
Nintedanib Induces Mesenchymal-to-Epithelial Transition and Reduces Subretinal Fibrosis Through Metabolic Reprogramming

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
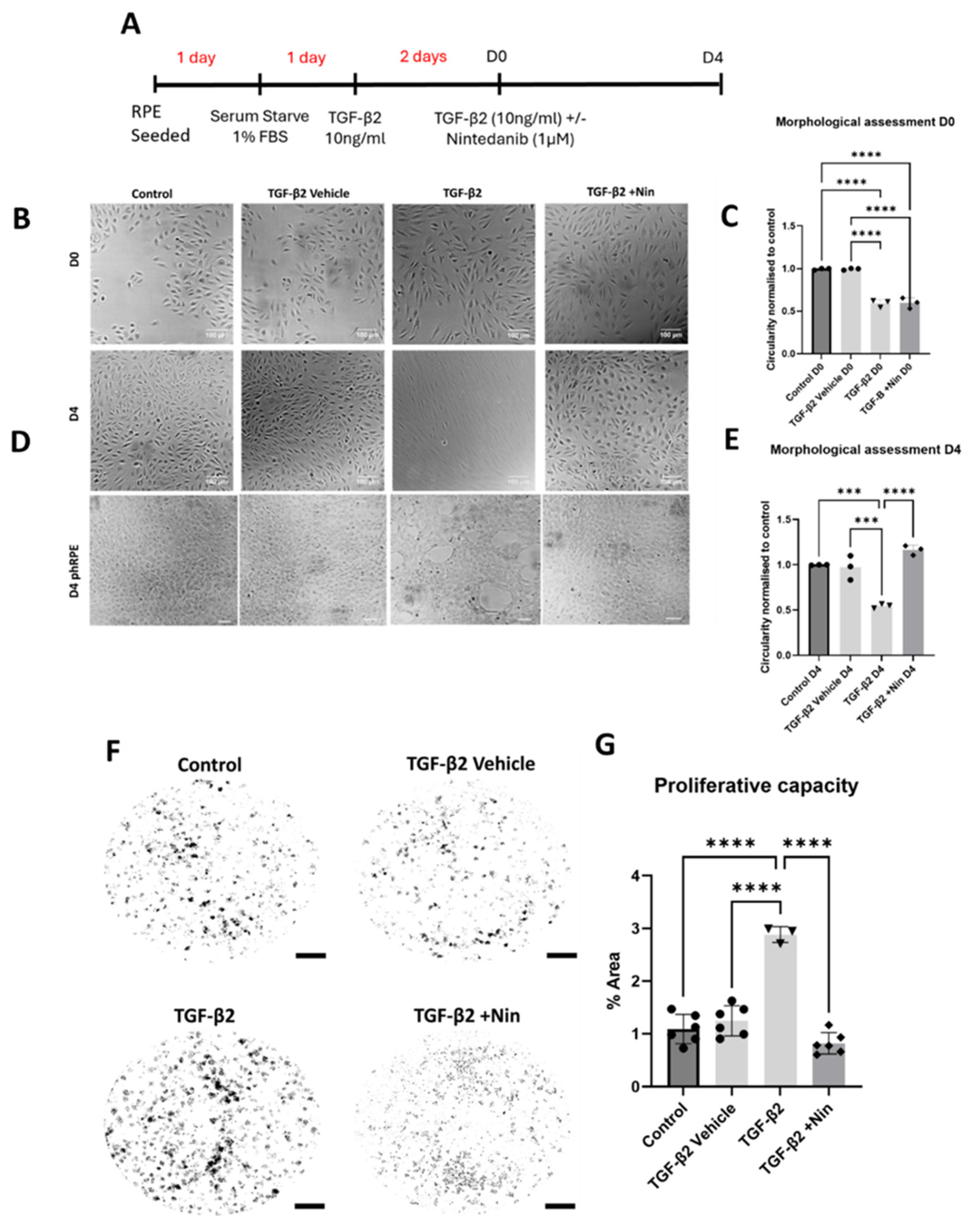
2.1. Nintedanib Reversed Transforming Growth Factor Beta 2 (TGF-β2) Induced Morphological Change and Reduced Proliferation in Retinal Pigment Epithelial (RPE) Cells
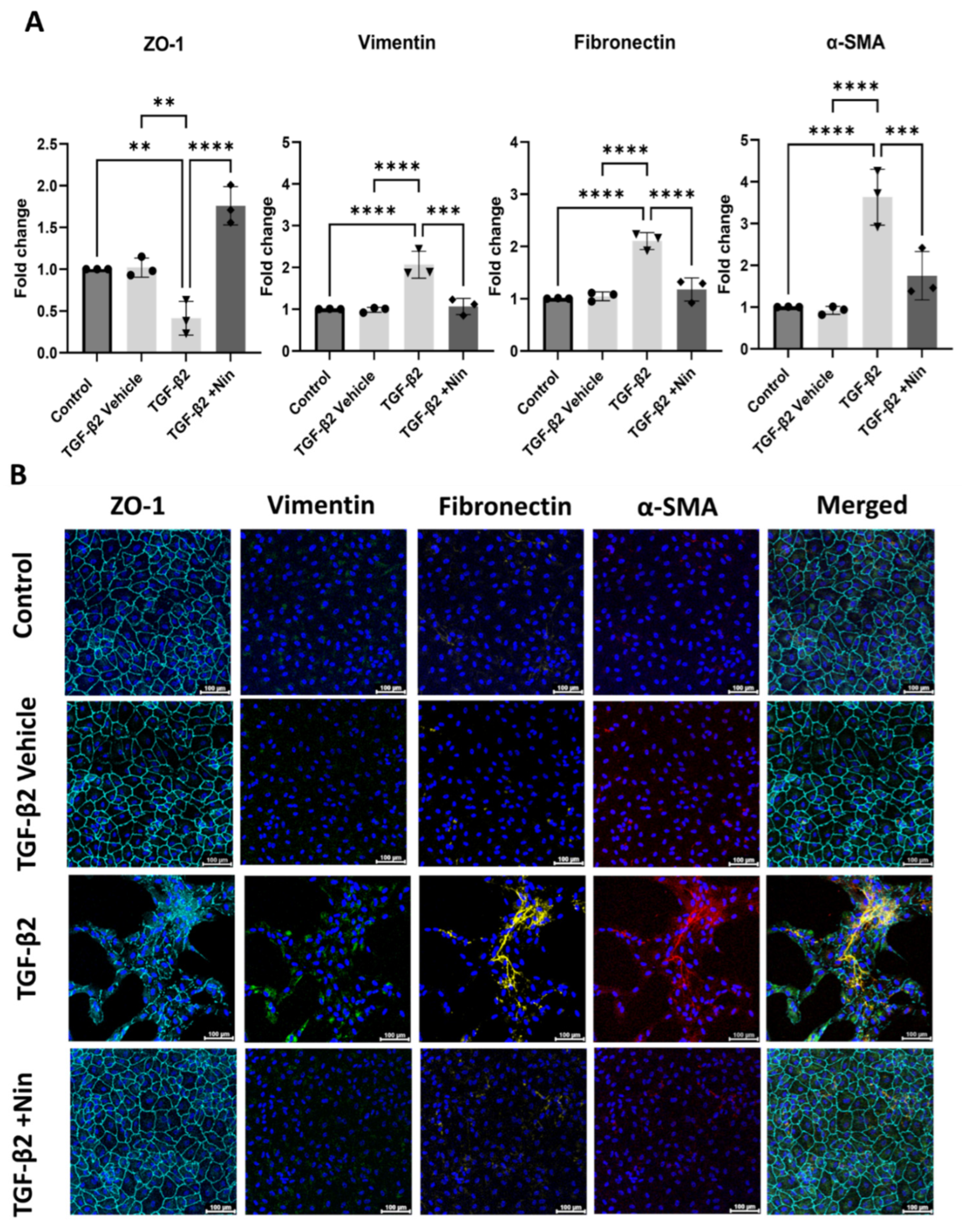
2.2. Nintedanib Restored Epithelial Markers and Reduced Epithelial Mesenchymal Transition (EMT) Markers in Transforming Growth Factor Beta 2 (TGF-β2) Treated Retinal Pigment Epithelial (RPE) Cells
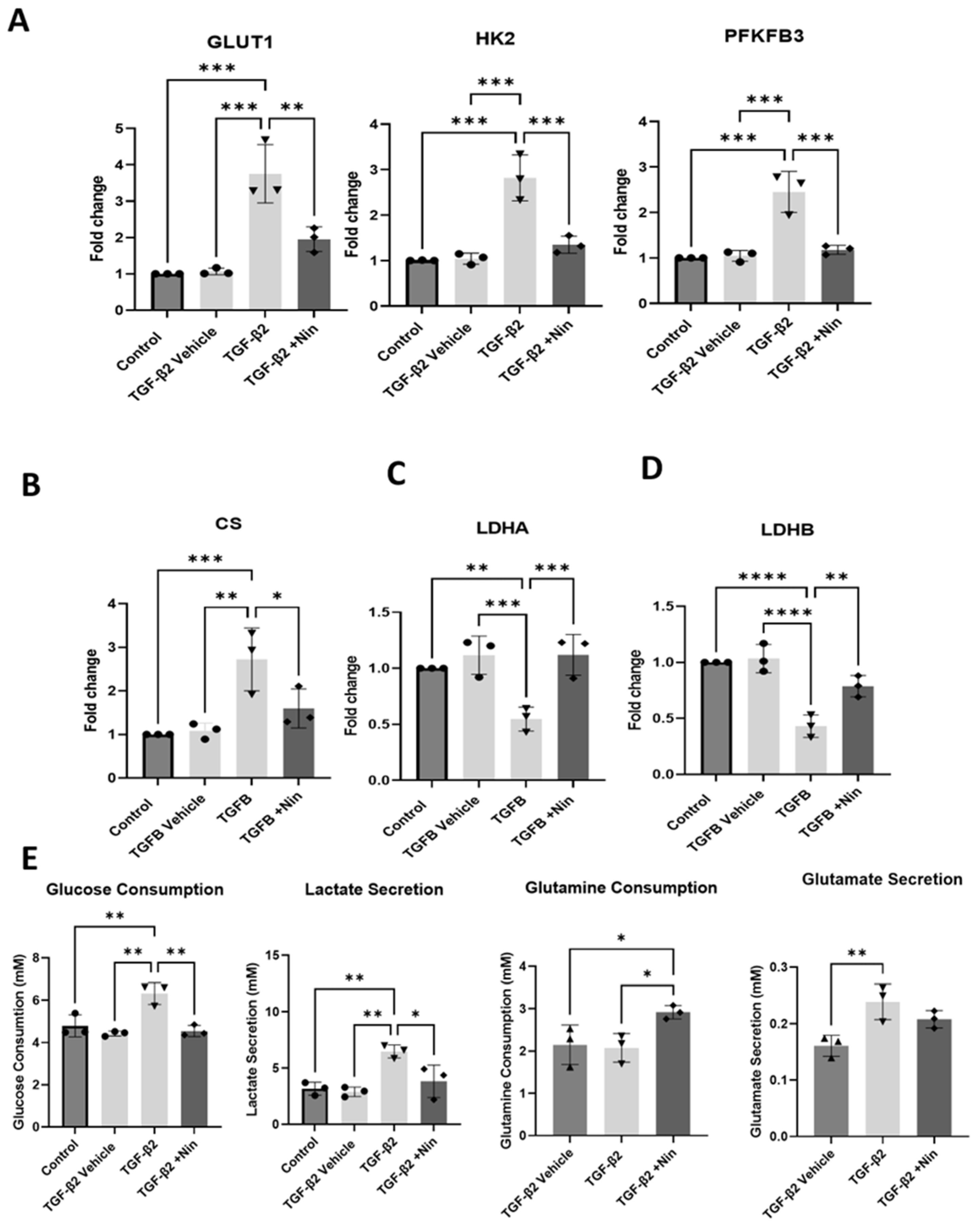
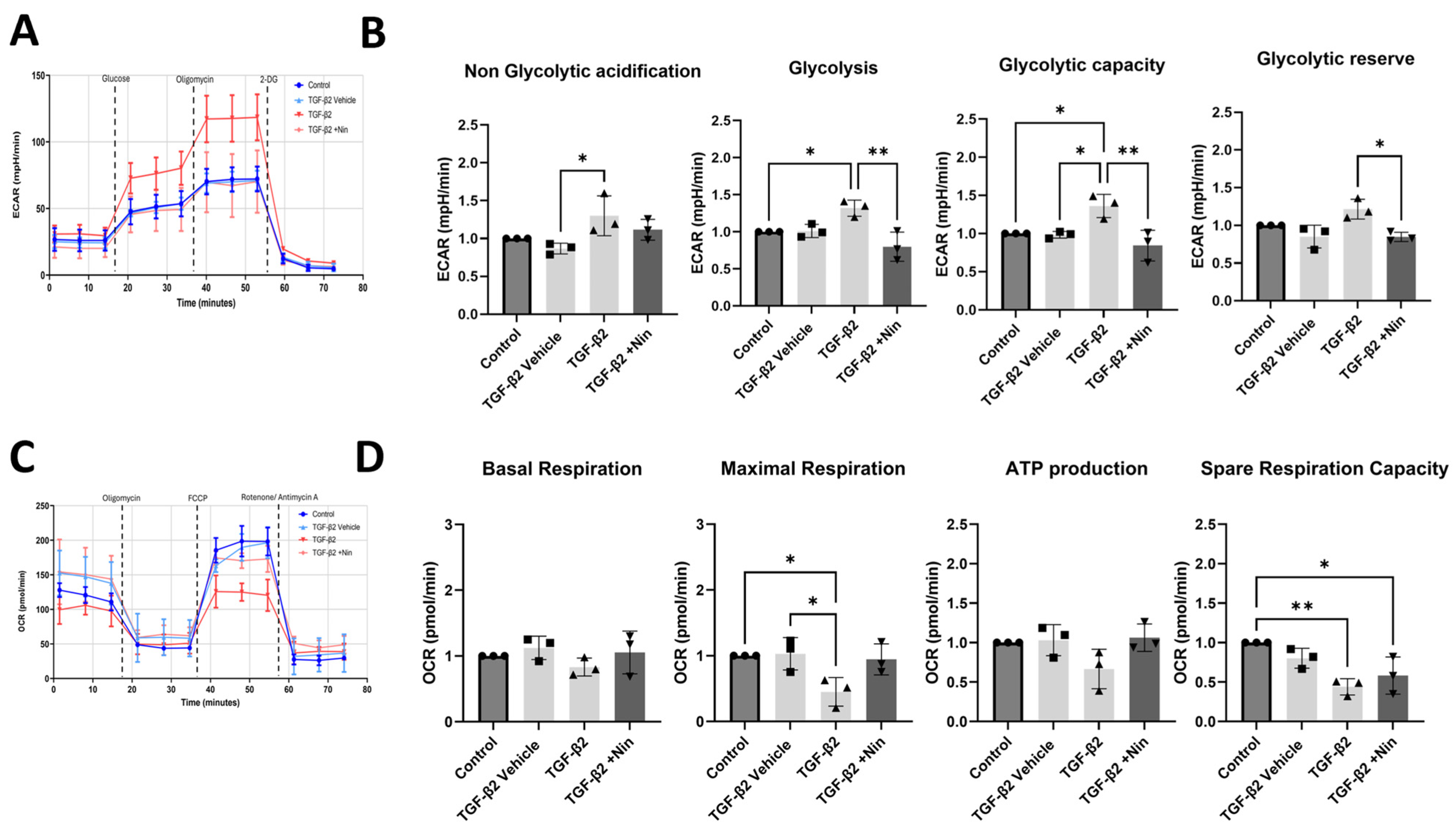
2.3. The Effect of Nintedanib on Metabolic Regulation in Transforming Growth Factor Beta 2 (TGF-β2) Treated Retinal Pigment Epithelial (RPE) Cells
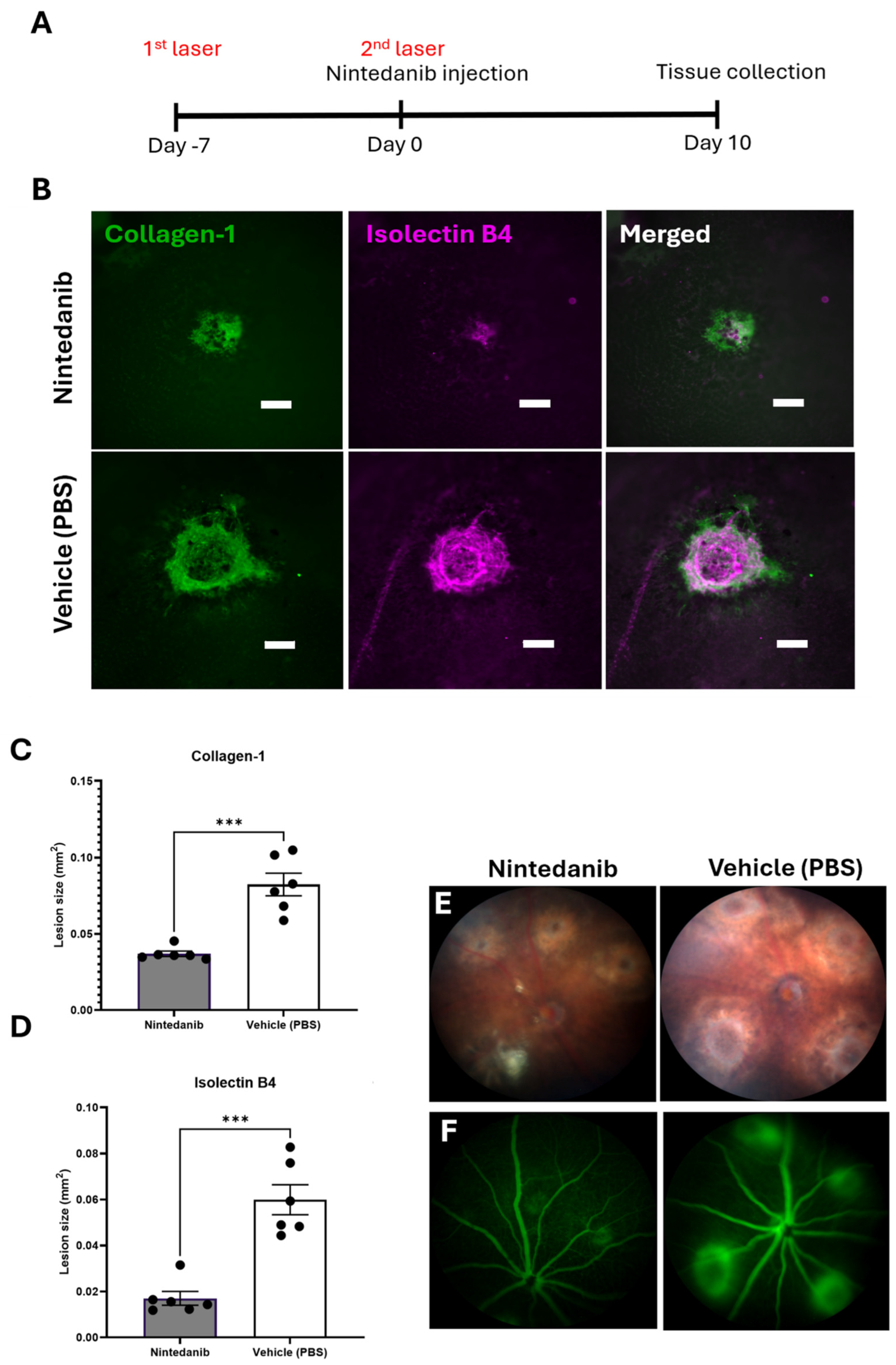
2.4. Nintedanib Reduces Fibrotic Lesions in the Mouse Model of Two-Stage Laser-Induced Subretinal Fibrosis
3. Discussion
4. Materials and Methods
4.1. Culture and Treatment of RPE Cells
4.2. Preparation of Nintedanib
4.3. Morphological Assessment
4.4. Clonogenic Assay
4.5. RNA Extraction
4.6. qPCR
4.7. Immunocytochemistry
4.8. Metabolic Assessments
4.9. Seahorse XFe96 Assay
4.10. Animals
4.11. Two-Stage Laser-Induced Subretinal Fibrosis
4.12. Intravitreal Injection of Nintedanib
4.13. Fundus Fluorescence Angiography
4.14. Immunohistochemistry
4.15. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ARPE-19 | Adult retinal pigment epithelial cell line |
| AMD | Age-related macular degeneration |
| ANOVA | Analysis of variance |
| CNV | Choroidal neovascularization |
| CTGF | Connective tissue growth factor |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle medium |
| ECAR | Extracellular acidification rate |
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| FBS | Fetal bovine serum |
| FGF | Fibroblast growth factor |
| FFA | Fundus fluorescein angiography |
| FGFR | Fibroblast growth factor receptor |
| FN | Fibronectin |
