 2007, 9, 2670–2682. [Google Scholar] [CrossRef]
- Wang, N.F.; Zhang, T.; Zhang, F.; Wang, E.T.; He, J.F.; Ding, H.; Zhang, B.T.; Liu, J.; Ran, X.B.; Zang, J.Y. Diversity and structure of soil bacterial communities in the Fildes Region (maritime Antarctica) as revealed by 454 pyrosequencing. Front. Microbiol. 2015, 6, 1188. [Google Scholar] [CrossRef]
- Oh, H.N.; Park, D.; Seong, H.J.; Kim, D.; Sul, W.J. Antarctic tundra soil metagenome as useful natural resources of cold-active lignocelluolytic enzymes. J. Microbiol. 2019, 57, 865–873. [Google Scholar] [CrossRef]
- Grond, K.; Sandercock, B.K.; Jumpponen, A.; Zeglin, L.H. The avian gut microbiota: Community, physiology and function in wild birds. J. Avian Biol. 2018, 49, e01788. [Google Scholar] [CrossRef]
- Sakamoto, M. The family porphyromonadaceae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 811–824. [Google Scholar]
- Newton, R.J.; Bootsma, M.J.; Morrison, H.G.; Sogin, M.L.; McLellan, S.L. A microbial signature approach to identify fecal pollution in the waters off an urbanized coast of Lake Michigan. Microb. Ecol. 2013, 65, 1011–1023. [Google Scholar] [CrossRef]
- Rabah, H.; Rosa do Carmo, F.L.; Jan, G. Dairy Propionibacteria: Versatile probiotics. Microorganisms 2017, 5, 24. [Google Scholar] [CrossRef]
- Kämpfer, P.; Groth, I. Intrasporangiaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 1–8. [Google Scholar]
- Wang, Z.; Yu, Z.; Zhao, J.; Zhuang, X.; Cao, P.; Guo, X.; Liu, C.; Xiang, W. Community composition, antifungal activity and chemical analyses of ant-derived Actinobacteria. Front. Microbiol. 2020, 11, 201. [Google Scholar] [CrossRef]
- Anjum, S.I.; Shah, A.H.; Aurongzeb, M.; Kori, J.; Azim, M.K.; Ansari, M.J.; Bin, L. Characterization of gut bacterial flora of Apis mellifera from north-west Pakistan. Saudi J. Biol. Sci. 2018, 25, 388–392. [Google Scholar] [CrossRef] [PubMed]
- De Salas, M.F.; Eriksen, R.; Davidson, A.T.; Wright, S.W. Protistan communities in the Australian sector of the Sub-Antarctic Zone during SAZ-Sense. Deep-Sea Res. II 2011, 58, 2135–2149. [Google Scholar] [CrossRef]
- Passmore, A.J.; Jarman, S.N.; Swadling, K.M.; Kawaguchi, S.; McMinn, A.; Nicol, S. DNA as a dietary biomarker in Antarctic Krill, Euphausia superba. Mar Biotechnol. 2006, 8, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Hagen, W.; Yoshida, T.; Virtue, P.; Kawaguchi, S.; Swadling, K.M.; Nicol, S.; Nichols, P.D. Effect of a carnivorous diet on the lipids, fatty acids and condition of Antarctic krill, Euphausia superba. Antarct. Sci. 2007, 19, 183–188. [Google Scholar] [CrossRef]
- Saunders, R.A.; Hill, S.L.; Tarling, G.A.; Murphy, E.J. Myctophid fish (family Myctophidae) are central consumers in the food web of the Scotia Sea (Southern Ocean). Front. Mar. Sci. 2019, 6, 530. [Google Scholar] [CrossRef]
- Schmidt, K.; Jónasdóttir, S.H. Nutritional quality of two cyanobacteria: How rich is ‘poor’ food? Mar. Ecol. Prog. Ser. 1997, 151, 1–10. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J. Bringing Planctomycetes into pure culture. Front. Microbiol. 2012, 3, 405. [Google Scholar] [CrossRef] [PubMed]
- Schlesner, H. The development of media suitable for the microorganisms morphologically resembling Planctomyces spp., Pirellula spp., and other Planctomycetales from various aquatic habitats using dilute media. Syst. Appl. Microbiol. 1994, 17, 135–145. [Google Scholar] [CrossRef]
- Yu, C.; Lee, A.M.; Bassler, B.L.; Roseman, S. Chitin utilization by marine bacteria. A physiological function for bacterial adhesion to immobilized carbohydrates. J. Biol. Chem. 1991, 266, 24260–24267. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fecal Sample | Average Reads | Average OTUs | Average Shannon | Average Simpson | Average Fisher’s Alpha | Average Ace | Average Chao 1 | Average Coverage |
|---|---|---|---|---|---|---|---|---|
| Adélie | 49733.00 ± 9503.48 | 266.37 ± 64.66 | 2.71 ± 0.55 | 0.16 ± 0.07 | 37.29 ±10.72 | 357.52 ± 108.91 | 336.08 ± 84.78 | 0.99 ±0 |
| Gentoo | 47314.66 ± 11939.60 | 394.75 ± 128.72 | 3.15 ± 0.81 | 0.13 ± 0.11 | 60.57 ± 23.28 | 503.06 ± 139.06 | 505.64 ± 148.39 | 0.99 ± 0 |
| Total | 47798.33 ± 11422.9 | 369.07 ± 128.94 | 3.06 ± 0.78 | 0.13 ± 0.10 | 55.91 ± 23.24 | 473.95 ± 144.83 | 471.73 ± 153.33 | 0.99 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.-X.; Li, H.-R.; Han, W.; Luo, W. Comparison of Gut Microbiota between Gentoo and Adélie Penguins Breeding Sympatrically on Antarctic Ardley Island as Revealed by Fecal DNA Sequencing. Diversity 2021, 13, 500. https://doi.org/10.3390/d13100500
Zeng Y-X, Li H-R, Han W, Luo W. Comparison of Gut Microbiota between Gentoo and Adélie Penguins Breeding Sympatrically on Antarctic Ardley Island as Revealed by Fecal DNA Sequencing. Diversity. 2021; 13(10):500. https://doi.org/10.3390/d13100500
Chicago/Turabian StyleZeng, Yin-Xin, Hui-Rong Li, Wei Han, and Wei Luo. 2021. "Comparison of Gut Microbiota between Gentoo and Adélie Penguins Breeding Sympatrically on Antarctic Ardley Island as Revealed by Fecal DNA Sequencing" Diversity 13, no. 10: 500. https://doi.org/10.3390/d13100500
APA StyleZeng, Y.-X., Li, H.-R., Han, W., & Luo, W. (2021). Comparison of Gut Microbiota between Gentoo and Adélie Penguins Breeding Sympatrically on Antarctic Ardley Island as Revealed by Fecal DNA Sequencing. Diversity, 13(10), 500. https://doi.org/10.3390/d13100500