
Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review


Abstract
1. Introduction
2. Traditional Biomarkers
2.1. HbA1c
2.2. Fructosamine
2.3. Glycated Albumin
2.4. Oral Glucose Tolerance Test

{kind=link}
| Biomarker | Expression | Target | Clinical Value | Reference |
|---|---|---|---|---|
| miRNA-375 | Up-/Downregulated | MTPN, PDK1 | Dysregulated levels can be detected early compared to traditional biomarkers in serum of high-risk groups. Can be utilized for investigative procedures for early detection of diabetes and prediabetes in high-risk groups. | [57,58,59] |
| miRNA-126 | Downregulated | IRS-1 | Investigative utility—may be used as part of screening tool for early detection of diabetes, as well as microvascular and retinal complications. Can further monitor levels to observe response to treatment. | [22] |
| miRNA-7a | Upregulated | IRS-2 | Increased expression observed in the serum aids in identifying dysregulated insulin signaling and may be used as an investigative tool to detect and monitor response to treatment. | [60,61] |
| Let-7 | Upregulated | IGF1R, INSR, IRS-2 | Increased levels in the plasma suggest impaired/decreased insulin secretion, a key factor in the pathogenesis of T2DM. Potential to aid in early detection of diabetes in clinic. | [62] |
3. MicroRNA
4. MicroRNAs Role in T2DM
4.1. miRNA-375
4.2. miRNA-126
4.3. miRNA-7a
4.4. Let-7
5. miRNAs in T2DM-Associated Complications
5.1. Diabetic Cardiomyopathy
5.2. Atherosclerosis
5.3. Diabetic Nephropathy
5.4. Diabetic Retinopathy
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association Professional Practice Committee. Summary of Revisions: Standards of Care in Diabetes—2024. Diabetes Care 2023, 47 (Suppl. 1), S5–S10.
- Mezza, T.; Cinti, F.; Cefalo, C.M.A.; Pontecorvi, A.; Kulkarni, R.N.; Giaccari, A. β-Cell Fate in Human Insulin Resistance and Type 2 Diabetes: A Perspective on Islet Plasticity. Diabetes 2019, 68, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Aghaei Zarch, S.M.; Dehghan Tezerjani, M.; Talebi, M.; Vahidi Mehrjardi, M.Y. Molecular biomarkers in diabetes mellitus (DM). Med. J. Islam. Repub. Iran 2020, 34, 28. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Agarwal, R. Pathogenesis of Diabetic Nephropathy. In Chronic Kidney Disease and Type 2 Diabetes; ADA Clinical Compendia Series; American Diabetes Association: Arlington, VA, USA, 2021; pp. 2–7. [Google Scholar]
- Ahmed, A.; Majeed, S.; Obaid, H.; Al-Hmmamy, S. Biochemistry and Molecular Cell Biology of Diabetic Complications. Syst. Rev. Pharm. 2020, 11, 850–860. [Google Scholar]
- Bansal, N. Prediabetes diagnosis and treatment: A review. World J. Diabetes 2015, 6, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Al-Muhtaresh, H.A.; Al-Kafaji, G. Evaluation of Two-Diabetes Related microRNAs Suitability as Earlier Blood Biomarkers for Detecting Prediabetes and type 2 Diabetes Mellitus. J. Clin. Med. 2018, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ohkuma, T.; Cooper, M.; Harrap, S.; Mancia, G.; Poulter, N.; Wang, J.-G.; Zoungas, S.; Woodward, M.; Chalmers, J. Effects of Intensive Glycemic Control on Clinical Outcomes Among Patients With Type 2 Diabetes With Different Levels of Cardiovascular Risk and Hemoglobin A1c in the ADVANCE Trial. Diabetes Care 2020, 43, 1293–1299. [Google Scholar] [CrossRef]
- Antwi, J. Precision Nutrition to Improve Risk Factors of Obesity and Type 2 Diabetes. Curr. Nutr. Rep. 2023, 12, 679–694. [Google Scholar] [CrossRef] [PubMed]
- Uusitupa, M.; Khan, T.A.; Viguiliouk, E.; Kahleova, H.; Rivellese, A.A.; Hermansen, K.; Pfeiffer, A.; Thanopoulou, A.; Salas-Salvadó, J.; Schwab, U.; et al. Prevention of Type 2 Diabetes by Lifestyle Changes: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 2611. [Google Scholar] [CrossRef]
- Su, J.; Luo, Y.; Hu, S.; Tang, L.; Ouyang, S. Advances in Research on Type 2 Diabetes Mellitus Targets and Therapeutic Agents. Int. J. Mol. Sci. 2023, 24, 13381. [Google Scholar] [CrossRef] [PubMed]
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Tuomilehto, J.; Schwarz, P.E. Preventing Diabetes: Early Versus Late Preventive Interventions. Diabetes Care 2016, 39 (Suppl. 2), S115–S120. [Google Scholar] [CrossRef] [PubMed]
- Califf, R.M. Biomarker definitions and their applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Sato Imuro, S.E.; Sabharwal, A.; Bevier, W.; Kerr, D. Evaluating HbA(1c)-to-average glucose conversion with patient-specific kinetic models for diverse populations. Sci. Rep. 2024, 14, 22098. [Google Scholar] [CrossRef]
