
Pharmacological Preconditioning with Diazoxide Upregulates HCN4 Channels in the Sinoatrial Node of Adult Rat Cardiomyocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
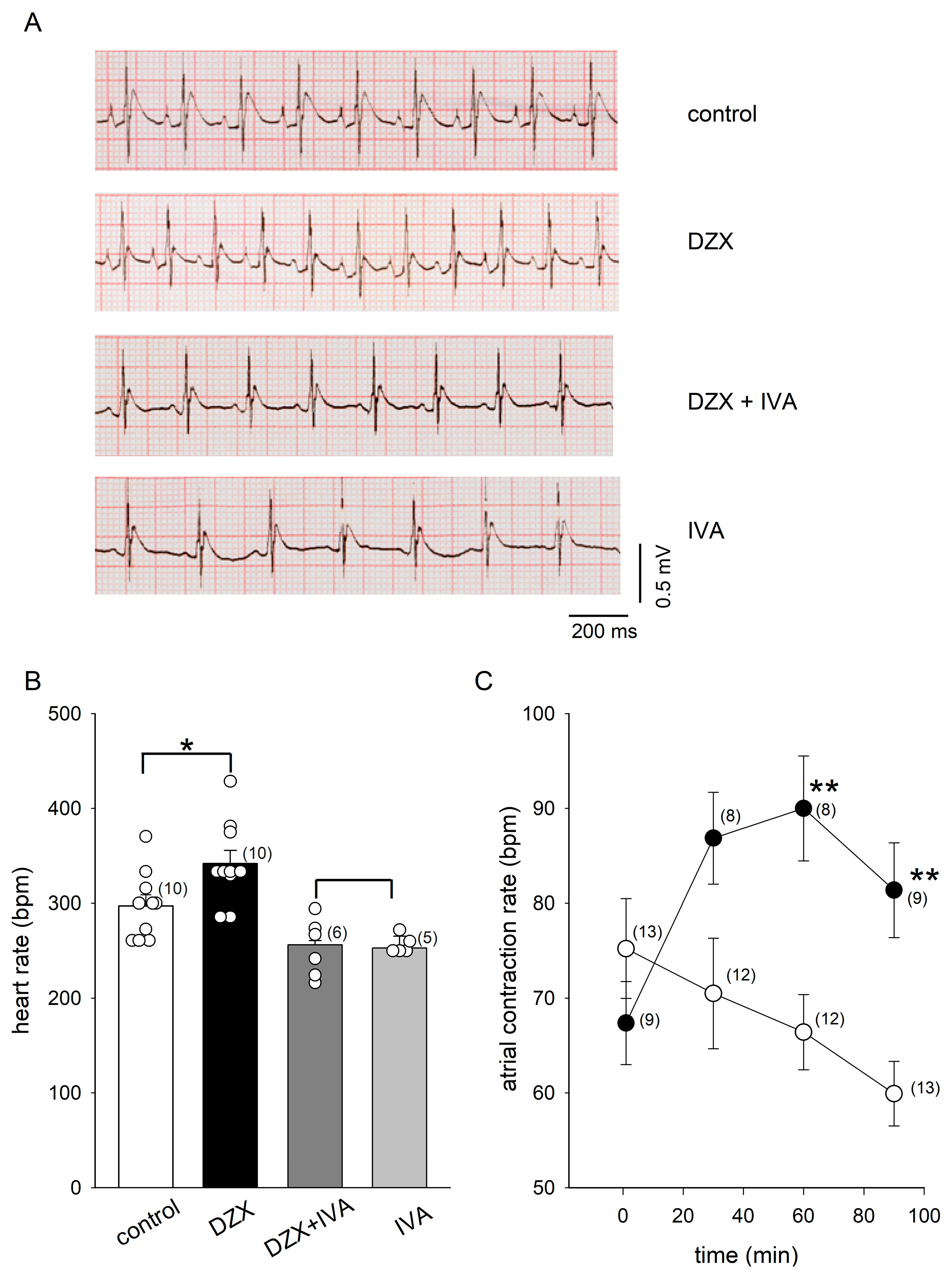
2.1. PPC Increased the Heart Rate and HCN4 Channel Currents
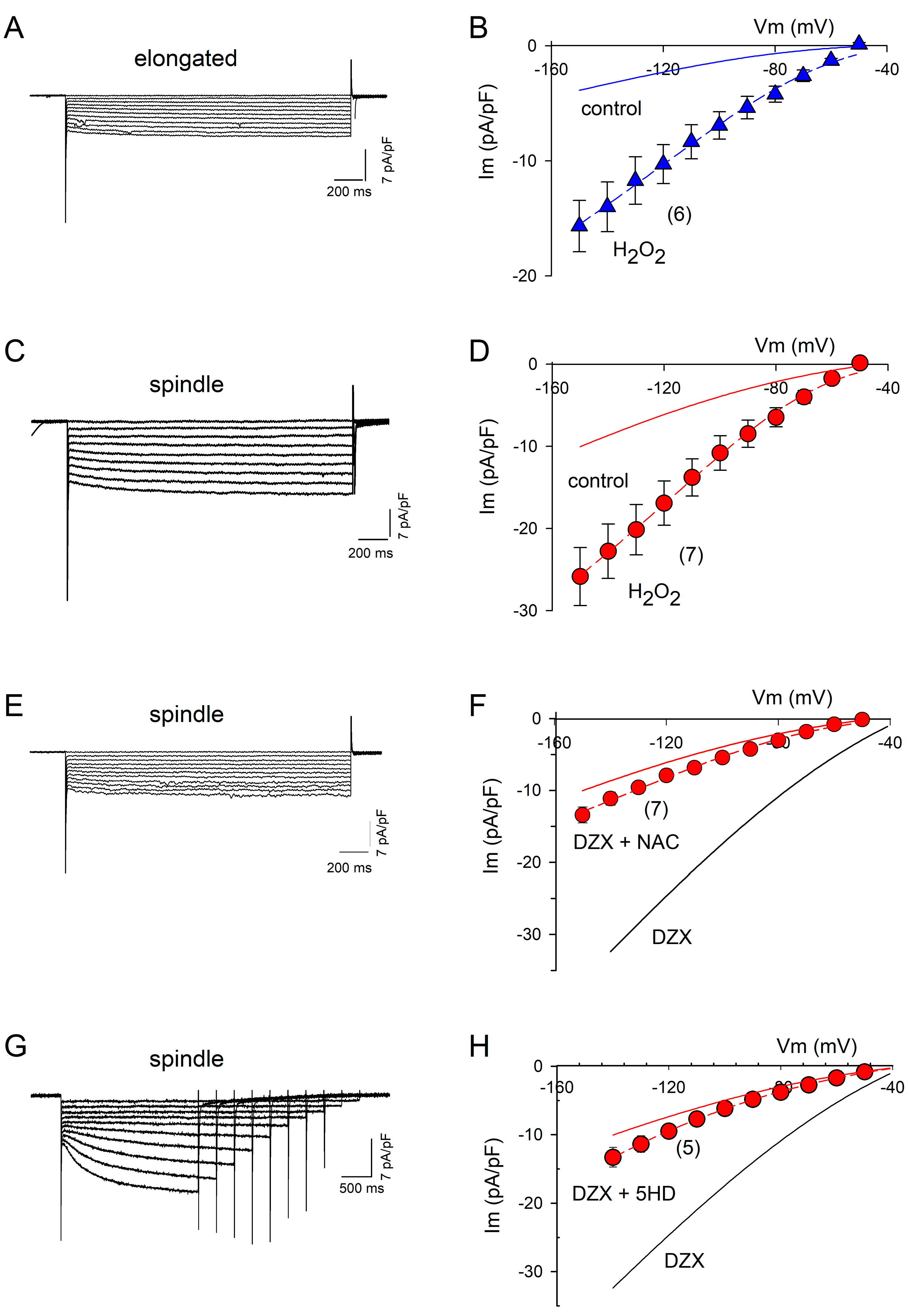
2.2. ROS Are Involved in PPC
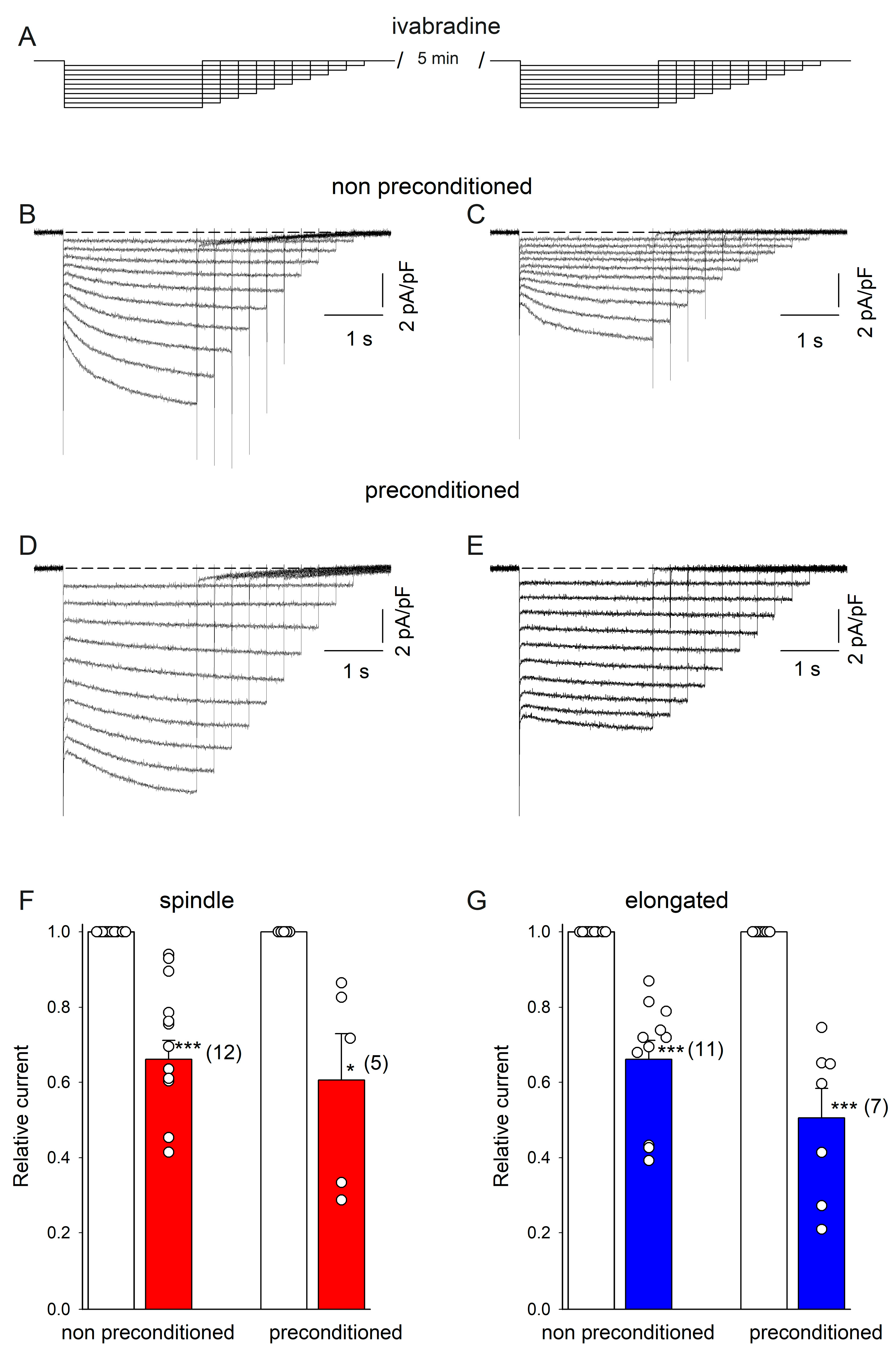
2.3. DZX Increases HCN4 Channel Conductance
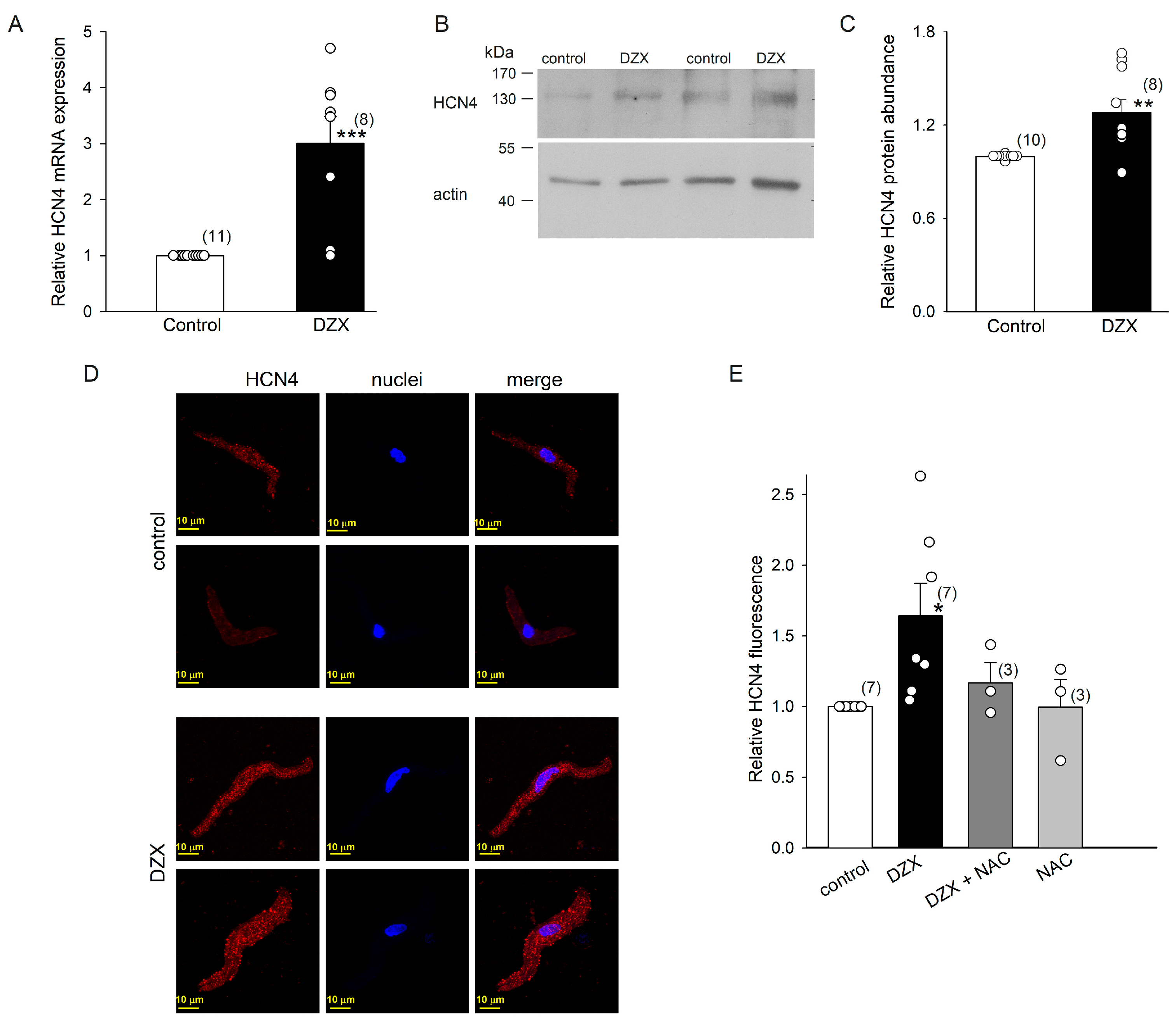
2.4. DZX Increased HCN4 Channel Expression
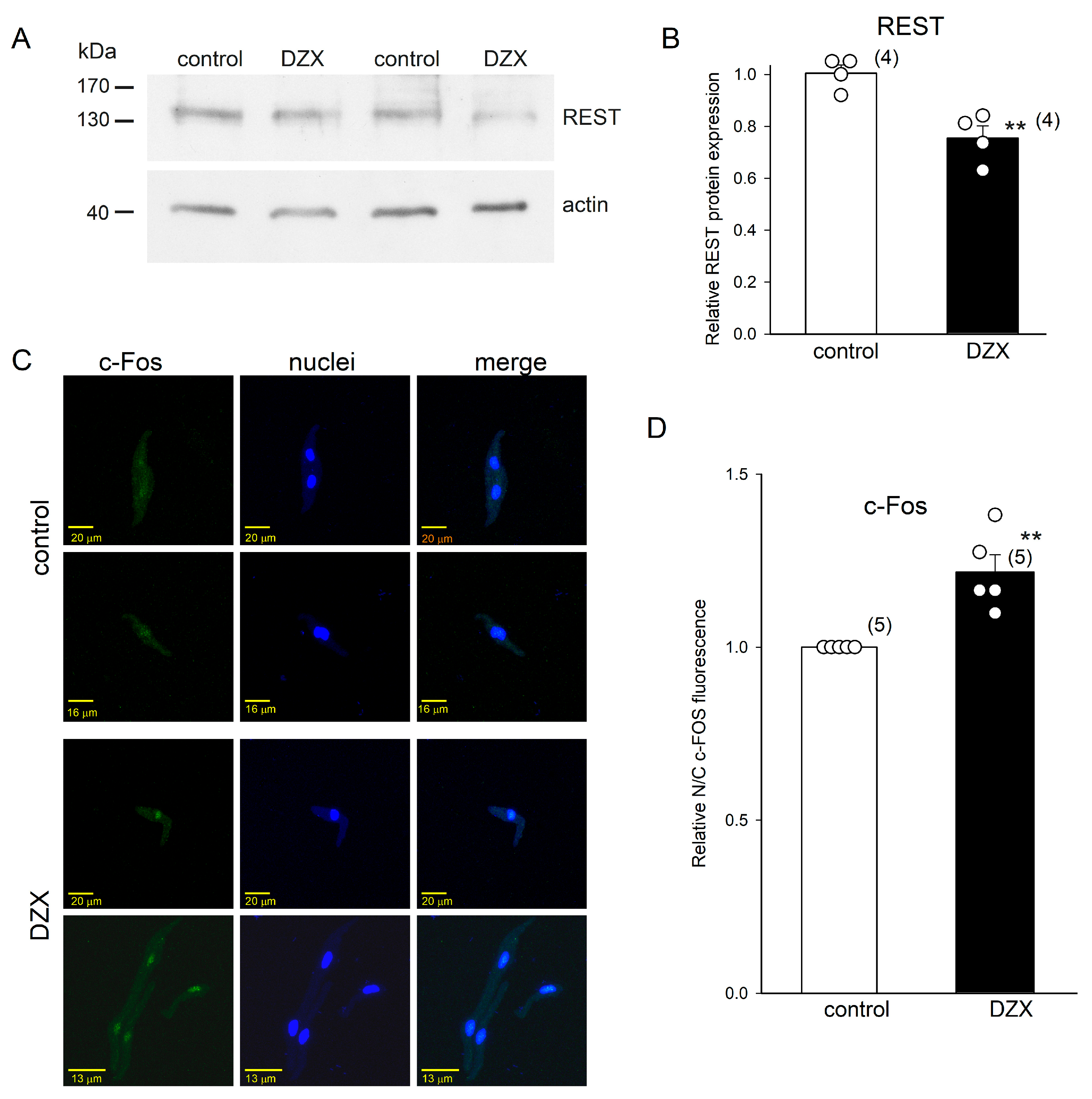
2.5. Effects of DZX on REST and c-Fos Transcription Factors
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. ECG and Atrial Contraction Rate Determination
4.3. SANC Isolation
4.4. Electrophysiological Analysis
4.5. Analysis of HCN4 Channel Currents
4.6. Measurement of Ca2+ Transients
4.7. qRT-PCR Assays
4.8. Immunofluorescence
4.9. Western Blotting
4.10. Statistical Analysis
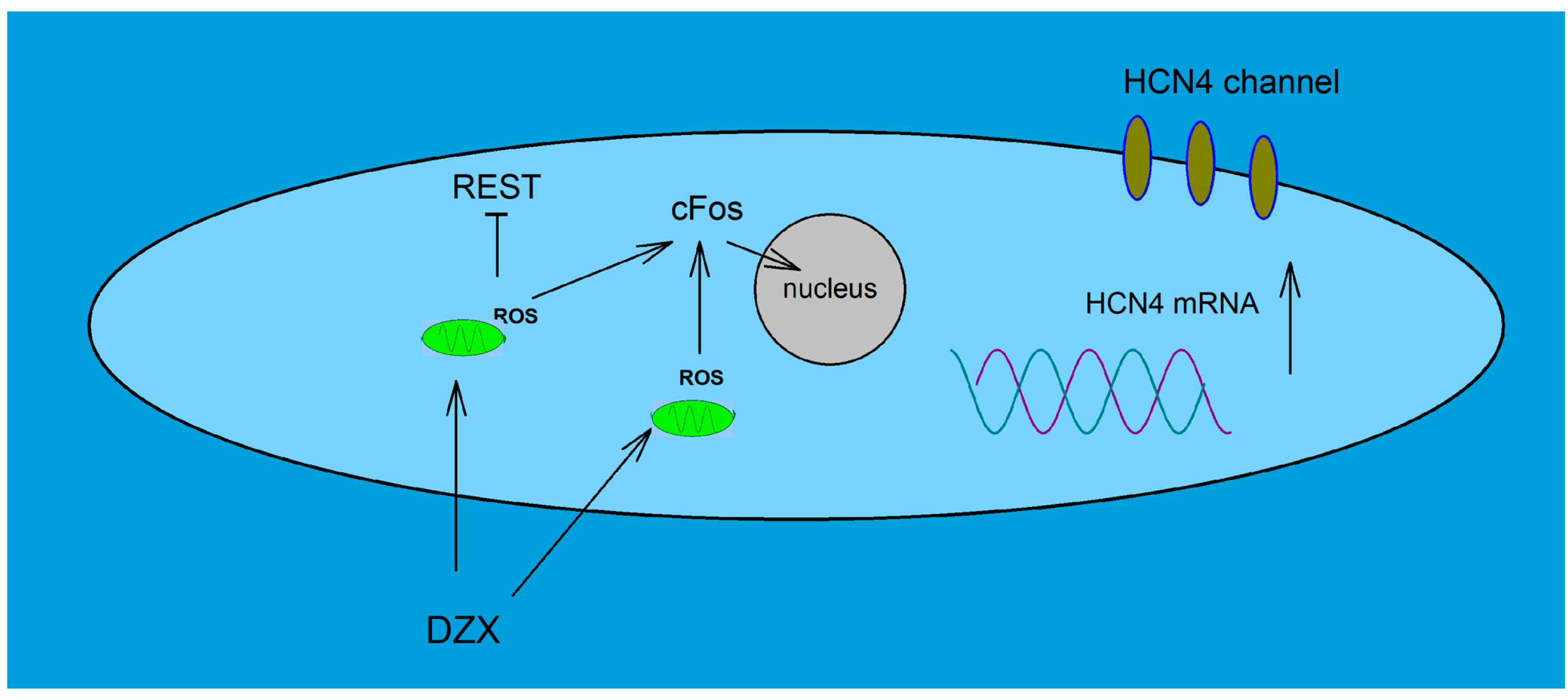
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DZX | Diazoxide |
| IVA | Ivabradine |
| HCN4 | Hyperpolarization-activated cyclic nucleotide potassium channel 4 |
| REST | RE1-silencing transcription factor |
| NRSF | Neuron-restrictive silencer factor |
References
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with Ischemia: A Delay of Lethal Cell Injury in Ischemic Myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef]
- Garlid, K.D.; Paucek, P.; Yarov-Yarovoy, V.; Murray, H.N.; Darbenzio, R.B.; D’Alonzo, A.J.; Lodge, N.J.; Smith, M.A.; Grover, G.J. Cardioprotective Effect of Diazoxide and Its Interaction with Mitochondrial ATP-Sensitive K+ Channels. Possible Mechanism of Cardioprotection. Circ. Res. 1997, 81, 1072–1082. [Google Scholar] [CrossRef]
