

Genetic Variants in Oxidative Stress-Related Genes and Their Impact on Prognosis and Treatment Response in Chronic Myeloid Leukemia Patients

 , , , ,
, , , ,  , and
, and
Abstract

1. Introduction
2. Results
2.1. Characteristics of CML Patients
2.2. Genetic Variants Associated with TKI Response and BCR::ABL1 Mutational Status
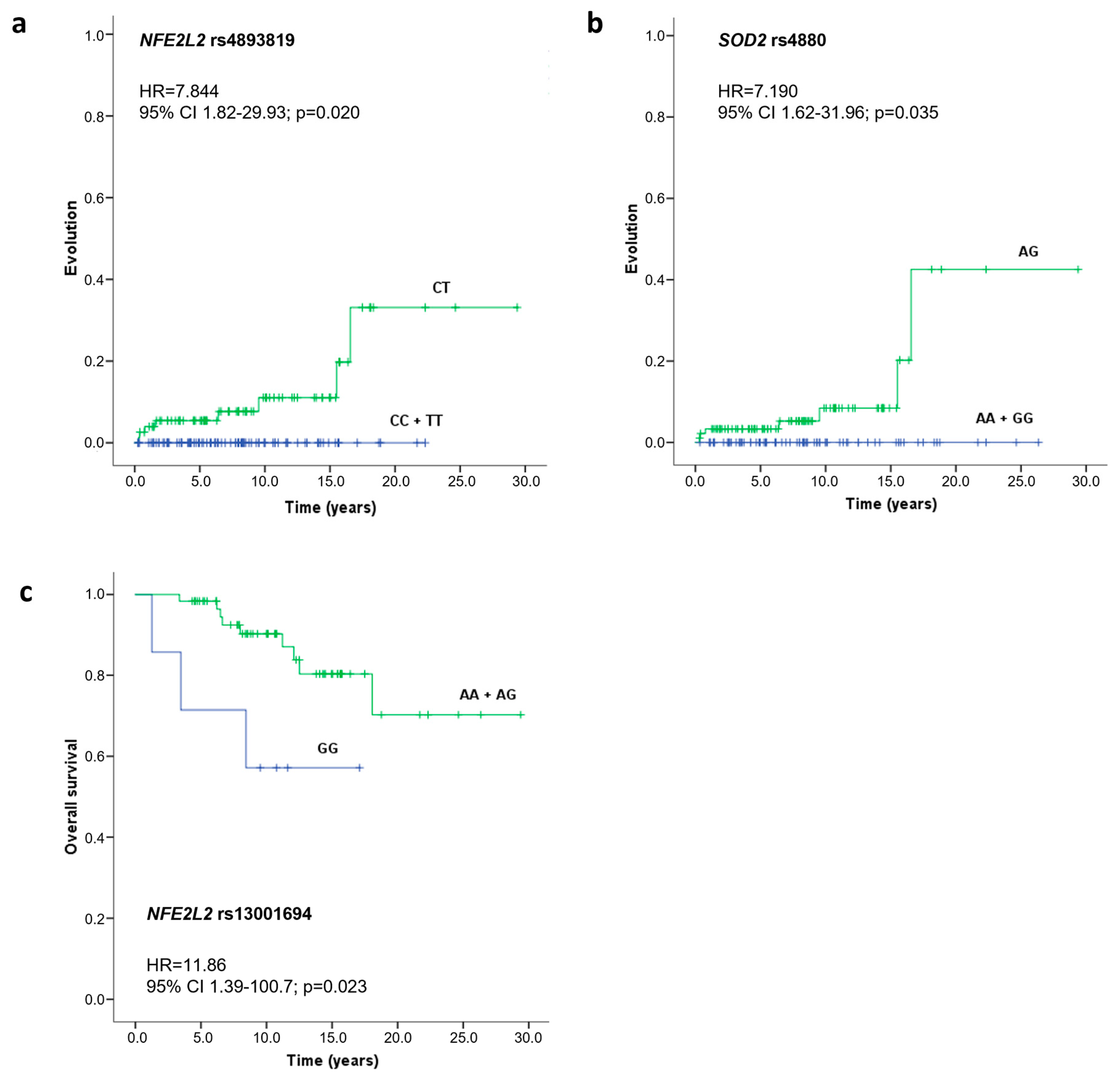
2.3. Impacts of Studied SNVs on Progression and Overall Survival
2.4. Influence of Studied SNVs on Gene Expression Levels and Protein Function
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Gene and SNV Selection
4.3. Genotyping
4.4. Genetic Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASO-PCR | Allele-specific oligonucleotide polymerase chain reaction |
| CAT | Catalase |
| CI | Confidence interval |
| CML | Chronic myeloid leukemia |
| ELN | European Leukemia Net |
| GP | Genotypic profile |
| GPX1 | Glutathione peroxidase 1 |
| HR | Hazard ratio |
| HWE | Hardy–Weinberg equilibrium |
| KEAP1 | Kelch-like ECH-associated protein 1 |
| M | Major allele |
| m | Minor allele |
| MAF | Frequency of the minor allele |
| MCD | Codominant model |
| MD | Dominant model |
| MgCL2 | Magnesium chloride |
| MOD | Overdominant model |
| MR | Recessive model |
| NFE2L2 | Nuclear factor erythroid 2-related factor 2 gene |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| OD | Odds ratio |
| OS | Oxidative stress |
| RFLP-PCR | Restriction fragment length polymorphism polymerase chain reaction |
| Ref | Reference allele or genotype |
| ROS | Reactive oxygen species |
| SNV | Single nucleotide variants |
| SOD2 | Superoxide dismutase [Mn], mitochondrial |
| Ta | Annealing temperature |
| TETRA-ARMS-PCR | Tetra-primer amplification refractory mutation system-polymerase chain reaction |
| TF | Transcription factor |
| TKI | Tyrosine kinase inhibitor |
| WHO | World Health Organization |
References
- Ali, M.A. Chronic Myeloid Leukemia in the Era of Tyrosine Kinase Inhibitors: An Evolving Paradigm of Molecularly Targeted Therapy. Mol. Diagn. Ther. 2016, 20, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Baccarani, M.; Castagnetti, F.; Gugliotta, G.; Rosti, G. A review of the European LeukemiaNet recommendations for the management of CML. Ann. Hematol. 2015, 94 (Suppl. S2), S141–S147. [Google Scholar] [CrossRef] [PubMed]
- Hochhaus, A.; Saussele, S.; Rosti, G.; Mahon, F.X.; Janssen, J.; Hjorth-Hansen, H.; Richter, J.; Buske, C.; Committee, E.G. Chronic myeloid leukaemia: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2017, 28, iv41–iv51. [Google Scholar] [CrossRef]