- Cs, L.; Aw, T.-C. HbA1c in the diagnosis and management of diabetes mellitus: An update. Diabetes Updates 2020, 6, 1–4. [Google Scholar] [CrossRef]
- Ling, C.; Bacos, K.; Rönn, T. Epigenetics of type 2 diabetes mellitus and weight change—A tool for precision medicine? Nat. Rev. Endocrinol. 2022, 18, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Panni, S.; Pizzolotto, R. Integrated Analysis of microRNA Targets Reveals New Insights into Transcriptional–Post-Transcriptional Regulatory Cross-Talk. Biology 2025, 14, 43. [Google Scholar] [CrossRef]
- Vaghf, A.; Khansarinejad, B.; Ghaznavi-Rad, E.; Mondanizadeh, M. The role of microRNAs in diseases and related signaling pathways. Mol. Biol. Rep. 2022, 49, 6789–6801. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, H.; Si, H.; Li, X.; Ding, X.; Sheng, Q.; Chen, P.; Zhang, H. Serum miR-23a, a potential biomarker for diagnosis of pre-diabetes and type 2 diabetes. Acta Diabetol. 2014, 51, 823–831. [Google Scholar] [CrossRef]
- Nigi, L.; Grieco, G.E.; Ventriglia, G.; Brusco, N.; Mancarella, F.; Formichi, C.; Dotta, F.; Sebastiani, G. MicroRNAs as Regulators of Insulin Signaling: Research Updates and Potential Therapeutic Perspectives in Type 2 Diabetes. Int. J. Mol. Sci. 2018, 19, 3705. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hng, T.-M. HbA1c: More than just a number. Aust. J. Gen. Pract. 2021, 50, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Mukonda, E.; van der Westhuizen, D.J.; Dave, J.A.; Cleary, S.; Hannan, L.; Rusch, J.A.; Lesosky, M. Understanding the relationship between the frequency of HbA1c monitoring, HbA1c changes over time, and the achievement of targets: A retrospective cohort study. BMC Endocr. Disord. 2025, 25, 3. [Google Scholar] [CrossRef]
- NICE. Type 2 Diabetes in Adults: Management: NICE. 2022. Available online: https://www.nice.org.uk/guidance/ng28 (accessed on 30 July 2024).
- Hussain, N. Implications of using HBA1(C) as a diagnostic marker for diabetes. Diabetol. Int. 2016, 7, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasamurthy, L. Evolution in Diagnosis and Classification of Diabetes. J. Diabetes Mellit. 2021, 11, 200–207. [Google Scholar] [CrossRef]
- Hellgren, M.; Hjörleifsdottir Steiner, K.; Bennet, L. Haemoglobin A1c as a screening tool for type 2 diabetes and prediabetes in populations of Swedish and Middle-East ancestry. Prim. Care Diabetes 2017, 11, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Karakaya, J.; Akin, S.; Karagaoglu, E.; Gurlek, A. The performance of hemoglobin A1c against fasting plasma glucose and oral glucose tolerance test in detecting prediabetes and diabetes. J. Res. Med. Sci. 2014, 19, 1051–1057. [Google Scholar]
- Hong, J.W.; Ku, C.R.; Noh, J.H.; Ko, K.S.; Rhee, B.D.; Kim, D.-J. Association Between the Presence of Iron Deficiency Anemia and Hemoglobin A1c in Korean Adults: The 2011–2012 Korea National Health and Nutrition Examination Survey. Medicine 2015, 94, e825. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, B.A.; Salamatullah, H.K.; Alsharm, F.S.; Baljoon, J.M.; Abukhodair, A.O.; Ahmed, M.E.; Malaikah, H.; Radi, S. The effect of different types of anemia on HbA1c levels in non-diabetics. BMC Endocr. Disord. 2023, 23, 24. [Google Scholar] [CrossRef] [PubMed]
- Seker, R.; Sahin, E.A.; Sener, A. HbA1c levels in iron deficiency anemia cases grouped according to hemoglobin levels. Ann. Med. Res. 2024, 31, 622–626. [Google Scholar] [CrossRef]
- Cavagnolli, G.; Pimentel, A.L.; Freitas, P.A.C.; Gross, J.L.; Camargo, J.L. Factors affecting A1C in non-diabetic individuals: Review and meta-analysis. Clin. Chim. Acta 2015, 445, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Floyd, C.N.; Brady, S.; Monteiro, D.; Nathan, Y.; Crook, M. Unexpected high HbA1c results due to an unusual haemoglobin variant: An important clinical lesson. Postgrad. Med. J. 2022, 98, 331–332. [Google Scholar] [CrossRef]
- Nansseu, J.R.N.; Fokom-Domgue, J.; Noubiap, J.J.N.; Balti, E.V.; Sobngwi, E.; Kengne, A.P. Fructosamine measurement for diabetes mellitus diagnosis and monitoring: A systematic review and meta-analysis protocol. BMJ Open 2015, 5, e007689. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E. Alternative biomarkers for assessing glycemic control in diabetes: Fructosamine, glycated albumin, and 1,5-anhydroglucitol. Ann. Pediatr. Endocrinol. Metab. 2015, 20, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, M.T.K.; Cukier, P.; Damascena, A.S.; Batista, R.L.; de Azevedo Correa, F.; Zanatta Kawahara, E.; Minanni, C.A.; Hoff, A.O.; Nery, M. Fructosamine and glycated hemoglobin as biomarkers of glycemic control in people with type 2 diabetes mellitus and cancer (GlicoOnco study). Clinics 2023, 78, 100240. [Google Scholar] [CrossRef] [PubMed]