- Pain, T.; Yang, X.M.; Critz, S.D.; Yue, Y.; Nakano, A.; Liu, G.S.; Heusch, G.; Cohen, M.V.; Downey, J.M. Opening of Mitochondrial K(ATP) Channels Triggers the Preconditioned State by Generating Free Radicals. Circ. Res. 2000, 87, 460–466. [Google Scholar] [CrossRef]
- González, G.; Zaldívar, D.; Carrillo, E.; Hernández, A.; García, M.; Sánchez, J. Pharmacological Preconditioning by Diazoxide Downregulates Cardiac L-Type Ca2+ Channels. Br. J. Pharmacol. 2010, 161, 1172–1185. [Google Scholar] [CrossRef]
- Ardehali, H.; O’Rourke, B. Mitochondrial K(ATP) Channels in Cell Survival and Death. J. Mol. Cell Cardiol. 2005, 39, 7–16. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Clarke, S.J.; Khaliulin, I. The Role of Mitochondria in Protection of the Heart by Preconditioning. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1767, 1007–1031. [Google Scholar] [CrossRef]
- Lesnefsky, E.J.; Chen, Q.; Tandler, B.; Hoppel, C.L. Mitochondrial Dysfunction and Myocardial Ischemia-Reperfusion: Implications for Novel Therapies. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 535–565. [Google Scholar] [CrossRef]
- Sampieri, R.; Fuentes, E.; Carrillo, E.D.; Hernández, A.; García, M.C.; Sánchez, J.A. Pharmacological Preconditioning Using Diazoxide Regulates Store-Operated Ca2 + Channels in Adult Rat Cardiomyocytes. Front. Physiol. 2019, 10, 1589. [Google Scholar] [CrossRef]
- Gavali, J.T.; Carrillo, E.D.; García, M.C.; Sánchez, J.A. The Mitochondrial K-ATP Channel Opener Diazoxide Upregulates STIM1 and Orai1 via ROS and the MAPK Pathway in Adult Rat Cardiomyocytes. Cell Biosci. 2020, 10, 96. [Google Scholar] [CrossRef]
- Lakatta, E.G.; DiFrancesco, D. What Keeps Us Ticking: A Funny Current, a Calcium Clock, or Both? J. Mol. Cell Cardiol. 2009, 47, 157–170. [Google Scholar] [CrossRef]