- Alves, R.; Goncalves, A.C.; Rutella, S.; Almeida, A.M.; De Las Rivas, J.; Trougakos, I.P.; Sarmento Ribeiro, A.B. Resistance to Tyrosine Kinase Inhibitors in Chronic Myeloid Leukemia-From Molecular Mechanisms to Clinical Relevance. Cancers 2021, 13, 4820. [Google Scholar] [CrossRef]
- Allegra, A.; Mirabile, G.; Caserta, S.; Stagno, F.; Russo, S.; Pioggia, G.; Gangemi, S. Oxidative Stress and Chronic Myeloid Leukemia: A Balance between ROS-Mediated Pro- and Anti-Apoptotic Effects of Tyrosine Kinase Inhibitors. Antioxidants 2024, 13, 461. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Parvez, S.; Long, M.J.C.; Poganik, J.R.; Aye, Y. Redox Signaling by Reactive Electrophiles and Oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef]
- Pascu, V.E.G.; AM, G.A. Involvement of Oxidative Stress in Resistance to Tyrosine-Kinase Inhibitors Therapy in Chronic Myeloid Leukemia. Curr. Health Sci. J. 2020, 46, 420–432. [Google Scholar]
- Hole, P.S.; Darley, R.L.; Tonks, A. Do reactive oxygen species play a role in myeloid leukemias? Blood 2011, 117, 5816–5826. [Google Scholar] [CrossRef] [PubMed]
- Slupianek, A.; Falinski, R.; Znojek, P.; Stoklosa, T.; Flis, S.; Doneddu, V.; Pytel, D.; Synowiec, E.; Blasiak, J.; Bellacosa, A.; et al. BCR-ABL1 kinase inhibits uracil DNA glycosylase UNG2 to enhance oxidative DNA damage and stimulate genomic instability. Leukemia 2013, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- de la Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Khodakarami, A.; Adibfar, S.; Karpisheh, V.; Abolhasani, S.; Jalali, P.; Mohammadi, H.; Gholizadeh Navashenaq, J.; Hojjat-Farsangi, M.; Jadidi-Niaragh, F. The molecular biology and therapeutic potential of Nrf2 in leukemia. Cancer Cell Int. 2022, 22, 241. [Google Scholar] [CrossRef]
- Scharfe, C.P.I.; Tremmel, R.; Schwab, M.; Kohlbacher, O.; Marks, D.S. Genetic variation in human drug-related genes. Genome Med. 2017, 9, 117. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Brozovic, A.; Goncalves, A.C.; Jurkovicova, D.; Line, A.; Machuqueiro, M.; Saponara, S.; Sarmento-Ribeiro, A.B.; Xavier, C.P.R.; Vasconcelos, M.H. The multi-factorial nature of clinical multidrug resistance in cancer. Drug Resist. Updates 2019, 46, 100645. [Google Scholar] [CrossRef]
- Goncalves, A.C.; Alves, R.; Baldeiras, I.; Cortesao, E.; Carda, J.P.; Branco, C.C.; Oliveiros, B.; Loureiro, L.; Pereira, A.; Nascimento Costa, J.M.; et al. Genetic variants involved in oxidative stress, base excision repair, DNA methylation, and folate metabolism pathways influence myeloid neoplasias susceptibility and prognosis. Mol. Carcinog. 2017, 56, 130–148. [Google Scholar] [CrossRef]