- Senapathi, S.H.; Bhavsar, R.; Kaur, R.; Kim, P.; Sachmechi, I. A Case Report of Fructosamine’s Unreliability as a Glycemic Control Assessment Tool in Nephrotic Syndrome. Cureus 2017, 9, e1694. [Google Scholar] [CrossRef] [PubMed]
- Malmström, H.; Walldius, G.; Grill, V.; Jungner, I.; Gudbjörnsdottir, S.; Hammar, N. Fructosamine is a useful indicator of hyperglycaemia and glucose control in clinical and epidemiological studies—Cross-sectional and longitudinal experience from the AMORIS cohort. PLoS ONE 2014, 9, e111463. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.; Loomis, S.J.; Venkataraghavan, S.; Zhang, J.; Tin, A.; Yu, B.; Chatterjee, N.; Selvin, E.; Duggal, P. Characterizing Common and Rare Variations in Nontraditional Glycemic Biomarkers Using Multivariate Approaches on Multiancestry ARIC Study. Diabetes 2024, 73, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Freitas, P.A.C.; Ehlert, L.R.; Camargo, J.L. Glycated albumin: A potential biomarker in diabetes. Arch. Endocrinol. Metab. 2017, 61, 296–304. [Google Scholar] [CrossRef]
- Kumari, N.; Vaishnav, M.; Srikanta, S.; Krishnaswamy, P.; Bhat, N. Exploring Glycated Sites in Human Serum Albumin: Impact of Sample Processing Techniques on Detection and Analysis. Anal. Methods 2024, 16, 5239–5247. [Google Scholar] [CrossRef] [PubMed]
- Moman, R.N.; Gupta, N.; Varacallo, M. Physiology, Albumin; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Fang, M.; Daya, N.; Coresh, J.; Christenson, R.H.; Selvin, E. Glycated Albumin for the Diagnosis of Diabetes in US Adults. Clin. Chem. 2022, 68, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Matsha, T.E.; Korf, M.; Erasmus, R.T.; Hoffmann, M.; Mapfumo, C.; Smit, F.; Zemlin, A.E. Reference interval determination for glycated albumin in defined subgroups of a South African population. Ann. Clin. Biochem. 2019, 56, 480–487. [Google Scholar] [CrossRef]
- Freitas, P.A.C.; Hernandez, M.K.; Camargo, J.L. Factors associated with glycated albumin in adults without diabetes. Med. Pharm. Rep. 2021, 94, 170–175. [Google Scholar] [CrossRef]
- Danese, E.; Montagnana, M.; Nouvenne, A.; Lippi, G. Advantages and pitfalls of fructosamine and glycated albumin in the diagnosis and treatment of diabetes. J. Diabetes Sci. Technol. 2015, 9, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chume, F.C.; Schiavenin, L.G.; Freitas, P.A.C.; Pimentel, A.L.; Camargo, J.L. The usefulness of glycated albumin in patients with diabetes and renal disease: A scoping review. J. Lab. Precis. Med. 2022, 7, 12. [Google Scholar] [CrossRef]
- Tang, M.; Berg, A.H.; Zheng, H.; Rhee, E.P.; Allegretti, A.S.; Nigwekar, S.U.; Karumanchi, S.A.; Lash, J.P.; Kalim, S. Glycated Albumin and Adverse Clinical Outcomes in Patients With CKD: A Prospective Cohort Study. Am. J. Kidney Dis. 2024, 84, 329–338. [Google Scholar] [CrossRef]
- Jagannathan, R.; Neves, J.S.; Dorcely, B.; Chung, S.T.; Tamura, K.; Rhee, M.; Bergman, M. The Oral Glucose Tolerance Test: 100 Years Later. Diabetes Metab. Syndr. Obes. 2020, 13, 3787–3805. [Google Scholar] [CrossRef] [PubMed]
- Thewjitcharoen, Y.; Jones Elizabeth, A.; Butadej, S.; Nakasatien, S.; Chotwanvirat, P.; Wanothayaroj, E.; Krittiyawong, S.; Himathongkam, T.; Himathongkam, T. Performance of HbA1c versus oral glucose tolerance test (OGTT) as a screening tool to diagnose dysglycemic status in high-risk Thai patients. BMC Endocr. Disord. 2019, 19, 23. [Google Scholar] [CrossRef]
- Eyth, E.; Basit, H.; Swift, C.J. Glucose Tolerance Test; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- American Diabetes Association Professional Practice Committee. 2. Diagnosis and Classification of Diabetes: Standards of Care in Diabetes—2024. Diabetes Care 2023, 47 (Suppl. 1), S20–S42. [Google Scholar]
- Rao, S.S.; Disraeli, P.; McGregor, T. Impaired glucose tolerance and impaired fasting glucose. Am. Fam. Physician 2004, 69, 1961–1968. [Google Scholar] [PubMed]
- Ligthart, S.; van Herpt, T.T.W.; Leening, M.J.G.; Kavousi, M.; Hofman, A.; Stricker, B.H.C.; van Hoek, M.; Sijbrands, E.J.G.; Franco, O.H.; Dehghan, A. Lifetime risk of developing impaired glucose metabolism and eventual progression from prediabetes to type 2 diabetes: A prospective cohort study. Lancet Diabetes Endocrinol. 2016, 4, 44–51. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2022. Diabetes Care 2022, 45 (Suppl. 1), S17–S38. [Google Scholar] [CrossRef]