| GA | Geographic atrophy |
| GLUT1 | Glucose transporter 1 |
| HK2 | Hexokinase 2 |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IPF | Idiopathic pulmonary fibrosis |
| LDHA | lactate dehydrogenase A |
| LDHB | lactate dehydrogenase B |
| MET | Mesenchymal–epithelial transition |
| nAMD | Neovascular age related macular degeneration |
| OCR | Oxygen consumption rate |
| P | Passages |
| PBS | Phosphate-buffered saline |
| PCR | Polymerase chain reaction |
| PDGF | Platelet derived growth factor |
| phRPE | Primary human retinal pigment epithelial cells |
| RPE | Retinal pigment epithelial cells |
| RT | Room temperature |
| PDGFR | Platelet derived growth factor receptor |
| PVR | proliferative vitreoretinopathy |
| RTKs | Receptor tyrosine kinases |
| SSc-ILD | Systemic sclerosis-associated interstitial lung disease |
| TCA | Tricarboxylic acid |
| TKI | Tyrosine kinase inhibitor |
| VEGF | Vascular endothelial growth factor; |
| VEGFR | Vascular endothelial growth factor receptor |
| αSMA | alpha-smooth muscle actin |
| TNF-α | Tumor necrosis factor alpha |
| TGF-β | Transforming growth factor beta |
| PFKFB3 | 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 |
| FCCP | Carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone |
References
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-Related Macular Degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Ling Wong, W.; Su, X.; Li, X.; Ming Cheung, C.G.; Klein, R.; Cheng, C.-Y.; Yin Wong, T. Articles Global Prevalence of Age-Related Macular Degeneration and Disease Burden Projection for 2020 and 2040: A Systematic Review and Meta-Analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, C.; Zhang, J.; Xu, G.-T.; Zhang, J. Molecular Pathogenesis of Subretinal Fibrosis in Neovascular AMD Focusing on Epithelial-Mesenchymal Transformation of Retinal Pigment Epithelium. Neurobiol. Dis. 2023, 185, 106250. [Google Scholar] [CrossRef] [PubMed]