- Alves, R.; Goncalves, A.C.; Jorge, J.; Marques, G.; Ribeiro, A.B.; Tenreiro, R.; Coucelo, M.; Diamond, J.; Oliveiros, B.; Pereira, A.; et al. Genetic Variants of ABC and SLC Transporter Genes and Chronic Myeloid Leukaemia: Impact on Susceptibility and Prognosis. Int. J. Mol. Sci. 2022, 23, 9815. [Google Scholar] [CrossRef]
- Yang, W.; Liu, H.; Duan, B.; Xu, X.; Carmody, D.; Luo, S.; Walsh, K.M.; Abbruzzese, J.L.; Zhang, X.; Chen, X.; et al. Three novel genetic variants in NRF2 signaling pathway genes are associated with pancreatic cancer risk. Cancer Sci. 2019, 110, 2022–2032. [Google Scholar] [CrossRef] [PubMed]
- Nunes Dos Santos, K.; Florentino, R.M.; Franca, A.; Lima Filho, A.C.M.; Santos, M.L.D.; Missiaggia, D.; Fonseca, M.C.; Brasil Costa, I.; Vidigal, P.V.T.; Nathanson, M.H.; et al. Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse. Int. J. Mol. Sci. 2019, 20, 3589. [Google Scholar] [CrossRef]
- Sjoblom, T.; Jones, S.; Wood, L.D.; Parsons, D.W.; Lin, J.; Barber, T.D.; Mandelker, D.; Leary, R.J.; Ptak, J.; Silliman, N.; et al. The consensus coding sequences of human breast and colorectal cancers. Science 2006, 314, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Sporn, M.B.; Liby, K.T. NRF2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef]
- Okano, Y.; Nezu, U.; Enokida, Y.; Lee, M.T.; Kinoshita, H.; Lezhava, A.; Hayashizaki, Y.; Morita, S.; Taguri, M.; Ichikawa, Y.; et al. SNP (-617C>A) in ARE-like loci of the NRF2 gene: A new biomarker for prognosis of lung adenocarcinoma in Japanese non-smoking women. PLoS ONE 2013, 8, e73794. [Google Scholar] [CrossRef] [PubMed]
- Hartikainen, J.M.; Tengstrom, M.; Kosma, V.M.; Kinnula, V.L.; Mannermaa, A.; Soini, Y. Genetic polymorphisms and protein expression of NRF2 and Sulfiredoxin predict survival outcomes in breast cancer. Cancer Res. 2012, 72, 5537–5546. [Google Scholar] [CrossRef]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; et al. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J. 2007, 21, 2237–2246. [Google Scholar] [CrossRef]
- Zazueta, C.; Jimenez-Uribe, A.P.; Pedraza-Chaverri, J.; Buelna-Chontal, M. Genetic Variations on Redox Control in Cardiometabolic Diseases: The Role of Nrf2. Antioxidants 2022, 11, 507. [Google Scholar] [CrossRef]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Hirata, I.; et al. Nrf2 gene promoter polymorphism and gastric carcinogenesis. Hepatogastroenterology 2008, 55, 750–754. [Google Scholar]
- Matana, A.; Ziros, P.G.; Chartoumpekis, D.V.; Renaud, C.O.; Polasek, O.; Hayward, C.; Zemunik, T.; Sykiotis, G.P. Rare and common genetic variations in the Keap1/Nrf2 antioxidant response pathway impact thyroglobulin gene expression and circulating levels, respectively. Biochem. Pharmacol. 2020, 173, 113605. [Google Scholar] [CrossRef]
