
Phaseolus coccineus Seed: A Valued Resource for Bioactive Compounds Targeting Health and Tumor Cells

 , ,
, ,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Extract Preparation
- PaP—it was treated with ViscozymeR + Papain;
- CtrlPaP—it underwent the same processes as PaP (1) without the addition of any enzyme;
- PaP100—it was treated with ViscozymeR + Papain and subsequently boiled;
- CtrlPaP100—it underwent the same processes as PaP100 (3) without the addition of any enzyme;
- AsP—it was treated with ViscozymeR + Aspergillopepsin I;
- CtrlAsP—it underwent the same processes as AsP (5) without the addition of any enzyme;
- AsP100—it was treated with ViscozymeR + Aspergillopepsin I and subsequently boiled;
- CtrlAsP100—it underwent the same processes as AsP100 (7) without the addition of any enzyme.
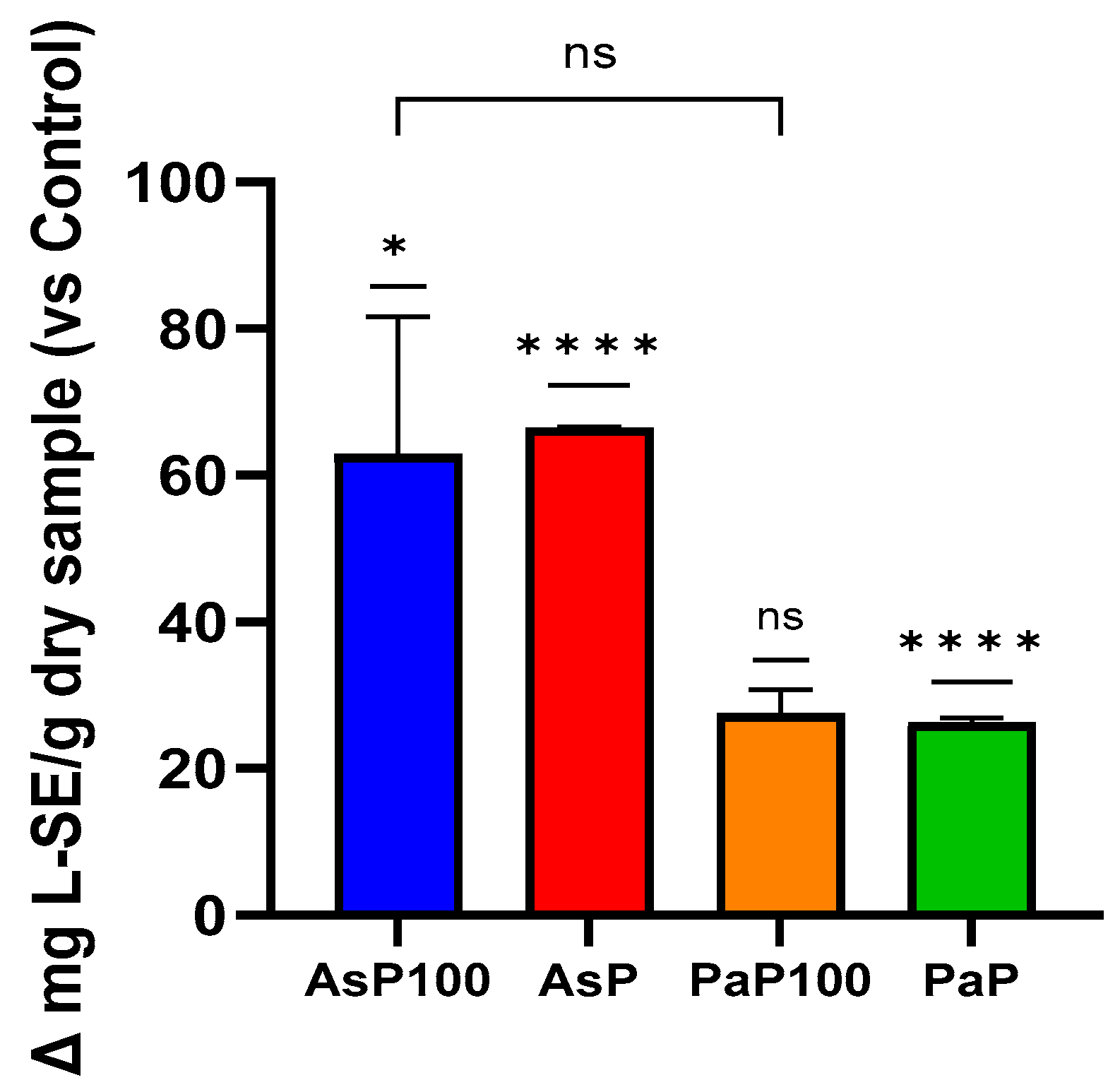
2.2. Peptide Content
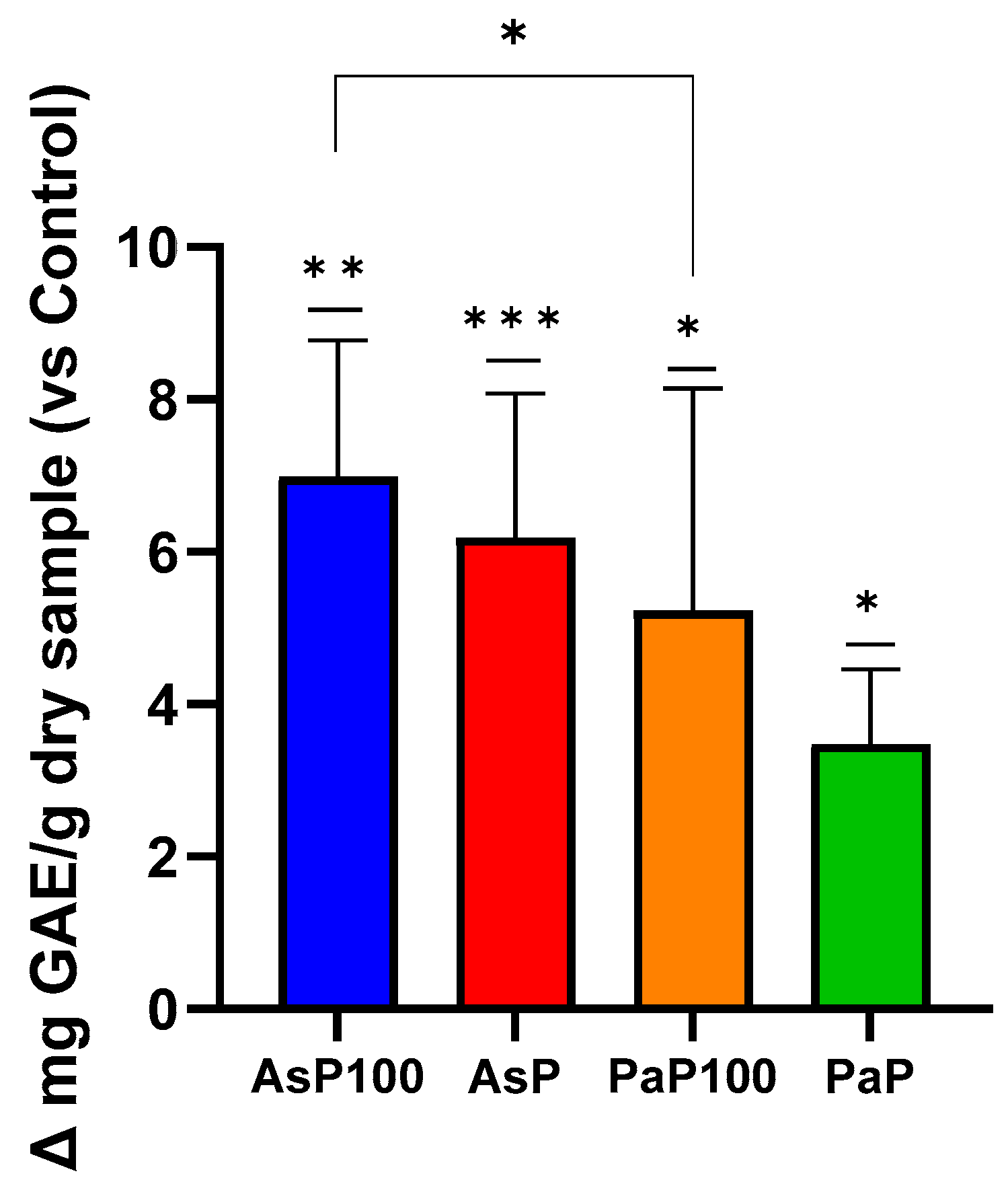
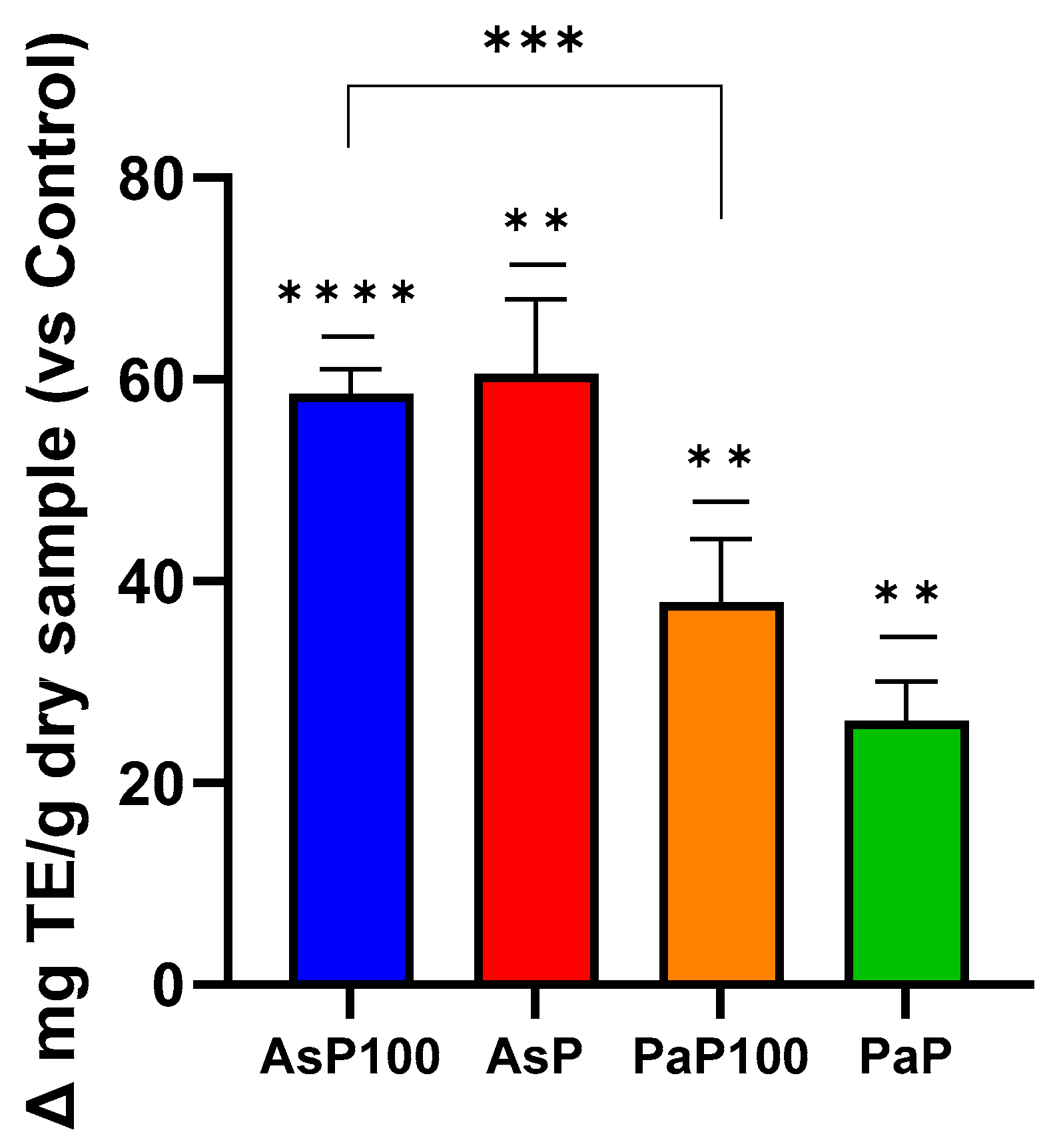
2.3. Total Polyphenol Content and In Vitro Antioxidant Activity
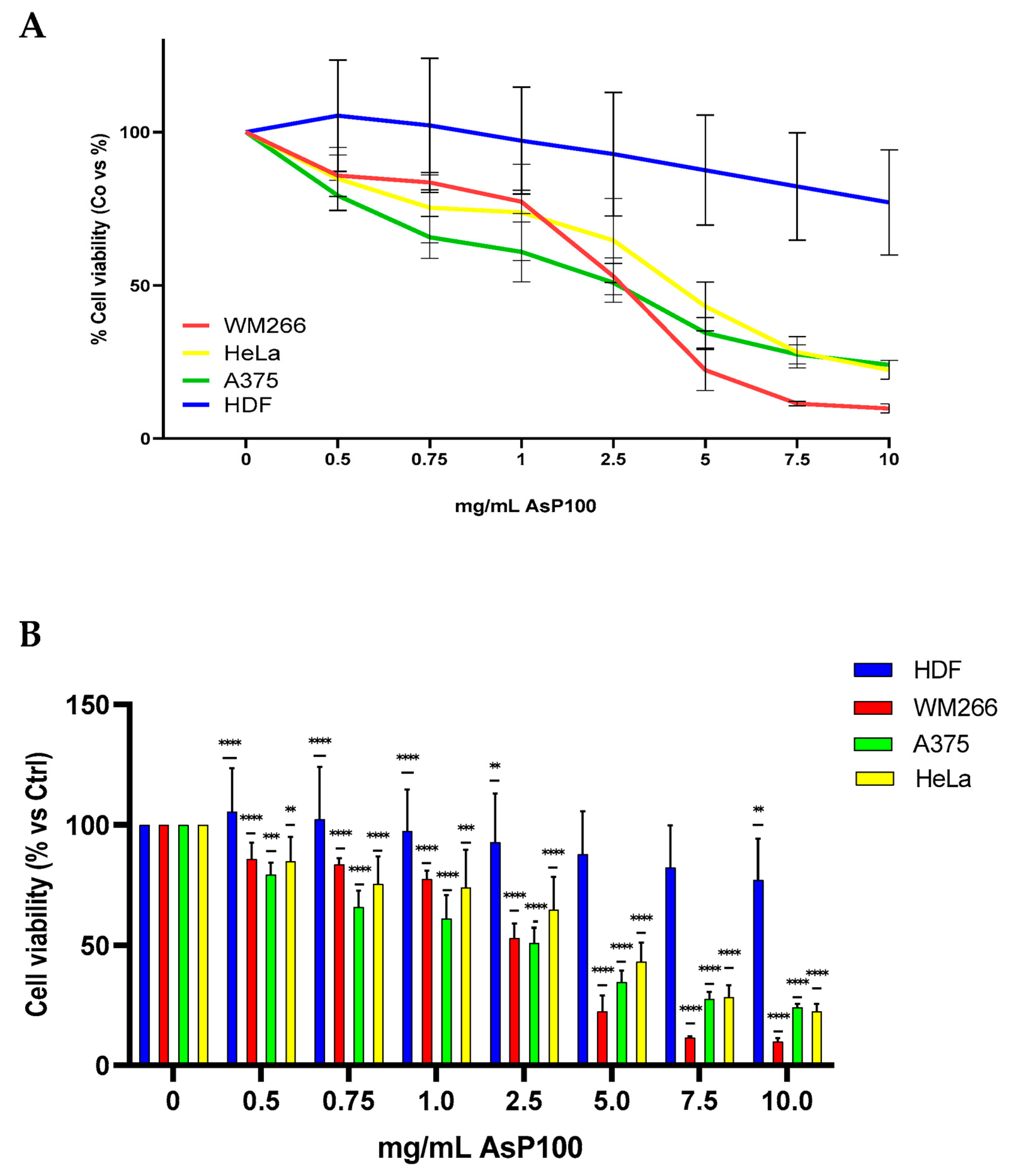
2.4. Evaluation of Antiproliferative Activity
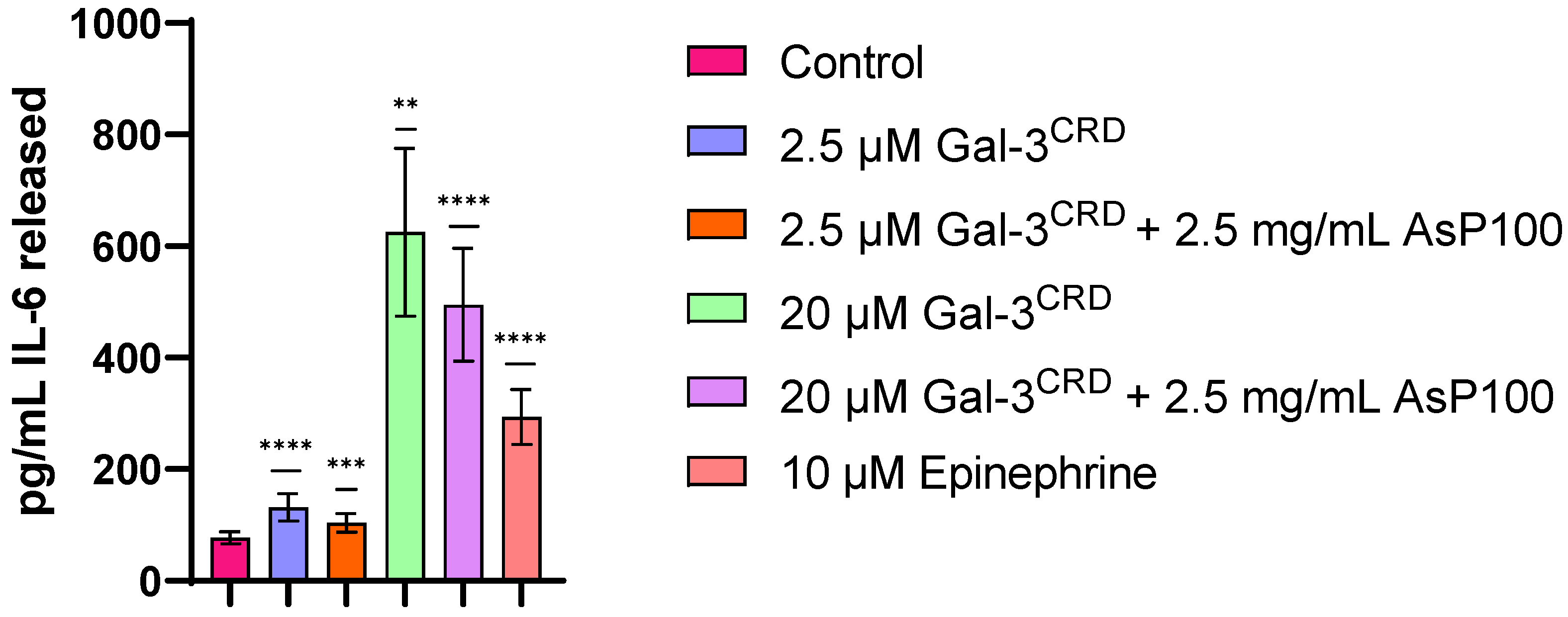
2.5. IL-6 Released Evaluation