- Irisawa, H.; Brown, H.F.; Giles, W. Cardiac Pacemaking in the Sinoatrial Node. Physiol. Rev. 1993, 73, 197–227. [Google Scholar] [CrossRef]
- DiFrancesco, D. The Pacemaker Current in the Sinus Node. Eur. Heart J. 1987, 8 (Suppl. L), 19–23. [Google Scholar] [CrossRef]
- Benzoni, P.; Bertoli, G.; Giannetti, F.; Piantoni, C.; Milanesi, R.; Pecchiari, M.; Barbuti, A.; Baruscotti, M.; Bucchi, A. The Funny Current: Even Funnier than 40 Years Ago. Uncanonical Expression and Roles of HCN/f Channels All over the Body. Prog. Biophys. Mol. Biol. 2021, 166, 189–204. [Google Scholar] [CrossRef]
- Moroni, A.; Gorza, L.; Beltrame, M.; Gravante, B.; Vaccari, T.; Bianchi, M.E.; Altomare, C.; Longhi, R.; Heurteaux, C.; Vitadello, M.; et al. Hyperpolarization-Activated Cyclic Nucleotide-Gated Channel 1 Is a Molecular Determinant of the Cardiac Pacemaker Current I(f). J. Biol. Chem. 2001, 276, 29233–29241. [Google Scholar] [CrossRef]
- Marger, L.; Mesirca, P.; Alig, J.; Torrente, A.; Dubel, S.; Engeland, B.; Kanani, S.; Fontanaud, P.; Striessnig, J.; Shin, H.-S.; et al. Pacemaker Activity and Ionic Currents in Mouse Atrioventricular Node Cells. Channels 2011, 5, 241–250. [Google Scholar] [CrossRef]
- Albarado-Ibañez, A.; Avelino-Cruz, J.E.; Velasco, M.; Torres-Jácome, J.; Hiriart, M. Metabolic Syndrome Remodels Electrical Activity of the Sinoatrial Node and Produces Arrhythmias in Rats. PLoS ONE 2013, 8, e76534. [Google Scholar] [CrossRef]
- Sato, T.; Sasaki, N.; Seharaseyon, J.; O’Rourke, B.; Marbán, E. Selective Pharmacological Agents Implicate Mitochondrial but Not Sarcolemmal K(ATP) Channels in Ischemic Cardioprotection. Circulation 2000, 101, 2418–2423. [Google Scholar] [CrossRef]