- Islam, S.S.; Qassem, K.; Islam, S.; Parag, R.R.; Rahman, M.Z.; Farhat, W.A.; Yeger, H.; Aboussekhra, A.; Karakas, B.; Noman, A.S.M. Genetic alterations of Keap1 confers chemotherapeutic resistance through functional activation of Nrf2 and Notch pathway in head and neck squamous cell carcinoma. Cell Death Dis. 2022, 13, 696. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.; Fassett, R.G.; Geraghty, D.P.; Kunde, D.A.; Ball, M.J.; Robertson, I.K.; Coombes, J.S. Relationships between single nucleotide polymorphisms of antioxidant enzymes and disease. Gene 2012, 501, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Sutton, A.; Imbert, A.; Igoudjil, A.; Descatoire, V.; Cazanave, S.; Pessayre, D.; Degoul, F. The manganese superoxide dismutase Ala16Val dimorphism modulates both mitochondrial import and mRNA stability. Pharm. Genomics 2005, 15, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Kinnula, V.L.; Crapo, J.D. Superoxide dismutases in malignant cells and human tumors. Free Radic. Biol. Med. 2004, 36, 718–744. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, Y.; Gu, D.; Lee, N.P.; Sun, S.; Gong, W.; Tan, Y.; Luk, J.M.; Chen, J. SOD2 rs4880 CT/CC genotype predicts poor survival for Chinese gastric cancer patients received platinum and fluorouracil based adjuvant chemotherapy. Am. J. Transl. Res. 2015, 7, 401–410. [Google Scholar] [CrossRef]
- Salimi, S.; Harati-Sadegh, M.; Eskandari, M.; Heidari, Z. The effects of the genetic polymorphisms of antioxidant enzymes on susceptibility to papillary thyroid carcinoma. IUBMB Life 2020, 72, 1045–1053. [Google Scholar] [CrossRef]
- Gallegos-Arreola, M.P.; Ramirez-Patino, R.; Sanchez-Lopez, J.Y.; Zuniga-Gonzalez, G.M.; Figuera, L.E.; Delgado-Saucedo, J.I.; Gomez-Meda, B.C.; Rosales-Reynoso, M.A.; Puebla-Perez, A.M.; Lemus-Varela, M.L.; et al. SOD2 Gene Variants (rs4880 and rs5746136) and Their Association with Breast Cancer Risk. Curr. Issues Mol. Biol. 2022, 44, 5221–5233. [Google Scholar] [CrossRef]
- Kucukgergin, C.; Sanli, O.; Amasyali, A.S.; Tefik, T.; Seckin, S. Genetic variants of MnSOD and GPX1 and susceptibility to bladder cancer in a Turkish population. Med. Oncol. 2012, 29, 1928–1934. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, R.; Chen, N.; Yang, L.; Wang, Y.; Sun, Y.; Huang, L.; Zhu, M.; Ji, Y.; Li, W. Association between glutathione peroxidase-1 (GPX1) Rs1050450 polymorphisms and cancer risk. Int. J. Clin. Exp. Pathol. 2017, 10, 9527–9540. [Google Scholar]