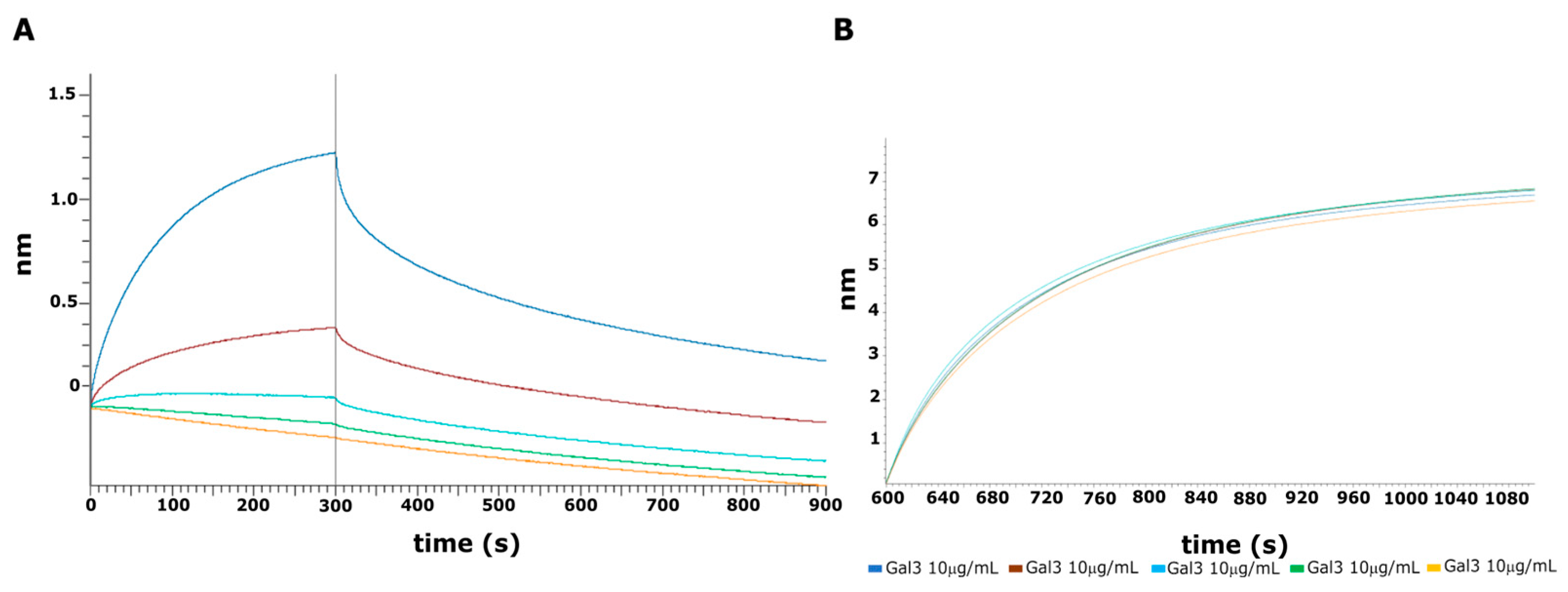
2.6. BLI Interaction Analysis
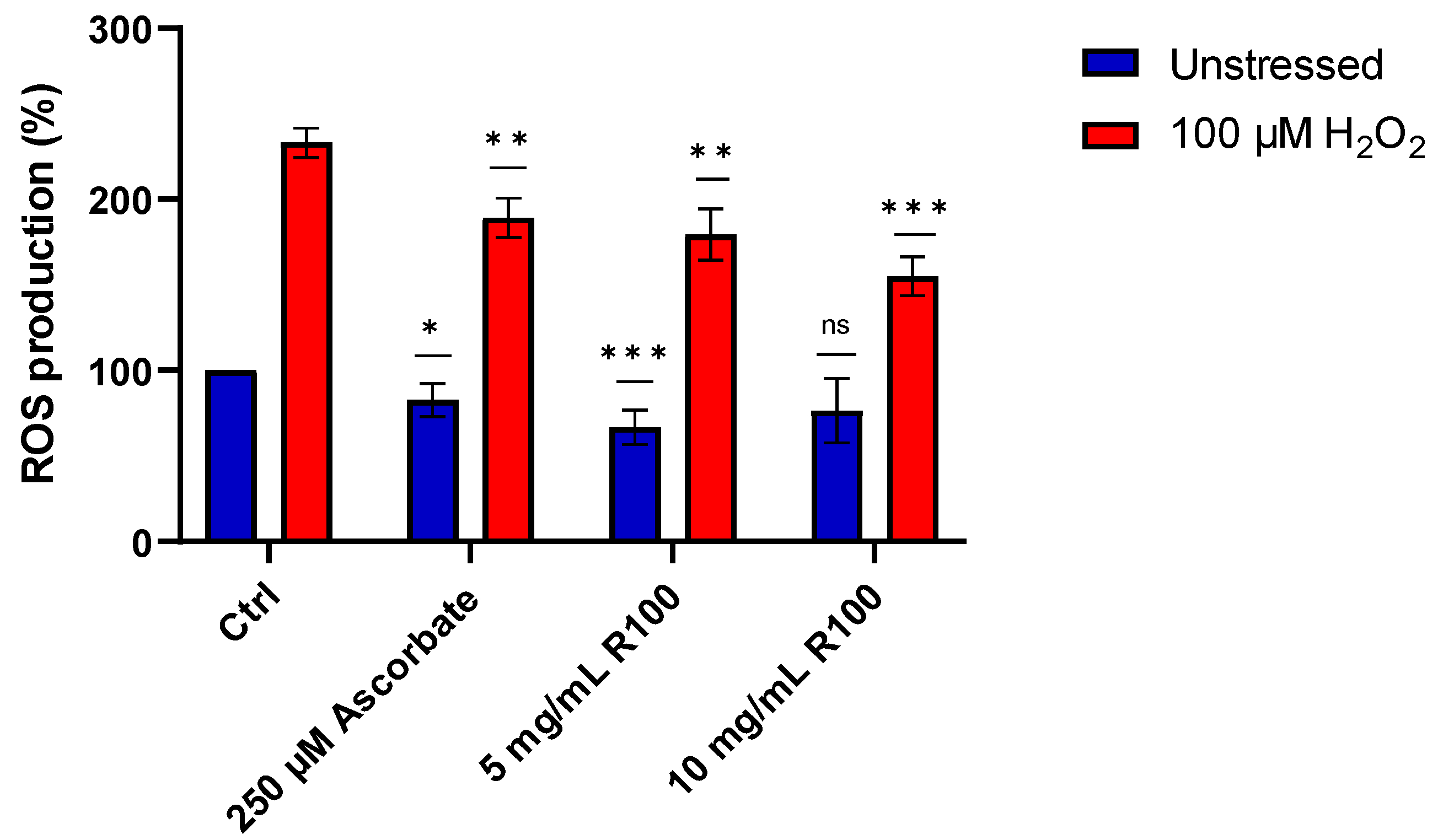
2.7. ROS Assay
3. Materials and Methods
3.1. Extract Preparation
3.2. Peptide Content
3.3. Total Polyphenol Content
3.4. In Vitro Antioxidant Activity
3.5. Cell Lines and Culture Condition
3.6. Cell Proliferation Assay
3.7. ELISA for IL-6 Detection
3.8. ROS Detection
3.9. Protein Expression and Purification
3.10. BLI Interaction Analysis
3.11. Statistical Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Gal-3 | Galectin-3 |
| CRD | Carbohydrate Recognition Domain |
| OPA | o-phthalaldehyde |
| HDF | Human dermal fibroblast |
| HeLa | Human cervical carcinoma |
References
- Lin, Y.-H.; Qiu, D.-C.; Chang, W.-H.; Yeh, Y.-Q.; Jeng, U.-S.; Liu, F.-T.; Huang, J. The Intrinsically Disordered N-Terminal Domain of Galectin-3 Dynamically Mediates Multisite Self-Association of the Protein through Fuzzy Interactions. J. Biol. Chem. 2017, 292, 17845–17856. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediat. Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef]