- Spaide, R.F.; Jaffe, G.J.; Sarraf, D.; Freund, K.B.; Sadda, S.R.; Staurenghi, G.; Waheed, N.K.; Chakravarthy, U.; Rosenfeld, P.J.; Holz, F.G.; et al. Consensus Nomenclature for Reporting Neovascular Age-Related Macular Degeneration Data Consensus on Neovascular Age-Related Macular Degeneration Nomenclature Study Group. Ophthalmology 2020, 127, 616–636. [Google Scholar] [CrossRef] [PubMed]
- Ambati, J.; Ambati, B.K.; Yoo, S.H.; Ianchulev, S.; Adamis, A.P. Age-Related Macular Degeneration: Etiology, Pathogenesis, and Therapeutic Strategies. Surv. Ophthalmol. 2003, 48, 257–293. [Google Scholar] [CrossRef] [PubMed]
- Bachmeier, I.; Armendariz, B.G.; Yu, S.; Jäger, R.J.; Ebneter, A.; Glittenberg, C.; Pauleikhoff, D.; Sadda, S.R.; Chakravarthy, U.; Fauser, S. Fibrosis in Neovascular Age-Related Macular Degeneration: A Review of Definitions Based on Clinical Imaging. Surv. Ophthalmol. 2023, 68, 835–843. [Google Scholar] [CrossRef]
- Little, K.; Ma, J.H.; Yang, N.; Chen, M.; Xu, H. Myofibroblasts in Macular Fibrosis Secondary to Neovascular Age-Related Macular Degeneration—The Potential Sources and Molecular Cues for Their Recruitment and Activation. eBioMedicine 2018, 38, 283–291. [Google Scholar] [CrossRef]
- Daniel, E.; Toth, C.A.; Grunwald, J.E.; Jaffe, G.J.; Martin, D.F.; Fine, S.L.; Huang, J.; Ying, G.-S.; Hagstrom, S.A.; Winter, K.; et al. Risk of Scar in the Comparison of Age-Related Macular Degeneration Treatments Trials. Ophthalmology 2014, 121, 656–666. [Google Scholar] [CrossRef]
- Little, K.; Llorián-Salvador, M.; Tang, M.; Du, X.; O’shaughnessy, Ó.; McIlwaine, G.; Chen, M.; Xu, H. A Two-Stage Laser-Induced Mouse Model of Subretinal Fibrosis Secondary to Choroidal Neovascularization. Transl. Vis. Sci. Technol. 2020, 9, 3. [Google Scholar] [CrossRef]
- Holz, F.G.; Schmitz-Valckenberg, S.; Fleckenstein, M. Recent Developments in the Treatment of Age-Related Macular Degeneration. J. Clin. Investig. 2014, 124, 1430–1438. [Google Scholar] [CrossRef]