- Ravn-Haren, G.; Olsen, A.; Tjonneland, A.; Dragsted, L.O.; Nexo, B.A.; Wallin, H.; Overvad, K.; Raaschou-Nielsen, O.; Vogel, U. Associations between GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis 2006, 27, 820–825. [Google Scholar] [CrossRef]
- Jablonska, E.; Gromadzinska, J.; Peplonska, B.; Fendler, W.; Reszka, E.; Krol, M.B.; Wieczorek, E.; Bukowska, A.; Gresner, P.; Galicki, M.; et al. Lipid peroxidation and glutathione peroxidase activity relationship in breast cancer depends on functional polymorphism of GPX1. BMC Cancer 2015, 15, 657. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, H.; Zhou, J.; Shao, Q. Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer. Cancers 2022, 14, 2560. [Google Scholar] [CrossRef]
- Kagita, S.; Digumarti, R.; Gundeti, S. Role of Antioxidant Gene Polymorphisms in Risk and Prognosis of Chronic Myeloid Leukemia. Asian Pac. J. Cancer Biol. 2021, 6, 27–36. [Google Scholar] [CrossRef]
- Hong, Z.; Tian, C.; Zhang, X. GPX1 gene Pro200Leu polymorphism, erythrocyte GPX activity, and cancer risk. Mol. Biol. Rep. 2013, 40, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Liu, X.; Wang, M.; Wang, X.; Kang, H.; Lin, S.; Yang, P.; Dai, C.; Xu, P.; Li, S.; et al. Two common functional catalase gene polymorphisms (rs1001179 and rs794316) and cancer susceptibility: Evidence from 14,942 cancer cases and 43,285 controls. Oncotarget 2016, 7, 62954–62965. [Google Scholar] [CrossRef]
- Galasso, M.; Dalla Pozza, E.; Chignola, R.; Gambino, S.; Cavallini, C.; Quaglia, F.M.; Lovato, O.; Dando, I.; Malpeli, G.; Krampera, M.; et al. The rs1001179 SNP and CpG methylation regulate catalase expression in chronic lymphocytic leukemia. Cell. Mol. Life Sci. 2022, 79, 521. [Google Scholar] [CrossRef]
- Eras, N.; Turkoz, G.; Tombak, A.; Tiftik, N.; Yalin, S.; Berkoz, M.; Erden, S.; Akbas, E. An investigation of the relation between catalase C262T gene polymorphism and catalase enzyme activity in leukemia patients. Arch. Med. Sci. 2021, 17, 928–933. [Google Scholar] [CrossRef]
- Ahn, J.; Nowell, S.; McCann, S.E.; Yu, J.; Carter, L.; Lang, N.P.; Kadlubar, F.F.; Ratnasinghe, L.D.; Ambrosone, C.B. Associations between catalase phenotype and genotype: Modification by epidemiologic factors. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1217–1222. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, L.; Zhao, J.; Huang, X.; Song, L.; Luo, J.; Ma, L.; Li, S.; Qin, X. Association between catalase gene polymorphisms and risk of chronic hepatitis B, hepatitis B virus-related liver cirrhosis and hepatocellular carcinoma in Guangxi population: A case-control study. Medicine 2015, 94, e702. [Google Scholar] [CrossRef]