- Kaur, P.; Kotru, S.; Singh, S.; Behera, B.S.; Munshi, A. Role of miRNAs in the pathogenesis of T2DM, insulin secretion, insulin resistance, and β cell dysfunction: The story so far. J. Physiol. Biochem. 2020, 76, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Raina, S.; Chugh, J.; Sharma, S. MIRNAS: Early Prognostic Biomarkers for Type 2 Diabetes Mellitus? Biomark. Med. 2015, 9, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Y.; Man, B.; Li, D. Assessing MicroRNA-375 Levels in Type 2 Diabetes Mellitus (T2DM) Patients and Their First-Degree Relatives with T2DM. Diabetes Metab. Syndr. Obes. 2021, 14, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Wang, J.; Wang, J.; Wu, J.; Song, J.; Zhang, C.-Y.; Zhang, C.; Wang, C.; Wang, J.-J. Increased serum miR-7 is a promising biomarker for type 2 diabetes mellitus and its microvascular complications. Diabetes Res. Clin. Pract. 2017, 130, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fu, L.L.; Wen, X.; Liu, B.; Huang, J.; Wang, J.H.; Wei, Y.Q. Oncogenic and tumor suppressive roles of microRNAs in apoptosis and autophagy. Apoptosis 2014, 19, 1177–1189. [Google Scholar] [CrossRef]
- Huo, J.-L.; Feng, Q.; Pan, S.; Fu, W.-J.; Liu, Z.; Liu, Z. Diabetic cardiomyopathy: Early diagnostic biomarkers, pathogenetic mechanisms, and therapeutic interventions. Cell Death Discov. 2023, 9, 256. [Google Scholar] [CrossRef]
- Hu, X.; Yin, G.; Zhang, Y.; Zhu, L.; Huang, H.; Lv, K. Recent advances in the functional explorations of nuclear microRNAs. Front. Immunol. 2023, 14, 1097491. [Google Scholar] [CrossRef]
- Pordzik, J.; Pisarz, K.; De Rosa, S.; Jones, A.D.; Eyileten, C.; Indolfi, C.; Malek, L.; Postula, M. The Potential Role of Platelet-Related microRNAs in the Development of Cardiovascular Events in High-Risk Populations, Including Diabetic Patients: A Review. Front. Endocrinol. 2018, 9, 74. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.C.; Gupta, A. MicroRNAs: Potential biomarkers for diagnosis and prognosis of different cancers. Transl. Cancer Res. 2020, 9, 5798–5818. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar]
- Steiman-Shimony, A.; Shtrikman, O.; Margalit, H. Assessing the functional association of intronic miRNAs with their host genes. RNA 2018, 24, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Talukder, A.; Cha, M.; Li, X.; Hu, H. Computational annotation of miRNA transcription start sites. Brief. Bioinform. 2021, 22, 380–392. [Google Scholar] [CrossRef]
- Eyileten, C.; Wicik, Z.; De Rosa, S.; Mirowska-Guzel, D.; Soplinska, A.; Indolfi, C.; Jastrzebska-Kurkowska, I.; Czlonkowska, A.; Postula, M. MicroRNAs as Diagnostic and Prognostic Biomarkers in Ischemic Stroke—A Comprehensive Review and Bioinformatic Analysis. Cells 2018, 7, 249. [Google Scholar] [CrossRef]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Pordzik, J.; Jakubik, D.; Jarosz-Popek, J.; Wicik, Z.; Eyileten, C.; De Rosa, S.; Indolfi, C.; Siller-Matula, J.M.; Czajka, P.; Postula, M. Significance of circulating microRNAs in diabetes mellitus type 2 and platelet reactivity: Bioinformatic analysis and review. Cardiovasc. Diabetol. 2019, 18, 113. [Google Scholar] [CrossRef]
- Sekar, D.; Venugopal, B.; Sekar, P.; Ramalingam, K. Role of microRNA 21 in diabetes and associated/related diseases. Gene 2016, 582, 14–18. [Google Scholar] [CrossRef]
- Agbu, P.; Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux. J. Clin. Investig. 2016, 126, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xing, W.; Xie, L. Regulatory Roles of MicroRNAs in Diabetes. Int. J. Mol. Sci. 2016, 17, 1729. [Google Scholar] [CrossRef] [PubMed]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Katsiki, N.; Behnam, B.; Iranpanah, H.; Sahebkar, A. MicroRNAs and type 2 diabetes mellitus: Molecular mechanisms and the effect of antidiabetic drug treatment. Metabolism 2018, 87, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, L. The small RNA miR-375—A pancreatic islet abundant miRNA with multiple roles in endocrine beta cell function. Mol. Cell. Endocrinol. 2017, 456, 95–101. [Google Scholar] [CrossRef]
- Lin, Y.C.; Huang, H.Y.; Shrestha, S.; Chou, C.H.; Chen, Y.H.; Chen, C.R.; Hong, H.C.; Li, J.; Chang, Y.A.; Chiew, M.Y.; et al. Multi-omics profiling reveals microRNA-mediated insulin signaling networks. BMC Bioinform. 2020, 21 (Suppl. 13), 389. [Google Scholar] [CrossRef]
- Macvanin, M.T.; Gluvic, Z.; Radovanovic, J.; Essack, M.; Gao, X.; Isenovic, E.R. Diabetic cardiomyopathy: The role of microRNAs and long non-coding RNAs. Front. Endocrinol. 2023, 14, 1124613. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sun, X.; Shan, P.F. MicroRNAs and Cardiovascular Disease in Diabetes Mellitus. Biomed. Res. Int. 2017, 2017, 4080364. [Google Scholar] [CrossRef]