- Gräfe, M.G.O.; van de Kreeke, J.A.; Willemse, J.; Braaf, B.; de Jong, Y.; Tan, H.S.; Verbraak, F.D.; de Boer, J.F. Subretinal Fibrosis Detection Using Polarization Sensitive Optical Coherence Tomography. Transl Vis. Sci. Technol. 2020, 9, 13. [Google Scholar] [CrossRef]
- Souied, E.H.; Addou-Regnard, M.; Ohayon, A.; Semoun, O.; Querques, G.; Blanco-Garavito, R.; Bunod, R.; Jung, C.; Sikorav, A.; Miere, A. Spectral-Domain Optical Coherence Tomography Analysis of Fibrotic Lesions in Neovascular Age-Related Macular Degeneration. Am. J. Ophthalmol. 2020, 214, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.L.; Ferris Iii, F.L.; Armstrong, J.; Hwang, T.S.; Chew, E.Y.; Bressler, S.B.; Chandra, S.R. Retinal Precursors and the Development of Geographic Atrophy in Age-Related Macular Degeneration. Ophthalmology 2008, 115, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Liu, J.; Deng, W.; Luo, C.; Qi, J.; Chen, M.; Xu, H. Macrophage Elastase (MMP12) Critically Contributes to the Development of Subretinal Fibrosis. J. Neuroinflamm. 2022, 19, 78. [Google Scholar] [CrossRef]
- Llorián-Salvador, M.; Byrne, E.M.; Szczepan, M.; Little, K.; Chen, M.; Xu, H. Complement Activation Contributes to Subretinal Fibrosis through the Induction of Epithelial-to-Mesenchymal Transition (EMT) in Retinal Pigment Epithelial Cells. J. Neuroinflamm. 2022, 19, 182. [Google Scholar] [CrossRef]
- Lechner, J.; Chen, M.; Hogg, R.E.; Toth, L.; Silvestri, G.; Chakravarthy, U.; Xu, H. Higher Plasma Levels of Complement C3a, C4a and C5a Increase the Risk of Subretinal Fibrosis in Neovascular Age-Related Macular Degeneration Complement Activation in AMD. Immun. Ageing 2016, 13, 4. [Google Scholar] [CrossRef]
- Chen, M.; Yang, N.; Lechner, J.; Toth, L.; Hogg, R.; Silvestri, G.; Chakravarthy, U.; Xu, H. Plasma Level of Lipocalin 2 Is Increased in Neovascular Age-Related Macular Degeneration Patients, Particularly Those with Macular Fibrosis. Immun. Ageing 2020, 17, 35. [Google Scholar] [CrossRef]