- Saadat, M.; Saadat, S. Genetic Polymorphism of CAT C-262 T and Susceptibility to Breast Cancer, a Case-Control Study and Meta-Analysis of the Literatures. Pathol. Oncol. Res. 2015, 21, 433–437. [Google Scholar] [CrossRef]
- Ebrahimpour, S.; Saadat, I. Association of CAT C-262T and SOD1 A251G single nucleotide polymorphisms susceptible to gastric cancer. Mol. Biol. Res. Commun. 2014, 3, 223–229. [Google Scholar] [PubMed]
- Banescu, C.; Trifa, A.P.; Voidazan, S.; Moldovan, V.G.; Macarie, I.; Benedek Lazar, E.; Dima, D.; Duicu, C.; Dobreanu, M. CAT, GPX1, MnSOD, GSTM1, GSTT1, and GSTP1 genetic polymorphisms in chronic myeloid leukemia: A case-control study. Oxid. Med. Cell Longev. 2014, 2014, 875861. [Google Scholar] [CrossRef]
- Ben Hassine, I.; Gharbi, H.; Soltani, I.; Ben Hadj Othman, H.; Farrah, A.; Amouri, H.; Teber, M.; Ghedira, H.; Ben Youssef, Y.; Safra, I.; et al. Molecular study of ABCB1 gene and its correlation with imatinib response in chronic myeloid leukemia. Cancer Chemother. Pharmacol. 2017, 80, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Neul, C.; Schaeffeler, E.; Sparreboom, A.; Laufer, S.; Schwab, M.; Nies, A.T. Impact of Membrane Drug Transporters on Resistance to Small-Molecule Tyrosine Kinase Inhibitors. Trends Pharmacol. Sci. 2016, 37, 904–932. [Google Scholar] [CrossRef]
- You, F.M.; Huo, N.; Gu, Y.Q.; Luo, M.C.; Ma, Y.; Hane, D.; Lazo, G.R.; Dvorak, J.; Anderson, O.D. BatchPrimer3: A high throughput web application for PCR and sequencing primer design. BMC Bioinform. 2008, 9, 253. [Google Scholar] [CrossRef]
- Mansur, Y.A.; Rojano, E.; Ranea, J.A.G.; Perkins, J.R. Chapter 7—Analyzing the Effects of Genetic Variation in Noncoding Genomic Regions. In Precision Medicine; Deigner, H.-P., Kohl, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 119–144. [Google Scholar]
- Lewis, C.M. Genetic association studies: Design, analysis and interpretation. Brief. Bioinform. 2002, 3, 146–153. [Google Scholar] [CrossRef]
- Setu, T.; Basak, T. An Introduction to Basic Statistical Models in Genetics. Open J. Stat. 2021, 11, 1017–1025. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Characteristics | CML | ||||||
|---|---|---|---|---|---|---|---|
| All Patients (n = 187) | TKI Responders (n = 138) | TKI Resistors (n = 49) | p Value | ||||
| Demographic Features | |||||||
| Gender (%) | |||||||
| Male | 110 | (58.8) | 78 | (56.5) | 32 | (65.3) | p = 0.294 * |
| Female | 77 | (41.2) | 60 | (43.5) | 17 | (34.7) | |
| Age (years) | |||||||
| Median | 54 | 54 | 50 | p = 0.408 # | |||
| Range | 15–86 | 15–86 | 18–79 | ||||
| Clinical Features | |||||||
| Phase of Disease | |||||||
| Chronic Phase (%) | 177 | (94.6) | 131 | (94.9) | 46 | (93.9) | p = 0.091 * |