- Pasdois, P.; Beauvoit, B.; Tariosse, L.; Vinassa, B.; Bonoron-Adèle, S.; Dos Santos, P. Effect of Diazoxide on Flavoprotein Oxidation and Reactive Oxygen Species Generation during Ischemia-Reperfusion: A Study on Langendorff-Perfused Rat Hearts Using Optic Fibers. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H2088–H2097. [Google Scholar] [CrossRef]
- Bezerra Palácio, P.; Brito Lucas, A.M.; Varlla de Lacerda Alexandre, J.; Oliveira Cunha, P.L.; Ponte Viana, Y.I.; Albuquerque, A.C.; Nunes Varela, A.L.; Facundo, H.T. Pharmacological and Molecular Docking Studies Reveal That Glibenclamide Competitively Inhibits Diazoxide-Induced Mitochondrial ATP-Sensitive Potassium Channel Activation and Pharmacological Preconditioning. Eur. J. Pharmacol. 2021, 908, 174379. [Google Scholar] [CrossRef]
- Guo, J.; Giles, W.R.; Ward, C.A. Effect of Hydrogen Peroxide on the Membrane Currents of Sinoatrial Node Cells from Rabbit Heart. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H992–H999. [Google Scholar] [CrossRef]
- Jin, L.; Liu, Y.; Wu, Y.; Huang, Y.; Zhang, D. REST Is Not Resting: REST/NRSF in Health and Disease. Biomolecules 2023, 13, 1477. [Google Scholar] [CrossRef]
- Bruce, A.W.; López-Contreras, A.J.; Flicek, P.; Down, T.A.; Dhami, P.; Dillon, S.C.; Koch, C.M.; Langford, C.F.; Dunham, I.; Andrews, R.M.; et al. Functional Diversity for REST (NRSF) Is Defined by in Vivo Binding Affinity Hierarchies at the DNA Sequence Level. Genome Res. 2009, 19, 994–1005. [Google Scholar] [CrossRef]
- Cavadas, M.A.S.; Mesnieres, M.; Crifo, B.; Manresa, M.C.; Selfridge, A.C.; Keogh, C.E.; Fabian, Z.; Scholz, C.C.; Nolan, K.A.; Rocha, L.M.A.; et al. REST Is a Hypoxia-Responsive Transcriptional Repressor. Sci. Rep. 2016, 6, 31355. [Google Scholar] [CrossRef]
- Kuratomi, S.; Kuratomi, A.; Kuwahara, K.; Ishii, T.M.; Nakao, K.; Saito, Y.; Takano, M. NRSF Regulates the Developmental and Hypertrophic Changes of HCN4 Transcription in Rat Cardiac Myocytes. Biochem. Biophys. Res. Commun. 2007, 353, 67–73. [Google Scholar] [CrossRef]