- Deng, W.; Yi, C.; Pan, W.; Liu, J.; Qi, J.; Chen, J.; Zhou, Z.; Duan, Y.; Ning, X.; Li, J.; et al. Vascular Cell Adhesion Molecule-1 (VCAM-1) Contributes to Macular Fibrosis in Neovascular Age-Related Macular Degeneration through Modulating Macrophage Functions. Immun. Ageing 2023, 20, 65. [Google Scholar] [CrossRef]
- Yi, C.; Liu, J.; Deng, W.; Luo, C.; Qi, J.; Chen, M.; Xu, H. Old Age Promotes Retinal Fibrosis in Choroidal Neovascularization through Circulating Fibrocytes and Profibrotic Macrophages. J. Neuroinflamm. 2023, 20, 45. [Google Scholar] [CrossRef]
- Ma, X.; Xie, Y.; Gong, Y.; Hu, C.; Qiu, K.; Yang, Y.; Shen, H.; Zhou, X.; Long, C.; Lin, X. Silibinin Prevents TGFβ-Induced EMT of RPE in Proliferative Vitreoretinopathy by Inhibiting Stat3 and Smad3 Phosphorylation. Investig. Ophthalmol. Vis. Science 2023, 64, 47. [Google Scholar] [CrossRef]
- Cunningham, F.; Van Bergen, T.; Canning, P.; Lengyel, I.; Feyen, J.H.M.; Stitt, A.W. The Placental Growth Factor Pathway and Its Potential Role in Macular Degenerative Disease. Curr. Eye Res. 2019, 44, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Rieg, A.D.; Anker, C.; Bünting, N.A.; Verjans, E.; Spillner, J.; Kalverkamp, S.; Von Stillfried, S.; Uhlig, S.; Martin, C. Platelet-Derived Growth Factor (PDGF)-BB Regulates the Airway Tone via Activation of MAP2K, Thromboxane, Actin Polymerisation and Ca 2+-Sensitisation. Respir. Res. 2020, 23, 189. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N. Nintedanib: A Review in Fibrotic Interstitial Lung Diseases. Drugs 2021, 81, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Beyer, C.; Palumbo-Zerr, K.; Zhang, Y.; Ramming, A.; Distler, A.; Gelse, K.; Distler, O.; Schett, G.; Wollin, L.; et al. Nintedanib Inhibits Fibroblast Activation and Ameliorates Fibrosis in Preclinical Models of Systemic Sclerosis. N. Engl. J. Med. 2014, 370, 22. [Google Scholar] [CrossRef]
- Hilberg, F.; Roth, G.J.; Krssak, M.; Kautschitsch, S.; Sommergruber, W.; Tontsch-Grunt, U.; Garin-Chesa, P.; Bader, G.; Zoephel, A.; Quant, J.; et al. BIBF 1120: Triple Angiokinase Inhibitor with Sustained Receptor Blockade and Good Antitumor Efficacy. Cancer Res. 2008, 68, 4774–4782. [Google Scholar] [CrossRef]