| Accelerate Phase (%) | 5 | (2.7) | 5 | (3.6) | – | – | |
| Blast Crisis (%) | 5 | (2.7) | 2 | (1.5) | 3 | (6.1) | |
| Scoring Systems | |||||||
| Sokal Score | (n = 138) | (n = 105) | (n = 33) | ||||
| Low Risk (%) | 75 | (54.4) | 59 | (56.2) | 16 | (48.5) | p = 0.712 * |
| Intermediate Risk (%) | 45 | (32.6) | 33 | (31.4) | 12 | (36.4) | |
| High Risk (%) | 18 | (13.0) | 13 | (12.4) | 5 | (15.1) | |
| Euro Score | (n = 138) | (n = 105) | (n = 33) | ||||
| Low Risk (%) | 100 | (72.5) | 79 | (75.2) | 21 | (63.7) | p = 0.266 * |
| Intermediate Risk (%) | 32 | (23.2) | 21 | (20.0) | 11 | (33.3) | |
| High Risk (%) | 6 | (4.3) | 5 | (4.8) | 1 | (3.0) | |
| EUTOS Score | (n = 136) | (n = 104) | (n = 32) | ||||
| Low Risk (%) | 119 | (87.5) | 92 | (88.5) | 27 | (84.4) | p = 0.528 * |
| High Risk (%) | 17 | (12.5) | 12 | (11.5) | 5 | (15.6) | |
| First-Line TKI | |||||||
| Imatinib (%) | 178 | (95.2) | 129 | (93.5) | 49 | (100.0) | p = 0.700 * |
| Other TKI (%) | 9 | (4.8) | 9 | (6.5) | – | – | |
| Number of TKIs during Treatment | |||||||
| 1 TKI (%) | 138 | (73.8) | 138 | (100.0) | – | – | p < 0.001 * |
| 2 TKIs (%) | 37 | (19.8) | – | – | 37 | (75.5) | |
| ≥3 TKIs (%) | 12 | (6.4) | – | – | 12 | (24.5) | |
| Mutations on BCR::ABL1 | (n = 104) | (n = 69) | (n = 35) | ||||
| Present (%) | 22 | (21.2) | 11 | (15.9) | 11 | (31.4) | p = 0.068 * |
| Absence (%) | 82 | (78.8) | 58 | (84.1) | 24 | (68.6) | |
| Disease Evolution | |||||||
| Progression (%) | 8 | (4.3) | – | – | 8 | (16.3) | p < 0.001 * |
| No Progression (%) | 179 | (95.7) | 138 | (100.0) | 41 | (83.7) | |
| Overall Survival | (n = 194) | (n = 138) | (n = 49) | ||||
| Death (%) | 27 | (14.4) | 12 | (8.7) | 15 | (30.6) | p < 0.001 * |
| Survive (%) | 160 | (85.6) | 126 | (91.3) | 34 | (69.4) | |
| GPX1 rs1050450 | KEAP1 rs113540846 | |||
|---|---|---|---|---|
| Allele G | Allele A * | Allele G | Allele A * | |
| TKI-sensitive | 155 | 115 | 190 | 44 |
| TKI-resisitant | 67 | 27 | 66 | 0 |
| OR (95% CI) | 1.841 (1.108–3.059) | 31.07 (1.886–511.9) | ||
| p value | 0.020 | <0.0001 | ||
| Gene: dbSNV | n | % | OR (95% CI) | p Value | n | % | ||
|---|---|---|---|---|---|---|---|---|
| GPX1 rs1050450 | TKI-resistant | TKI-sensitive | ||||||
| GG | 25 | 53.2 | Ref. | 46 | 34.1 | |||
| GA | 17 | 36.2 | 0.497 (0.241–1.024) | 0.058 | 63 | 46.7 | ||
| AA | 5 | 10.6 | 0.354 (0.121–1.036) | 0.058 | 26 | 19.3 | ||
| GG (MD) | 2.199 (1.120–4.316) | 0.022 | ||||||
| AA (MR) | 0.499 (0.180–1.386) | 0.182 | ||||||
| GA (MOD) | 0.648 (0.327–1.284) | 0.213 | ||||||
| NFE2L2 rs4893819 | 3 or more TKIs | 2 TKIs | ||||||
| CC | 3 | 37.3 | Ref. | 11 | 35.5 | |||