- Jiang, Q.; Lyu, X.-M.; Yuan, Y.; Wang, L. Plasma miR-21 expression: An indicator for the severity of Type 2 diabetes with diabetic retinopathy. Biosci. Rep. 2017, 37, BSR20160589. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lu, C.; Lv, C.; Wu, C.; Wang, Q. The Expression of miR-192 and Its Significance in Diabetic Nephropathy Patients with Different Urine Albumin Creatinine Ratio. J. Diabetes Res. 2016, 2016, 6789402. [Google Scholar] [CrossRef] [PubMed]
- Pei, C.Z.; Liu, B.; Li, Y.T.; Fang, L.; Zhang, Y.; Li, Y.G.; Meng, S. MicroRNA-126 protects against vascular injury by promoting homing and maintaining stemness of late outgrowth endothelial progenitor cells. Stem Cell Res. Ther. 2020, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Maxwell, A.P.; Simpson, D.A.; McKay, G.J. Differential Expression of Urinary Exosomal MicroRNAs miR-21-5p and miR-30b-5p in Individuals with Diabetic Kidney Disease. Sci. Rep. 2019, 9, 10900. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, Y.; Li, G.; Lu, H. Identification of potential markers for type 2 diabetes mellitus via bioinformatics analysis. Mol. Med. Rep. 2020, 22, 1868–1882. [Google Scholar] [CrossRef] [PubMed]
- Poy, M.N.; Eliasson, L.; Krutzfeldt, J.; Kuwajima, S.; Ma, X.; Macdonald, P.E.; Pfeffer, S.; Tuschl, T.; Rajewsky, N.; Rorsman, P.; et al. A pancreatic islet-specific microRNA regulates insulin secretion. Nature 2004, 432, 226–230. [Google Scholar] [CrossRef]
- Li, X. miR-375, a microRNA related to diabetes. Gene 2014, 533, 1–4. [Google Scholar] [CrossRef]
- Dumortier, O.; Fabris, G.; Pisani, D.F.; Casamento, V.; Gautier, N.; Hinault, C.; Lebrun, P.; Duranton, C.; Tauc, M.; Dalle, S.; et al. microRNA-375 regulates glucose metabolism-related signaling for insulin secretion. J. Endocrinol. 2020, 244, 189–200. [Google Scholar] [CrossRef]
- Zhu, H.; Leung, S.W. Identification of microRNA biomarkers in type 2 diabetes: A meta-analysis of controlled profiling studies. Diabetologia 2015, 58, 900–911. [Google Scholar] [CrossRef]
- Drokow, E.K.; Sun, K.; Ahmed, H.A.W.; Akpabla, G.S.; Song, J.; Shi, M. Circulating microRNA as diagnostic biomarkers for haematological cancers: A systematic review and meta-analysis. Cancer Manag. Res. 2019, 11, 4313–4326. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Dhawan, S.; Hoang, J.; Cory, M.; Zeng, K.; Fritsch, H.; Meier, J.J.; Rizza, R.A.; Butler, P.C. β-Cell Deficit in Obese Type 2 Diabetes, a Minor Role of β-Cell Dedifferentiation and Degranulation. J. Clin. Endocrinol. Metab. 2016, 101, 523–532. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, Q.; Xing, X.; Yuan, T.; Li, P. Clinical research progress on β-cell dysfunction in T2DM development in the Chinese population. Rev. Endocr. Metab. Disord. 2024. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, G.M.; Azoz, N.M.A.; El Zohne, R.A.; Abdellatif, H.; Saleem, T.H.; Emam, W.A.; Mohammed, A.R.; Mohamed, S.A.; Muhammed, A.A.; Abd el-Rady, N.M.; et al. Dysregulated miRNA-375, IL-17, TGF-β, and Microminerals Are Associated with Calpain-10 SNP 19 in Diabetic Patients: Correlation with Diabetic Nephropathy Stages. Int. J. Mol. Sci. 2023, 24, 17446. [Google Scholar] [CrossRef] [PubMed]
- Sangali, P.; Abdullahi, S.; Nosrati, M.; Khosravi-Asrami, O.F.; Mahrooz, A.; Bagheri, A. Altered expression of miR-375 and miR-541 in type 2 diabetes patients with and without coronary artery disease (CAD): The potential of miR-375 as a CAD biomarker. J. Diabetes Metab. Disord. 2024, 23, 1101–1106. [Google Scholar] [CrossRef]
- Chang, X.; Li, S.; Li, J.; Yin, L.; Zhou, T.; Zhang, C.; Chen, X.; Sun, K. Ethnic differences in microRNA-375 expression level and DNA methylation status in type 2 diabetes of Han and Kazak populations. J. Diabetes Res. 2014, 2014, 761938. [Google Scholar] [CrossRef]
- Nathan, G.; Kredo-Russo, S.; Geiger, T.; Lenz, A.; Kaspi, H.; Hornstein, E.; Efrat, S. MiR-375 Promotes Redifferentiation of Adult Human β Cells Expanded In Vitro. PLoS ONE 2015, 10, e0122108. [Google Scholar] [CrossRef]
- Liao, L.; Tang, Y.; Zhou, Y.; Meng, X.; Li, B.; Zhang, X. MicroRNA-126 (MiR-126): Key roles in related diseases. J. Physiol. Biochem. 2024, 80, 277–286. [Google Scholar] [CrossRef]