- Rangarajan, S.; Kurundkar, A.; Kurundkar, D.; Bernard, K.; Sanders, Y.Y.; Ding, Q.; Antony, V.B.; Zhang, J.; Zmijewski, J.; Thannickal, V.J. ORIGINAL RESEARCH Novel Mechanisms for the Antifibrotic Action of Nintedanib. Am. J. Respir. Cell Mol. Biol. 2016, 54, 51–59. [Google Scholar] [CrossRef]
- Richeldi, L.; Du Bois, R.M.; Raghu, G.; Azuma, A.; Brown, K.K.; Costabel, U.; Cottin, V.; Flaherty, K.R.; Hansell, D.M.; Inoue, Y.; et al. Efficacy and Safety of Nintedanib in Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2014, 22, 2071–2082. [Google Scholar] [CrossRef]
- Flaherty, K.R.; Wells, A.U.; Cottin, V.; Devaraj, A.; Walsh, S.L.F.; Inoue, Y.; Richeldi, L.; Kolb, M.; Tetzlaff, K.; Stowasser, S.; et al. Nintedanib in Progressive Fibrosing Interstitial Lung Diseases. N. Engl. J. Med. 2019, 381, 1718–1727. [Google Scholar] [CrossRef]
- Li, L.F.; Kao, K.C.; Liu, Y.Y.; Lin, C.W.; Chen, N.H.; Lee, C.S.; Wang, C.W.; Yang, C.T. Nintedanib Reduces Ventilation-augmented Bleomycin-induced Epithelial–Mesenchymal Transition and Lung Fibrosis through Suppression of the Src Pathway. J. Cell Mol. Med. 2017, 21, 2937–2949. [Google Scholar] [CrossRef]
- Ihara, H.; Mitsuishi, Y.; Kato, M.; Takahashi, F.; Tajima, K.; Hayashi, T.; Hidayat, M.; Winardi, W.; Wirawan, A.; Hayakawa, D.; et al. Nintedanib Inhibits Epithelial-Mesenchymal Transition in A549 Alveolar Epithelial Cells through Regulation of the TGF-b/Smad Pathway. Respir. Investig. 2020, 58, 275–284. [Google Scholar] [CrossRef]
- Nishijima, N.; Seike, M.; Soeno, C.; Chiba, M.; Miyanaga, A.; Noro, R.; Sugano, T.; Matsumoto, M.; Kubota, K.; Gemma, A. MiR-200/ZEB Axis Regulates Sensitivity to Nintedanib in Non-Small Cell Lung Cancer Cells. Int. J. Oncol. 2016, 48, 937–944. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, S.; Pu, L.; Luo, J.; Liu, H.; Wu, W. Nintedanib Prevents TGF-Β2-Induced Epithelial-Mesenchymal Transition in Retinal Pigment Epithelial Cells. Biomed. Pharmacother. 2023, 161, 114543. [Google Scholar] [CrossRef] [PubMed]