- Pirone, L.; Lenza, M.P.; Di Gaetano, S.; Capasso, D.; Filocaso, M.; Russo, R.; Di Carluccio, C.; Saviano, M.; Silipo, A.; Pedone, E. Biophysical and Structural Characterization of the Interaction between Human Galectin-3 and the Lipopolysaccharide from Pseudomonas Aeruginosa. Int. J. Mol. Sci. 2024, 25, 2895. [Google Scholar] [CrossRef]
- Bouffette, S.; Botez, I.; Ceuninck, F.D. Targeting Galectin-3 in Inflammatory and Fibrotic Diseases. Trends Pharmacol. Sci. 2023, 44, 519–531. [Google Scholar] [CrossRef]
- Machado, C.M.L.; Andrade, L.N.S.; Teixeira, V.R.; Costa, F.F.; Melo, C.M.; Dos Santos, S.N.; Nonogaki, S.; Liu, F.; Bernardes, E.S.; Camargo, A.A.; et al. Galectin-3 Disruption Impaired Tumoral Angiogenesis by Reducing VEGF Secretion from TGF β 1-induced Macrophages. Cancer Med. 2014, 3, 201–214. [Google Scholar] [CrossRef]
- Fortuna-Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S.G. Extracellular Galectin-3 in Tumor Progression and Metastasis. Front. Oncol. 2014, 4, 138. [Google Scholar] [CrossRef]
- Srejovic, I.; Selakovic, D.; Jovicic, N.; Jakovljević, V.; Lukic, M.L.; Rosic, G. Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior. Biomolecules 2020, 10, 798. [Google Scholar] [CrossRef]