- Ye, P.; Liu, J.; He, F.; Xu, W.; Yao, K. Hypoxia-induced deregulation of miR-126 and its regulative effect on VEGF and MMP-9 expression. Int. J. Med. Sci. 2014, 11, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Nammian, P.; Razban, V.; Tabei Mohammad Bagher, S.; Asadi-Yousefabad, S.-L. MicroRNA-126: Dual Role in Angiogenesis Dependent Diseases. Curr. Pharm. Des. 2020, 26, 4883–4893. [Google Scholar] [CrossRef] [PubMed]
- Alique, M.; Bodega, G.; Giannarelli, C.; Carracedo, J.; Ramírez, R. MicroRNA-126 regulates Hypoxia-Inducible Factor-1α which inhibited migration, proliferation, and angiogenesis in replicative endothelial senescence. Sci. Rep. 2019, 9, 7381. [Google Scholar] [CrossRef]
- Monfared, Y.K.; Mirzaii-Dizgah, M.-R.; Khodabandehloo, E.; Sarookhani, M.R.; Hashemipour, S.; Mirzaii-Dizgah, I. Salivary microRNA-126 and 135a: A potentially non-invasive diagnostic biomarkers of type- 2 diabetes. J. Diabetes Metab. Disord. 2021, 20, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Rezk, N.A.; Sabbah, N.A.; Saad, M.S.S. Role of MicroRNA 126 in screening, diagnosis, and prognosis of diabetic patients in Egypt. IUBMB Life 2016, 68, 452–458. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, G.; Yang, C.; Zhou, K.; Shen, B.; Liang, H.; Jiang, X. The Role of Circulating MicroRNA-126 (miR-126): A Novel Biomarker for Screening Prediabetes and Newly Diagnosed Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2014, 15, 10567–10577. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.L.; An, M.X.; Liu, Y.L.; Xu, H.C.; Lu, Z.Q. MicroRNA-126: A promising novel biomarker in peripheral blood for diabetic retinopathy. Int. J. Ophthalmol. 2017, 10, 530–534. [Google Scholar]
- Fernández-de Frutos, M.; Galán-Chilet, I.; Goedeke, L.; Kim, B.; Pardo-Marqués, V.; Pérez-García, A.; Herrero, J.I.; Fernández-Hernando, C.; Kim, J.; Ramírez, C.M. MicroRNA 7 Impairs Insulin Signaling and Regulates Aβ Levels through Posttranscriptional Regulation of the Insulin Receptor Substrate 2, Insulin Receptor, Insulin-Degrading Enzyme, and Liver X Receptor Pathway. Mol. Cell. Biol. 2019, 39, e00170-19. [Google Scholar] [CrossRef]
- Horsham, J.L.; Ganda, C.; Kalinowski, F.C.; Brown, R.A.M.; Epis, M.R.; Leedman, P.J. MicroRNA-7: A miRNA with expanding roles in development and disease. Int. J. Biochem. Cell Biol. 2015, 69, 215–224. [Google Scholar] [CrossRef]
- Zhang, H.M.; Kuang, S.; Xiong, X.; Gao, T.; Liu, C.; Guo, A.Y. Transcription factor and microRNA co-regulatory loops: Important regulatory motifs in biological processes and diseases. Brief. Bioinform. 2015, 16, 45–58. [Google Scholar] [CrossRef]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci. Rep. 2015, 5, 12453. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Luo, J.; Su, T.; Chen, C.; Su, Y. miR-7a Targets Insulin Receptor Substrate-2 Gene and Suppresses Viability and Invasion of Cells in Diabetic Retinopathy Mice via PI3K-Akt-VEGF Pathway. Diabetes Metab. Syndr. Obes. 2021, 14, 719–728. [Google Scholar] [CrossRef]
- Cao, Y.L.; Liu, D.J.; Zhang, H.G. MiR-7 regulates the PI3K/AKT/VEGF pathway of retinal capillary endothelial cell and retinal pericytes in diabetic rat model through IRS-1 and inhibits cell proliferation. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4427–4430. [Google Scholar] [PubMed]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S. A Regulator of Metabolic Reprogramming: MicroRNA Let-7. Transl. Oncol. 2019, 12, 1005–1013. [Google Scholar] [CrossRef]
- Katayama, M.; Sjögren Rasmus, J.O.; Egan, B.; Krook, A. miRNA let-7 expression is regulated by glucose and TNF-α by a remote upstream promoter. Biochem. J. 2015, 472, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Fan, L.; Shan, A.; Wang, W.; Ning, G.; Cao, Y.; Jiang, X. Let7b-5p inhibits insulin secretion and decreases pancreatic β-cell mass in mice. Mol. Cell. Endocrinol. 2022, 540, 111506. [Google Scholar] [CrossRef]
- Zhou, Q.; Frost, R.J.A.; Anderson, C.; Zhao, F.; Ma, J.; Yu, B.; Wang, S. let-7 Contributes to Diabetic Retinopathy but Represses Pathological Ocular Angiogenesis. Mol. Cell. Biol. 2017, 37, e00001-17. [Google Scholar] [CrossRef] [PubMed]
- Angelescu, M.A.; Andronic, O.; Dima, S.O.; Popescu, I.; Meivar-Levy, I.; Ferber, S.; Lixandru, D. miRNAs as Biomarkers in Diabetes: Moving towards Precision Medicine. Int. J. Mol. Sci. 2022, 23, 12843. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; El-Mahdy, H.A.; Eldeib, M.G.; Doghish, A.S. miRNAs as cornerstones in diabetic microvascular complications. Mol. Genet. Metab. 2023, 138, 106978. [Google Scholar] [CrossRef] [PubMed]
- Adnan, H. MicroRNA Expression in Diabetic Complications [Internet]. 2025. Available online: https://BioRender.com/y62j230 (accessed on 11 January 2025).