- Hajari Case, A.; Johnson, P. Clinical Use of Nintedanib in Patients with Idiopathic Pulmonary Fibrosis. BMJ Open Respir. Res. 2017, 4, e000192. [Google Scholar] [CrossRef] [PubMed]
- Senabouth, A.; Daniszewski, M.; Lidgerwood, G.E.; Liang, H.H.; Hernández, D.; Mirzaei, M.; Keenan, S.N.; Zhang, R.; Han, X.; Neavin, D.; et al. Transcriptomic and Proteomic Retinal Pigment Epithelium Signatures of Age-Related Macular Degeneration. Nat. Commun. 2022, 13, 4233. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Kim, H.; Lee, S.; Youn, H.; Youn, B. Role of Metabolic Reprogramming in Epithelial–Mesenchymal Transition (EMT). Int. J. Mol. Sci. 2019, 20, 2042. [Google Scholar] [CrossRef]
- Schwager, S.C.; Mosier, J.A.; Padmanabhan, R.S.; White, A.; Xing, Q.; Hapach, L.A.; Taufalele, P.V.; Ortiz, I.; Reinhart-King, C.A. Link between Glucose Metabolism and Epithelial-to-Mesenchymal Transition Drives Triple-Negative Breast Cancer Migratory Heterogeneity. iScience 2022, 25, 105190. [Google Scholar] [CrossRef]
- Shu, D.Y.; Butcher, E.R.; Saint-Geniez, M.; John, H.; Paulson, A. Molecular Sciences Suppression of PGC-1α Drives Metabolic Dysfunction in TGFβ2-Induced EMT of Retinal Pigment Epithelial Cells. Int. J. Mol. Sci 2021, 22, 4701. [Google Scholar] [CrossRef]
- Martusewicz-Boros, M.; Górska, K. Nintedanib—Efficacy, Safety and Practical Aspects of Treatment for Patients with Idiopathic Pulmonary Fibrosis. Adv. Respir. Med. 2020, 88, 599–607. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinants/Molecules | |||
| Cat. No. | Company | ||
| Recombinant Mouse Transforming Growth Factor Beta 2 (TGF-β2) | 7346-B2-005 | R&D Systems, Abingdon, Oxfordshire, UK | |
| Primary Antibodies | |||
| Cat. No. | Company | Host | |
| CoraLite® Plus 488-conjugated ZO-1 Polyclonal antibody | CL488-21773 | Proteintech, Manchester, UK | rabbit |
| CoraLite®555-conjugated Vimentin Recombinant antibody | CL555-80232 | Proteintech, Manchester, UK | rabbit |