- Blanchard, H.; Yu, X.; Collins, P.M.; Bum-Erdene, K. Galectin-3 Inhibitors: A Patent Review (2008–Present). Expert Opin. Ther. Pat. 2014, 24, 1053–1065. [Google Scholar] [CrossRef]
- Denavit, V.; Lainé, D.; Tremblay, T.; St-Gelais, J.; Giguère, D. Synthetic Inhibitors of Galectins: Structures and Syntheses. Trends Glycosci. Glycotechnol. 2018, 30, SE21–SE40. [Google Scholar] [CrossRef]
- Slack, R.J.; Hirani, N.; Gibbons, M.A.; Simpson, A.J.; Ford, P.; Leffler, H.; Nilsson, U.J.; Sethi, T.; Pedersen, A.; Schambye, H.; et al. Translational Pharmacology of TD139, an Inhaled Small Molecule Galectin-3 (Gal-3) Inhibitor for the Treatment of Idiopathic Pulmonary Fibrosis (IPF). FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Di Gaetano, S.; Pirone, L.; Galdadas, I.; Traboni, S.; Iadonisi, A.; Pedone, E.; Saviano, M.; Gervasio, F.L.; Capasso, D. Design, Synthesis, and Anticancer Activity of a Selenium-Containing Galectin-3 and Galectin-9N Inhibitor. Int. J. Mol. Sci. 2022, 23, 2581. [Google Scholar] [CrossRef]
- Traber, P.G.; Zomer, E. Therapy of Experimental NASH and Fibrosis with Galectin Inhibitors. PLoS ONE 2013, 8, e83481. [Google Scholar] [CrossRef]