- Wang, S.; Ding, L.; Ji, H.; Xu, Z.; Liu, Q.; Zheng, Y. The Role of p38 MAPK in the Development of Diabetic Cardiomyopathy. Int. J. Mol. Sci. 2016, 17, 1037. [Google Scholar] [CrossRef]
- Guo, R.; Nair, S. Role of microRNA in diabetic cardiomyopathy: From mechanism to intervention. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2070–2077. [Google Scholar] [CrossRef]
- Poznyak, A.; Grechko, A.V.; Poggio, P.; Myasoedova, V.A.; Alfieri, V.; Orekhov, A.N. The Diabetes Mellitus-Atherosclerosis Connection: The Role of Lipid and Glucose Metabolism and Chronic Inflammation. Int. J. Mol. Sci. 2020, 21, 1835. [Google Scholar] [CrossRef] [PubMed]
- Katakami, N. Mechanism of Development of Atherosclerosis and Cardiovascular Disease in Diabetes Mellitus. J. Atheroscler. Thromb. 2018, 25, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Krecke, K.N.; Bapat, A.S.; Yang, T.; Lopresti, M.W.; Mashek, D.G.; Kelekar, A. Phosphatase PHLPP2 regulates the cellular response to metabolic stress through AMPK. Cell Death Dis. 2021, 12, 904. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Lin, J.; Zhang, Y.; Kang, S.; Belkin, N.; Wara, A.K.; Icli, B.; Hamburg, N.M.; Li, D.; Feinberg, M.W. MicroRNA-181b Improves Glucose Homeostasis and Insulin Sensitivity by Regulating Endothelial Function in White Adipose Tissue. Circ. Res. 2016, 118, 810–821. [Google Scholar] [CrossRef]
- Simpson, K.; Wonnacott, A.; Fraser, D.J.; Bowen, T. MicroRNAs in Diabetic Nephropathy: From Biomarkers to Therapy. Curr. Diabetes Rep. 2016, 16, 35. [Google Scholar] [CrossRef]
- Gallagher, H.; Suckling, R.J. Diabetic nephropathy: Where are we on the journey from pathophysiology to treatment? Diabetes Obes. Metab. 2016, 18, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Gan, H.; Zhang, H.; Tang, W.; Sun, Y.; Tang, X.; Kong, D.; Zhou, J.; Wang, Y.; Zhu, Y. MicroRNA-21 inhibits SMAD7 expression through a target sequence in the 3′ untranslated region and inhibits proliferation of renal tubular epithelial cells. Mol. Med. Rep. 2014, 10, 707–712. [Google Scholar] [CrossRef]
- Dhas, Y.; Arshad, N.; Biswas, N.; Jones, L.D.; Ashili, S. MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies. Biomedicines 2023, 11, 2583. [Google Scholar] [CrossRef]
- Kato, M.; Natarajan, R. MicroRNAs in diabetic nephropathy: Functions, biomarkers, and therapeutic targets. Ann. N. Y. Acad. Sci. 2015, 1353, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Eissa, S.; Matboli, M.; Bekhet, M.M. Clinical verification of a novel urinary microRNA panal: 133b, -342 and -30 as biomarkers for diabetic nephropathy identified by bioinformatics analysis. Biomed. Pharmacother. 2016, 83, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Moon, S.; Lee, K.; Park Ie, B.; Lee Dae, H.; Nam, S. Urinary and Blood MicroRNA-126 and -770 are Potential Noninvasive Biomarker Candidates for Diabetic Nephropathy: A Meta-Analysis. Cell. Physiol. Biochem. 2018, 46, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Xu, G.T.; Zhang, J.F. Inflammation in diabetic retinopathy: Possible roles in pathogenesis and potential implications for therapy. Neural Regen. Res. 2023, 18, 976–982. [Google Scholar] [PubMed]
- Lima, V.C.; Cavalieri, G.C.; Lima, M.C.; Nazario, N.O.; Lima, G.C. Risk factors for diabetic retinopathy: A case-control study. Int. J. Retin. Vitr. 2016, 2, 21. [Google Scholar] [CrossRef]
- Corcóstegui, B.; Durán, S.; González-Albarrán, M.O.; Hernández, C.; Ruiz-Moreno, J.M.; Salvador, J.; Udaondo, P.; Simó, R. Update on Diagnosis and Treatment of Diabetic Retinopathy: A Consensus Guideline of the Working Group of Ocular Health (Spanish Society of Diabetes and Spanish Vitreous and Retina Society). J. Ophthalmol. 2017, 2017, 8234186. [Google Scholar] [CrossRef]
- Rezazadeh-Gavgani, E.; Oladghaffari, M.; Bahramian, S.; Majidazar, R.; Dolati, S. MicroRNA-21: A critical underestimated molecule in diabetic retinopathy. Gene 2023, 859, 147212. [Google Scholar] [CrossRef] [PubMed]