- Schweizer, P.A.; Yampolsky, P.; Malik, R.; Thomas, D.; Zehelein, J.; Katus, H.A.; Koenen, M. Transcription Profiling of HCN-Channel Isotypes throughout Mouse Cardiac Development. Basic. Res. Cardiol. 2009, 104, 621–629. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, H.; Yin, H.; Zhao, X.; Zhang, Y. Emerging Roles of B56 Phosphorylation and Binding Motif in PP2A-B56 Holoenzyme Biological Function. Int. J. Mol. Sci. 2024, 25, 3185. [Google Scholar] [CrossRef]
- Inazumi, H.; Kuwahara, K. NRSF/REST-Mediated Epigenomic Regulation in the Heart: Transcriptional Control of Natriuretic Peptides and Beyond. Biology 2022, 11, 1197. [Google Scholar] [CrossRef]
- D’Souza, A.; Bucchi, A.; Johnsen, A.B.; Logantha, S.J.R.J.; Monfredi, O.; Yanni, J.; Prehar, S.; Hart, G.; Cartwright, E.; Wisloff, U.; et al. Exercise Training Reduces Resting Heart Rate via Downregulation of the Funny Channel HCN4. Nat. Commun. 2014, 5, 3775. [Google Scholar] [CrossRef]
- Averill-Bates, D. Reactive Oxygen Species and Cell Signaling. Review. Biochim. Biophys. Acta Mol. Cell Res. 2024, 1871, 119573. [Google Scholar] [CrossRef]
- Kuratomi, S.; Ohmori, Y.; Ito, M.; Shimazaki, K.; Muramatsu, S.-I.; Mizukami, H.; Uosaki, H.; Yamashita, J.K.; Arai, Y.; Kuwahara, K.; et al. The Cardiac Pacemaker-Specific Channel Hcn4 Is a Direct Transcriptional Target of MEF2. Cardiovasc. Res. 2009, 83, 682–687. [Google Scholar] [CrossRef]
- Priya Dharshini, L.C.; Vishnupriya, S.; Sakthivel, K.M.; Rasmi, R.R. Oxidative Stress Responsive Transcription Factors in Cellular Signalling Transduction Mechanisms. Cell Signal 2020, 72, 109670. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.H.; Shih, N.L.; Chen, S.Y.; Wang, D.L.; Chen, J.J. Reactive Oxygen Species Modulate Endothelin-I-Induced c-Fos Gene Expression in Cardiomyocytes. Cardiovasc. Res. 1999, 41, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Koenigsberger, C.; Chicca, J.J.; Amoureux, M.C.; Edelman, G.M.; Jones, F.S. Differential Regulation by Multiple Promoters of the Gene Encoding the Neuron-Restrictive Silencer Factor. Proc. Natl. Acad. Sci. USA 2000, 97, 2291–2296. [Google Scholar] [CrossRef] [PubMed]
- Shea-Eaton, W.; Sandhoff, T.W.; Lopez, D.; Hales, D.B.; McLean, M.P. Transcriptional Repression of the Rat Steroidogenic Acute Regulatory (StAR) Protein Gene by the AP-1 Family Member c-Fos. Mol. Cell Endocrinol. 2002, 188, 161–170. [Google Scholar] [CrossRef]
- Guida, N.; Laudati, G.; Anzilotti, S.; Secondo, A.; Montuori, P.; Di Renzo, G.; Canzoniero, L.M.T.; Formisano, L. Resveratrol via Sirtuin-1 Downregulates RE1-Silencing Transcription Factor (REST) Expression Preventing PCB-95-Induced Neuronal Cell Death. Toxicol. Appl. Pharmacol. 2015, 288, 387–398. [Google Scholar] [CrossRef]
- Reno, C.M.; Bayles, J.; Skinner, A.; Fisher, S.J. Glibenclamide Prevents Hypoglycemia-Induced Fatal Cardiac Arrhythmias in Rats. Endocrinology 2018, 159, 2614–2620. [Google Scholar] [CrossRef]
- DiFrancesco, D.; Borer, J.S. The Funny Current: Cellular Basis for the Control of Heart Rate. Drugs 2007, 67 (Suppl. 2), 15–24. [Google Scholar] [CrossRef]
- Longobardi, S.; Sher, A.; Niederer, S.A. Quantitative Mapping of Force-pCa Curves to Whole-Heart Contraction and Relaxation. J. Physiol. 2022, 600, 3497–3516. [Google Scholar] [CrossRef]
- Narasimhan, G.; Carrillo, E.D.; Hernández, A.; García, M.C.; Sánchez, J.A. Protective Action of Diazoxide on Isoproterenol-Induced Hypertrophy Is Mediated by Reduction in MicroRNA-132 Expression. J. Cardiovasc. Pharmacol. 2018, 72, 222–230. [Google Scholar] [CrossRef]