- Traber, P.G.; Chou, H.; Zomer, E.; Hong, F.; Klyosov, A.; Fiel, M.I.; Friedman, S.L. Regression of Fibrosis and Reversal of Cirrhosis in Rats by Galectin Inhibitors in Thioacetamide-Induced Liver Disease. PLoS ONE 2013, 8, e75361. [Google Scholar] [CrossRef]
- Gao, X.; Zhi, Y.; Zhang, T.; Xue, H.; Wang, X.; Foday, A.D.; Tai, G.; Zhou, Y. Analysis of the Neutral Polysaccharide Fraction of MCP and Its Inhibitory Activity on Galectin-3. Glycoconj. J. 2012, 29, 159–165. [Google Scholar] [CrossRef]
- Galectin Therapeutics Inc. A Seamless, Adaptive, Phase 2b/3, Double-Blind, Randomized, Placebo-Controlled, Multicenter, International Study Evaluating the Efficacy and Safety of Belapectin (GR MD-02) for the Prevention of Esophageal Varices in NASH Cirrhosis; Galectin Therapeutics Inc.: Norcross, GA, USA, 2023. [Google Scholar]
- Tito, A.; Colantuono, A.; Pirone, L.; Pedone, E.; Intartaglia, D.; Giamundo, G.; Conte, I.; Vitaglione, P.; Apone, F. Pomegranate Peel Extract as an Inhibitor of SARS-CoV-2 Spike Binding to Human ACE2 Receptor (in Vitro): A Promising Source of Novel Antiviral Drugs. Front. Chem. 2021, 9, 638187. [Google Scholar] [CrossRef]