- Woerner, J.; Sriram, V.; Nam, Y.; Verma, A.; Kim, D. Uncovering genetic associations in the human diseasome using an endophenotype-augmented disease network. Bioinformatics 2024, 40, btae126. [Google Scholar] [CrossRef]
- Alamro, H.; Bajic, V.; Macvanin, M.T.; Isenovic, E.R.; Gojobori, T.; Essack, M.; Gao, X. Type 2 Diabetes Mellitus and its comorbidity, Alzheimer’s disease: Identifying critical microRNA using machine learning. Front. Endocrinol. 2023, 13, 1084656. [Google Scholar] [CrossRef] [PubMed]
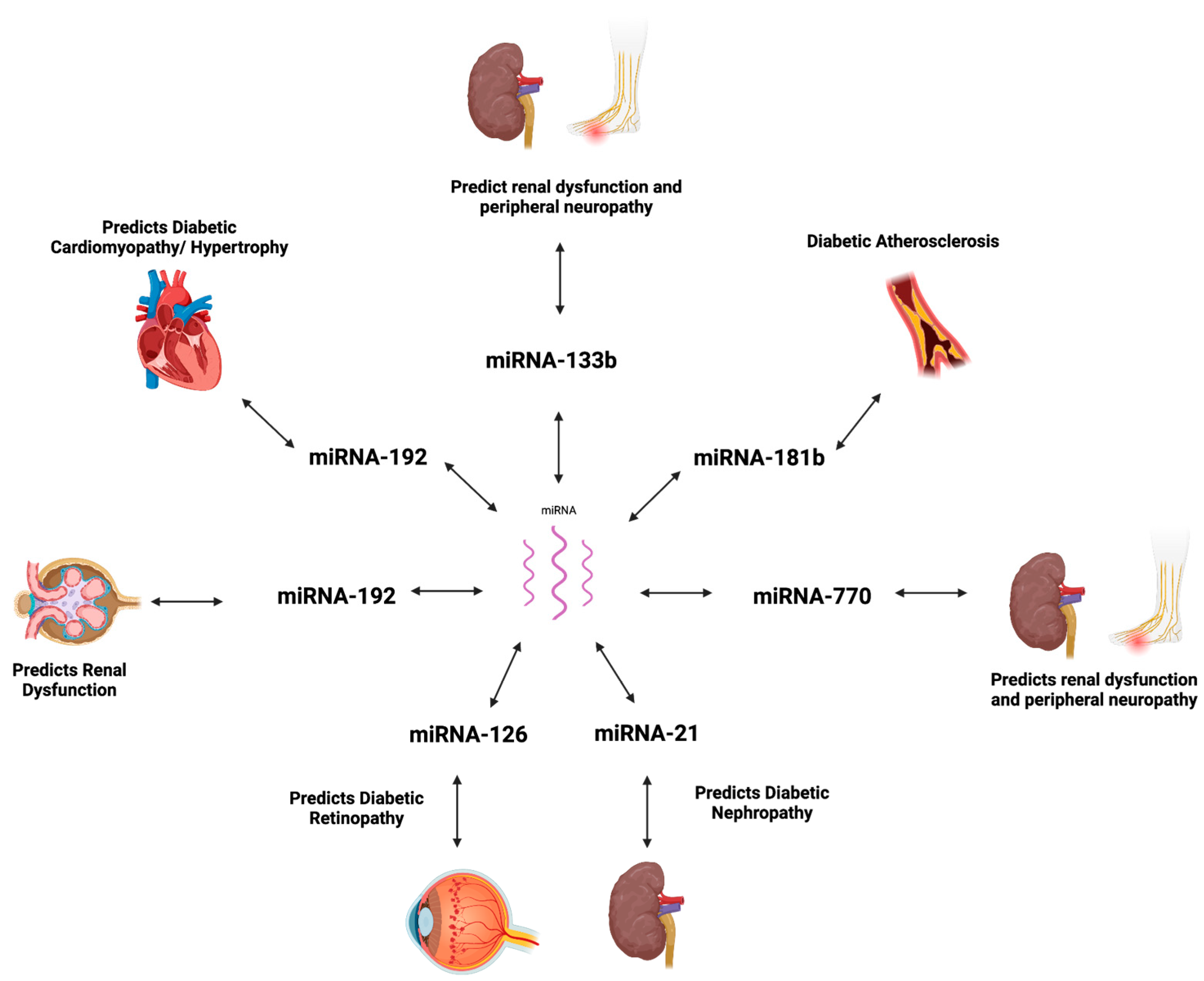
| miRNA | Expression | Clinical Value | Reference |
|---|---|---|---|
| miRNA-373 | Decreased expression is linked to hypertrophy mediated by MEF2C. | Diagnostic biomarker of diabetic cardiomyopathy, with dysfunction leading to hypertrophy of cardiomyocyte; early detection of decreased levels in myocardial cells can aid in timely diagnosis in high-risk groups. | [83] |
| miRNA-181b | Decreased expression is linked with increased macrophage accumulation and pathogenesis of diabetic atherosclerosis. | Decreased expression can be identified in white adipose tissue, allowing early detection to guide diagnosis and treatment. | [84] |
| miRNA-21 | Increased expression is linked to fibrosis and inflammation mediated by TGF-β and suppression of SMAD7. | Detecting upregulated levels in kidney/renal tubular epithelial cells can indicate early stages of diabetic nephropathy, aiding diagnosis, and can further be targeted for therapeutic purposes to decrease fibrosis and inflammation. | [85] |
| miRNA-192 | Decreased expression is linked to decreased urine albumin ratio. It is mediated by TGF-β and is associated with fibrosis. | Levels are decreased in early states of renal injury and can be identified in peripheral blood, allowing viability as an early diagnostic biomarker for diabetic nephropathy. | [86] |
| miRNA-133b | Dysregulated expression is linked with renal dysfunction and mediates fibrosis in renal tissue. | Dysregulated expression can be detected in the urine in micro or normal albuminuria states, with the potential for use as an early detection tool for diabetic nephropathy and further guiding treatment. | [87] |
| miRNA-342-3p | Upregulated/downregulated expression is linked with renal dysfunction and associated with injury to the renal tissue. | Altered levels can be identified in urine during early states of albuminuria to aid in the early detection of diabetic renal injury with potential progression to diabetic nephropathy. | [87] |
| miRNA-30b-5p | Dysregulated expression indicates renal dysfunction and may be used as a biomarker of DN. | Diagnostic urinary biomarker for diabetic nephropathy with potential for early detection of renal injury that can aid in monitoring response to treatment. | [88] |
| miRNA-126 | Decreased expression can serve as a biomarker for DR. | Decreased expression can be identified in serum of prediabetic and diabetic patients for early detection of potential progression to diabetic retinopathy and aid in monitoring response to treatment. | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.; Adnan, H.; Khawaja, M.A.; Butler, A.E. Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review. Int. J. Mol. Sci. 2025, 26, 753. https://doi.org/10.3390/ijms26020753
Ahmed S, Adnan H, Khawaja MA, Butler AE. Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review. International Journal of Molecular Sciences. 2025; 26(2):753. https://doi.org/10.3390/ijms26020753
Chicago/Turabian StyleAhmed, Sara, Haroon Adnan, Maryam A. Khawaja, and Alexandra E. Butler. 2025. "Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review" International Journal of Molecular Sciences 26, no. 2: 753. https://doi.org/10.3390/ijms26020753
APA StyleAhmed, S., Adnan, H., Khawaja, M. A., & Butler, A. E. (2025). Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review. International Journal of Molecular Sciences, 26(2), 753. https://doi.org/10.3390/ijms26020753