- Sharpe, E.J.; St Clair, J.R.; Proenza, C. Methods for the Isolation, Culture, and Functional Characterization of Sinoatrial Node Myocytes from Adult Mice. J. Vis. Exp. 2016, 116, 54555. [Google Scholar] [CrossRef]
- Garlid, K.D.; Paucek, P.; Yarov-Yarovoy, V.; Sun, X.; Schindler, P.A. The Mitochondrial K Channel as a Receptor for Potassium Channel Openers. J. Biol. Chem. 1996, 271, 8796–8799. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Sato, T.; Seharaseyon, J.; Liu, Y.; Johns, D.C.; O’Rourke, B.; Marbán, E. Pharmacological and Histochemical Distinctions between Molecularly Defined Sarcolemmal KATP Channels and Native Cardiac Mitochondrial KATP Channels. Mol. Pharmacol. 1999, 55, 1000–1005. [Google Scholar] [CrossRef]
- Bucchi, A.; Baruscotti, M.; Nardini, M.; Barbuti, A.; Micheloni, S.; Bolognesi, M.; DiFrancesco, D. Identification of the Molecular Site of Ivabradine Binding to HCN4 Channels. PLoS ONE 2013, 8, e53132. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A Quantitative Description of Membrane Current and Its Application to Conduction and Excitation in Nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Holden, A.V.; Kodama, I.; Honjo, H.; Lei, M.; Varghese, T.; Boyett, M.R. Mathematical Models of Action Potentials in the Periphery and Center of the Rabbit Sinoatrial Node. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H397–H421. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.O.; Wilders, R. Hyperpolarization-Activated Current, If, in Mathematical Models of Rabbit Sinoatrial Node Pacemaker Cells. Biomed. Res. Int. 2013, 2013, 872454. [Google Scholar] [CrossRef]