- Tang, B.; Jin, C.; Li, M.; Liu, S.; Zhang, X.; Li, J.; Ding, K.; Zang, Y. A Novel Pectin-like Polysaccharide from Crocus Sativus Targets Galectin-3 to Inhibit Hepatic Stellate Cells Activation and Liver Fibrosis. Carbohydr. Polym. 2025, 348, 122826. [Google Scholar] [CrossRef]
- Baeza-Jiménez, R.; López-Martínez, L.X. Changes in Phenolic Composition and Bioactivities of Ayocote Beans under Boiling (Phaseolus coccineus L.). Molecules 2024, 29, 3744. [Google Scholar] [CrossRef]
- Alcázar-Valle, M.; Lugo-Cervantes, E.; Mojica, L.; Morales-Hernández, N.; Reyes-Ramírez, H.; Enríquez-Vara, J.N.; García-Morales, S. Bioactive Compounds, Antioxidant Activity, and Antinutritional Content of Legumes: A Comparison between Four Phaseolus Species. Molecules 2020, 25, 3528. [Google Scholar] [CrossRef]
- AAT_Bioquest_Inc. Quest Graph™ IC50_Calculator._AAT_Bioquest. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 17 February 2025).
- Adascălului, M.; Multescu, M.; Mihai, A.L.; Bobea, S.A.; Florea, C.; Belc, N. Cytotoxicity Assessment and Nutritional Profiling of Bio-Active Compounds Obtained from FoodWaste. Processes 2023, 11, 89. [Google Scholar] [CrossRef]