| CoraLite® Plus 647-conjugated smooth muscle actin specific Monoclonal antibody | CL647-67735 | Proteintech, Manchester, UK | rabbit |
| CoraLite®594-conjugated Fibronectin Polyclonal antibody | CL594-15613 | Proteintech, Manchester, UK | rabbit |
| Anti-Collagen Type I Antibody | AB758 | Sigma-Aldrich, Gillingham, Dorest, UK | Goat |
| Griffonia Simplicifolia Lectin I (GSL I) Isolectin B4, Biotinylated | B-1205-5 | Vector Laboratories, Newark, CA, USA | |
| Secondary Antibodies | ||
|---|---|---|
| Cat. No. | Company | |
| Alexa Fluor® 488 AffiniPure™ Donkey Anti-Goat IgG (H+L) | 705-545-147 | Jackson ImmunoResearch, Ely, Cambridgeshire, UK |
| Alexa Fluor® 594 Streptavidin | 016-580-084 | Jackson ImmunoResearch, Ely, Cambridgeshire, UK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hughes, D.; Prestle, J.; Zippel, N.; McFetridge, S.; Szczepan, M.; Neubauer, H.; Xu, H.; Chen, M. Nintedanib Induces Mesenchymal-to-Epithelial Transition and Reduces Subretinal Fibrosis Through Metabolic Reprogramming. Int. J. Mol. Sci. 2025, 26, 7131. https://doi.org/10.3390/ijms26157131
Hughes D, Prestle J, Zippel N, McFetridge S, Szczepan M, Neubauer H, Xu H, Chen M. Nintedanib Induces Mesenchymal-to-Epithelial Transition and Reduces Subretinal Fibrosis Through Metabolic Reprogramming. International Journal of Molecular Sciences. 2025; 26(15):7131. https://doi.org/10.3390/ijms26157131
Chicago/Turabian StyleHughes, David, Jüergen Prestle, Nina Zippel, Sarah McFetridge, Manon Szczepan, Heike Neubauer, Heping Xu, and Mei Chen. 2025. "Nintedanib Induces Mesenchymal-to-Epithelial Transition and Reduces Subretinal Fibrosis Through Metabolic Reprogramming" International Journal of Molecular Sciences 26, no. 15: 7131. https://doi.org/10.3390/ijms26157131
APA StyleHughes, D., Prestle, J., Zippel, N., McFetridge, S., Szczepan, M., Neubauer, H., Xu, H., & Chen, M. (2025). Nintedanib Induces Mesenchymal-to-Epithelial Transition and Reduces Subretinal Fibrosis Through Metabolic Reprogramming. International Journal of Molecular Sciences, 26(15), 7131. https://doi.org/10.3390/ijms26157131