- Sarai, N.; Matsuoka, S.; Kuratomi, S.; Ono, K.; Noma, A. Role of Individual Ionic Current Systems in the SA Node Hypothesized by a Model Study. Jpn. J. Physiol. 2003, 53, 125–134. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kelley, J.B.; Paschal, B.M. Fluorescence-Based Quantification of Nucleocytoplasmic Transport. Methods 2019, 157, 106–114. [Google Scholar] [CrossRef]
- Carrillo, E.D.; Alvarado, J.A.; Hernández, A.; Lezama, I.; García, M.C.; Sánchez, J.A. Thyroid Hormone Upregulates Cav1.2 Channels in Cardiac Cells via the Downregulation of the Channels’ Β4 Subunit. Int. J. Mol. Sci. 2024, 25, 10798. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orea, W.; Carrillo, E.D.; Hernández, A.; Moreno, R.; García, M.C.; Sánchez, J.A. Pharmacological Preconditioning with Diazoxide Upregulates HCN4 Channels in the Sinoatrial Node of Adult Rat Cardiomyocytes. Int. J. Mol. Sci. 2025, 26, 6062. https://doi.org/10.3390/ijms26136062
Orea W, Carrillo ED, Hernández A, Moreno R, García MC, Sánchez JA. Pharmacological Preconditioning with Diazoxide Upregulates HCN4 Channels in the Sinoatrial Node of Adult Rat Cardiomyocytes. International Journal of Molecular Sciences. 2025; 26(13):6062. https://doi.org/10.3390/ijms26136062
Chicago/Turabian StyleOrea, Wilibaldo, Elba D. Carrillo, Ascención Hernández, Rubén Moreno, María C. García, and Jorge A. Sánchez. 2025. "Pharmacological Preconditioning with Diazoxide Upregulates HCN4 Channels in the Sinoatrial Node of Adult Rat Cardiomyocytes" International Journal of Molecular Sciences 26, no. 13: 6062. https://doi.org/10.3390/ijms26136062
APA StyleOrea, W., Carrillo, E. D., Hernández, A., Moreno, R., García, M. C., & Sánchez, J. A. (2025). Pharmacological Preconditioning with Diazoxide Upregulates HCN4 Channels in the Sinoatrial Node of Adult Rat Cardiomyocytes. International Journal of Molecular Sciences, 26(13), 6062. https://doi.org/10.3390/ijms26136062