- Lee, K.S.; Drescher, D.G. Fluorometric Amino-Acid Analysis with o-Phthaldialdehyde (OPA). Int. J. Biochem. 1978, 9, 457–467. [Google Scholar] [CrossRef]
- Box, J.D. Investigation of the Folin-Ciocalteau Phenol Reagent for the Determination of Polyphenolic Substances in Natural Waters. Water Res. 1983, 17, 511–525. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Donato, M.T.; Gómez-Lechón, M.J. General Cytotoxicity Assessment by Means of the MTT Assay. In Protocols in In Vitro Hepatocyte Research; Vinken, M., Rogiers, V., Eds.; Springer: New York, NY, USA, 2015; pp. 333–348. ISBN 978-1-4939-2074-7. [Google Scholar]
- Oparka, M.; Walczak, J.; Malinska, D.; van Oppen, L.M.P.E.; Szczepanowska, J.; Koopman, W.J.H.; Wieckowski, M.R. Quantifying ROS Levels Using CM-H2DCFDA and HyPer. Methods 2016, 109, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Pirone, L.; Esposito, C.; Correale, S.; Graziano, G.; Di Gaetano, S.; Vitagliano, L.; Pedone, E. Thermal and Chemical Stability of Two Homologous POZ/BTB Domains of KCTD Proteins Characterized by a Different Oligomeric Organization. Biomed. Res. Int. 2013, 2013, 162674. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, R.; Colantuono, A.; Di Gaetano, S.; Capasso, D.; Tito, A.; Pedone, E.; Pirone, L. Phaseolus coccineus Seed: A Valued Resource for Bioactive Compounds Targeting Health and Tumor Cells. Int. J. Mol. Sci. 2025, 26, 2189. https://doi.org/10.3390/ijms26052189
Russo R, Colantuono A, Di Gaetano S, Capasso D, Tito A, Pedone E, Pirone L. Phaseolus coccineus Seed: A Valued Resource for Bioactive Compounds Targeting Health and Tumor Cells. International Journal of Molecular Sciences. 2025; 26(5):2189. https://doi.org/10.3390/ijms26052189
Chicago/Turabian StyleRusso, Rita, Antonio Colantuono, Sonia Di Gaetano, Domenica Capasso, Annalisa Tito, Emilia Pedone, and Luciano Pirone. 2025. "Phaseolus coccineus Seed: A Valued Resource for Bioactive Compounds Targeting Health and Tumor Cells" International Journal of Molecular Sciences 26, no. 5: 2189. https://doi.org/10.3390/ijms26052189
APA StyleRusso, R., Colantuono, A., Di Gaetano, S., Capasso, D., Tito, A., Pedone, E., & Pirone, L. (2025). Phaseolus coccineus Seed: A Valued Resource for Bioactive Compounds Targeting Health and Tumor Cells. International Journal of Molecular Sciences, 26(5), 2189. https://doi.org/10.3390/ijms26052189