| CT | 8 | 72.7 | 2.933 (0.605–14.231) | 0.182 | 10 | 32.3 | ||
| TT | 0 | 0.0 | - | - | 10 | 32.3 | ||
| CC (MD) | 0.682 (0.150–3.109) | 0.621 | ||||||
| TT (MR) | - | - | ||||||
| CT (MOD) | 5.600 (1.218–25.751) | 0.027 | ||||||
| NFE2L2 rs13001694 | With mutation | Without mutation | ||||||
| AA | 1 | 7.7 | Ref. | 15 | 35.7 | |||
| AG | 12 | 92.3 | 8.571 (1.004–73.210) | 0.050 | 21 | 50 | ||
| GG | 0 | 0.0 | - | - | 6 | 14.3 | ||
| AA (MD) | 0.150 (0.018–1.269) | 0.082 | ||||||
| GG (MR) | - | - | ||||||
| AG (MOD) | 12.000 (1.429–100.754) | 0.022 | ||||||
| Gene: dbSNV | n | % | OR (95% CI) | p-Value | n | % | ||
|---|---|---|---|---|---|---|---|---|
| CAT rs1001179 | Death | Survival | ||||||
| CC | 4 | 33.3 | Ref. | 5 | 8.9 | |||
| CT | 3 | 25.0 | 0.163 (0.027–0.969) | 0.046 | 23 | 41.1 | ||
| TT | 5 | 41.7 | 0.223 (0.044–1.131) | 0.07 | 28 | 50.0 | ||
| CC (MD) | 5.100 (1.125–23.117) | 0.035 | ||||||
| TT (MR) | 0.714 (0.202–2.522) | 0.601 | ||||||
| CT (MOD) | 0.478 (0.117–1.961) | 0.306 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, R.; Ventura, F.; Jorge, J.; Marques, G.; Coucelo, M.; Diamond, J.; Oliveiros, B.; Pereira, A.; Freitas-Tavares, P.; Almeida, A.M.; et al. Genetic Variants in Oxidative Stress-Related Genes and Their Impact on Prognosis and Treatment Response in Chronic Myeloid Leukemia Patients. Int. J. Mol. Sci. 2025, 26, 5682. https://doi.org/10.3390/ijms26125682
Alves R, Ventura F, Jorge J, Marques G, Coucelo M, Diamond J, Oliveiros B, Pereira A, Freitas-Tavares P, Almeida AM, et al. Genetic Variants in Oxidative Stress-Related Genes and Their Impact on Prognosis and Treatment Response in Chronic Myeloid Leukemia Patients. International Journal of Molecular Sciences. 2025; 26(12):5682. https://doi.org/10.3390/ijms26125682
Chicago/Turabian StyleAlves, Raquel, Filipa Ventura, Joana Jorge, Gilberto Marques, Margarida Coucelo, Joana Diamond, Bárbara Oliveiros, Amélia Pereira, Paulo Freitas-Tavares, António M. Almeida, and et al. 2025. "Genetic Variants in Oxidative Stress-Related Genes and Their Impact on Prognosis and Treatment Response in Chronic Myeloid Leukemia Patients" International Journal of Molecular Sciences 26, no. 12: 5682. https://doi.org/10.3390/ijms26125682
APA StyleAlves, R., Ventura, F., Jorge, J., Marques, G., Coucelo, M., Diamond, J., Oliveiros, B., Pereira, A., Freitas-Tavares, P., Almeida, A. M., Gonçalves, A. C., & Sarmento-Ribeiro, A. B. (2025). Genetic Variants in Oxidative Stress-Related Genes and Their Impact on Prognosis and Treatment Response in Chronic Myeloid Leukemia Patients. International Journal of Molecular Sciences, 26(12), 5682. https://doi.org/10.3390/ijms26125682